Tempol Supplementation Restores Diaphragm Force and Metabolic Enzyme Activities in mdx Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Animals
2.3. Muscle Physiology
2.3.1. Ex Vivo Muscle Preparation
2.3.2. Isometric Protocol
2.3.3. Isotonic Protocol
2.4. Muscle Biochemistry
2.4.1. Tissue Preparation
2.4.2. Metabolic Enzyme Assays
2.5. Data Analysis
2.6. Statistical Analysis
3. Results
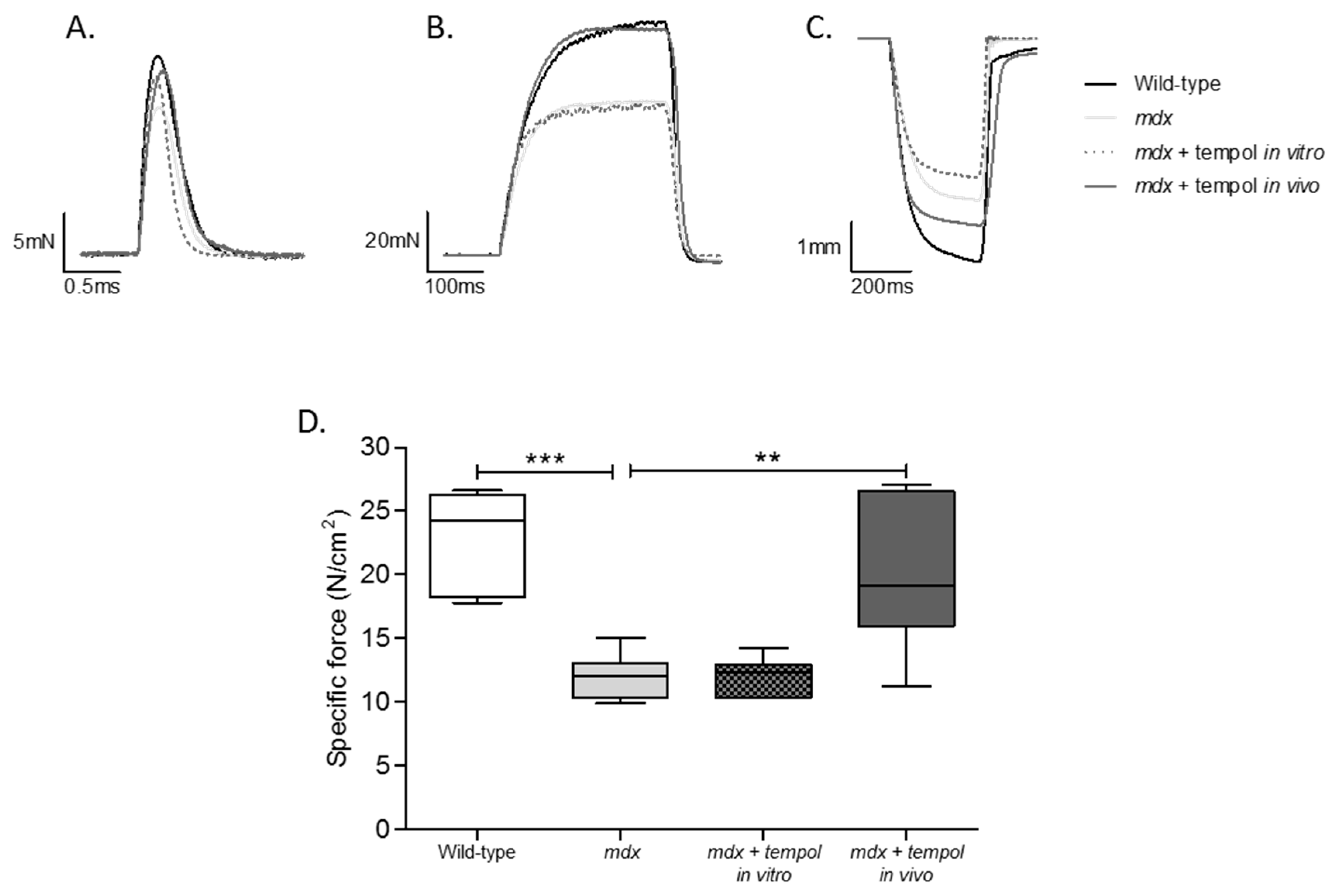
3.1. Isometric Force and Twitch Contractile Kinetics
3.2. Isotonic Contractile Parameters and Kinetics
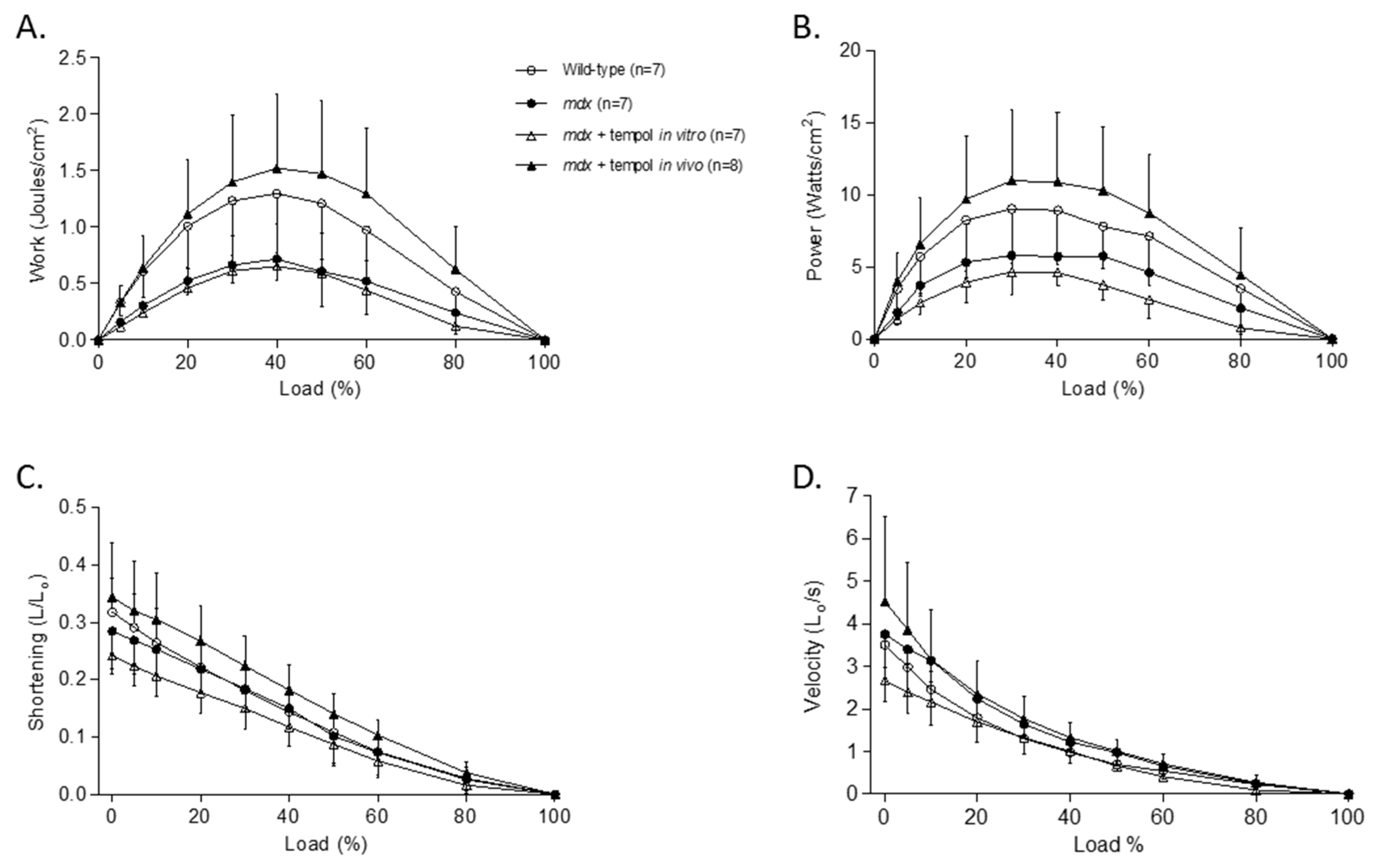
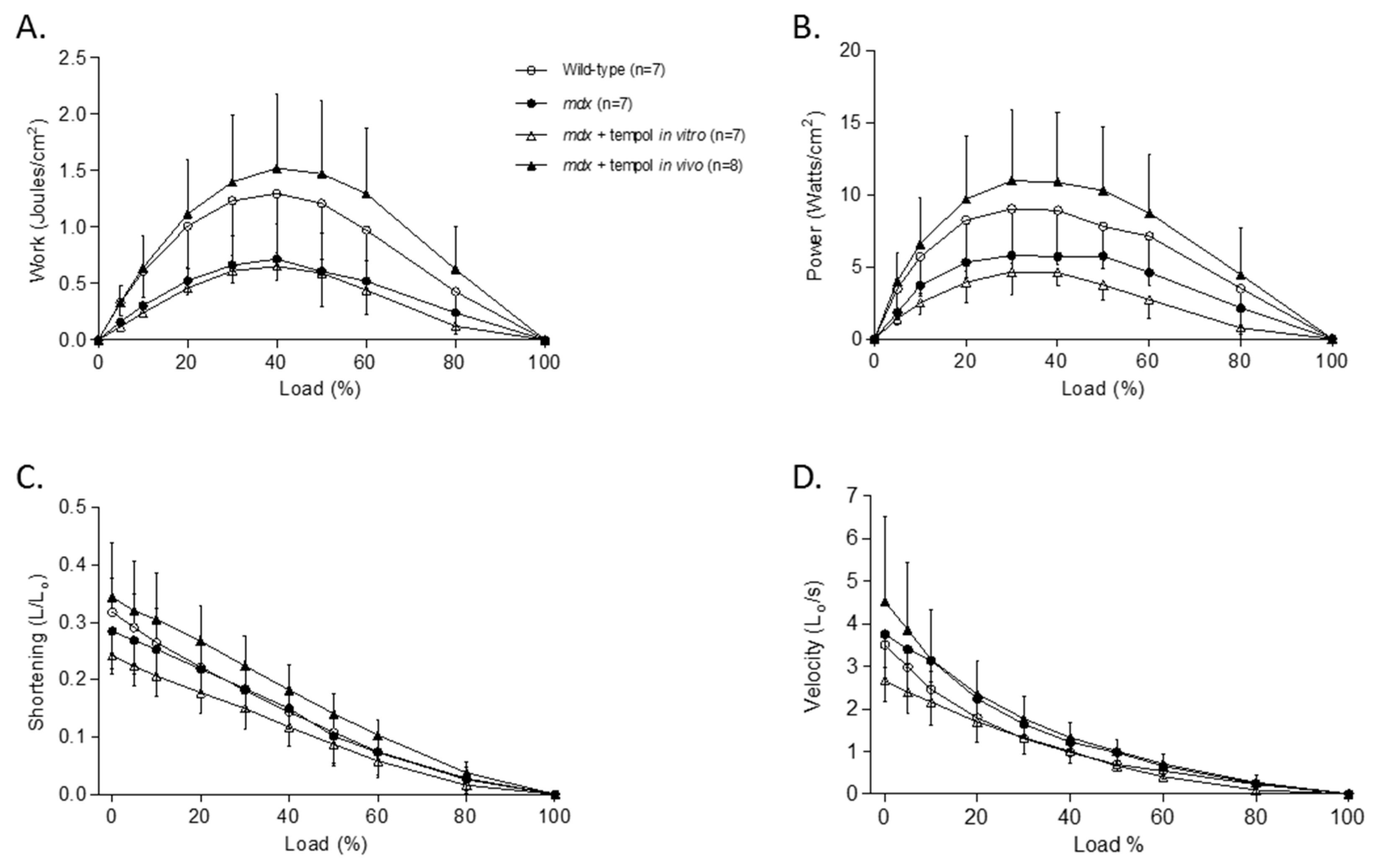
3.3. Isotonic Load Relationships
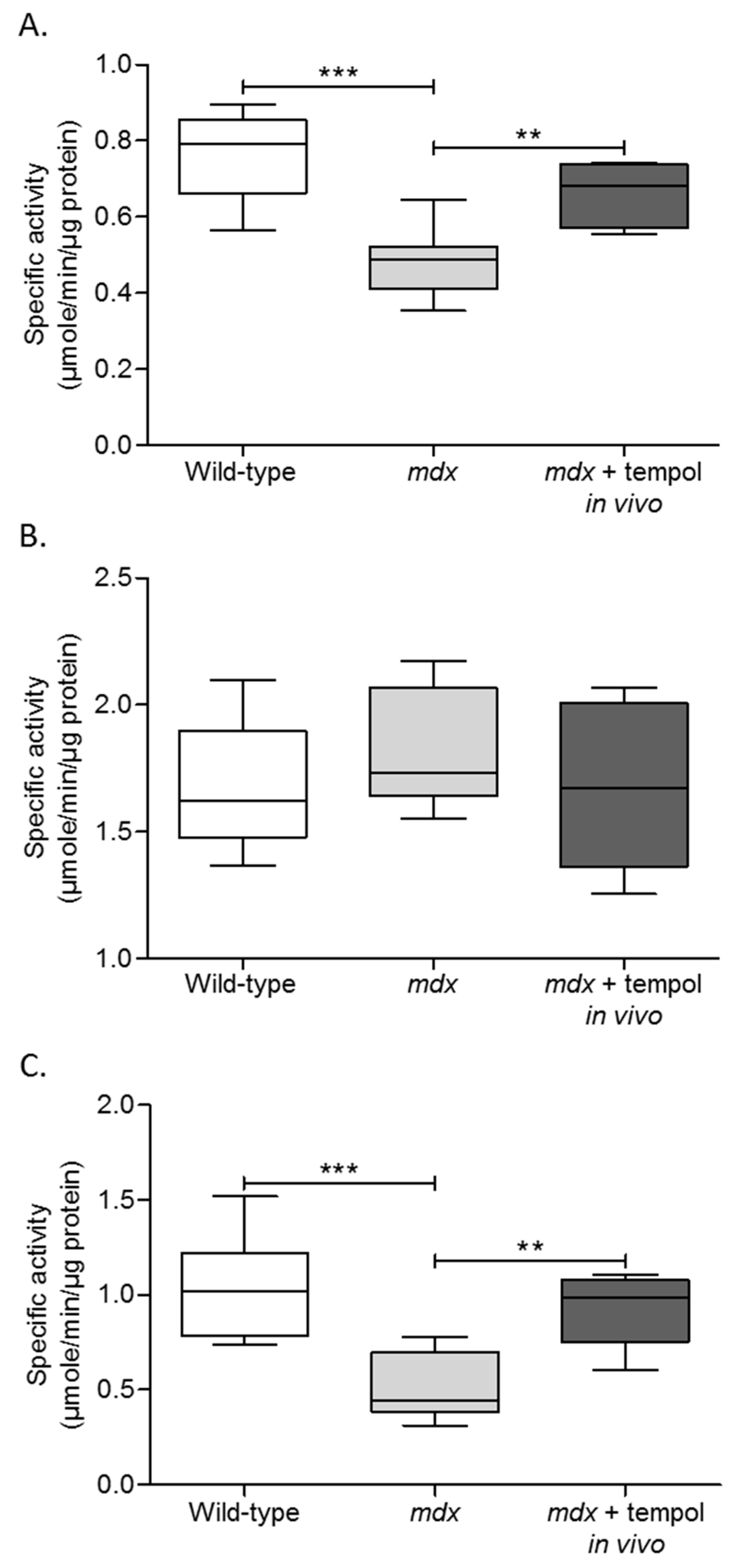
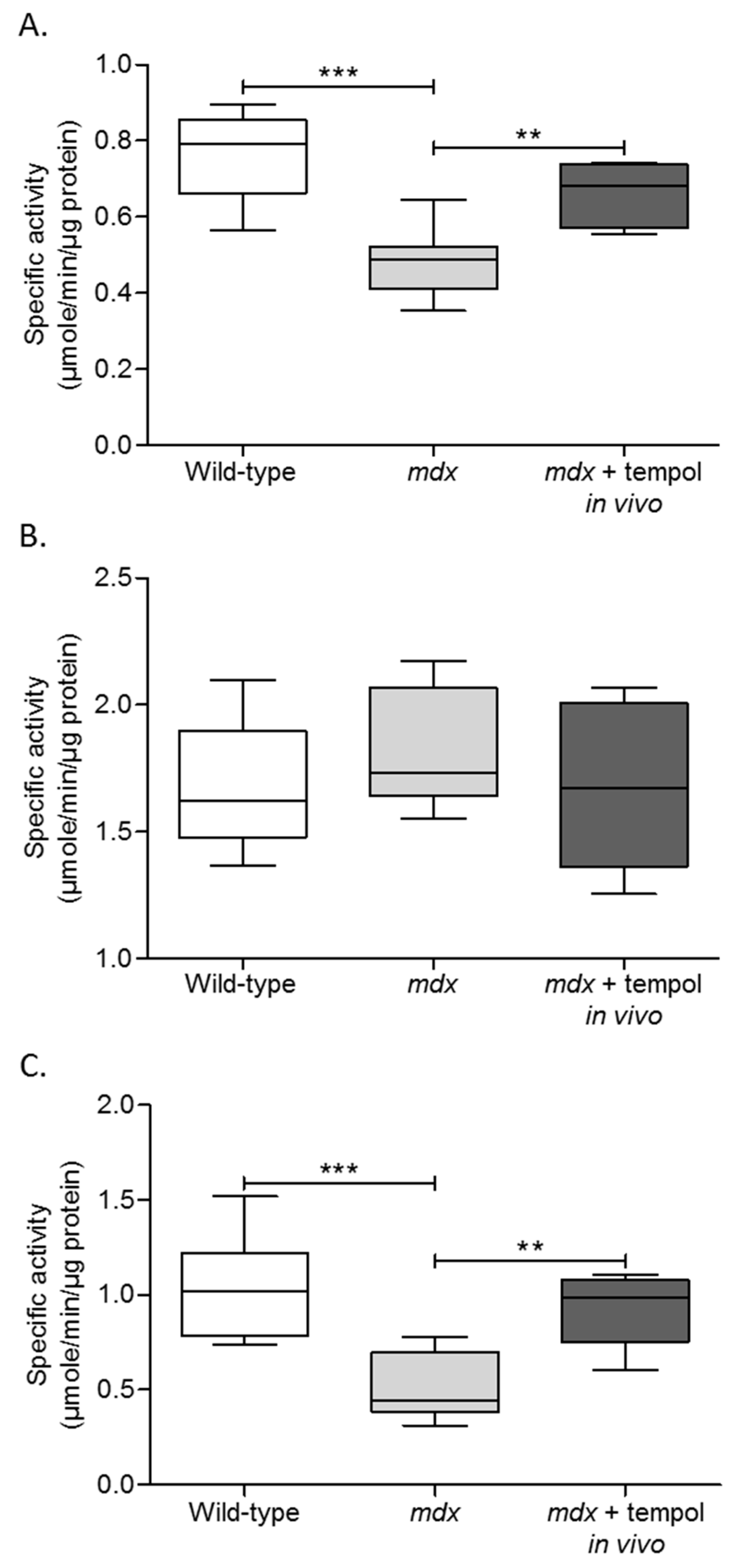
3.4. Metabolic Enzyme Activity
4. Discussion
Limitations
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Emery, A.E. Population frequencies of inherited neuromuscular diseases—A world survey. Neuromuscul. Disord. 1991, 1, 19–29. [Google Scholar] [CrossRef]
- Blake, D.J.; Weir, A.; Newey, S.E.; Davies, K.E. Function and genetics of dystrophin and dystrophin-related proteins in muscle. Physiol. Rev. 2002, 82, 291–329. [Google Scholar] [CrossRef] [PubMed]
- Ehmsen, J.; Poon, E.; Davies, K. The dystrophin-associated protein complex. J. Cell Sci. 2002, 115, 2801–2803. [Google Scholar] [PubMed]
- Kanagawa, M.; Toda, T. The genetic and molecular basis of muscular dystrophy: Roles of cell-matrix linkage in the pathogenesis. J. Hum. Genet. 2006, 51, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.P.; Yeung, E.W.; Allen, D.G. Muscle damage in mdx (dystrophic) mice: Role of calcium and reactive oxygen species. Clin. Exp. Pharmacol. Physiol. 2006, 33, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Yiu, E.M.; Kornberg, A.J. Duchenne muscular dystrophy. Neurol. India 2008, 56, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Bach, J.R.; Delaubier, A.; Renardel-Irani, A.; Guillou, C.; Rideau, Y. Clinical implications of maximal respiratory pressure determinations for individuals with duchenne muscular dystrophy. Arch. Phys. Med. Rehabil. 1997, 78, 1–6. [Google Scholar] [CrossRef]
- Khirani, S.; Ramirez, A.; Aubertin, G.; Boulé, M.; Chemouny, C.; Forin, V.; Fauroux, B. Respiratory muscle decline in duchenne muscular dystrophy. Pediatr. Pulmonol. 2014, 49, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.E.; Edwards, R.H.; Calverley, P.M. Ventilation and breathing pattern during sleep in duchenne muscular dystrophy. Chest 1989, 96, 1346–1351. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, E.; Sestili, P. Reactive oxygen species in skeletal muscle signaling. J. Signal Transduct. 2012, 2012, 982794. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.B. Invited review: Redox modulation of skeletal muscle contraction: What we know and what we don’t. J. Appl. Physiol. 2001, 90, 724–731. [Google Scholar] [PubMed]
- Allen, D.G.; Whitehead, N.P. Duchenne muscular dystrophy—What causes the increased membrane permeability in skeletal muscle? Int. J. Biochem. Cell Biol. 2011, 43, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.J. Control of reactive oxygen species production in contracting skeletal muscle. Antioxid. Redox Signal. 2011, 15, 2477–2486. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.G.; Gervasio, O.L.; Yeung, E.W.; Whitehead, N.P. Calcium and the damage pathways in muscular dystrophy. Can. J. Physiol. Pharmacol. 