Metabolic Changes in Synechocystis PCC6803 upon Nitrogen-Starvation: Excess NADPH Sustains Polyhydroxybutyrate Accumulation

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
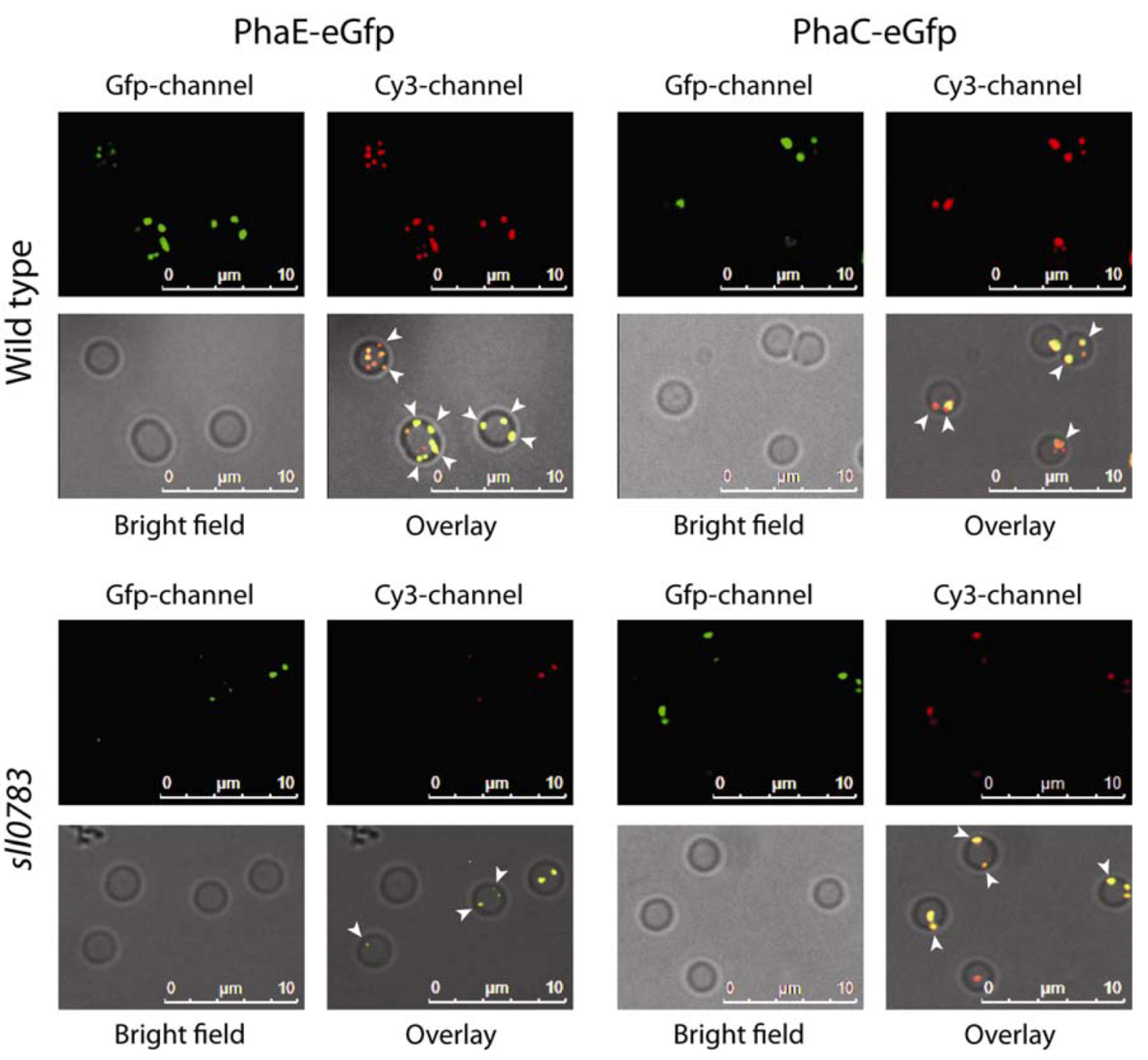
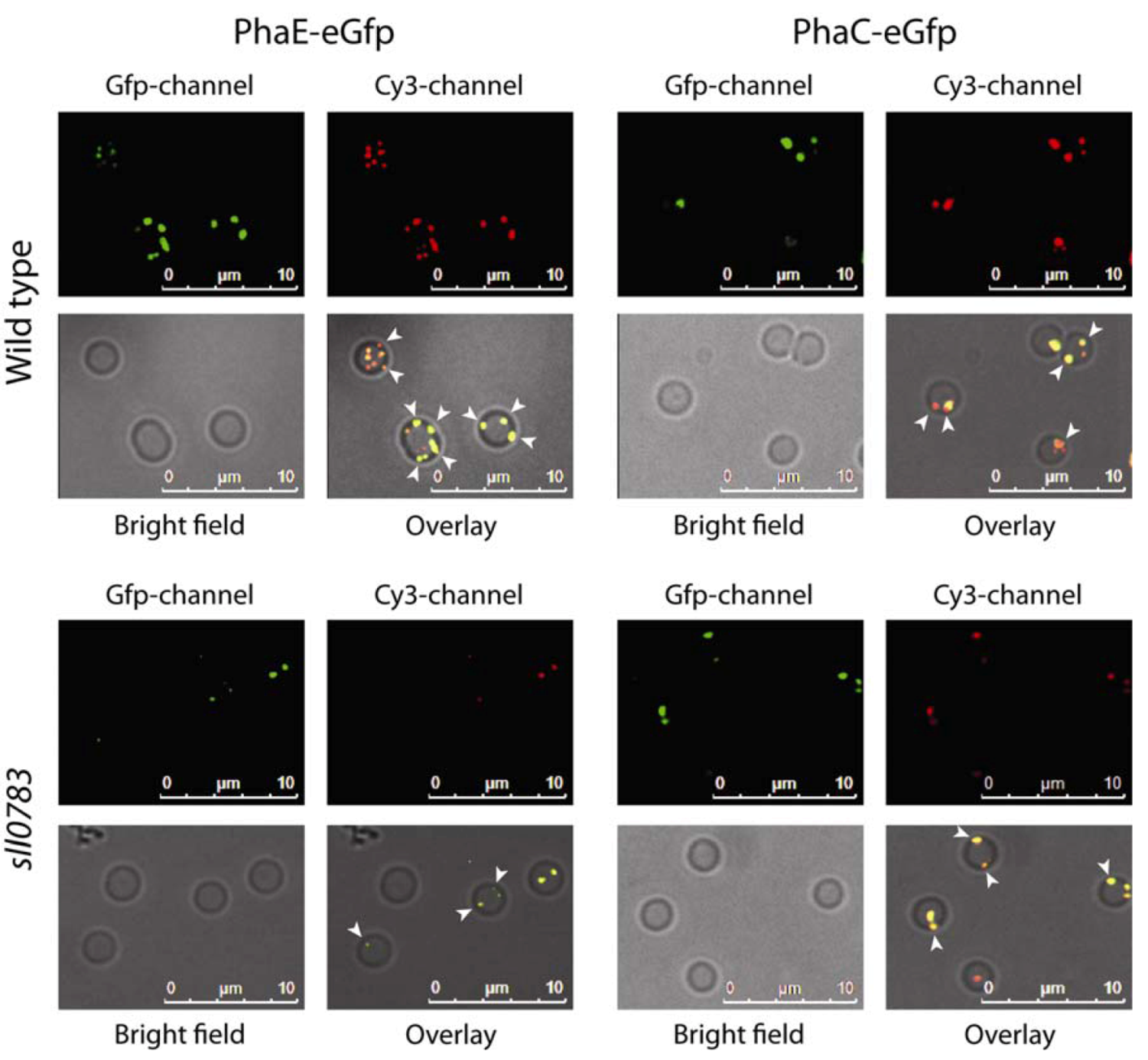
2.1. Intracellular