1. Introduction
Cowpea (
Vigna unguiculata (L.) Walp.) is an important legume crop produced and consumed in Sub-Saharan Africa by the majority of smallholder farmers, especially women. Cowpea is diploid, with 2n = 22 and a genome size of about 620 Mb [
1,
2,
3]. The crop is autogamous, but outcrossing of up to 5% has been reported in the cultivated varieties, mainly due to insect activities [
4]. It is a crucial food crop for meeting dietary protein needs and preventing micronutrient deficiencies [
5], especially among women and children [
6].
Cowpea whole grains contain 23–32% protein, 50–60% carbohydrates, and 1% fat [
7], as well as considerable amounts of phytochemicals, dietary fiber, minerals (calcium, iron, and zinc), and vitamins. Iron and zinc are essential for human well-being. Their adequate supply helps prevent anemia and boost the immune system, two common problems in developing countries [
8]. The composition of nutrients can vary due to several factors, including varietal differences, climatic conditions, and agronomic practices [
9]. Some studies have reported mature grains to contain lower concentrations of most minerals [
10] than immature green pods [
11] and leaves [
12].
Despite its enormous economic potential, the productivity of the crop is considerably low in Malawi, which is estimated at an average of less than 500 kg/ha among smallholder farmers [
13]. This low productivity is attributed to low adoption levels of improved varieties (currently at 10%) and continued use of landraces, which tend to be low-yielding and highly susceptible to biotic and abiotic stress. Improved varieties, however, have been blamed for lacking important quality traits such as grain size, cooking time, and broth thickness. Low adoption of improved varieties and a lack of farmer and consumer-preferred varieties have been attributed to the failure of national breeding programs to involve farmers in the process of designing and developing varieties to meet their priorities and preferences [
14].
Although cowpeas are nutritionally rich, the longer cooking time makes the legume unacceptable to some consumers. In a trait preference study conducted in Malawi in 2021 [
15], short cooking time and broth thickness were among the priority traits for consumers to adopt and utilize a particular cowpea variety. It has also been reported that broth thickness is an important trait in the determination of cooking quality [
16]. Traore et al. [
17] stated that cooking qualities must be combined with other consumer preferences to encourage cowpea utilization. Other traits that influence cowpea marketing and utilization include seed coat color, seed shape, and grain size [
18,
19,
20,
21,
22].
Considerable genetic variability has been reported for cowpea’s physical, cooking, and nutritional quality traits [
19,
20,
23,
24,
25,
26]. In Malawi, reports on the genetic variability for grain quality (grain color, seed size, seed shape), cooking quality (cooking time, broth thickness), and nutritional quality (protein, zinc, and iron) traits on a diverse panel of cowpea genotypes are rare and non-existent. The degree of association among these quality traits has neither been fully dissected nor exploited to inform cowpea breeding programs. The present study was therefore conducted to evaluate the genetic variability of cowpea genotypes for grain, cooking, and nutritional quality traits. Further, this study was conducted to understand the degree of associations among the cowpea quality traits so that they can be exploited in a breeding program to develop cowpea varieties with consumer-preferred, market-driven, and resilient inclusive traits to enhance the food, income, and nutrition security of smallholder farmers in East and Southern Africa.
4. Discussion
Most resource-constrained smallholder farmers in sub-Saharan Africa, especially women, rely on cowpeas as their major source of protein and other mineral elements such as iron and zinc, thereby contributing to their improved nutritional status and health well-being. In dryland areas, cowpea is a major source of income to support their daily livelihoods. At the same time, cowpea is one of the crops used to adapt to climate-related changes due to its resilience in these areas. Current cowpea breeding strategies in the region must, therefore, take into consideration its grain, nutritional value, and cooking quality, which influence farmers/consumers’ choices and consumption.
This study observed significant variations in the genotypes for grain quality, cooking quality, and nutritional quality traits (
Table 1,
Table 3 and
Table 5). This suggests that genetic variability exists in the current set of cowpea germplasm in Malawi, which could be exploited in breeding programs aimed at developing high-performing varieties for the said traits. Assessment of genetic variability for different traits is an important pre-breeding step as it provides an opportunity for plant breeders to develop new and improved varieties with desirable characteristics that are both farmer- and consumer-preferred. Earlier studies [
6,
40,
41] reported wide genetic variation in grain quality, crude protein, and micronutrient contents in cowpeas, suggesting that the nutritional value of varieties could be improved.
In the study, seed coat color among the genotypes was quite variable, predominated by brown color (43.14%), followed by white (16.39%), and reddish-brown (13.40%). In cowpeas, seed coat color is an important aspect of consumer preference, which directly influences the marketability of the grain. This implies that variety development programs need to respond to specific and unique market preferences for improved uptake of the varieties. Trait preference studies in different regions and countries reported these unique preferences [
19,
21,
22,
42,
43]. For instance, in Malawi, Mozambique, and Tanzania, through a trait preference survey [
15] among farmers, consumers, traders, and processors, seed coat color was among the most preferred characteristics.