2010, 88, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Coirault, C.; Pignol, B.; Cooper, R.N.; Butler-Browne, G.; Chabrier, P.E.; Lecarpentier, Y. Severe muscle dysfunction precedes collagen tissue proliferation in mdx mouse diaphragm. J. Appl. Physiol. (1985) 2003, 94, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Stedman, H.H.; Sweeney, H.L.; Shrager, J.B.; Maguire, H.C.; Panettieri, R.A.; Petrof, B.; Narusawa, M.; Leferovich, J.M.; Sladky, J.T.; Kelly, A.M. The mdx mouse diaphragm reproduces the degenerative changes of duchenne muscular dystrophy. Nature 1991, 352, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Moraes, L.H.; de Burgos, R.R.; Macedo, A.B.; de Almeida Hermes, T.; de Faria, F.M.; Minatel, E. Reduction of oxidative damage and inflammatory response in the diaphragm muscle of mdx mice using iron chelator deferoxamine. Biol. Trace Elem. Res. 2015, 167, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.P.; Roy, A.; Lucking, E.F.; McDonald, F.B.; Gray, S.; Wilson, R.J.; Edge, D.; O’Halloran, K.D. Sensorimotor control of breathing in the mdx mouse model of duchenne muscular dystrophy. J. Physiol. 2017, 595, 6653–6672. [Google Scholar] [CrossRef] [PubMed]
- Williams, I.A.; Allen, D.G. The role of reactive oxygen species in the hearts of dystrophin-deficient mdx mice. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1969–H1977. [Google Scholar] [CrossRef] [PubMed]
- Kaczor, J.J.; Hall, J.E.; Payne, E.; Tarnopolsky, M.A. Low intensity training decreases markers of oxidative stress in skeletal muscle of mdx mice. Free Radic. Biol. Med. 2007, 43, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.P.; Yeung, E.W.; Froehner, S.C.; Allen, D.G. Skeletal muscle NADPH oxidase is increased and triggers stretch-induced damage in the mdx mouse. PLoS ONE 2010, 5, e15354. [Google Scholar] [CrossRef] [PubMed]
- Gordon, B.S.; Delgado-Diaz, D.C.; Carson, J.; Fayad, R.; Wilson, L.B.; Kostek, M.C. Resveratrol improves muscle function but not oxidative capacity in young mdx mice. Can. J. Physiol. Pharmacol. 2014, 92, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Comim, C.M.; Cassol, O.J., Jr.; Constantino, L.C.; Constantino, L.S.; Petronilho, F.; Tuon, L.; Vainzof, M.; Dal-Pizzol, F.; Quevedo, J. Oxidative variables and antioxidant enzymes activities in the mdx mouse brain. Neurochem. Int. 2009, 55, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.R.; Treuer, A.V.; Lamirault, G.; Mayo, V.; Cao, Y.; Dulce, R.A.; Hare, J.M. NADPH oxidase-2 inhibition restores contractility and intracellular calcium handling and reduces arrhythmogenicity in dystrophic cardiomyopathy. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H710–H721. [Google Scholar] [CrossRef] [PubMed]
- Ismail, H.M.; Scapozza, L.; Ruegg, U.T.; Dorchies, O.M. Diapocynin, a dimer of the NADPH oxidase inhibitor apocynin, reduces ROS production and prevents force loss in eccentrically contracting dystrophic muscle. PLoS ONE 2014, 9, e110708. [Google Scholar] [CrossRef] [PubMed]
- Lawler, J.M. Exacerbation of pathology by oxidative stress in respiratory and locomotor muscles with duchenne muscular dystrophy. J. Physiol. 2011, 589, 2161–2170. [Google Scholar] [CrossRef] [PubMed]
