
Solid-State Fermentation Using Bacillus licheniformis-Driven Changes in Composition, Viability and In Vitro Protein Digestibility of Oilseed Cakes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material and Microorganism
2.3. Solid-State Fermentation (SSF)
2.4. Microbial Viability, Spore Number, Colony Morphology, and pH Value
2.4.1. Determination of Microbial Viability
2.4.2. Determination of Spore Number
2.4.3. Determination of pH Value
2.5. Proximate Composition
2.6. Fatty Acid Profile
2.7. Reducing Sugars
2.8. Soluble Proteins and Peptides, Extraction, and Quantification
2.8.1. Determination of Soluble Proteins
2.8.2. Determination of Peptides
2.9. In Vitro Protein Digestibility (IVPD) and Microorganism Viability under Simulated Conditions
2.10. Statistical Analysis
3. Results
3.1. Effect of Fermentation on Proximate Composition and Fatty Acid Profile
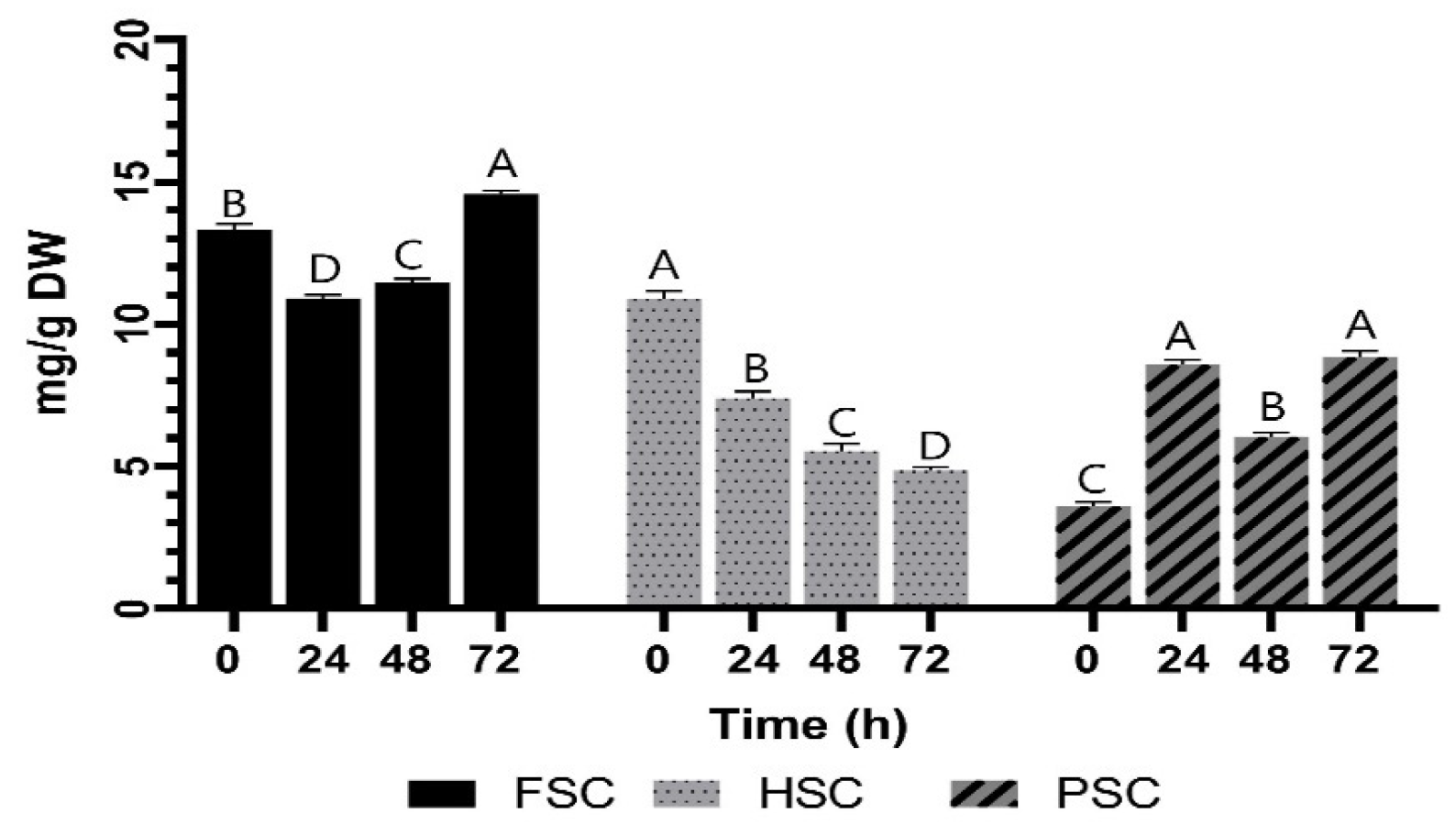
3.2. Effect of Fermentation pH and Reducing Sugars
3.3. Effect of Fermentation on Growth Dynamics and Spore Formation
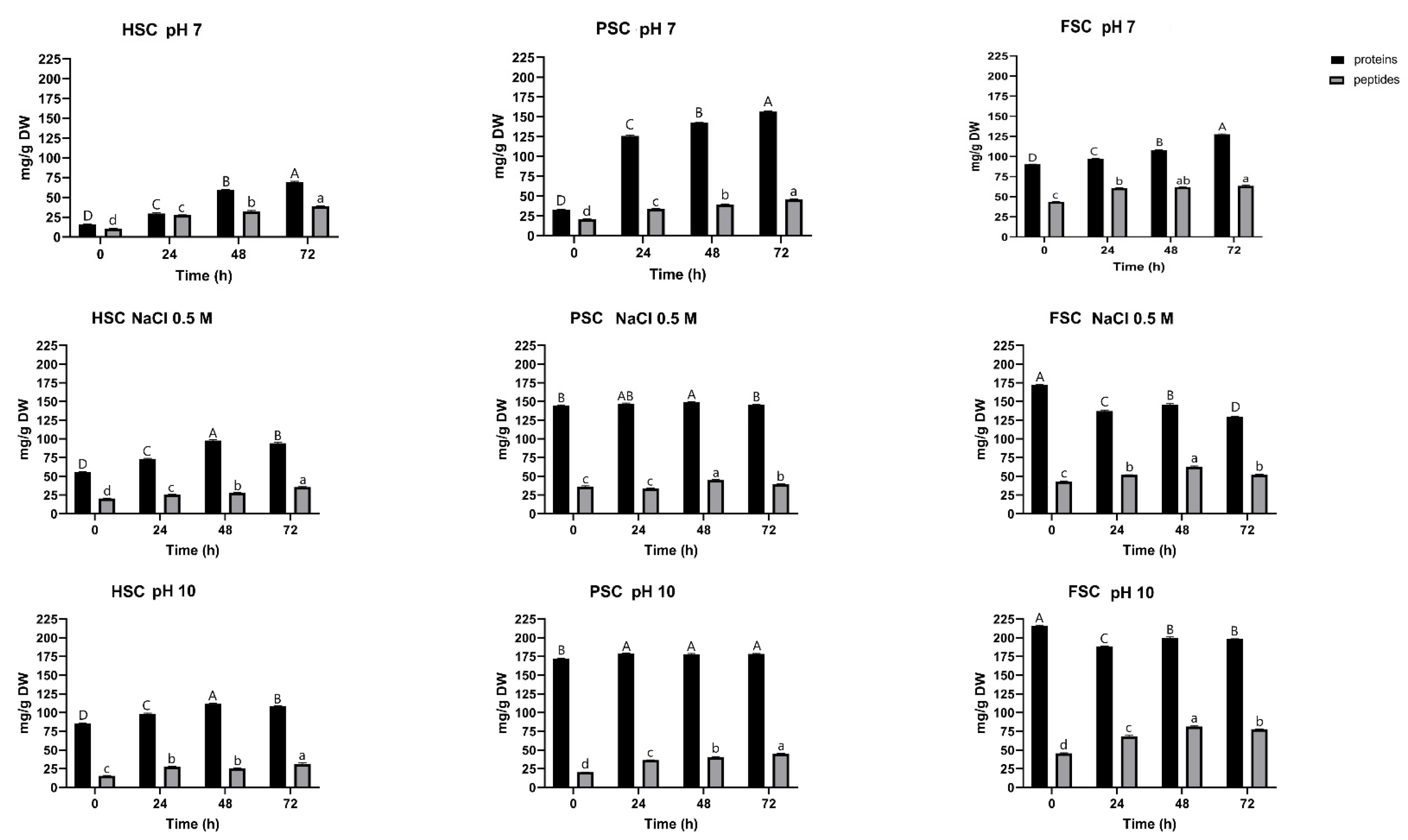
3.4. Effect of Fermentation on Protein Solubility and Peptide Content
3.5. Bacillus licheniformis Survival under Simulated GI Conditions
3.6. In Vitro Protein Digestibility
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| HSC | hemp seed cake |
| PSC | pumpkin seed cake |
| FSC | flaxseed cake |
References
- Singh, R.; Langyan, S.; Sangwan, S.; Rohtagi, B.; Khandelwal, A.; Shrivastava, M. Protein for human consumption from oilseed cakes: A Review. Front. Sustain. Food Syst. 2022, 6, 856401. [Google Scholar] [CrossRef]
- Dozier, W.A.; Hess, J.B. Soybean meal quality and analytical techniques. In Soybean and Nutrition; InTech Open: London, UK, 2011; pp. 111–124. [Google Scholar]
- Keller, M.; Reidy, B.; Scheurer, A.; Eggerschwiler, L.; Morel, I.; Giller, K. Soybean meal can be replaced by faba beans, pumpkin seed cake, spirulina or be completely omitted in a forage-based diet for fattening bulls to achieve comparable performance, carcass and meat quality. Animals 2021, 11, 1588. [Google Scholar] [CrossRef] [PubMed]
- Ancuța, P.; Sonia, A. Oil Press-Cakes and Meals Valorization through Circular Economy Approaches: A Review. Appl. Sci. 2020, 10, 7432. [Google Scholar] [CrossRef]
