

LanCL2 Implicates in Testicular Redox Homeostasis and Acrosomal Maturation

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Quantitative Real-Time PCR
2.3. Western Blotting
2.4. Immunochemical Staining
2.5. Immunofluorescence Staining
2.6. H&E Staining and Luxol Fast Blue Staining
2.7. Sperm Parameter Analysis and Acrosome Reaction Assay
2.8. Cell Culture
2.9. Biochemical Assays
2.10. Behavioral Tests
2.10.1. Mouse Gait Analysis
2.10.2. Accelerated Rotarod Test
2.10.3. Pole Test
2.11. Statistical Analysis
3. Results
3.1. Age-Dependent Expression of LanCL2 Correlates with Testicular Maturation
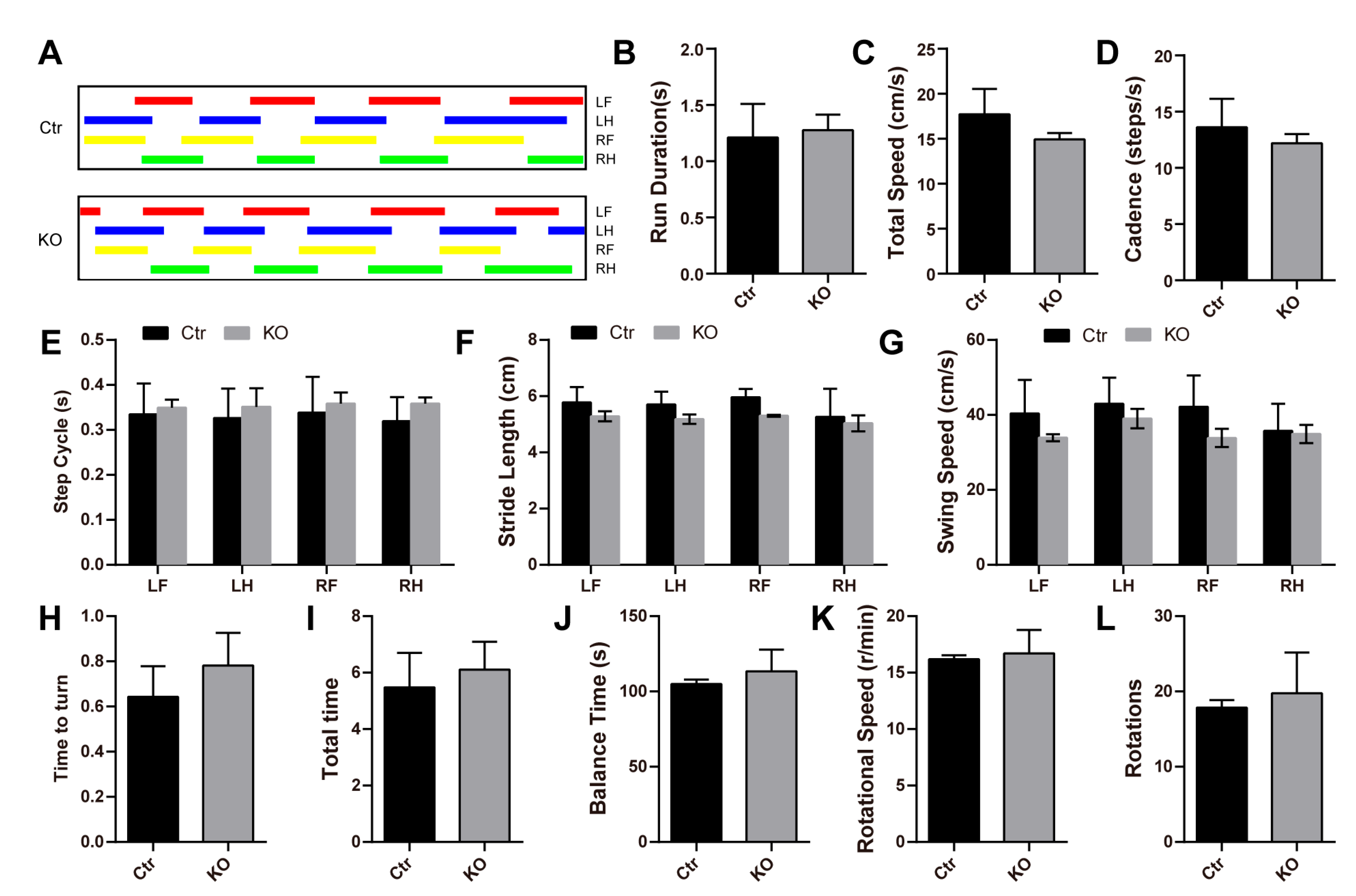
3.2. Loss of LanCL2 Does Not Affect Normal Brain Development
3.3. LanCL2 Deficiency Results in Oligoasthenozoospermia
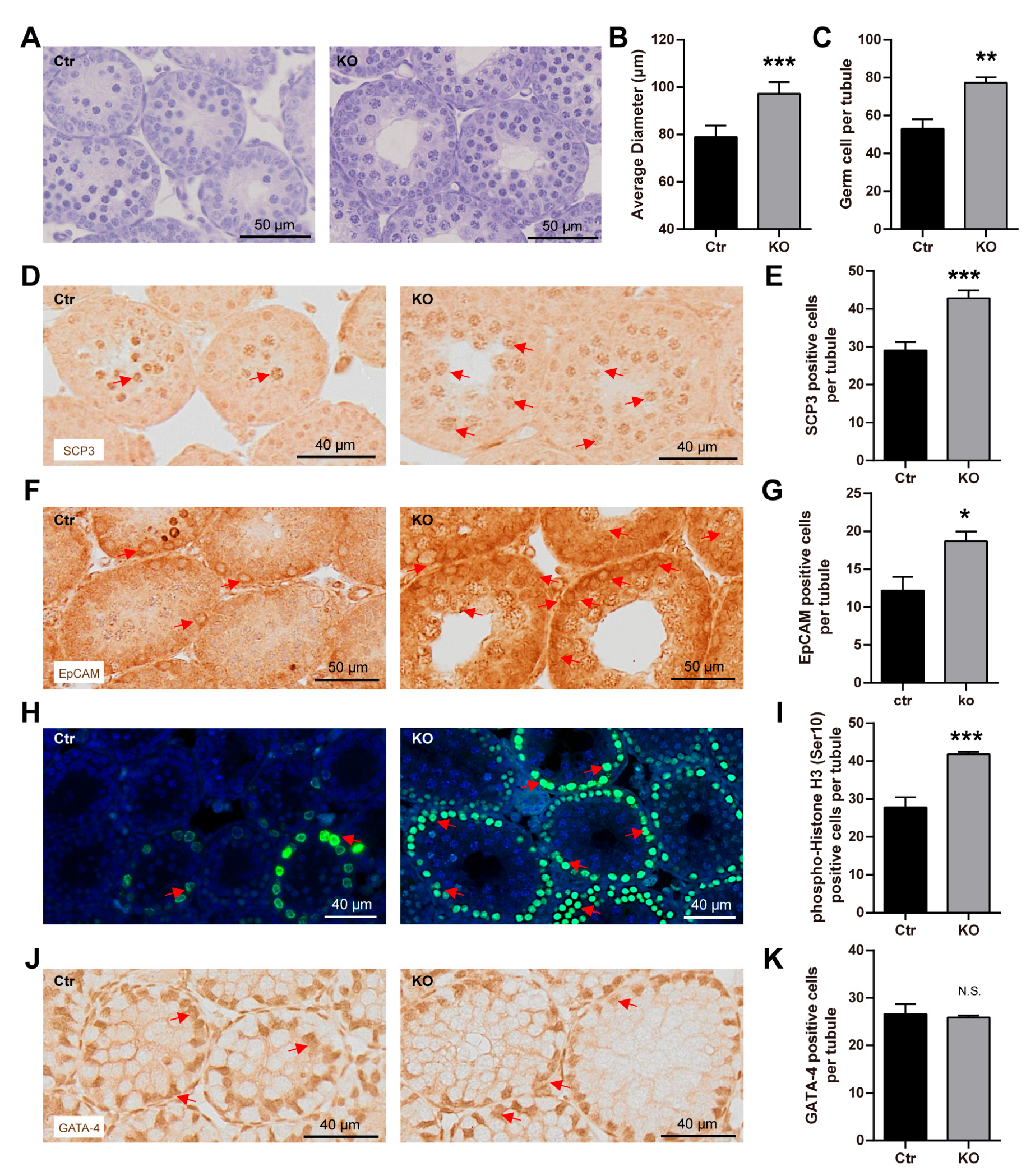
3.4. The Loss of LanCL2 Results in the Pre-Maturation of Spermatogonia
3.5. LanCL2 Is Important for Acrosomal Maturation in Spermiogenesis
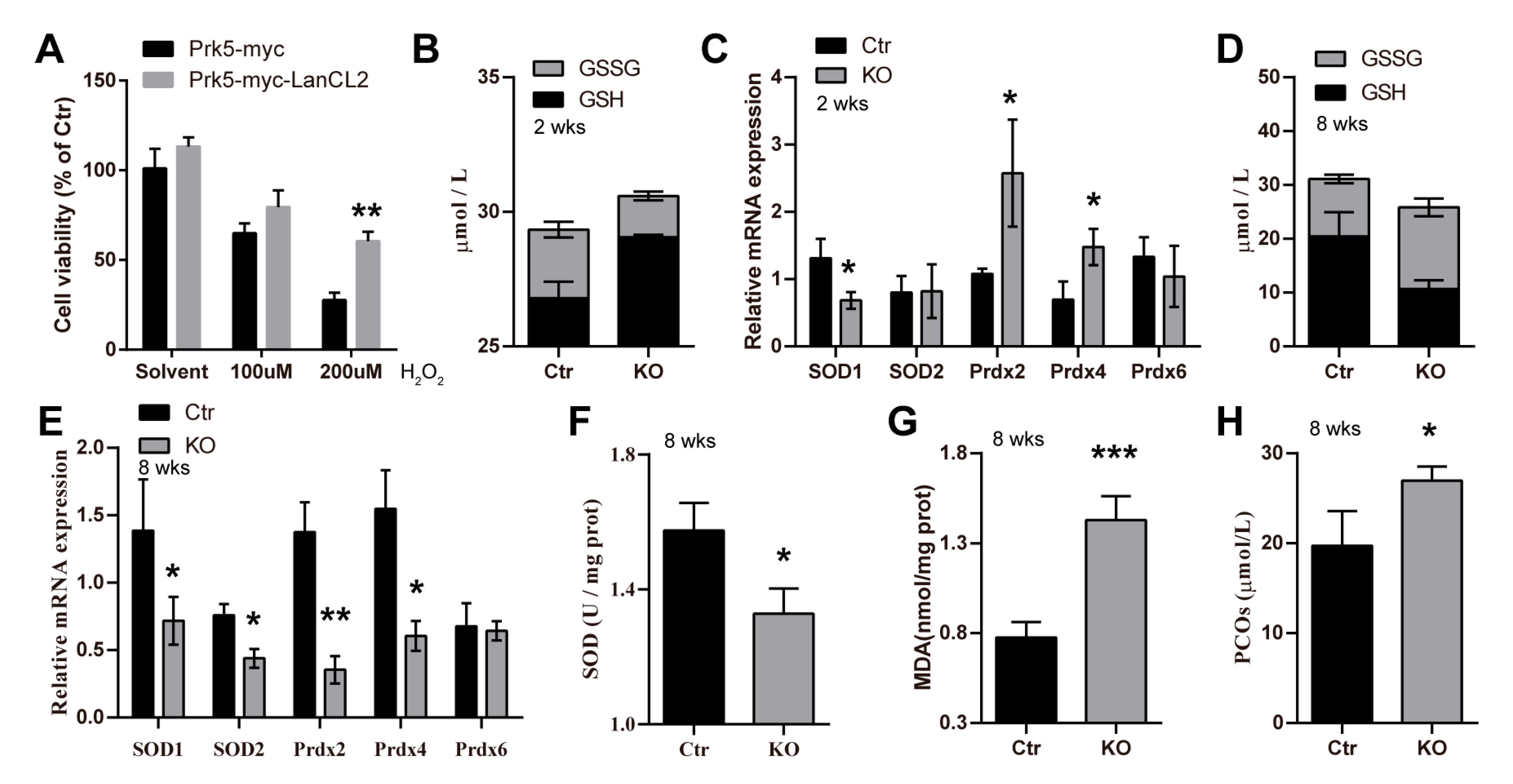
3.6. LanCL2 Is Implicated in Regulating Spermatogenic Redox Homeostasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, C.W.; Bowles, J.; Koopman, P. Control of mammalian germ cell entry into meiosis. Mol. Cell. Endocrinol. 2014, 382, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis. Curr. Biol. CB 2017, 27, R988–R994. [Google Scholar] [CrossRef]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The role of mitochondria in energy production for human sperm motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Baker, M.A. Oxidative stress, sperm survival and fertility control. Mol. Cell. Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Roman, S.D. Antioxidant systems and oxidative stress in the testes. Oxidative Med. Cell. Longev. 2008, 1, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Trocchia, S.; Abdel-Gawad, F.K.; Ciarcia, G. Roles of reactive oxygen species in the spermatogenesis regulation. Front. Endocrinol. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Imai, H.J.S. Redox reactions in mammalian spermatogenesis and the potential targets of reactive oxygen species under oxidative stress. Spermatogenesis 2014, 4, e979108. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Adwas, A.A.; Elsayed, A.S.I.; Azab, A.E.; Quwaydir, F.A.J.J.o.B. Oxidative stress and antioxidant mechanisms in human body. J. Biotechnol. 2019, 6, 43–47. [Google Scholar]
