
Hydrogen Sulfide, Oxidative Stress and Periodontal Diseases: A Concise Review


Abstract
:1. Introduction
2. H2S—Production

3. H2S Biological Roles

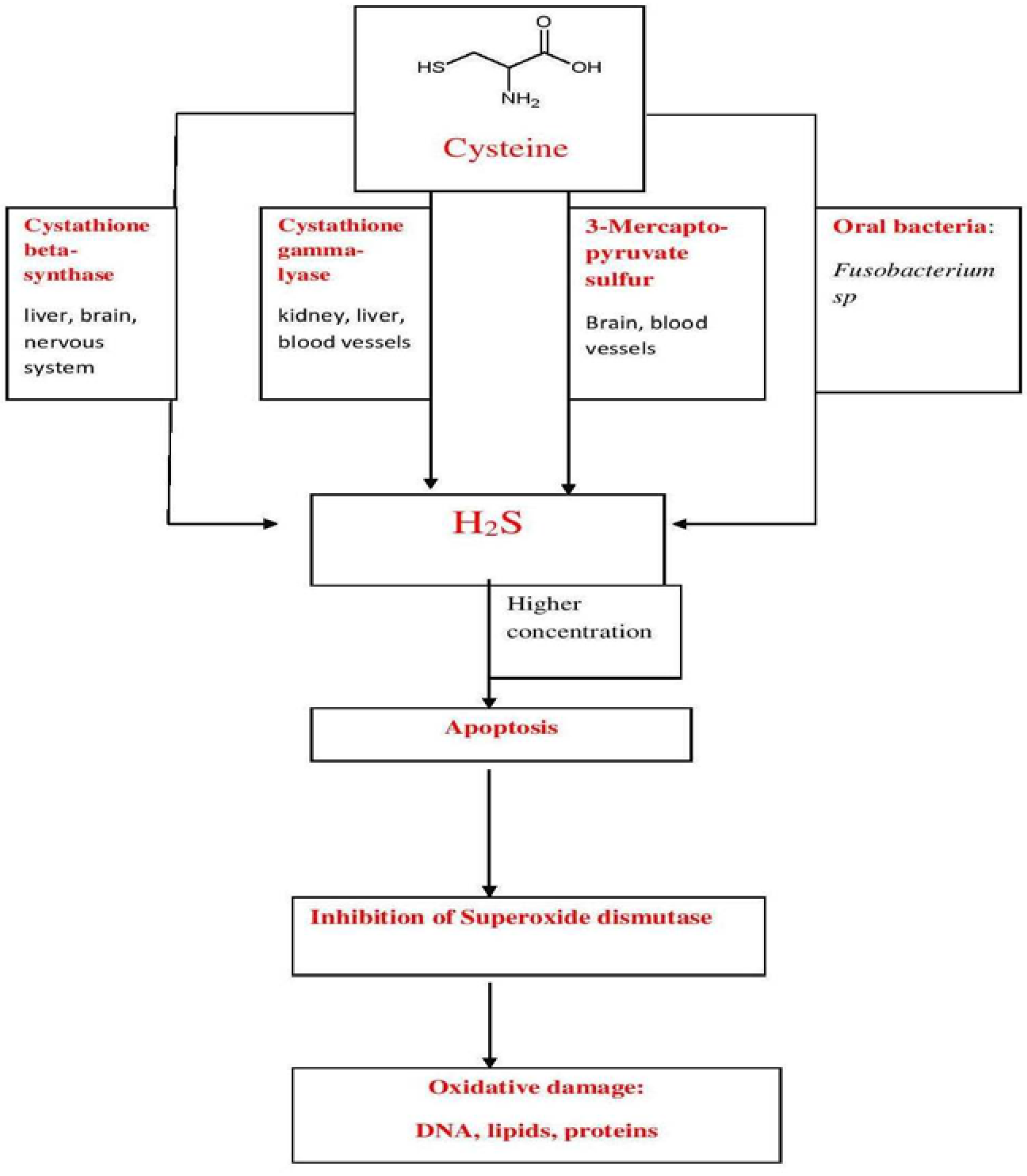{kind=link}
| Biological Event | H2S—Effect |
|---|---|
| Angiogenesis | Increases blood flow Decreases the risk of tissue injury |
| Mitochondrial respiration | Decreases the function Cytoprotection |
| Vasodilatation | Regulates blood pressure |
| Leukocyte adhesion | Anti-inflammatory effect |
| Apoptosis | Decreases apoptosis—cytoprotective effect |
| Antioxidant | Up-regulation of antioxidant molecules |
| Tissue | Cells | Origin | Biological Event |
|---|---|---|---|
| Oral Epithelia | Normal keratinocytes | Ca9-22 cell line | Apoptosis—mitochondrial pathway activated; DNA damage |
| Keratinocyte stem cells | Human skin cell line | Apoptosis—mitochondrial pathway activated; DNA damage; p53 and Bax activity increased | |
| Keratinocyte stem cells | Human oral mucosa | Apoptosis—mitochondrial pathway activated; DNA damage; Activation of genes from p53 pathway connected with DNA repair, cell cycle arrest | |
| Keratinocyte cells | Animal oral mucosa | Increases the permeability of the epithelium | |
| Oral Dermis | Fibroblasts | Human oral mucosa | Apoptosis—mitochondrial pathway activated; DNA damage |
| Collagen Fibers | Extracellular matrix | Increases collagen degradation/decreases collagen synthesis | |
| Dental Pulp | Dental pulp stem cells | Human dental pulp | Apoptosis—mitochondrial pathway activated; DNA damage |
| Bone | Osteoblasts | Mouse calvaria | Apoptosis—mitochondrial and death ligand pathway activated; DNA damage; Bone resorption |
| Tissue/Cells | Origin | Biological Event | |
|---|---|---|---|
| Dental Pulp | Dental pulp cells | Human pulp | Differentiation to hepatic like cells |
| Dental pulp cells | Human pulp | Differentiation of pancreatic like cells | |
| Bone | Osteoclasts | Mouse | Osteoclast activation followed by bone resorption |
4. Oxidative Stress and Periodontal Diseases
5. H2S—Involvement in Periodontal Diseases
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ju, Y.; Zhang, W.; Pei, Y.; Yang, G. H2S signaling in redox regulation of cellular functions. Can. J. Physiol. Pharmacol. 2013, 91, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Edlund, M.B.; Claesson, R.; Carlsson, J. The formation of hydrogen sulfide and methyl mercaptan by oral bacteria. Oral Microbiol. Immunol. 1990, 5, 195–201. [Google Scholar] [CrossRef]
- Persson, S. Hydrogen sulfide and methyl mercaptan in periodontal pockets. Oral Microbiol. Immunol. 1992, 7, 378–379. [Google Scholar] [CrossRef] [PubMed]
- Biju, T.; Shabeer, M.M.; Amitha, R.; Rajendra, B.P.; Suchetha, K. Comparative evaluation of serum superoxide dismutase and glutathione levels in periodontally diseased patients: An interventional study. Indian J. Dent. Res. 2014, 25, 613–616. [Google Scholar] [PubMed]