- Capogrosso, R.F.; Cozzoli, A.; Mantuano, P.; Camerino, G.M.; Massari, A.M.; Sblendorio, V.T.; De Bellis, M.; Tamma, R.; Giustino, A.; Nico, B.; et al. Assessment of resveratrol, apocynin and taurine on mechanical-metabolic uncoupling and oxidative stress in a mouse model of duchenne muscular dystrophy: A comparison with the gold standard, α-methyl prednisolone. Pharmacol. Res. 2016, 106, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Skelly, J.R.; Bradford, A.; Jones, J.F.; O’Halloran, K.D. Superoxide scavengers improve rat pharyngeal dilator muscle performance. Am. J. Respir. Cell Mol. Biol. 2010, 42, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Skelly, J.R.; Edge, D.; Shortt, C.M.; Jones, J.F.; Bradford, A.; O’Halloran, K.D. Tempol ameliorates pharyngeal dilator muscle dysfunction in a rodent model of chronic intermittent hypoxia. Am. J. Respir. Cell Mol. Biol. 2012, 46, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Shortt, C.M.; Fredsted, A.; Chow, H.B.; Williams, R.; Skelly, J.R.; Edge, D.; Bradford, A.; O’Halloran, K.D. Reactive oxygen species mediated diaphragm fatigue in a rat model of chronic intermittent hypoxia. Exp. Physiol. 2014, 99, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.; Sheehan, D.; Soares, R.; Varela Coelho, A.; O’Halloran, K.D. Chronic sustained hypoxia-induced redox remodeling causes contractile dysfunction in mouse sternohyoid muscle. Front. Physiol. 2015, 6, 122. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.; O’Halloran, K.D. Diaphragm muscle adaptation to sustained hypoxia: Lessons from animal models with relevance to high altitude and chronic respiratory diseases. Front. Physiol. 2016, 7, 623. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.N.; Macdonald, W.A.; van der Poel, C.; Stephenson, D.G. O2•− production at 37 °C plays a critical role in depressing tetanic force of isolated rat and mouse skeletal muscle. Am. J. Physiol. Cell Physiol. 2007, 293, C650–C660. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.P.; O’Halloran, K.D. Evidence of hypoxic tolerance in weak upper airway muscle from young mdx mice. Respir. Physiol. Neurobiol. 2016, 226, 68–75. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, K.D. Effects of nicotine on rat sternohyoid muscle contractile properties. Respir. Physiol. Neurobiol. 2006, 150, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.P.; Rowland, J.; Canavan, L.; Murphy, K.H.; Brannock, M.; O’Malley, D.; O’Halloran, K.D.; Edge, D. Restoration of pharyngeal dilator muscle force in dystrophin-deficient (mdx) mice following co-treatment with neutralizing interleukin-6 receptor antibodies and urocortin 2. Exp. Physiol. 2017, 102, 1177–1193. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.; Sheehan, D.; Soares, R.; Coelho, A.V.; O’Halloran, K.D. Redox remodeling is pivotal in murine diaphragm muscle adaptation to chronic sustained hypoxia. Am. J. Respir. Cell Mol. Biol. 2016, 55, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Weinberg, J.; Hamnegård, C.H.; Spahija, J.; Olofson, J.; Grimby, G.; Sinderby, C. Diaphragmatic function in advanced duchenne muscular dystrophy. Neuromuscul. Disord. 2006, 16, 161–167. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, P.F.; Ueki, J.; Bush, A.; Khan, Y.; Watson, A.; Pride, N.B. Diaphragm thickness and inspiratory strength in patients with duchenne muscular dystrophy. Thorax 1997, 52, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Baydur, A.; Gilgoff, I.; Prentice, W.; Carlson, M.; Fischer, D.A. Decline in respiratory function and experience with long-term assisted ventilation in advanced Duchenne’s muscular dystrophy. Chest 1990, 97, 884–889. [Google Scholar] [PubMed]
- Manning, J.; Buckley, M.M.; O’Halloran, K.D.; O’Malley, D. Combined XIL-6R and urocortin-2 treatment restores MDX diaphragm muscle force. Muscle Nerve 2017, 56, E134–E140. [Google Scholar] [CrossRef] [PubMed]