- Gong, S.; San Gabriel, M.C.; Zini, A.; Chan, P.; O’Flaherty, C. Low amounts and high thiol oxidation of peroxiredoxins in spermatozoa from infertile men. J. Androl. 2012, 33, 1342–1351. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Lu, J.; Tang, Q.; Zhang, S.; Yuan, B.; Li, J.; Di, W.; Sun, H.; Lu, C.; Xia, Y.; et al. GSTM1 and GSTT1 null polymorphisms and male infertility risk: An updated meta-analysis encompassing 6934 subjects. Sci. Rep. 2013, 3, 2258. [Google Scholar] [CrossRef] [PubMed]
- Naghavi, A.; Fazeli, F.; Salimi, S.; Nemati, B.M. Glutathione-S-transferase P1 Ile105Val polymorphism and idiopathic male infertility. Eur. Urol. Suppl. 2013, 12, e1133-S25. [Google Scholar] [CrossRef]
- Faure, C.; Leveille, P.; Dupont, C.; Julia, C.; Chavatte-Palmer, P.; Alifert, G.; Sutton, A.; Levy, R. Are superoxide dismutase 2 and nitric oxide synthase polymorphisms associated with idiopathic infertility? Antioxid. Redox Signal. 2014, 21, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Gu, A.; Wang, Y.; Huang, C.; Hu, F.; Zhou, Y.; Song, L.; Wang, X. Genetic variants in antioxidant genes are associated with sperm DNA damage and risk of male infertility in a Chinese population. Free. Radic. Biol. Med. 2012, 52, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Matsuki, S.; Iuchi, Y.; Okada, F.; Toyosaki, S.; Tomita, Y.; Ikeda, Y.; Fujii, J. Accelerated impairment of spermatogenic cells in SOD1-knockout mice under heat stress. Free. Radic. Res. 2005, 39, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.B.; Baker, M.A.; Connaughton, H.S.; Habenicht, U.; Aitken, R.J. Functional deletion of Txndc2 and Txndc3 increases the susceptibility of spermatozoa to age-related oxidative stress. Free. Radic. Biol. Med. 2013, 65, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.S.; Gargano, M.; Cao, J.; Bronson, R.T.; Heimler, I.; Hutz, R.J. Reduced fertility in female mice lacking copper-zinc superoxide dismutase. J. Biol. Chem. 1998, 273, 7765–7769. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, L.M.; Jonsson, J.; Edlund, T.; Marklund, S.L. Mice lacking extracellular superoxide dismutase are more sensitive to hyperoxia. Proc. Natl. Acad. Sci. USA 1995, 92, 6264–6268. [Google Scholar] [CrossRef]