Localization of PHB Synthase
2.2. Metabolome Analysis of Wild Type Synechocystis sp. PCC 6803 and sll0783 Mutant
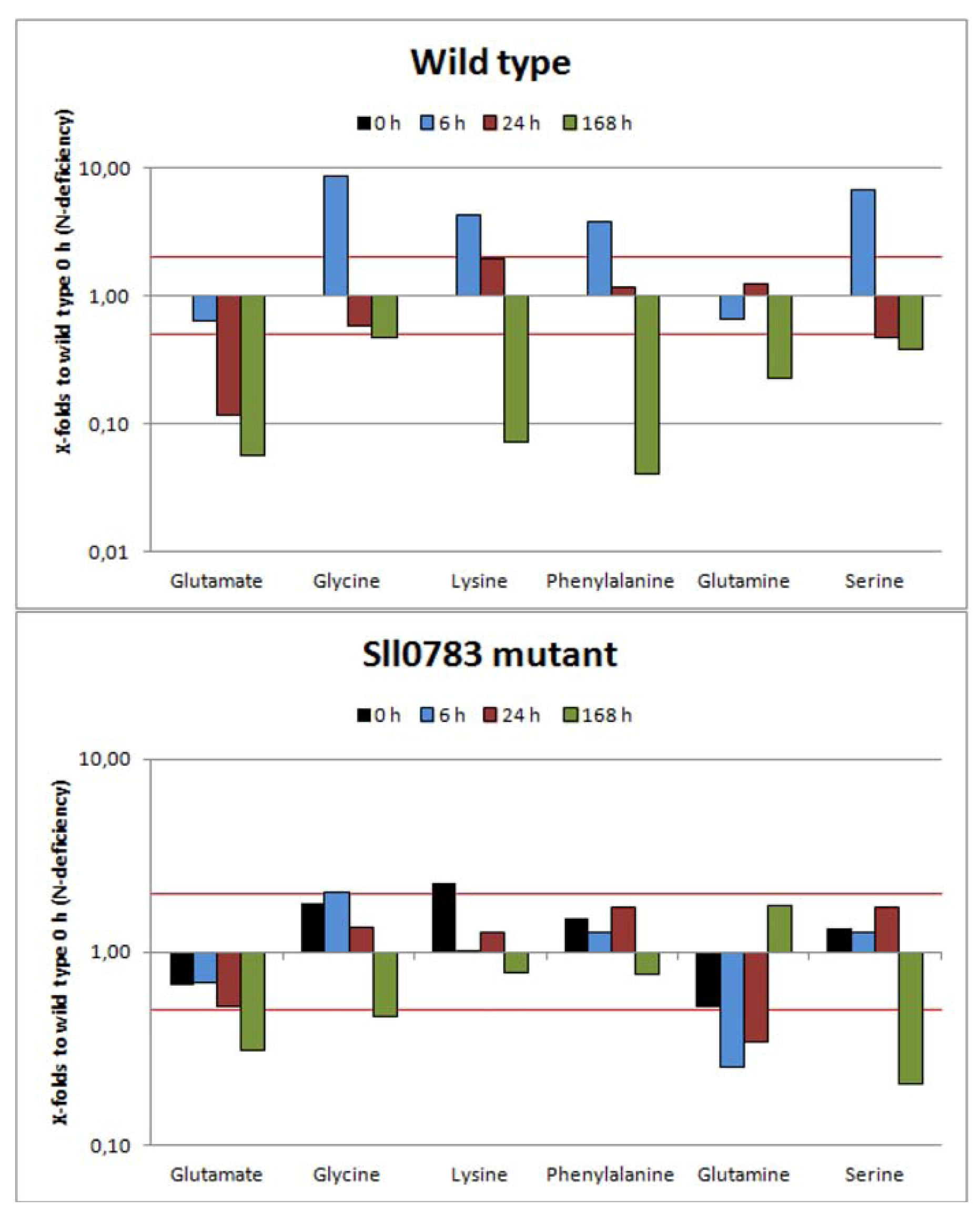
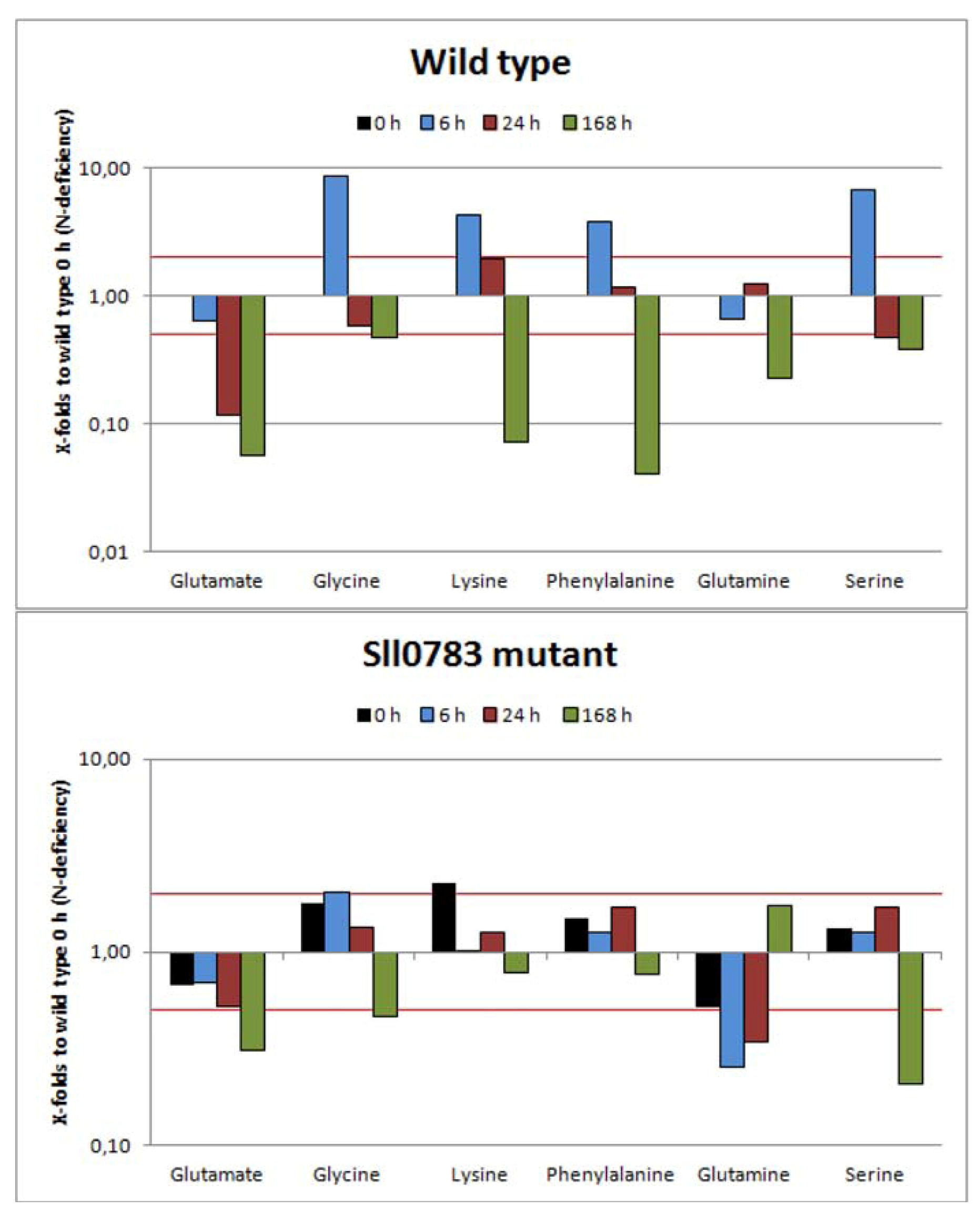