- Brand, K.A.; Hermfisse, U. Aerobic glycolysis by proliferating cells: A protective strategy against reactive oxygen species. FASEB J. 1997, 11, 388–395. [Google Scholar] [PubMed]
- Le, A.; Cooper, C.R.; Gouw, A.M.; Dinavahi, R.; Maitra, A.; Deck, L.M.; Royer, R.E.; Vander Jagt, D.L.; Semenza, G.L.; Dang, C.V. Inhibition of lactate dehydrogenase a induces oxidative stress and inhibits tumor progression. Proc. Natl. Acad. Sci. USA 2010, 107, 2037–2042. [Google Scholar] [CrossRef] [PubMed]
- Comim, C.M.; Hoepers, A.; Ventura, L.; Freiberger, V.; Dominguini, D.; Mina, F.; Mendonça, B.P.; Scaini, G.; Vainzof, M.; Streck, E.L.; et al. Activity of krebs cycle enzymes in mdx mice. Muscle Nerve 2016, 53, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.P.; Kim, M.J.; Bible, K.L.; Adams, M.E.; Froehner, S.C. A new therapeutic effect of simvastatin revealed by functional improvement in muscular dystrophy. Proc. Natl. Acad. Sci. USA 2015, 112, 12864–12869. [Google Scholar] [CrossRef] [PubMed]
- Hukins, C.A.; Hillman, D.R. Daytime predictors of sleep hypoventilation in duchenne muscular dystrophy. Am. J. Respir. Crit. Care Med. 2000, 161, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Burns, D.; Edge, D.; O’Malley, D.; O’Halloran, K. Respiratory control in the mdx mouse model of duchenne muscular dystrophy. Adv. Exp. Med. Biol. 2015, 860, 239–244. [Google Scholar] [PubMed]
- Mosqueira, M.; Baby, S.M.; Lahiri, S.; Khurana, T.S. Ventilatory chemosensory drive is blunted in the mdx mouse model of Duchenne Muscular Dystrophy (DMD). PLoS ONE 2013, 8, e69567. [Google Scholar] [CrossRef] [PubMed]
- Barbé, F.; Quera-Salva, M.A.; McCann, C.; Gajdos, P.; Raphael, J.C.; de Lattre, J.; Agustí, A.G. Sleep-related respiratory disturbances in patients with Duchenne muscular dystrophy. Eur. Respir. J. 1994, 7, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.E.; Edwards, R.H.; Calverley, P.M. Oxygen treatment of sleep hypoxaemia in Duchenne muscular dystrophy. Thorax 1989, 44, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Farkas, G.A.; McCormick, K.M.; Gosselin, L.E. Episodic hypoxia exacerbates respiratory muscle dysfunction in DMD(mdx) mice. Muscle Nerve 2007, 36, 708–710. [Google Scholar] [CrossRef] [PubMed]
- Shortt, C.M.; O’Halloran, K.D. Hydrogen peroxide alters sternohyoid muscle function. Oral. Dis. 2014, 20, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lawler, J.M. Amplification of proinflammatory phenotype, damage, and weakness by oxidative stress in the diaphragm muscle of mdx mice. Free Radic. Biol. Med. 2012, 52, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Tonon, E.; Ferretti, R.; Shiratori, J.H.; Santo Neto, H.; Marques, M.J.; Minatel, E. Ascorbic acid protects the diaphragm muscle against myonecrosis in mdx mice. Nutrition 2012, 28, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Hnia, K.; Hugon, G.; Rivier, F.; Masmoudi, A.; Mercier, J.; Mornet, D. Modulation of p38 mitogen-activated protein kinase cascade and metalloproteinase activity in diaphragm muscle in response to free radical scavenger administration in dystrophin-deficient mdx mice. Am. J. Pathol. 2007, 170, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Hermes, T.e.A.; Macedo, A.B.; Fogaça, A.R.; Moraes, L.H.; de Faria, F.M.; Kido, L.A.; Cagnon, V.H.; Minatel, E. Beneficial cilostazol therapeutic effects in mdx dystrophic skeletal muscle. Clin. Exp. Pharmacol. Physiol. 2016, 43, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Mâncio, R.D.; Hermes, T.A.; Macedo, A.B.; Mizobuti, D.S.; Rupcic, I.F.; Minatel, E. Dystrophic phenotype improvement in the diaphragm muscle of mdx mice by diacerhein. PLoS ONE 2017, 12, e0182449. [Google Scholar] [CrossRef] [PubMed]