- Ho, Y.S.; Magnenat, J.L.; Bronson, R.T.; Cao, J.; Gargano, M.; Sugawara, M.; Funk, C.D. Mice deficient in cellular glutathione peroxidase develop normally and show no increased sensitivity to hyperoxia. J. Biol. Chem. 1997, 272, 16644–16651. [Google Scholar] [CrossRef]
- Neumann, C.A.; Krause, D.S.; Carman, C.V.; Shampa, D.; Dubey, D.P.; Abraham, J.L.; Bronson, R.T.; Yuko, F.; Orkin, S.H.; Van Etten, R.A. Essential role for the peroxiredoxin Prdx1 in erythrocyte antioxidant defence and tumour suppression. Nature 2003, 424, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Argyropoulou, V.; Goemaere, J.; Clippe, A.; Lefort, C.; Tissir, F.; Schakman, O.; Gailly, P.; Ahn, M.-T.; Guiot, Y.; Galant, C. Peroxiredoxin-5 as a Novel Actor in Inflammation and Tumor Suppression. Free. Radic. Biol. Med. 2016, 100, S92. [Google Scholar] [CrossRef]
- Henderson, C.J.; Smith, A.G.; Ure, J.; Brown, K.; Bacon, E.J.; Wolf, C.R. Increased skin tumorigenesis in mice lacking pi class glutathione S-transferases. Proc. Natl. Acad. Sci. USA 1998, 95, 5275–5280. [Google Scholar] [CrossRef] [PubMed]
- Engle, M.R.; Singh, S.P.; Czernik, P.J.; Gaddy, D.; Montague, D.C.; Ceci, J.D.; Yang, Y.; Awasthi, S.; Awasthi, Y.C.; Zimniak, P. Physiological role of mGSTA4-4, a glutathione S-transferase metabolizing 4-hydroxynonenal: Generation and analysis of mGsta4 null mouse. Toxicol. Appl. Pharmacol. 2004, 194, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.E.; Matthaei, K.I.; Blackburn, A.C.; Davis, R.P.; Dahlstrom, J.E.; Koina, M.E.; Anders, M.W.; Board, P.G. Mice deficient in glutathione transferase zeta/maleylacetoacetate isomerase exhibit a range of pathological changes and elevated expression of alpha, mu, and pi class glutathione transferases. Am. J. Pathol. 2004, 165, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Chen, M.; Pang, D.; Bi, D.; Zou, Y.; Xia, X.; Yang, W.; Luo, L.; Deng, R.; Tan, H.; et al. Developmental and Activity-Dependent Expression of LanCL1 Confers Antioxidant Activity Required for Neuronal Survival. Dev. Cell 2014, 30, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Yang, C.; Pang, D.; Li, C.; Gong, H.; Cao, X.; He, X.; Chen, X.; Mu, B.; Cui, Y.; et al. Animal models of male subfertility targeted on LanCL1-regulated