- Mancini, S.; Romanelli, R.; Laschinger, C.A.; Overall, C.M.; Sodek, J.; McCulloch, C.A. Assessment of a novel screening test for neutrophil collagenase activity in the diagnosis of periodontal diseases. J. Periodontol. 1999, 70, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Vacek, T.P.; Gillespie, W.; Tyagi, N.; Vacek, J.C.; Tyagi, S.C. Hydrogen sulfide protects against vascular remodeling from endothelial damage. Amino Acids 2010, 39, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Calvert, J.W.; Jha, S.; Gundewar, S.; Elrod, J.W.; Ramachandran, A.; Pattillo, C.B.; Kevil, C.G.; Lefer, D.J. Hydrogen sulfide mediates cardioprotection through Nrf2 signaling. Circ. Res. 2009, 105, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, W.; Chen, Q.; Jiang, Y.; Lu, X.; Zhao, X. Hydrogen sulfide accelerates wound healing in diabetic rats. Int. J. Clin. Exp. Pathol. 2015, 8, 5097–5104. [Google Scholar] [PubMed]
- Zanardo, R.C.; Brancaleone, V.; Distrutti, E.; Fiorucci, S.; Cirino, G.; Wallace, J.L. Hydrogen sulfide is an endogenous modulator of leukocyte-mediated inflammation. FASEB J. 2006, 20, 2118–2120. [Google Scholar] [CrossRef] [PubMed]
- Predmore, B.L.; Lefer, D.J.; Gojon, G. Hydrogen sulfide in biochemistry and medicine. Antioxid. Redox Signal. 2012, 17, 119–140. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen sulfide and polysulfides as signaling molecules. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2015, 91, 131–159. [Google Scholar] [CrossRef] [PubMed]
- Kasparek, M.S.; Linden, D.R.; Kreis, M.E.; Sarr, M.G. Gasotransmitters in the gastrointestinal tract. Surgery 2008, 143, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.; Palinkas, Z.; Nagy, A.; Budai, B.; Toth, I.; Vasas, A. Chemical aspects of hydrogen sulfide measurements in physiological samples. Biochim. Biophys. Acta 2014, 1840, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, D.; Liu, Y.; Zhang, C.; Wang, J.; Wang, S. Physiologic levels of endogenous hydrogen sulfide maintain the proliferation and differentiation capacity of periodontal ligament stem cells. J. Periodontol. 2015, 86, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, P.A.; Johnson, P.W. The relationship between oral malodor, gingivitis, and periodontitis. A review. J. Periodontol. 1999, 70, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Sigusch, B.W. Periodontitis as manifestation of Crohn’s disease in primary dentition: A case report. J. Dent. Child. 2004, 71, 193–196. [Google Scholar]
- Calvert, J.W.; Coetzee, W.A.; Lefer, D.J. Novel insights into hydrogen sulfide—Mediated cytoprotection. Antioxid. Redox Signal. 2010, 12, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Pae, H.O.; Lee, Y.C.; Jo, E.K.; Chung, H.T. Subtle interplay of endogenous bioactive gases (NO, CO and H2S) in inflammation. Arch. Pharm. Res. 2009, 32, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; Blackler, R.W.; Chan, M.V.; Da Silva, G.J.; Elsheikh, W.; Flannigan, K.L.; Gamaniek, I.; Manko, A.; Wang, L.; Motta, J.P.; et al. Anti-inflammatory and cytoprotective actions of hydrogen sulfide: Translation to therapeutics. Antioxid. Redox Signal. 2015, 22, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Hadadha, M.; Vakili, A.; Bandegi, A.R. Effect of the inhibition of hydrogen sulfide synthesis on ischemic injury and oxidative stress biomarkers in a transient model of focal cerebral ischemia in rats. J. Stroke Cerebrovasc. Dis. 2015, 24, 2676–2684. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhao, T.; Yuan, Y.; Hu, N.; Tang, X. Hydrogen sulfide (H2S) attenuates uranium-induced acute nephrotoxicity through oxidative stress and inflammatory response via Nrf2-NF-kB pathways. Chem. Biol. Interact. 