The genotypes in the study also varied for seed shape, with over 51.67% being ovoid and 32.03% rhomboid. This finding is similar to Kim et al. [
44], who reported that out of the 245 cowpea genotypes in the study, 66.9% were egg-shaped (ovoid) and 24.9% were rectangular (rhomboid). This suggests that the ovoid shape is a dominant seed shape in cowpeas.
The other important trait in cowpea breeding is grain size. Seed size is directly correlated with grain yield, and as such, it is considered an essential market trait [
18]. Seed size is measured as grams per 100 seeds, and Moses and Zibokere [
24] grouped seed size into small (10–15 g), medium (15.1–20 g), large (20.1–25), and very large (greater than 25 g). In the present study, genotypes exhibited wide genetic variability for seed size, which ranged from 6 to 26.13 g. This range is within most of the reported seed sizes [
23,
26]. It was noted that most of the landraces were larger-seeded than the improved varieties. As noted before, most farmers prefer large-seeded varieties, and this might explain the reasons behind the low adoption levels of improved varieties in Malawi since all the improved varieties have small grain sizes.
With cowpeas being an important source of protein and other elements for many smallholder farmers, breeding programs must continue to develop and release more nutritious varieties. The genetic variability for nutritional quality properties such as protein, zinc, and iron exhibited in the study is a sure way to set the trajectory towards sustainable, improved variety development that meets the nutritional needs of farmers, especially women and children. Genetic variability for nutritional quality in cowpeas is a common phenomenon [
6,
12,
20,
40,
41], which means that selection for these traits in cowpeas can increase genetic gain.
Cooking time is usually associated with energy sources. In sub-Saharan Africa, where the predominant energy source is fuel wood, most households struggle to prepare meals that require more energy. Shorter cooking varieties are usually a strategy for energy savings. The findings of the study established some considerable genetic variation among genotypes, with some taking a minimum of 39 min to cook. Some studies have reported a minimum of 35 min [
45] to cook. Contrastingly, some genotypes in the study took 191 min, which suggests that they are less economical in terms of energy consumption. Related to cooking quality is broth viscosity, which very few studies have looked at in cowpeas as a genetically controlled trait. This study suggests that the variability recorded is genetically controlled, as considerable variation was observed in the genotypes, which ranged from 1.6 to 5.5 cP. In Malawi, it has been reported that broth thickness is one of the key determinants of the cooking quality of the legume [
16], and if a variety lacks this trait, it can rarely be taken up by farmers/consumers.
Significant positive correlations were detected among the 306 genotypes for protein content with iron and zinc (
Table 7). This is in agreement with the positive correlation between crude protein and Fe contents in 11 genotypes reported by Moura et al. [
25] and Boukar et al. [
6,
40]. Since the presence of genetic variability has already been elucidated for these traits, it is, therefore, possible to improve protein, iron, and zinc contents without adverse interactions, as also espoused by Nielsen et al. [
46] and Jean Baptiste et al. [
47], who indicated some heritability for crude protein content in cowpea.
On the other hand, nutritional quality traits did not exhibit any association with grain quality or cooking quality traits. The non-significant associations between grain quality and nutritional quality traits suggest that desirable traits can be incorporated through trait introgression techniques. This is in agreement with several reported [
25,
41,
48] results where cooking time was not associated with protein content in their study. However, cooking quality traits were shown to be significantly and positively correlated with grain quality traits. For example, cooking time is correlated with seed length, seed width, and 100 seed weight (cooking time increases with seed size). The implication is that as breeding programs strive to develop varieties that are large-seeded with short cooking times to meet market demands, there will be a trade-off between these two traits. There is a need to devise breeding strategies that maximize both of these traits, as well as a careful selection of parental materials. In addition to this, there is a need to dissect the genetic mechanisms of these traits and determine whether any of them can be amenable to trait integration techniques.