spermatogenic redox homeostasis. Lab Anim. 2022, 51, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Fresia, C.; Vigliarolo, T.; Guida, L.; Booz, V.; Bruzzone, S.; Sturla, L.; Di Bona, M.; Pesce, M.; Usai, C.; De Flora, A.; et al. G-protein coupling and nuclear translocation of the human abscisic acid receptor LANCL2. Sci. Rep. 2016, 6, 26658. [Google Scholar] [CrossRef]
- Park, S.; James, C.D. Lanthionine synthetase components C-like 2 increases cellular sensitivity to adriamycin by decreasing the expression of P-glycoprotein through a transcription-mediated mechanism. Cancer Res. 2003, 63, 723–727. [Google Scholar] [PubMed]
- Sturla, L.; Fresia, C.; Guida, L.; Grozio, A.; Vigliarolo, T.; Mannino, E.; Millo, E.; Bagnasco, L.; Bruzzone, S.; De Flora, A.; et al. Binding of abscisic acid to human LANCL2. Biochem. Biophys. Res. Commun. 2011, 415, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Van Der Donk, W.A.; Chen, J. Lanthionine synthetase C–like protein 2 (LanCL2) is a novel regulator of Akt. Mol. Biol. Cell 2014, 25, 3954–3961. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Biggers, J.D.; Whitten, W.K.; Whittingham, D.G. The culture of mouse embryos in vitro. In Methods in Mammalian Embryology; W. H. Freeman & Co., Ltd.: San Francisco, CA, USA, 1971. [Google Scholar]
- Soria-Tiedemann, M.; Michel, G.; Urban, I.; Aldrovandi, M.; O’Donnell, V.B.; Stehling, S.; Kuhn, H.; Borchert, A. Unbalanced Expression of Glutathione Peroxidase 4 and Arachidonate 15-Lipoxygenase Affects Acrosome Reaction and In Vitro Fertilization. Int. J. Mol. Sci. 2022, 23, 9907. [Google Scholar] [CrossRef] [PubMed]
- Mayer, H.; Pongratz, M.; Prohaska, R. Molecular cloning, characterization, and tissue-specific expression of human LANCL2, a novel member of the LanC-like protein family. DNA Seq. J. DNA Seq. Mapp. 2001, 12, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Griswold, M.D. Spermatogenesis: The Commitment to Meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ernst, C.; Eling, N.; Martinez-Jimenez, C.P.; Marioni, J.C.; Odom, D.T. Staged developmental mapping and X chromosome transcriptional dynamics during mouse spermatogenesis. Nat. Commun. 2019, 10, 1251. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, S.G. Testicular biopsy score count--a method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Hormones 1970, 1, 2–25. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Freinkman, E.; de Rooij, D.G.; Page, D.C. Periodic production of retinoic acid by meiotic and somatic cells coordinates four transitions in mouse spermatogenesis. Proc. Natl. Acad. Sci. USA 2017, 114, E10132–E10141. [Google Scholar] [CrossRef]
- O’Donnell, L. Mechanisms of spermiogenesis and spermiation and how they are disturbed. Spermatogenesis 2014, 4, e979623. [Google Scholar] [CrossRef]
- Huang, C.; Gong, H.; Mu, B.; Lan, X.; Yang, C.; Tan, J.; Liu, W.; Zou, Y.; Li, L.; Feng, B.; et al. BAF-L Modulates Histone-to-Protamine Transition during Spermiogenesis. Int. J. Mol. Sci. 2022, 23, 1985. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Iwata, K.; Ogonuki, N.; Inoue, K.; Atsuo, O.; Kanatsu-Shinohara, M.; Morimoto, T.; Yabe-Nishimura, C.; Shinohara, T. ROS are required for mouse spermatogonial stem cell self-renewal. Cell Stem Cell 2013, 12, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Kanastu-Shinohara, M.; Ogonuki, N.; Kamimura, S.; Ogura, A.; Yabe-Nishimura, C.; Mori, Y.; Morimoto, T.; Watanabe, S.; Otsu, K.; et al. ROS amplification drives mouse spermatogonial stem cell self-renewal. Life Sci. Alliance 2019, 2. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, H.; Kanatsu-Shinohara, M.; Shinohara, T. ROS-Generating Oxidase Nox3 Regulates the Self-Renewal of Mouse Spermatogonial Stem Cells. Biol. Reprod. 2015, 92, 147. [Google Scholar] [CrossRef]
- de Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13 (Suppl. S1), 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fallah, H.P.; Tovo-Neto, A.; Yeung, E.C.; Nóbrega, R.H.; Habibi, H.R. Paracrine/autocrine control of spermatogenesis by gonadotropin-inhibitory hormone. Mol. Cell. Endocrinol. 2019, 492, 110440. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-M.; Li, Z.-F.; Yang, W.-X.; Tan, F.-Q. Follicle-stimulating hormone signaling in Sertoli cells: A licence to the early stages of spermatogenesis. Reprod. Biol. Endocrinol. 2022, 20, 97. [Google Scholar] [CrossRef] [PubMed]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Hansen, M.A.; Jørgensen, M.; Tanaka, M.; Almstrup, K.; Skakkebaek, N.E.; Leffers, H. Germ cell differentiation-dependent and stage-specific expression of LANCL1 in rodent testis. Eur. J. Histochem. 2003, 47, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Liao, J.; Qin, M.; Li, H.; Zhang, Q.; Chen, Y.; Cheng, J. Single-Cell RNAseq Resolve the Potential Effects of LanCL1 Gene in the Mouse Testis. Cells 2022, 11, 4135. [Google Scholar] [CrossRef] [PubMed]