2015, 242, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Olah, G.; Szczesny, B.; Wood, M.E.; Whiteman, M.; Szabo, C. Ap39, a mitochondrially-targeted hydrogen sulfide donor, exerts protective effects in renal epithelial cells subjected to oxidative stress in vitro and in acute renal injury in vivo. Shock 2016, 45, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Shibuya, N.; Kimura, Y. Hydrogen sulfide is a signaling molecule and a cytoprotectant. Antioxid. Redox Signal. 2012, 17, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rose, P.; Moore, P.K. Hydrogen sulfide and cell signaling. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Huertas, J.R.; Palomino, N.; Ochoa, J.J.; Quiles, J.L.; Ramirez-Tortosa, M.C.; Battino, M.; Robles, R.; Mataix, J. Lipid peroxidation and antioxidants in erythrocyte membranes of full-term and preterm newborns. BioFactors 1998, 8, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Miricescu, D.; Totan, A.; Calenic, B.; Mocanu, B.; Didilescu, A.; Mohora, M.; Spinu, T.; Greabu, M. Salivary biomarkers: Relationship between oxidative stress and alveolar bone loss in chronic periodontitis. Acta Odontol. Scand. 2014, 72, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Greabu, M.; Calenic, B. Salivary biomarkers of oxidative stress associated with periodontal diseases. In Studies on Periodontal Disease; Ekuni, D., Battino, M., Tomofuji, T., Putnins, E.E., Eds.; Springer: New York, NY, USA, 2014; pp. 329–343. [Google Scholar]
- Singer, R.E.; Moss, K.; Kim, S.J.; Beck, J.D.; Offenbacher, S. Oxidative stress and IgG antibody modify periodontitis-CRP association. J. Dent. Res. 2015, 94, 1698–1705. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Gornitsky, M.; Velly, A.M.; Yu, H.; Benarroch, M.; Schipper, H.M. Salivary DNA, lipid, and protein oxidation in nonsmokers with periodontal disease. Free Radic. Biol. Med. 2009, 46, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Mohora, M.; Greabu, M.; Totan, A.; Mitrea, N.; Battino, M. Redox-sensitive signaling factors and antioxidants. Farmacia 2009, 57, 399–411. [Google Scholar]
- Totan, A.; Miricescu, D.; Parlatescu, I.; Mohora, M.; Greabu, M. Possible salivary and serum biomarkers for oral lichen planus. Biotechnic. Histochem. 2015, 90, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Miricescu, D.; Greabu, M.; Totan, A.; Mohora, M.; Didilescu, A.; Mitrea, N.; Arsene, A.; Spinu, T.; Totan, C.; Rădulescu, R. Oxidative stress—A possible link between systemic and oral diseases. Farmacia 2011, 59, 329–337. [Google Scholar]
- Jenifer, H.D.; Bhola, S.; Kalburgi, V.; Warad, S.; Kokatnur, V.M. The influence of cigarette smoking on blood and salivary super oxide dismutase enzyme levels among smokers and nonsmokers—A cross sectional study. J. Tradit. Complement. Med. 2015, 5, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Chapple, I.L.; Mason, G.I.; Garner, I.; Matthews, J.B.; Thorpe, G.H.; Maxwell, S.R.; Whitehead, T.P. Enhanced chemiluminescent assay for measuring the total antioxidant capacity of serum, saliva and crevicular fluid. Ann. Clin. Biochem. 