- Lekha, P.K.; Lonsane, B.K. Production and application of tannin acyl hydrolase: State of the art. Adv. Appl. Microbiol. 1997, 44, 215–260. [Google Scholar] [CrossRef] [PubMed]
- Lizardi-Jiménez, M.A.; Hernández-Martínez, R. Solid state fermentation (SSF): Diversity of applications to valorize waste and biomass. 3 Biotech 2017, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Cerda, A.; Artola, A.; Barrena, R.; Font, X.; Gea, T.; Sánchez, A. Innovative production of bioproducts from organic waste through solid-state fermentation. Front. Sustain. Food Syst. 2019, 3, 63. [Google Scholar] [CrossRef]
- Soares, V.F.; Castilho, L.R.; Bon, E.P.; Freire, D.M. High-yield Bacillus subtilis protease production by solid-state fermentation. Appl. Biochem. Biotechnol. 2005, 121–124, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Jatuwong, K.; Kumla, J.; Suwannarach, N.; Matsui, K.; Lumyong, S. Bioprocessing of agricultural residues as substrates and optimal conditions for phytase production of Chestnut Mushroom, Pholiota adiposa, in solid state fermentation. J. Fungi 2020, 6, 384. [Google Scholar] [CrossRef]
- Sadh, P.K.; Kumar, S.; Chawla, P.; Duhan, J.S. Fermentation: A boon for production of bioactive compounds by processing of food industries wastes (by-products). Molecules 2018, 23, 2560. [Google Scholar] [CrossRef]
- Olukomaiya, O.; Fernando, C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Solid-state fermented plant protein sources in the diets of broiler chickens: A review. Anim. Nutr. 2019, 5, 319–330. [Google Scholar] [CrossRef]
- Manan, M.A.; Webb, C. Design aspects of solid state fermentation as applied to microbial bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 511–532. [Google Scholar] [CrossRef]
- Ramirez-Olea, H.; Reyes-Ballesteros, B.; Chavez-Santoscoy, R.A. Potential application of the probiotic Bacillus licheniformis as an adjuvant in the treatment of diseases in humans and animals: A systematic review. Front. Microbiol. 2022, 26, 993451. [Google Scholar] [CrossRef]
- Bernardeau, M.; Lehtinen, M.J.; Forssten, S.D.; Nurminen, P. Importance of the gastrointestinal life cycle of Bacillus for probiotic functionality. J. Food Sci. Technol. 2017, 54, 2570–2584. [Google Scholar] [CrossRef]