- O, W.S.; Chen, H.; Chow, P.H. Male genital tract antioxidant enzymes--their ability to preserve sperm DNA integrity. Mol. Cell. Endocrinol. 2006, 250, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Khawar, M.B.; Gao, H.; Li, W. Mechanism of acrosome biogenesis in mammals. Front. Cell Dev. Biol. 2019, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-J.; Allam, J.-P.; Duan, Y.-G.; Haidl, G. Influence of reactive oxygen species on human sperm functions and fertilizing capacity including therapeutical approaches. Arch. Gynecol. Obstet. 2013, 288, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D. Physiological role of reactive oxygen species in male reproduction. In Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction; Elsevier: Amsterdam, The Netherlands, 2019; pp. 65–78. [Google Scholar]
- Sasagawa, I.; Matsuki, S.; Suzuki, Y.; Iuchi, Y.; Tohya, K.; Kimura, M.; Nakada, T.; Fujii, J. Possible involvement of the membrane-bound form of peroxiredoxin 4 in acrosome formation during spermiogenesis of rats. Eur. J. Biochem. 2001, 268, 3053–3061. [Google Scholar] [CrossRef]
- Iuchi, Y.; Okada, F.; Tsunoda, S.; Kibe, N.; Shirasawa, N.; Ikawa, M.; Okabe, M.; Ikeda, Y.; Fujii, J. Peroxiredoxin 4 knockout results in elevated spermatogenic cell death via oxidative stress. Biochem. J. 2009, 419, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Hakkaku, N.; Iwamoto, R.; Suzuki, J.; Suzuki, T.; Tajima, Y.; Konishi, K.; Minami, S.; Ichinose, S.; Ishizaka, K. Depletion of selenoprotein GPx4 in spermatocytes causes male infertility in mice. J. Biol. Chem. 2009, 284, 32522–32532. [Google Scholar] [CrossRef]
- Ursini, F.; Heim, S.; Kiess, M.; Maiorino, M.; Roveri, A.; Wissing, J.; Flohe, L. Dual function of the selenoprotein PHGPx during sperm maturation. Science 1999, 285, 1393–1396. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Aitken, R.J. The importance of redox regulated pathways in sperm cell biology. Mol. Cell. Endocrinol. 2004, 216, 47–54. [Google Scholar] [CrossRef]