- Nakae, Y.; Hirasaka, K.; Goto, J.; Nikawa, T.; Shono, M.; Yoshida, M.; Stoward, P.J. Subcutaneous injection, from birth, of epigallocatechin-3-gallate, a component of green tea, limits the onset of muscular dystrophy in mdx mice: A quantitative histological, immunohistochemical and electrophysiological study. Histochem. Cell Biol. 2008, 129, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Buyse, G.M.; Van der Mieren, G.; Erb, M.; D’hooge, J.; Herijgers, P.; Verbeken, E.; Jara, A.; Van Den Bergh, A.; Mertens, L.; Courdier-Fruh, I.; et al. Long-term blinded placebo-controlled study of SNT-MC17/idebenone in the dystrophin deficient mdx mouse: Cardiac protection and improved exercise performance. Eur. Heart J. 2009, 30, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.J.; Barbin, I.C.; Taniguti, A.P.; Oggian, D.S.; Ferretti, R.; Santo Neto, H. Myocardial fibrosis is unaltered by long-term administration of L-arginine in dystrophin deficient mdx mice: A histomorphometric analysis. Acta Biol. Hung. 2010, 61, 168–174. [Google Scholar] [CrossRef] [PubMed]
- De Senzi Moraes Pinto, R.; Ferretti, R.; Moraes, L.H.; Neto, H.S.; Marques, M.J.; Minatel, E. N-acetylcysteine treatment reduces TNF-α levels and myonecrosis in diaphragm muscle of mdx mice. Clin. Nutr. 2013, 32, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Kuno, A.; Hori, Y.S.; Hosoda, R.; Tanno, M.; Miura, T.; Shimamoto, K.; Horio, Y. Resveratrol improves cardiomyopathy in dystrophin-deficient mice through Sirt1 protein-mediated modulation of p300 protein. J. Biol. Chem. 2013, 288, 5963–5972. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, L.E.; Williams, J.E. Pentoxifylline fails to attenuate fibrosis in dystrophic (mdx) diaphragm muscle. Muscle Nerve 2006, 33, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Burdi, R.; Rolland, J.F.; Fraysse, B.; Litvinova, K.; Cozzoli, A.; Giannuzzi, V.; Liantonio, A.; Camerino, G.M.; Sblendorio, V.; Capogrosso, R.F.; et al. Multiple pathological events in exercised dystrophic mdx mice are targeted by pentoxifylline: Outcome of a large array of in vivo and ex vivo tests. J. Appl. Physiol. 2009, 106, 1311–1324. [Google Scholar] [CrossRef] [PubMed]
- Graham, K.M.; Singh, R.; Millman, G.; Malnassy, G.; Gatti, F.; Bruemmer, K.; Stefanski, C.; Curtis, H.; Sesti, J.; Carlson, C.G. Excessive collagen accumulation in dystrophic (mdx) respiratory musculature is independent of enhanced activation of the NF-kappaB pathway. J. Neurol. Sci. 2010, 294, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hollinger, K.; Shanely, R.A.; Quindry, J.C.; Selsby, J.T. Long-term quercetin dietary enrichment decreases muscle injury in mdx mice. Clin. Nutr. 2015, 34, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Selsby, J.T.; Ballmann, C.G.; Spaulding, H.R.; Ross, J.W.; Quindry, J.C. Oral quercetin administration transiently protects respiratory function in dystrophin-deficient mice. J. Physiol. 2016, 594, 6037–6053. [Google Scholar] [CrossRef] [PubMed]