- Solanki, P.; Putatunda, C.; Kumar, A.; Bhatia, R.; Walia, A. Microbial proteases: Ubiquitous enzymes with innumerable uses. 3 Biotech 2021, 11, 428. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Yan, P.; Xu, X.; Huang, L.; Dabbour, M.; Benjamin, K.M.; He, R.; Ma, H. Effect of single and two-stage fermentation on the antioxidative activity of soybean meal, and the structural and interfacial characteristics of its protein. LWT 2023, 183, 114938. [Google Scholar] [CrossRef]
- Liu, D.; Guo, Y.; Ma, H. Production of value-added peptides from agro-industrial residues by solid-state fermentation with a new thermophilic protease-producing strain. Food Biosci. 2023, 53, 102534. [Google Scholar] [CrossRef]
- Wu, P.; Guo, Y.; Golly, M.K.; Ma, H.; He, R.; Luo, S.; Zhang, C.; Zhang, L.; Zhu, J. Feasibility study on direct fermentation of soybean meal by Bacillus stearothermophilus under non-sterile conditions. JSFAAE 2018, 99, 3291–3298. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Hou, Y.; Xu, H.; Huang, L.; Dabbour, M.; Mintah, B.K.; He, R.; Ma, H. Effect of solid-state fermentation by three different Bacillus species on composition and protein structure of soybean meal. J. Sci. Food Agric. 2022, 102, 557–566. [Google Scholar] [CrossRef]
- Dai, C.; Ma, H.; He, R.; Huang, L.; Zhu, S.; Ding, Q.; Luo, L. Improvement of nutritional value and bioactivity of soybean meal by solid-state fermentation with Bacillus subtilis. LWT 2017, 86, 1–7. [Google Scholar] [CrossRef]
- González Pereyra, M.L.; Di Giacomo, A.L.; Lara, A.L.; Martínez, M.P.; Cavaglieri, L.; Gonz, M.L. Aflatoxin-degrading Bacillus sp. strains degrade zearalenone and produce proteases, amylases and cellulases of agro-industrial interest. Toxicon 2020, 180, 43–48. [Google Scholar] [CrossRef]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Yi, Z.; Zhu, H.; Pan, K.; Jing, B.; et al. Bacillus licheniformis normalize the ileum microbiota of chickens infected with necrotic enteritis. Sci. Rep. 2018, 8, 1744. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.-R.; Cheng, Y.-H.; Hsiao, F.S.-H.; Proskura, W.S.; Dybus, A.; Yu, Y.-H. Optimization of solid-state fermentation conditions of Bacillus licheniformis and its effects on Clostridium perfringens-induced necrotic enteritis in broilers. Rev. Bras. Zootec. 2019, 48, e20170298. [Google Scholar] [CrossRef]