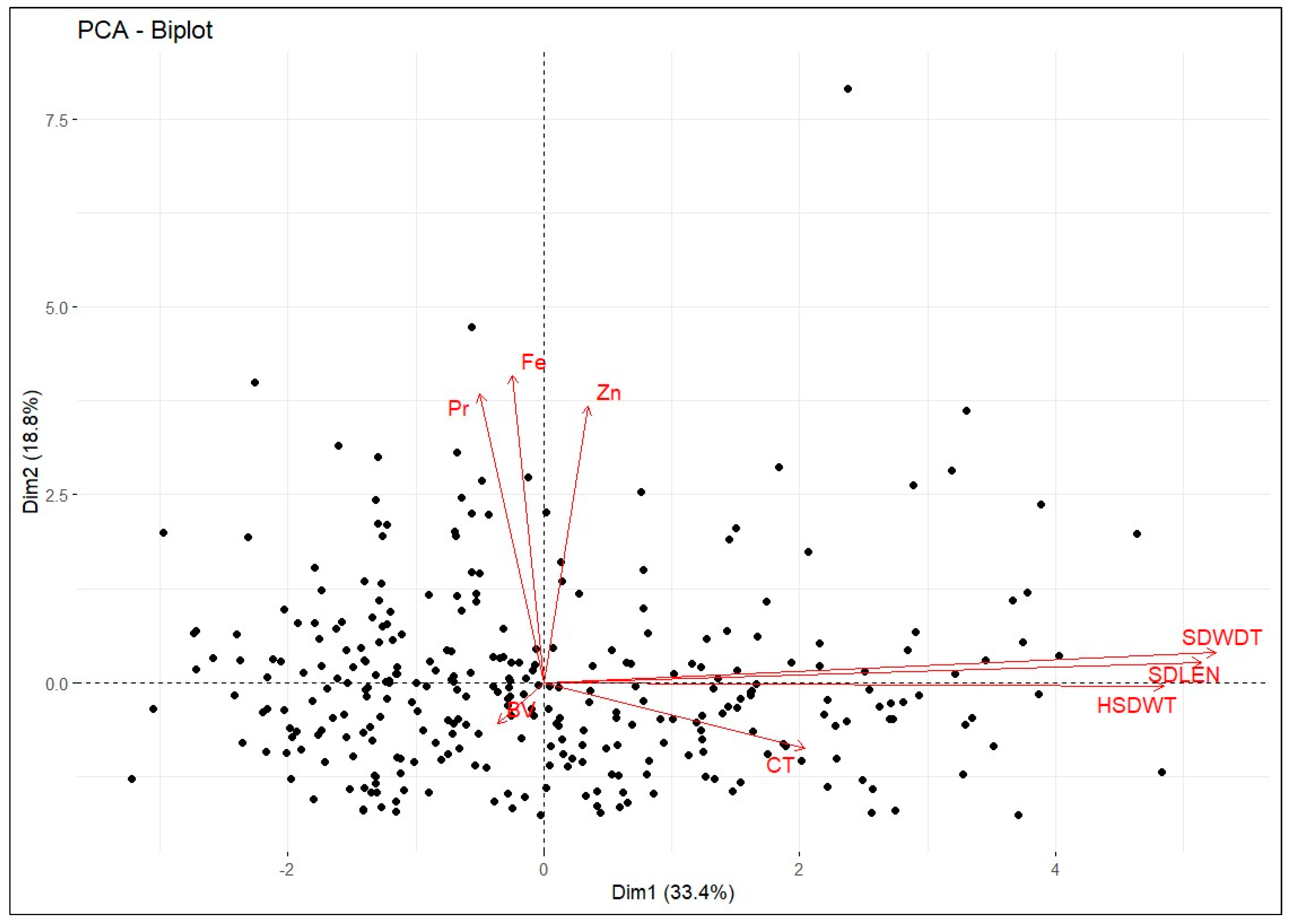
A genotype by trait biplot was produced using the first two PCs, which explained 52.19% of the variability (
Figure 3). This was used to identify the best-performing and genetically dissimilar genotypes that could be further explored for breeding. The random scattering of the cowpea genotypes across the quadrant, as seen in the biplot, suggests that the genotypes were genetically different. The genotypes clustered based on their performance for the traits; thus, those genotypes located close together had relatively similar values for specific traits. The biplot shows that many genotypes clustered near the origin, so they are likely derived from the same parents. Genotypes located further from the biplot origin had more extreme values for a specific trait than genotypes closer to the origin. For example, genotypes MWcp03 and MWcp36 showed extreme values for seed length, width, and 100 seed weight, whereas TVu-3533 showed higher values for protein, iron, and zinc concentrations. On the other hand, TVu-322 had the highest broth viscosity, and MWcp46 had the longest cooking time. Furthermore, MWcp37 was distant from the rest of the genotypes, indicating that the genotype is genetically dissimilar from most of the genotypes. Concentrations of protein, iron, and zinc were independent of broth viscosity, cooking time, seed length, width, and 100 seed weight. This suggests that protein, iron, and zinc can be improved without altering the rest of the traits. Noticeably, broth viscosity was not associated with cooking time or the quantitative grain quality traits. However, cooking time was positively related to the quantitative grain quality traits. This result further suggests that increasing the seed size in cowpeas may increase the cooking time but not the broth viscosity. As already elucidated, the landraces from Malawi were generally large-seeded with longer cooking times, but these tend to be common among smallholder farmers. This was further confirmed through cluster analysis, which clustered the majority of the landraces from Malawi into Cluster II, which had large-seeded genotypes with relatively higher protein and iron content. Nevertheless, genotypes in Cluster I were small-seeded, and within the cluster were most of the improved varieties and advanced lines from the IITA, which is an indication that they were selected for similar traits, mostly small seed size, which is contrary to farmer preferences.







{kind=link}
{kind=link}
{kind=link}