- Ballmann, C.; Denney, T.; Beyers, R.J.; Quindry, T.; Romero, M.; Selsby, J.T.; Quindry, J.C. Long-term dietary quercetin enrichment as a cardioprotective countermeasure in mdx mice. Exp. Physiol. 2017, 102, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Ballmann, C.; Hollinger, K.; Selsby, J.T.; Amin, R.; Quindry, J.C. Histological and biochemical outcomes of cardiac pathology in mdx mice with dietary quercetin enrichment. Exp. Physiol. 2015, 100, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Percival, J.M.; Whitehead, N.P.; Adams, M.E.; Adamo, C.M.; Beavo, J.A.; Froehner, S.C. Sildenafil reduces respiratory muscle weakness and fibrosis in the mdx mouse model of Duchenne muscular dystrophy. J. Pathol. 2012, 228, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Mâncio, R.D.; Hermes, T.A.; Macedo, A.B.; Mizobuti, D.S.; Valduga, A.H.; Rupcic, I.F.; Minatel, E. Vitamin E treatment decreases muscle injury in mdx mice. Nutrition 2017, 43–44, 39–46. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Wild-Type (n = 7) | Mdx (n = 7) | mdx + Tempol In Vitro (n = 7) | mdx + Tempol In Vivo (n = 8) | Student’s t Test | |
|---|---|---|---|---|---|
| CT (ms) | 18.0 ± 1.8 | 20.5 ± 4.5 | 17.9 ± 1.5 | 20.2 ± 3.1 | $: p = 0.2196; †: p = 0.1796; £: p = 0.8777 |
| ½ RT (ms) | 23.5 ± 0.6 | 23.5 ± 0.5 | 23.2 ± 0.2 | 17.8 ± 3.9 | $: p = 0.9803; †: p = 0.1197; £: p = 0.0046 |
| Pt (N/cm2) | 5.1 ± 1.7 | 2.5 ± 0.7 | 2.7 ± 0.7 | 4.0 ± 1.2 | $: p = 0.0066; †: p = 0.4821; £: p = 0.0111 |
| Wmax (J/cm2) | 1.3 ± 0.5 | 0.7 ± 0.2 | 0.7 ± 0.4 | 1.5 ± 0.7 | $: p = 0.0276; †: p = 0.6852; £: p = 0.0085 |
| Pmax (W/cm2) | 9.0 ± 3.8 | 5.8 ± 0.9 | 4.7 ± 1.6 | 11.0 ± 4.9 | $: p = 0.0709; †: p = 0.1329; £: p = 0.0217 |
| Smax (L/Lo) | 0.32 ± 0.06 | 0.28 ± 0.07 | 0.24 ± 0.03 | 0.34 ± 0.10 | $: p = 0.3457; †: p = 0.1532; £: p = 0.1936 |
| Vmax (Lo/s) | 3.5 ± 1.3 | 3.8 ± 0.8 | 2.7 ± 0.9 | 4.5 ± 2.0 | $: p = 0.6766; †: p = 0.0353; £: p = 0.3433 |
| Antioxidant | Author | Classification | Model | Age | Dose/Method of Delivery | Tissue Examined | Results |
|---|---|---|---|---|---|---|---|
| α-lipoic acid/L-carnitine | Hnia K. et al., 2007 [55] | Free radical scavenger | mdx mouse | 5 weeks old | 250 mg/kg α-lipoic acid/L-carnitine i.p injection for 14 days | Diaphragm | α-lipoic acid/L-carnitine decreased plasma CK levels and decreased muscle fibre central nucleation and fibre variance, antioxidant activity, lipid peroxidation, NF-kB and matrix metalloproteinase activity in mdx diaphragm. Β-dystroglycan expression was increased in mdx diaphragm following α-lipoic acid/L-carnitine. |
| Apocynin | Gonzalez D.R. et al., 2014 [24] | NADPH oxidase inhibitor | mdx cardiac myocytes | - | 100 µM apocynin in vitro | Isolated cardiac myocytes | Apocynin restored contractility in mdx cardiac myocytes and normalised the amplitude of evoked intracellular Ca2+ concentration transients and total SR Ca2+ content. The production of spontaneous diastolic Ca2+ release events was decreased and SR Ca2+ leakage was decreased, thus apocynin improved SR Ca2+ handling and contractility in mdx cardiac myocytes. |
| Ascorbic acid (vitamin C) | Tonon E. et al., 2012 [54] | Antioxidant | mdx mouse | 14 days old | Ascorbic acid 200 mg/kg via oral gavage daily for 14 days | Diaphragm | Ascorbic acid decreased plasma CK levels and diaphragm myonecrosis, inflammation, TNF-α and 4-HNE levels and Evans blue dye staining in mdx mice. |
| Cilostazol | Hermes Tde A.E. et al., 2016 [56] | PDE3 inhibitor | mdx mouse | 14 days old | Cilostazol 100 mg/kg/day for 14 days | Diaphragm | Cilostazol reduced plasma CK and diaphragm myonecrosis, inflammatory cell area and macrophage infiltration, NF-kB and TNF-α content, ROS production and oxidative stress in mdx mice. |
| Diacerhein | Mâncio R.D. et al., 2017 [57] | IL-1β inhibitor | mdx mouse | 14 days old | 20 mg/kg/day diacerhein via oral gavage for 14 days | Diaphragm | Diacerhin reduced plasma CK levels, diaphragm muscle fibre damage and central nucleation, inflammatory mediators, oxidative stress and lipid peroxidation in mdx mice. |