- Ozkosem, B.; Feinstein, S.I.; Fisher, A.B.; O’Flaherty, C. Absence of Peroxiredoxin 6 Amplifies the Effect of Oxidant Stress on Mobility and SCSA/CMA3 Defined Chromatin Quality and Impairs Fertilizing Ability of Mouse Spermatozoa. Biol. Reprod. 2016, 94, 68. [Google Scholar] [CrossRef] [PubMed]
- Bumanlag, E.; Scarlata, E.; O’Flaherty, C. Peroxiredoxin 6 Peroxidase and Ca-Independent Phospholipase A Activities Are Essential to Support Male-Mouse Fertility. Antioxidants 2022, 11, 226. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| β-actin | F | 5′-AGA GGG AAA TCG TGC GTG AC-3′ |
| R | 5′-CAA TAG TGA TGA CCT GGC CGT-3′ | |
| LanCL2 | F | 5′-GTG TAG CGA TGT GAT TTG GC-3′ |
| R | 5′-AAT GCT GGA AAC CGT GAT GT-3′ | |
| Prdx2 | F | 5′-CAT TCC AGT TCT CGC TGA CA-3′ |
| R | 5′-GTT TTG TGA TGG GTC GAT GA-3′ | |
| Prdx4 | F | 5′-AGA GGA GTG CCA CTT CTA CG-3′ |
| R | 5′-GGA AAT CTT CGC TTT GCT TAG GT-3′ | |
| Prdx6 | F | 5′-GAC TCA TGG GGC ATT CTC TTC-3′ |
| R | 5′-CAA GCT CCC GAT TCC TAT CAT C-3′ | |
| SOD1 | F | 5′-AAC CAG TTG TGT TGT CAG GAC-3′ |
| R | 5′-CCA CCA TGT TTC TTA GAG TGA GG-3′ | |
| SOD2 | F | 5′-TGG ACA AAC CTG AGC CCT AAG-3′ |
| R | 5′-CCC AAA GTC ACG CTT GAT AGC-3′ | |
| FAM71F1 | F | 5′-ATG ATG ACA TCA GTT CCA CCT AGA AAG TC-3′ |
| R | 5′-TAT AGA GTT TCC TCC AGT TAG GGA CAG CC-3′ | |
| FAM209 | F | 5′-TGC CTG TTC TTG TCT CTG TG-3′ |
| R | 5′-TCA CCA ATT CCA TCT CGA GC-3′ |
| Antibodies | Source | Identifier/Application |
|---|---|---|
| EpCAM Rabbit mAb | ABclonal (Wuhan, China) | Cat#A19301, IHC: 1:500 |
| GATA-4(D3A3M) Rabbit mAb | Cell Signaling Technology (Danvers, MA, USA) | Cat#36966, IHC: 1:800 |
| ACRV1 Rabbit pAb | Proteintech (Wuhan, China) | Cat#14040-1-AP, IHC: 1:500 |
| SCP3 Rabbit pAb | Abcam (Cambridge, MA, USA) | Cat#ab15093, IHC: 1:500 |
| Phospho-Histone H3-S10 | ABclonal (Wuhan, China) | Cat#AP0002, IF: 1:200 |
| LanCL2 Rabbit pAb | Generated by immunizing rabbits with full-length GST fusion proteins of mouse LanCL2 | IHC: 1:250, WB: 1:50 |
| β-Actin Rabbit mAb | Abclonal (Wuhan, China) | Cat#AC026, WB: 1:100,000 |
| aRab-488 Alexa Fluor | Jackson ImmuneResearch (West Grove, PA, USA) | 711-547-003, IF: 1:50 |
| aRab-594 Alexa Fluor | Invitrogen (Carlsbad, CA,USA) | A-11037, IF: 1:500 |