2.2.1. Amino Acids
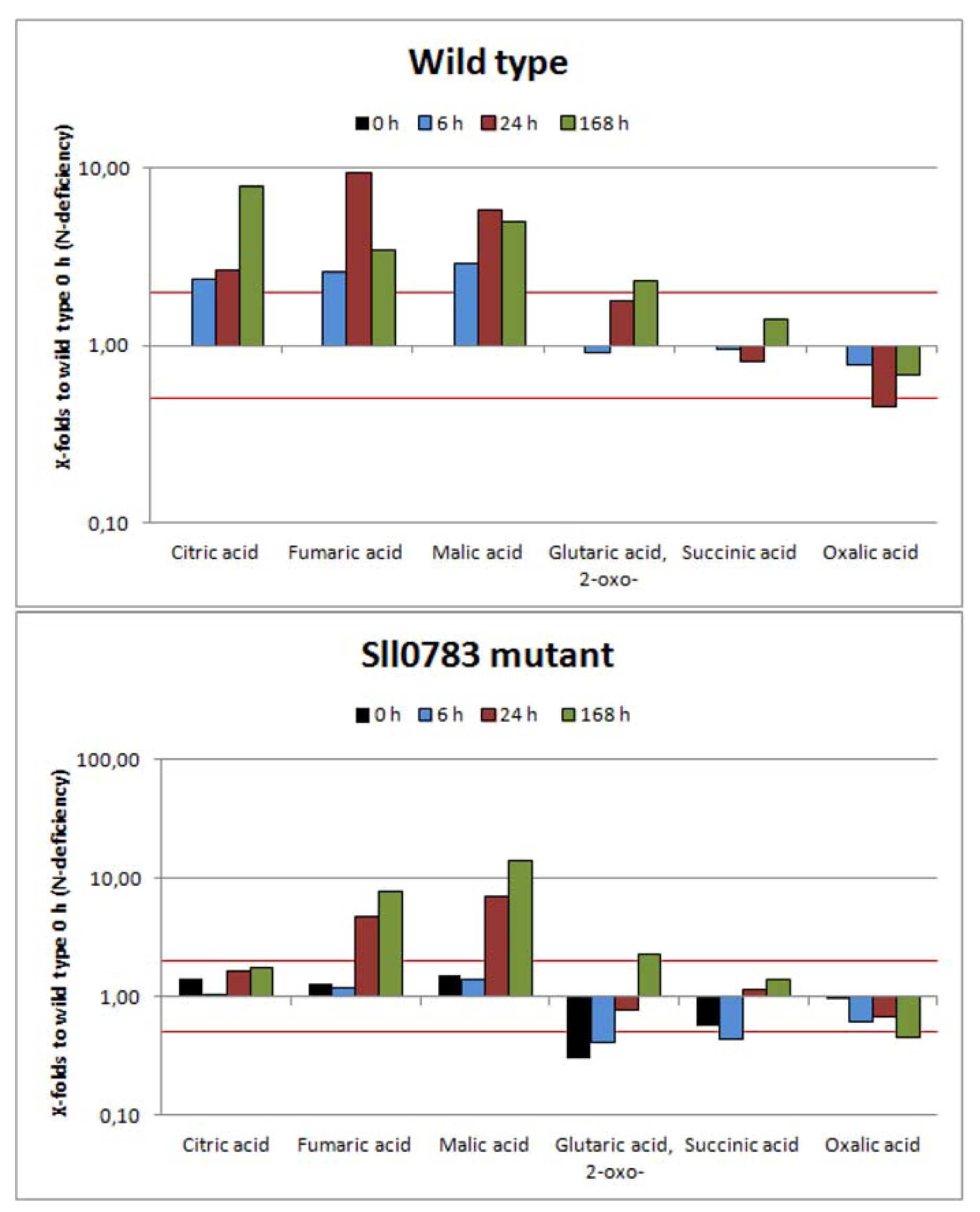
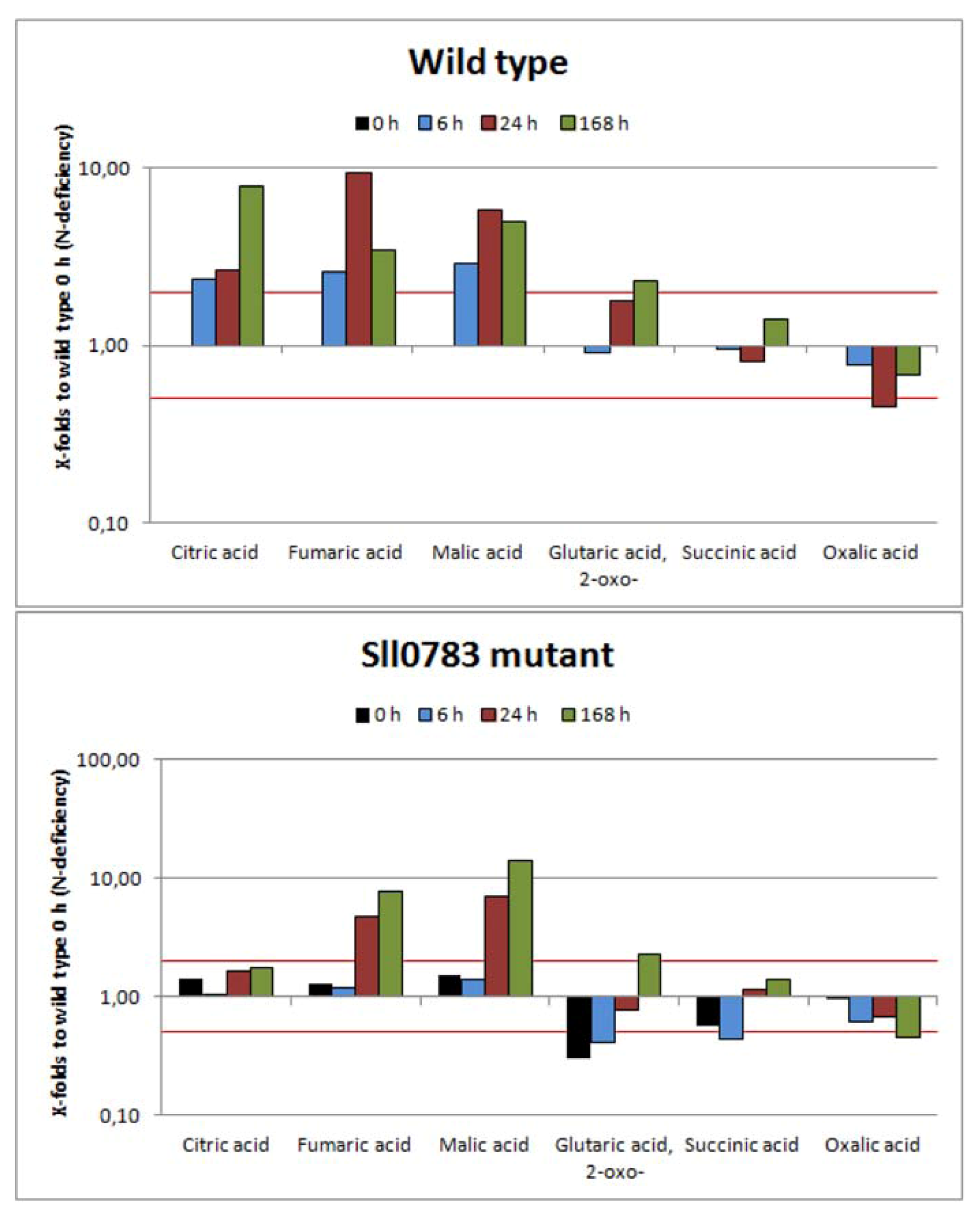