- Yang, L.; Zeng, X.; Qiao, S. Advances in research on solid-state fermented feed and its utilization: The pioneer of private customization for intestinal microorganisms. Anim. Nutr. 2021, 7, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Hung, D.-Y.; Cheng, Y.-H.; Chen, W.-J.; Hua, K.-F.; Pietruszka, A.; Dybus, A.; Lin, C.-S.; Yu, Y.-H. Bacillus licheniformis-Fermented Products Reduce Diarrhea Incidence and Alter the Fecal Microbiota Community in Weaning Piglets. Animals 2019, 9, 1145. [Google Scholar] [CrossRef]
- Yi, W.; Liu, Y.; Fu, S.; Zhuo, J.; Wang, J.; Shan, T. Dietary novel alkaline protease from Bacillus licheniformis improves broiler meat nutritional value and modulates intestinal microbiota and metabolites. Anim Microbiome 2024, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, K.; Ma, L.; Zhao, J.; Zhao, Z.M. Effects of drying strategies on sporulation and titer of microbial ecological agents with Bacillus subtilis. Front. Nutr. 2022, 9, 1025248. [Google Scholar] [CrossRef]
- AOAC. Association of Official Agricultural Chemists, 17th ed.; Official Methods of Analysis of AOAC International: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Vlaicu, P.A.; Untea, A.E.; Turcu, R.P.; Saracila, M.; Panaite, T.D.; Cornescu, G.M. Nutritional composition and bioactive compounds of basil, thyme and sage plant additives and their functionality on broiler thigh meat quality. Foods 2022, 11, 1105. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Lefter, N.A.; Toma, S.M.; Dumitru, M.; Cismileanu, A.; Surdu, I.; Gheorghe, A.; Dragomir, C.; Untea, A. Changes in ileum and cecum volatile fatty acids and their relationship with microflora and enteric methane in pigs fed different fiber levels. Agriculture 2022, 12, 451. [Google Scholar] [CrossRef]
- Ciurescu, G.; Vasilachi, A.; Idriceanuand, L.; Dumitru, M. Effects of corn replacement by sorghum in broiler chickens diets on performance, blood chemistry, and meat quality. Ital. J. Anim. Sci. 2023, 12, 537–547. [Google Scholar] [CrossRef]
- ISO/TS 17764-2:2008; Animal Feeding Stuffs. ISO: Geneva, Switzerland, 2008.