1997, 34, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Tonguc, M.O.; Ozturk, O.; Sutcu, R.; Ceyhan, B.M.; Kilinc, G.; Sonmez, Y.; Yetkin Ay, Z.; Sahin, U.; Baltacioglu, E.; Kirzioglu, F.Y. The impact of smoking status on antioxidant enzyme activity and malondialdehyde levels in chronic periodontitis. J. Periodontol. 2011, 82, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Sculley, D.V.; Langley-Evans, S.C. Salivary antioxidants and periodontal disease status. Proc. Nutr. Soc. 2002, 61, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Nagler, R.M.; Klein, I.; Zarzhevsky, N.; Drigues, N.; Reznick, A.Z. Characterization of the differentiated antioxidant profile of human saliva. Free Radic. Biol. Med. 2002, 32, 268–277. [Google Scholar] [CrossRef]
- Amerongen, A.V.; Veerman, E.C. Saliva—The defender of the oral cavity. Oral Dis. 2002, 8, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Liskmann, S.; Vihalemm, T.; Salum, O.; Zilmer, K.; Fischer, K.; Zilmer, M. Characterization of the antioxidant profile of human saliva in peri-implant health and disease. Clin. Oral Implant. Res. 2007, 18, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, I.; Kohen, R.; Shalish, M.; Varon, D.; Shai, E.; Koren, E. The oxidant-scavenging abilities in the oral cavity may be regulated by a collaboration among antioxidants in saliva, microorganisms, blood cells and polyphenols: A chemiluminescence-based study. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Wu, Y.; Shu, R.; Luo, L.J.; Ge, L.H.; Xie, Y.F. Initial comparison of proteomic profiles of whole unstimulated saliva obtained from generalized aggressive periodontitis patients and healthy control subjects. J. Periodontal Res. 2009, 44, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, R.; Pandurang, P.; Kamath, S.U.; Goyal, R.; Ballal, S.; Shanbhogue, A.Y.; Kamath, U.; Bhat, G.S.; Bhat, K.M. Association of cigarette smoking with superoxide dismutase enzyme levels in subjects with chronic periodontitis. J. Periodontol. 2009, 80, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Canakci, C.F.; Cicek, Y.; Canakci, V. Reactive oxygen species and human inflammatory periodontal diseases. Biochemistry 2005, 70, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.T. Salivary diagnostics powered by nanotechnologies, proteomics and genomics. JADA 2006, 137, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Garon, E.; Wong, D. Salivary diagnostics. Orthod. Craniofac. Res. 2009, 12, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef]
- Matsuyama, T.; Kawai, T.; Izumi, Y.; Taubman, M.A. Expression of major histocompatibility complex class II and CD80 by gingival epithelial cells induces activation of CD4+ T cells in response to bacterial challenge. Infect. Immun. 2005, 73, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.; Haake, S.K.; Kim, J.W.; Park, N.H. Differential expression of interleukin-8 and intercellular adhesion molecule-1 by human gingival epithelial cells in response to Actinobacillus actinomycetemcomitans or Porphyromonas gingivalis infection. Oral Microbiol. Immunol. 1998, 13, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Asai, Y.; Ohyama, Y.; Gen, K.; Ogawa, T. Bacterial fimbriae and their peptides activate human gingival epithelial cells through toll-like receptor 2. Infect. Immun. 2001, 69, 7387–7395. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, I.; Hosokawa, Y.; Komatsuzawa, H.; Goncalves, R.B.; Karimbux, N.; Napimoga, M.H.; Seki, M.; Ouhara, K.; Sugai, M.; Taubman, M.A.; et al. Innate immune peptide LL-37 displays distinct expression pattern from beta-defensins in inflamed gingival tissue. Clin. Exp. Immunol. 2006, 146, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Almerich-Silla, J.M.; Montiel-Company, J.M.; Pastor, S.; Serrano, F.; Puig-Silla, M.; Dasi, F. Oxidative stress parameters in saliva and its association with periodontal disease and types of bacteria. Dis. Markers 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Oktay, S.; Chukkapalli, S.S.; Rivera-Kweh, M.F.; Velsko, I.M.; Holliday, L.S.; Kesavalu, L. Periodontitis in rats induces systemic oxidative stress that is controlled by bone-targeted antiresorptives. J. Periodontol. 2015, 86, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Golz, L.; Memmert, S.; Rath-Deschner, B.; Jager, A.; Appel, T.; Baumgarten, G.; Gotz, W.; Frede, S. LPS from P. gingivalis and hypoxia increases oxidative stress in periodontal ligament fibroblasts and contributes to periodontitis. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, Y.; Song, Y.; Zhang, X.; Wang, S.; Wang, Z. Systemic oxidative stress biomarkers in chronic periodontitis: A meta-analysis. Dis. Markers 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sree, S.L.; Sethupathy, S. Evaluation of the efficacy of taurine as an antioxidant in the management of patients with chronic periodontitis. Dent. Res. J. 2014, 11, 228–233. [Google Scholar]
- Wu, W.; Yang, N.; Feng, X.; Sun, T.; Shen, P.; Sun, W. Effect of vitamin C administration on hydrogen peroxide-induced cytotoxicity in periodontal ligament cells. Mol. Med. Rep. 2015, 11, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Ghallab, N.A.; Hamdy, E.; Shaker, O.G. Malondialdehyde, superoxide dismutase and melatonin levels in GCF of aggressive and chronic periodontitis patients. Aust. Dent. J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.; Rao, A.; Prasad, B.R.; Kumari, S. Serum levels of antioxidants and superoxide dismutase in periodontitis patients with diabetes type 2. J. Indian Soc. Periodontol. 2014, 18, 451–455. [Google Scholar] [PubMed]
- Thomas, B.; Madani, S.M.; Prasad, B.R.; Kumari, S. Comparative evaluation of serum antioxidant levels in periodontally diseased patients: An interventional study. Contemp. Clin. Dent. 2014, 5, 340–344. [Google Scholar] [PubMed]
- Monea, A.; Mezei, T.; Popsor, S.; Monea, M. Oxidative stress: A link between diabetes mellitus and periodontal disease. Int. J. Endocrinol. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Pushparani, D.S.; Anandan, S.N.; Theagarayan, P. Serum zinc and magnesium concentrations in type 2 diabetes mellitus with periodontitis. J. Indian Soc. Periodontol. 2014, 18, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Gurav, A.N. The association of periodontitis and metabolic syndrome. Dent. Res. J. 2014, 11, 1–10. [Google Scholar]