| aGoat-488 Alexa Fluor | Invitrogen (Carlsbad, CA,USA) | A-11055, IF: 1:1000 |
| aM-594 Alexa Fluor | Invitrogen (Carlsbad, CA,USA) | A-21203, IF: 1:1000 |
| Score | Morphology |
|---|---|
| 10 | Complete spermatogenesis |
| 9 | Slightly damaged spermatogenesis, large number of late-stage sperm cells, and disorder of spermatogenic epithelium |
| 8 | Less than 5 sperm in the lumen and a small amount of late-stage sperm cells |
| 7 | Mo sperm or late-stage sperm cells in the lumen but a large number of early-stage sperm cells |
| 6 | No sperm or late-stage sperm cells in the lumen with a small amount of early-stage sperm cells |
| 5 | No sperm or sperm cells in the lumen but a large number of spermatocytes |
| 4 | No sperm or sperm cells in the lumen with a small amount of spermatocytes |
| 3 | Only spermatogonia in the lumen |
| 2 | Only Sertoli cells in the lumen and no germ cells |
| 1 | No seminiferous epithelium |
| Component | Concentration |
|---|---|
| NaCl | 95 mM |
| NaHCO3 | 25 mM |
| HEPES | 10 mM |
| Glucose | 5 mM |
| KCl | 4.8 mM |
| Lactic acid | 2.0 mM |
| CaCl2 | 1.3 mM |
| MgSO4 | 1.2 mM |
| KH2PO4 | 1.2 mM |
| Na-pyruvate | 0.25 mM |
| BSA | 3 mg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Wang, J.; Shi, S.; Lan, X.; Cheng, X.; Li, L.; Zou, Y.; Jia, L.; Liu, W.; Luo, Q.; et al. LanCL2 Implicates in Testicular Redox Homeostasis and Acrosomal Maturation. Antioxidants 2024, 13, 534. https://doi.org/10.3390/antiox13050534
Zhao Y, Wang J, Shi S, Lan X, Cheng X, Li L, Zou Y, Jia L, Liu W, Luo Q, et al. LanCL2 Implicates in Testicular Redox Homeostasis and Acrosomal Maturation. Antioxidants. 2024; 13(5):534. https://doi.org/10.3390/antiox13050534
Chicago/Turabian StyleZhao, Yanling, Jichen Wang, Shuai Shi, Xinting Lan, Xiangyu Cheng, Lixia Li, Yuanfeng Zou, Lanlan Jia, Wentao Liu, Qihui Luo, and et al. 2024. "LanCL2 Implicates in Testicular Redox Homeostasis and Acrosomal Maturation" Antioxidants 13, no. 5: 534. https://doi.org/10.3390/antiox13050534
APA StyleZhao, Y., Wang, J., Shi, S., Lan, X., Cheng, X., Li, L., Zou, Y., Jia, L., Liu, W., Luo, Q., Chen, Z., & Huang, C. (2024). LanCL2 Implicates in Testicular Redox Homeostasis and Acrosomal Maturation. Antioxidants, 13(5), 534. https://doi.org/10.3390/antiox13050534