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, M.; Sun, W. Effect of protein oxidation on the in vitro digestibility of soy protein isolate. Food Chem. 2013, 141, 3224–3229. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Marttinen, M.; Anjum, M.; Saarinen, M.T.; Ahonen, I.; Lehtinen, M.J.; Nurminen, P.; Laitila, A. Enhancing bioaccessibility of plant protein using probiotics: An in vitro study. Nutrients 2023, 15, 3905. [Google Scholar] [CrossRef]
- Coelho, M.C.; Ribeiro, T.B.; Oliveira, C.; Batista, P.; Castro, P.; Monforte, A.R.; Rodrigues, A.S.; Teixeira, J.; Pintado, M. In Vitro gastrointestinal digestion impact on the bioaccessibility and antioxidant capacity of bioactive compounds from tomato flours obtained after conventional and ohmic heating extraction. Foods 2021, 10, 554. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Ogawa, Y. In vitro protein digestibility and biochemical characteristics of soaked, boiled and fermented soybeans. Sci. Rep. 2021, 11, 14257. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.E.; Moraru, C.I. Effect of High pressure processing and heat treatment on in vitro digestibility and trypsin inhibitor activity in lentil and faba bean protein concentrates. LWT 2011, 152, 112342. [Google Scholar] [CrossRef]
- Allagheny, N.; Obanu, Z.A.; Campbell-Platt, G.; Owens, J.D. Control of ammonia formation during Bacillus subtilis fermentation of legumes. Int. J. Food Microbiol. 1996, 29, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Akanni, G.B.; De Kock, H.L.; Naudé, Y.; Buys, E.M. Volatile compounds produced by Bacillus species alkaline fermentation of bambara groundnut (Vigna subterranean (L.) Verdc) into a dawadawa-type African food condiment using headspace solid-phase microextraction and GC × GC–TOFMS. Int. J. Food Prop. 2018, 21, 929–941. [Google Scholar] [CrossRef]
- Bernacchia, R.; Preti, R.; Vinci, G. Chemical composition and health benefits of flaxseed. Austin. J. Nutri. Food Sci. 2014, 2, 1045. [Google Scholar]
- Habib, A.; Biswas, S.; Siddique, A.H.; Mohammad, M.; Uddin, B.; Hasan, S.; MMH, K.; Uddin, M.; Islam, M.; Hasan, M.; et al. Nutritional and lipid composition analysis of pumpkin seed (Cucurbita maxima Linn.). J. Nutr. Food Sci. 2015, 5, 4. [Google Scholar] [CrossRef]
- Montero, L.; Ballesteros-Vivas, D.; Gonzalez-Barrios, A.F.; Sánchez-Camargo, A.D.P. Hemp seeds: Nutritional value, associated bioactivities and the potential food applications in the colombian context. Front. Nutr. 2023, 11, 1039180. [Google Scholar] [CrossRef] [PubMed]
- Diomandé, S.E.; Guinebretière, M.H.; Broussolle, V.; Brillard, J. Role of fatty acids in Bacillus environmental adaptation. Front. Microbiol. 2015, 6, 813. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Dong, Z.; Chen, X.; Cai, K.; Chen, Z.; Wang, H.; Jin, P.; Liu, X.; Permaul, K.; Singh, S.; Wang, Z.-X. Exploring the Metabolomic Responses of Bacillus licheniformis to Temperature Stress by Gas Chromatography/Mass Spectrometry. J. Microbiol. Biotechnol. 2018, 28, 473–481. [Google Scholar] [CrossRef]
- Lopes, C.; Barbosa, J.; Maciel, E.; Costa, E.; Alves, E.; Ricardo, F.; Domingues, P.; Mendo, S.; Domingues, M.R.M. Decoding the Fatty Acid Profile of Bacillus li-cheniformis I89 and Its Adaptation to Different Growth Conditions to Investigate Possible Biotechnological Applications. Lipids 2019, 54, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]
- Logan, N.A.; Vos, P.D. Bacillus. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–163. [Google Scholar] [CrossRef]
- Vlamakis, H.; Aguilar, C.; Losick, R.; Kolter, R. Control of cell fate by the formation of an architecturally complex bacterial community. Genes Dev. 2008, 22, 945–953. [Google Scholar] [CrossRef]
- Tasaki, S.; Nakayama, M.; Shoji, W. Morphologies of Bacillus subtilis communities responding to environmental variation. DGD 2017, 59, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Elisashvili, V.; Kachlishvili, E.; Chikindas, M.L. Recent advances in the physiology of spore formation for Bacillus probiotic production. Probiotics Antimicrob. Prot. 2019, 11, 731–747. [Google Scholar] [CrossRef]
- Gray, D.A.; Dugar, G.; Gamba, P.; Strahl, H.; Jonker, M.J.; Hamoen, L.W. Extreme slow growth as alternative strategy to survive deep starvation in bacteria. Nat. Commun. 2019, 10, 890. [Google Scholar] [CrossRef]
- Zhao, S.; Deng, L.; Hu, N.; Zhao, B.; Liang, Y. Cost-effective production of Bacillus licheniformis using simple netting bag solid bioreactor. World J. Microb. Biotechnol. 2008, 24, 2859–2863. [Google Scholar] [CrossRef]
- Chistyakov, V.; Mlnikov, V.; Chikindas, M.L.; Khutsishvili, M.; Chagelishvili, A.; Bren, A.; Kostina, N.; Cavera, V.; Elisashvili, V. Poultry-beneficial solid-state Bacillus amyloliquefaciens B-1895 fermented soybean formulation. Biosci. Microbiota Food Health 2015, 34, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Golnari, M.; Bahrami, N.; Milanian, Z.; Rabbani Khorasgani, M.; Asadollahi, M.A.; Shafiei, R.; Fatemi, S.S.A. Isolation and characterization of novel Bacillus strains with superior probiotic potential: Comparative analysis and safety evaluation. Sci. Rep. 2024, 14, 1457. [Google Scholar] [CrossRef] [PubMed]
- Flach, J.; van der Waal, M.B.; van den Nieuwboer, M.; Claassen, E.; Larsen, O.F.A. The underexposed role of food matrices in probiotic products: Reviewing the relationship between carrier matrices and product parameters. Crit. Rev. Food Sci. Nutr. 2018, 58, 2570–2584. [Google Scholar] [CrossRef] [PubMed]
- Branlard, G.; Bancel, E. Protein extraction from cereal seeds. Plant Proteom. 2007, 355, 15–26. [Google Scholar] [CrossRef]
- Osborne, T.B. The Vegetable Proteins; Longmans: London, UK, 1924; Available online: https://wellcomecollection.org/works/p8hecz9k/items?canvas=32 (accessed on 14 February 2024).