- Barnes, V.M.; Kennedy, A.D.; Panagakos, F.; Devizio, W.; Trivedi, H.M.; Jonsson, T.; Guo, L.; Cervi, S.; Scannapieco, F.A. Global metabolomic analysis of human saliva and plasma from healthy and diabetic subjects, with and without periodontal disease. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.F.; Ho, K.Y.; Ho, Y.P.; Wu, Y.M.; Yang, Y.H.; Tsai, C.C. The investigation of glutathione peroxidase, lactoferrin, myeloperoxidase and interleukin-1beta in gingival crevicular fluid: Implications for oxidative stress in human periodontal diseases. J. Periodontal Res. 2004, 39, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Miricescu, D.; Greabu, M.; Totan, A.; Didilescu, A.; Rădulescu, R. The antioxidant potential of saliva: Clinical significance in oral diseases. Ther. Pharmacol. Clin. Toxicol. 2011, 15, 139–143. [Google Scholar]
- Kanzaki, H.; Shinohara, F.; Kajiya, M.; Kodama, T. The Keap1/Nrf2 protein axis plays a role in osteoclast differentiation by regulating intracellular reactive oxygen species signaling. J. Biol. Chem. 2013, 288, 23009–23020. [Google Scholar] [CrossRef] [PubMed]
- Alagl, A.S.; Bhat, S.G. Ascorbic acid: New role of an age-old micronutrient in the management of periodontal disease in older adults. Geriatr. Gerontol. Int. 2015, 15, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Mendi, A.; Aslim, B. Antioxidant lactobacilli could protect gingival fibroblasts against hydrogen peroxide: A preliminary in vitro study. Probiotics Antimicrob. Proteins 2014, 6, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Roman-Malo, L.; Marin-Aguilar, F.; Alvarez-Suarez, J.M.; Giampieri, F.; Battino, M.; Cordero, M.D. Lipophilic antioxidants prevent lipopolysaccharide-induced mitochondrial dysfunction through mitochondrial biogenesis improvement. Pharmacol. Res. 2015, 91, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.A. The possible role of hydrogen sulfide in human periodontal disease. I. Hydrogen sulfide production in periodontal pockets. Periodontics 1967, 5, 233–236. [Google Scholar] [PubMed]
- Morhart, R.E.; Mata, L.J.; Sinskey, A.J.; Harris, R.S. A microbiological and biochemical study of gingival crevice debris obtained from Guatemalan Mayan Indians. J. Periodontol. 1970, 41, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Folke, L.E. Hydrogen sulfide production in the periodontal environment. J. Periodontol. 1973, 44, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Kajiya, M.; Giro, G.; Ouhara, K.; Mackler, H.E.; Mawardi, H.; Boisvert, H.; Duncan, M.J.; Sato, K.; Kawai, T. Bacteria-derived hydrogen sulfide promotes IL-8 production from epithelial cells. Biochem. Biophys. Res. Commun. 2010, 391, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Ii, H.; Yaegaki, K.; Murata, T.; Sato, T.; Kamoda, T. Oral malodorous compound inhibits osteoblast proliferation. J. Periodontol. 2009, 80, 2028–2034. [Google Scholar] [CrossRef] [PubMed]
- Sivarajah, A.; Collino, M.; Yasin, M.; Benetti, E.; Gallicchio, M.; Mazzon, E.; Cuzzocrea, S.; Fantozzi, R.; Thiemermann, C. Anti-apoptotic and anti-inflammatory effects of hydrogen sulfide in a rat model of regional myocardial I/R. Shock 2009, 31, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Searcy, D.G.; Whitehead, J.P.; Maroney, M.J. Interaction of Cu, Zn superoxide dismutase with hydrogen sulfide. Arch. Biochem. Biophys. 1995, 318, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Calenic, B.; Yaegaki, K.; Kozhuharova, A.; Imai, T. Oral malodorous compound causes oxidative stress and p53-mediated programmed cell death in keratinocyte stem cells. J. Periodontol. 