| EUK-134 | Kim J.H. and Lawler J.M. 2012 [53] | Superoxide dismutase mimetic | mdx mouse | 20 days old | 30 mg/kg/day EUK-134 i.p. injection for 8 days | Diaphragm | EUK-134 reduced 4-HNE, total hydroperoxides, positive staining of macrophages and T-cells, activation of NF-κB, p65 protein abundance and the number of centralised myonuclei and variability of fibre size in diaphragm muscle from mdx mice. Diaphragm contractile force was partially rescued following EUK-134 and increased citrate synthase activity in mdx mice. |
| Epigallocatechin-3-gallate (EGCG) | Nakae Y. et al., 2008 [58] | Green tea extract/antioxidant/Polyphenol | mdx mouse | From birth | 5 mg/kg EGCG s.c. injection 4 times per week for 8 weeks | Diaphragm | EGCG had no effect on body weight and no observable toxic effects in the liver and kidney. EGCG decreased plasma CK and decreased the number of lipofuscin granules, necrotic muscle fibres and connective tissue in mdx diaphragm and increased utrophin expression. EGCG did not affect diaphragm isometric force. |
| SNT-NC17/Idebenone | Buyse G.M. et al., 2009 [59] | Antioxidant | mdx mouse | 4 weeks old | 200 mg/kg SNT-MC17/idebenone for 9 months | Heart | SNT-NC17/Idebenone corrected cardiac diastolic dysfunction, improved contractile reserve and voluntary running and decreased cardiac inflammation and fibrosis in mdx mice. |
| L-arginine | Marques M.J. et al., 2010 [60] | Amino acid | mdx mouse | 6 months old | L-arginine in drinking water for 6 months | Heart | L-arginine had no effect on myocardial fibrosis but reduced the density of inflammatory cells in the mdx heart. |
| N-acetylcysteine (NAC) | Williams I.A. and Allen D.G. 2007 [19] | Glutathione precursor | mdx mouse | 3 weeks old | 1% NAC in drinking water for 6 weeks | Heart | NAC reduced DHE levels in mdx hearts, reduced abnormalities in mdx cardiomyocyte Ca2+ handling, returned mdx fractional shortening to WT values but did not affect Ca2+ sensitivity. NAC returned collagen type III and CD68 expression in mdx hearts to WT values. |
| N-acetylcysteine (NAC) | de Senzi Moraes Pinto R. et al., 2013 [61] | Glutathione precursor | mdx mouse | 14 days old | 150 mg/kg NAC i.p. daily for 14 days | Diaphragm | NAC reduced plasma CK levels and reduced TNF-α and 4-HNE protein adduct levels, inflammation, Evans blue dye staining and myonecrosis in mdx diaphragm muscle. |
| Resveratrol | Kuno A. et al., 2013 [62] | SIRT1 activator | mdx mouse | 9 weeks old | 4 g/kg resveratrol enriched diet for 32 weeks | Heart | Resveratrol downregulated the pro-hypertrophic co-activator p300 protein level in the mdx heart thus inhibiting fibre hypertrophy. Resveratrol also suppressed cardiac fibrosis and preserved cardiac diastolic function in mdx hearts. |
| Pentoxifylline | Gosselin L.E. and Williams J.E. 2006 [63] | PDE inhibitor | mdx mouse | 4 weeks old | 16 mg/kg/day pentoxyifylline for 4 weeks | Diaphragm | Pentoxyifylline had no effect on mdx diaphragm force, hydroxyproline concentration, type I and III procollagen mRNA and TGF-β mRNA. |
| Pentoxifylline | Burdi R. et al., 2009 [64] | PDE inhibitor | mdx mouse | 4–5 weeks old | 50 mg/kg/day pentoxyifylline i.p. injection for 4–8 weeks | Diaphragm | Pentoxifylline modestly increased mdx diaphragm isometric tetanic force. |