- Hadnađev, M.S.; Hadnađev, D.T.; Pojić, M.M.; Šarić, B.M.; Mišan, A.Č.; Jovanov, P.T.; Sakač, M.B. Progress in vegetable proteins isolation techniques: A review. Food Feed. Res. 2017, 44, 11–21. [Google Scholar] [CrossRef]
- Gouseti, O.; Larsen, M.E.; Amin, A.; Bakalis, S.; Petersen, I.L.; Lametsch, R.; Jensen, P.E. Applications of enzyme technology to enhance transition to plant proteins: A Review. Foods 2023, 12, 2518. [Google Scholar] [CrossRef]
- Chen, H.; Xu, B.; Wang, Y.; Li, W.; He, D.; Zhang, Y.; Xing, X. Emerging natural hemp seed proteins and their functions for nutraceutical applications. Food Sci. Hum. Wellness 2023, 12, 929–941. [Google Scholar] [CrossRef]
- Pham, T.T.; Tran, T.T.T.; Ton, N.M.N.; Le, V.V.M. Effects of pH and salt concentration on functional properties of pumpkin seed protein fractions. J. Food Process. Preserv. 2016, 41, e13073. [Google Scholar] [CrossRef]
- Friedman, M.; Gumbmann, M.R.; Masters, P.M. Protein-alkali reactions: Chemistry, toxicology, and nutritional consequences. Nutr. Toxicol. Asp. Food Saf. 1984, 177, 367–412. [Google Scholar] [CrossRef]
- Del Mar Contreras, M.; Lama-Muñoz, A.; Manuel Gutiérrez-Pérez, J.; Espínola, F.; Moya, M.; Castro, E. Protein extraction from agri-food residues for integration in biorefinery: Potential techniques and current status. Bioresour. Technol. 2019, 280, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, T.; Guo, G.; Liu, Z.; Hao, N. Production of Nattokinase from Hemp Seed Meal by Solid-State Fermentation and Improvement of Its Nutritional Quality. Fermentation 2023, 9, 469. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B. Fermentation with Edible Rhizopus Strains to Enhance the Bioactive Potential of Hull-Less Pumpkin Oil Cake. Molecules 2020, 25, 5782. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.-F.; Chen, Y.-A.; Luo, C.; Chiang, W.-D. Antioxidant and antibacterial activities of peptide fractions from flaxseed protein hydro-lysed by protease from Bacillus altitudinis HK02. Int. J. Food Sci. Technol. 2016, 51, 681–689. [Google Scholar] [CrossRef]
- Li, X.; Cheng, Y.; Yi, C.; Hua, Y.; Yang, C.; Cui, S. Effect of ionic strength on the heat-induced soy protein aggregation and the phase separation of soy protein aggregate/dextran mixtures. Food Hydrocoll. 2009, 23, 1015–1023. [Google Scholar] [CrossRef]
- Song, P.; Zhang, X.; Wang, S.; Xu, W.; Wang, F.; Fu, R.; Wei, F. Microbial proteases and their applications. Front. Microbiol. 2023, 14, 1236368. [Google Scholar] [CrossRef]
- Ayad, A. Characterization and Properties of Flaxseed Protein Fractions. 2010. Available online: https://www.google.com.hk/url?sa=t&source=web&rct=j&opi=89978449&url=https://escholarship.mcgill.ca/downloads/hh63sw24h.pdf&ved=2ahUKEwiZp5L_is2FAxVsk1YBHT30BvEQFnoECBwQAQ&usg=AOvVaw2eSSYehMgfvPYYv7M3kLSb (accessed on 14 February 2024).