2010, 81, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Calenic, B.; Amann, A. Detection of volatile malodorous compounds in breath: Current analytical techniques and implications in human disease. Bioanalysis 2014, 6, 357–376. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, I.; Calenic, B.; Imai, T.; Ii, H.; Yaegaki, K. Oral malodorous compound causes caspase-8 and -9 mediated programmed cell death in osteoblasts. J. Periodontal Res. 2012, 47, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.S.; Coimbra, L.S.; da Silva, A.R.; Teixeira, S.A.; Costa, S.K.; Wallace, J.L.; Spolidorio, L.C.; Muscara, M.N. The H2S-releasing naproxen derivative, ATB-346, inhibits alveolar bone loss and inflammation in rats with ligature-induced periodontitis. Med. Gas. Res. 2015, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, T.; Yaegaki, K.; Qian, W.; Herai, M.; Calenic, B.; Imai, T.; Sato, T.; Tanaka, T.; Kamoda, T.; Ii, H. Hydrogen sulfide induces apoptosis in epithelial cells derived from human gingiva. J. Breath Res. 2008, 2. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Browne, G.; Melino, G.; Cohen, G.M. Ordering of caspases in cells undergoing apoptosis by the intrinsic pathway. Cell Death Differ. 2009, 16, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Pianotti, R.; Lachette, S.; Dills, S. Desulfuration of cysteine and methionine by fusobacterium nucleatum. J. Dent. Res. 1986, 65, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Yoshimura, M.; Ohara, N.; Yoshimura, S.; Nagashima, S.; Takehara, T.; Nakayama, K. Hydrogen sulfide production from cysteine and homocysteine by periodontal and oral bacteria. J. Periodontol. 2009, 80, 1845–1851. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, J.; Larsen, J.T.; Edlund, M.B. Peptostreptococcus micros has a uniquely high capacity to form hydrogen sulfide from glutathione. Oral Microbiol. Immunol. 1993, 8, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Dong, Z.; Xu, X.; Cochran, D.L.; Ebersole, J.L. Role of glutathione metabolism of Treponema denticola in bacterial growth and virulence expression. Infect. Immun. 2002, 70, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Ebersole, J.L.; Kurzban, G.P.; Holt, S.C. Cystalysin, a 46-kilodalton cysteine desulfhydrase from Treponema denticola, with hemolytic and hemoxidative activities. Infect. Immun. 1997, 65, 3231–3238. [Google Scholar] [PubMed]
- Chu, L.; Ebersole, J.L.; Kurzban, G.P.; Holt, S.C. Cystalysin, a 46-kda l-cysteine desulfhydrase from Treponema denticola: Biochemical and biophysical characterization. Clin. Infect. Dis. 1999, 28, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Ebersole, J.L.; Holt, S.C. Hemoxidation and binding of the 46-kda cystalysin of Treponema denticola leads to a cysteine-dependent hemolysis of human erythrocytes. Oral Microbiol. Immunol. 1999, 14, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Basic, A.; Dahlen, G. Hydrogen sulfide production from subgingival plaque samples. Anaerobe 2015, 35, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hagewald, S.; Bernimoulin, J.P.; Kottgen, E.; Kage, A. Total IgA and Porphyromonas gingivalis-reactive IgA in the saliva of patients with generalised early-onset periodontitis. Eur. J. Oral Sci. 2000, 108, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, B.W.; Darveau, R.P. Porphyromonas gingivalis lipopolysaccharide: An unusual pattern recognition receptor ligand for the innate host defense system. Acta Odontol. Scand. 2001, 59, 131–138. [Google Scholar] [CrossRef] [PubMed]