| Pyrrolidine dithiocarbamate (PDTC) or ursodeoxycholic acid(UDCA) | Graham K.M. et al., 2010 [65] | NF-κB inhibitors | mdx mouse | 30 days old | 50 mg/kg/day PDTC i.p. injection for one month 40 mg/kg/day UDCA i.p. injection for one month | Diaphragm | Neither PDTC or UDCA influenced collagen deposition or TGF-β1 expression in mdx diaphragm. |
| Quercetin | Hollinger K. et al., 2015 [66] | PGC-1α pathway activator | mdx mouse | 3 months old | 0.2% quercetin-enriched diet for 6 months | Diaphragm | Quercetin preserved diaphragm muscle fibres and reduced centralised nuclei, infiltrating immune cells, TNF-α gene expression and muscle fibrosis in mdx mice. Genes associated with oxidative metabolism were increased following quercetin. |
| Quercetin | Selsby J.T. et al., 2016 [67] | PGC-1α pathway activator | mdx mouse | 2 months old | 0.2% quercetin-enriched diet for 12 months | Diaphragm | Quercetin protected respiratory function in mdx mice during the first 4–6 months and declined thereafter. Mdx diaphragm muscle function and histology were not preserved following 12 months of quercetin treatment. |
| Quercetin | Ballmann C. et al., 2017 [68] | PGC-1α pathway activator | mdx mouse | 2 months old | 0.2% quercetin-enriched diet for 12 months | Heart | Quercetin decreased fibronectin, inflammation and indices of tissue damage while mitochondrial biogenesis and antioxidant enzymes were improved, and quercetin facilitated the assembly of the DAPC in mdx hearts. |
| Quercetin | Ballmann C. et al., 2015 [69] | PGC-1α pathway activator | mdx mouse | 3 weeks old 3 months old | 0.2% quercetin-enriched diet for 6 months | Heart | 3 weeks old: Quercetin increased cytochrome-c and superoxide dismutase 2 protein expression, increased utrophin and decreased matrix metalloproteinase 9 abundance in mdx heart. 3 months old: Quercetin decreased relative and absolute heart weights, damage indicators and TGFβ-1 in mdx heart. |
| Sildenafil | Percival J.M. et al., 2012 [70] | PDE-5 inhibitor | mdx mouse | 3 weeks old | 400 mg/L sildenafil citrate in drinking water for 14 weeks | Diaphragm | Sildenafil modestly increased diaphragm force generating capacity and reduced fibronectin, TNF-α, matrix metalloproteinase 13 and Evans blue dye staining in the mdx diaphragm. Fatigue resistance and TGF-β were unaffected. |
| Vitamin E | Mancio R.D. et al., 2017 [71] | Peroxyl radical scavenger | mdx mouse | 14 day old | 40 mg vitamin E/kg daily via oral gavage for 14 days | Diaphragm | Vitamin E reduced muscle fibre damage, oxidative stress and inflammation processes in mdx diaphragm. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burns, D.P.; Ali, I.; Rieux, C.; Healy, J.; Jasionek, G.; O’Halloran, K.D. Tempol Supplementation Restores Diaphragm Force and Metabolic Enzyme Activities in mdx Mice. Antioxidants 2017, 6, 101. https://doi.org/10.3390/antiox6040101
Burns DP, Ali I, Rieux C, Healy J, Jasionek G, O’Halloran KD. Tempol Supplementation Restores Diaphragm Force and Metabolic Enzyme Activities in mdx Mice. Antioxidants. 2017; 6(4):101. https://doi.org/10.3390/antiox6040101
Chicago/Turabian StyleBurns, David P., Izza Ali, Clement Rieux, James Healy, Greg Jasionek, and Ken D. O’Halloran. 2017. "Tempol Supplementation Restores Diaphragm Force and Metabolic Enzyme Activities in mdx Mice" Antioxidants 6, no. 4: 101. https://doi.org/10.3390/antiox6040101
APA StyleBurns, D. P., Ali, I., Rieux, C., Healy, J., Jasionek, G., & O’Halloran, K. D. (2017). Tempol Supplementation Restores Diaphragm Force and Metabolic Enzyme Activities in mdx Mice. Antioxidants, 6(4), 101. https://doi.org/10.3390/antiox6040101