- Pontonio, E.; Verni, M.; Dingeo, C.; Diaz-de-Cerio, E.; Pinto, D.; Rizzello, C.G. Impact of Enzymatic and Microbial Bioprocessing on Antioxidant Properties of Hemp (Cannabis sativa L.). Antioxidants 2020, 9, 1258. [Google Scholar] [CrossRef] [PubMed]
- Giami, S.Y. Effect of fermentation on the seed proteins, nitrogenous constituents, antinutrients and nutritional quality of fluted pumpkin (Telfairia occidentalis Hook). Food Chem. 2004, 88, 397–404. [Google Scholar] [CrossRef]
- Marambe, H.K.; Shand, P.J.; Wanasundara, J.P.D. In vitrodigestibility of flaxseed (Linum usitatissimum L.) protein: Effect of seed mucilage, oil and thermal processing. Int. J. Food Sci. Technol. 2012, 48, 628–635. [Google Scholar] [CrossRef]
- Wu, M.; Wang, L.; Li, D.; Wang, Y.; Mao, Z.; Chen, X.D. The Digestibility and Thermal Properties of Fermented Flaxseed Protein. Int. J. Food Eng. 2012, 8. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | HSC T 0 h | HSC T 72 h | PSC T 0 h | PSC T 72 h | FSC T 0 h | FSC T 72 h |
|---|---|---|---|---|---|---|
| DM (%) | 93.58 ± 0.38 | 95.82 ± 0.1 | 90.91 ± 0.1 | 95.17 ± 0.03 | 90.54 ± 0.06 | 96.44 ± 0.05 |
| CP (% DM) | 38.00 ± 0.22 | 35.95 ± 0.08 | 45.27 ± 0.1 | 44.65 ± 0.17 | 33.35 ± 0.13 | 32.53 ± 0.07 |
| Crude fat (% DM) | 8.44 ± 0.11 | 8.53 ± 0.08 | 15.60 ± 0.09 | 15.05 ± 0.1 | 18.82 ± 0.09 | 19.15 ± 0.14 |
| Carbohydrates (% DM) | 17.10 ± 0.05 | 18.24 ± 0.27 | 1.55 ± 0.58 | 3.45 ± 0.47 | 34.07 ± 0.149 | 31.21 ± 0.56 |
| CF (% DM) | 30.23 ± 0.09 | 30.14 ± 0.04 | 31.18 ± 0.29 | 32.27 ± 0.15 | 10.44 ± 0.23 | 11.78 ± 0.22 |
| NDF (% DM) | 43.98 ± 0.13 | 47.16 ± 0.1 | 45.28 ± 0.28 | 40.64 ± 0.33 | 27.32 ± 0.17 | 28.22 ± 0.24 |
| ADF (% DM) | 29.96 ± 0.25 | 33.83 ± 0.12 | 29.60 ± 0.17 | 30.92 ± 0.19 | 13.99 ± 0.21 | 16.82 ± 0.13 |
| Ash (% DM) | 6.20 ± 0.06 | 7.11 ± 0.06 | 6.37 ± 0.09 | 4.56 ± 0.05 | 4.11 ± 0.09 | 4.48 ± 0.06 |
| FA (g/100 g) | HSC T 0 h | HSC F 72 h | PSC T 0 h | PSC F 72 h | FSC T 0 h | FSC F 72 h |
|---|---|---|---|---|---|---|
| Myristic C14:0 | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.12 ± 0.02 | 0.11 ± 0.01 | 0.09 ± 0.02 | 0.05 ± 0.01 |
| Pentadecanoic C15:0 | 0.04 ± 0.01 | 0.11 ± 0.02 | 0.04 ± 0.01 | 0.19 ± 0.02 | ND | 0.22 ± 0.02 |
| Palmitic C16:0 | 8.61 ± 0.05 | 9.29 ± 0.04 | 14.53 ± 0.09 | 12.65 ± 0.15 | 7.67 ± 0.13 | 7.51 ± 0.19 |
| Heptadecanoic C17:0 | 0.72 ± 0.04 | ND | 0.03 ± 0.0 | 0.05 ± 0.0 | ND | 0.08 ± 0.01 |
| Stearic C18:0 | 2.66 ± 0.04 | 3.05 ± 0.06 | 5.03 ± 0.07 | 5.45 ± 0.06 | 3.32 ± 0.08 | 3.99 ± 0.2 |
| Arachic C20:0 | 1.85 ± 0.03 | 1.75 ± 0.05 | ND | ND | ND | ND |