- McBride, B.C.; Joe, A.; Singh, U. Cloning of bacteroides gingivalis surface antigens involved in adherence. Arch. Oral Biol. 1990, 35 (Suppl. S59–S68). [Google Scholar] [CrossRef]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.P.; Ouyang, X.Y.; Wang, Y.X. Hydrogen sulfide synergistically upregulates Porphyromonas gingivalis lipopolysaccharide-induced expression of IL-6 and IL-8 via NF-kB signalling in periodontal fibroblasts. Arch. Oral Biol. 2014, 59, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Li, L.; Rose, P.; Tan, C.H.; Parkinson, D.B.; Moore, P.K. The effect of hydrogen sulfide donors on lipopolysaccharide-induced formation of inflammatory mediators in macrophages. Antioxid. Redox Signal. 2010, 12, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.; Schantz, J.T.; Haigh, R.; Wood, M.E.; Moore, P.K.; Viner, N.; Spencer, J.P.; Winyard, P.G.; Whiteman, M. Inducible hydrogen sulfide synthesis in chondrocytes and mesenchymal progenitor cells: Is H2S a novel cytoprotective mediator in the inflamed joint? J. Cell Mol. Med. 2012, 16, 896–910. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, K.; Oppenheim, J.J. Interleukin 8 and MCAF: Novel inflammatory cytokines inducible by IL1 and TNF. Cytokine 1989, 1, 2–13. [Google Scholar] [CrossRef]
- Jacquot, J.; Tabary, O.; Le Rouzic, P.; Clement, A. Airway epithelial cell inflammatory signalling in cystic fibrosis. Int. J. Biochem. Cell Biol. 2008, 40, 1703–1715. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Wong, F.L.; Fu, D.; Lau, H.Y.; Moochhala, S.M.; Moore, P.K. Role of hydrogen sulfide in acute pancreatitis and associated lung injury. FASEB J. 2005, 19, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bhatia, M.; Zhu, Y.Z.; Zhu, Y.C.; Ramnath, R.D.; Wang, Z.J.; Anuar, F.B.; Whiteman, M.; Salto-Tellez, M.; Moore, P.K. Hydrogen sulfide is a novel mediator of lipopolysaccharide-induced inflammation in the mouse. FASEB J. 2005, 19, 1196–1198. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greabu, M.; Totan, A.; Miricescu, D.; Radulescu, R.; Virlan, J.; Calenic, B. Hydrogen Sulfide, Oxidative Stress and Periodontal Diseases: A Concise Review. Antioxidants 2016, 5, 3. https://doi.org/10.3390/antiox5010003
Greabu M, Totan A, Miricescu D, Radulescu R, Virlan J, Calenic B. Hydrogen Sulfide, Oxidative Stress and Periodontal Diseases: A Concise Review. Antioxidants. 2016; 5(1):3. https://doi.org/10.3390/antiox5010003
Chicago/Turabian StyleGreabu, Maria, Alexandra Totan, Daniela Miricescu, Radu Radulescu, Justina Virlan, and Bogdan Calenic. 2016. "Hydrogen Sulfide, Oxidative Stress and Periodontal Diseases: A Concise Review" Antioxidants 5, no. 1: 3. https://doi.org/10.3390/antiox5010003
APA StyleGreabu, M., Totan, A., Miricescu, D., Radulescu, R., Virlan, J., & Calenic, B. (2016). Hydrogen Sulfide, Oxidative Stress and Periodontal Diseases: A Concise Review. Antioxidants, 5(1), 3. https://doi.org/10.3390/antiox5010003