| TOTAL SATURATED | 13.97 ± 0.08 | 14.28 ± 0.18 | 19.76 ± 0.19 | 18.45 ± 0.22 | 11.09 ± 0.24 | 12.04 ± 0.38 |
| Pentadecenoic C15:1 | 0.12 ± 0.02 | ND | 0.07 ± 0.02 | ND | 0.05 ± 0 | ND |
| Palmitoleic C16:1 | 0.19 ± 0.02 | 0.12 ± 0.01 | 0.11 ± 0.03 | 0.05 ± 0.0 | 0.08 ± 0.01 | 0.06 ± 0.01 |
| Heptadecenoic C17:1 | 0.81 ± 0.07 | ND | ND | 0.05 ± 0.0 | ND | 0.73 ± 0.07 |
| Oleic C18:1 | 11.49 ± 0.05 | 11.71 ± 0.1 | 28.4 ± 0.3 | 29.51 ± 0.13 | 21.9 ± 0.37 | 22.77 ± 0.08 |
| TOTAL MONO- UNSATURATED | 12.61 ± 0.16 | 11.84 ± 0.12 | 28.59 ± 0.31 | 29.61 ± 0.13 | 22.04 ± 0.38 | 23.56 ± 0.13 |
| Linoleic Cis C18:2n6 | 54.13 ± 0.07 | 53.96 ± 0.08 | 48.48 ± 0.41 | 50.23 ± 0.24 | 15.05 ± 0.08 | 14.74 ± 0.06 |
| Linolenic γ C18:3n6 | 0.09 ± 0.02 | ND | ND | ND | 0.21 ± 0.03 | ND |
| α Linolenic C18:3n3 | 17.4 ± 0.04 | 16.33 ± 0.06 | 0.39 ± 0.04 | 0.3 ± 0.04 | 51.14 ± 0.07 | 49.34 ± 0.67 |
| Octadecatetraenoic C18:4n3 | 0.66 ± 0.03 | 1.7 ± 0.08 | 0.24 ± 0.02 | 0.33 ± 0.06 | 0.16 ± 0.02 | 0.24 ± 0.02 |
| Eicosadienoic C20:2n6 | 0.33 ± 0.03 | 0 ± 0 | 0.38 ± 0.04 | 0.14 ± 0.04 | 0.07 ± 0.0 | ND |
| Eicosatrienoic C20:3n6 | ND | ND | ND | ND | ND | ND |
| Arachidonic C20:4n6 | 0.07 ± 0.0 | 0.47 ± 0.04 | 0.17 ± 0 | ND | ND | ND |
| Eicosapentaenoic C20:5n3 | 0.02 ± 0.0 | ND | 0.22 ± 0.04 | ND | ND | ND |
| Docosapentaenoic C22:5n3 | ND | 0.12 ± 0.03 | ND | ND | ND | ND |
| Docosadienoic C22:2n6 | 0.22 ± 0.02 | 0.68 ± 0.04 | 0.16 ± 0.02 | 0.14 ± 0 | ND | ND |
| Other Fatty Acids | 0.49 ± 0.02 | 0.68 ± 0.06 | 0.6 ± 0.08 | ND | 0.11 ± 0.0 | ND |
| TOTAL POLY- UNSATURATED | 73.44 ± 0.22 | 73.95 ± 0.41 | 50.81 ± 0.54 | 51.43 ± 0.42 | 66.76 ± 0.21 | 64.33 ± 0.72 |
| n-6/n-3 ratio | 0.75 | 0.75 | 0.98 | 0.99 | 0.23 | 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rambu, D.; Dumitru, M.; Ciurescu, G.; Vamanu, E. Solid-State Fermentation Using Bacillus licheniformis-Driven Changes in Composition, Viability and In Vitro Protein Digestibility of Oilseed Cakes. Agriculture 2024, 14, 639. https://doi.org/10.3390/agriculture14040639
Rambu D, Dumitru M, Ciurescu G, Vamanu E. Solid-State Fermentation Using Bacillus licheniformis-Driven Changes in Composition, Viability and In Vitro Protein Digestibility of Oilseed Cakes. Agriculture. 2024; 14(4):639. https://doi.org/10.3390/agriculture14040639
Chicago/Turabian StyleRambu, Dan, Mihaela Dumitru, Georgeta Ciurescu, and Emanuel Vamanu. 2024. "Solid-State Fermentation Using Bacillus licheniformis-Driven Changes in Composition, Viability and In Vitro Protein Digestibility of Oilseed Cakes" Agriculture 14, no. 4: 639. https://doi.org/10.3390/agriculture14040639