2.2.2. TCA-Cycle
2.2.3. Sugar Metabolism
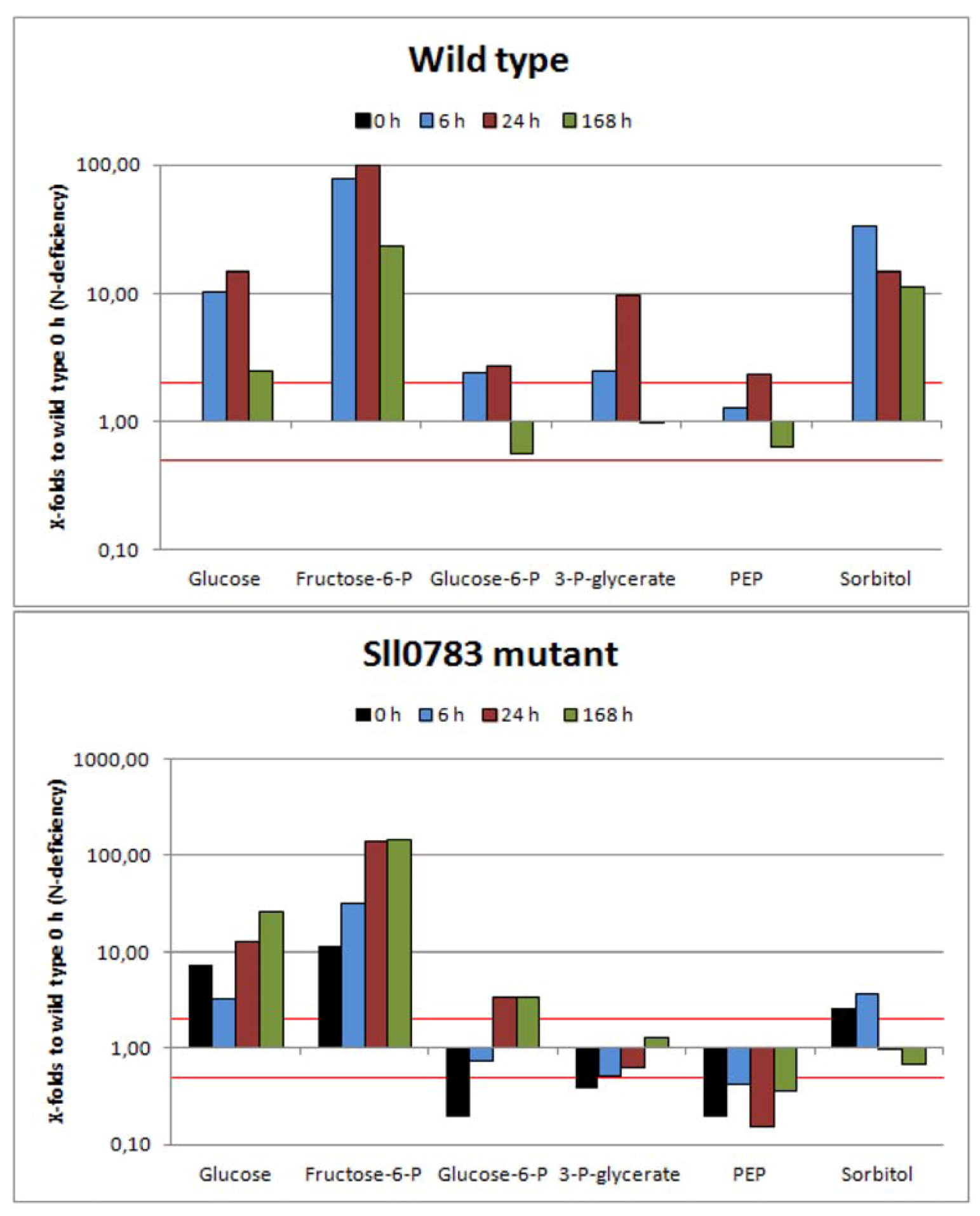
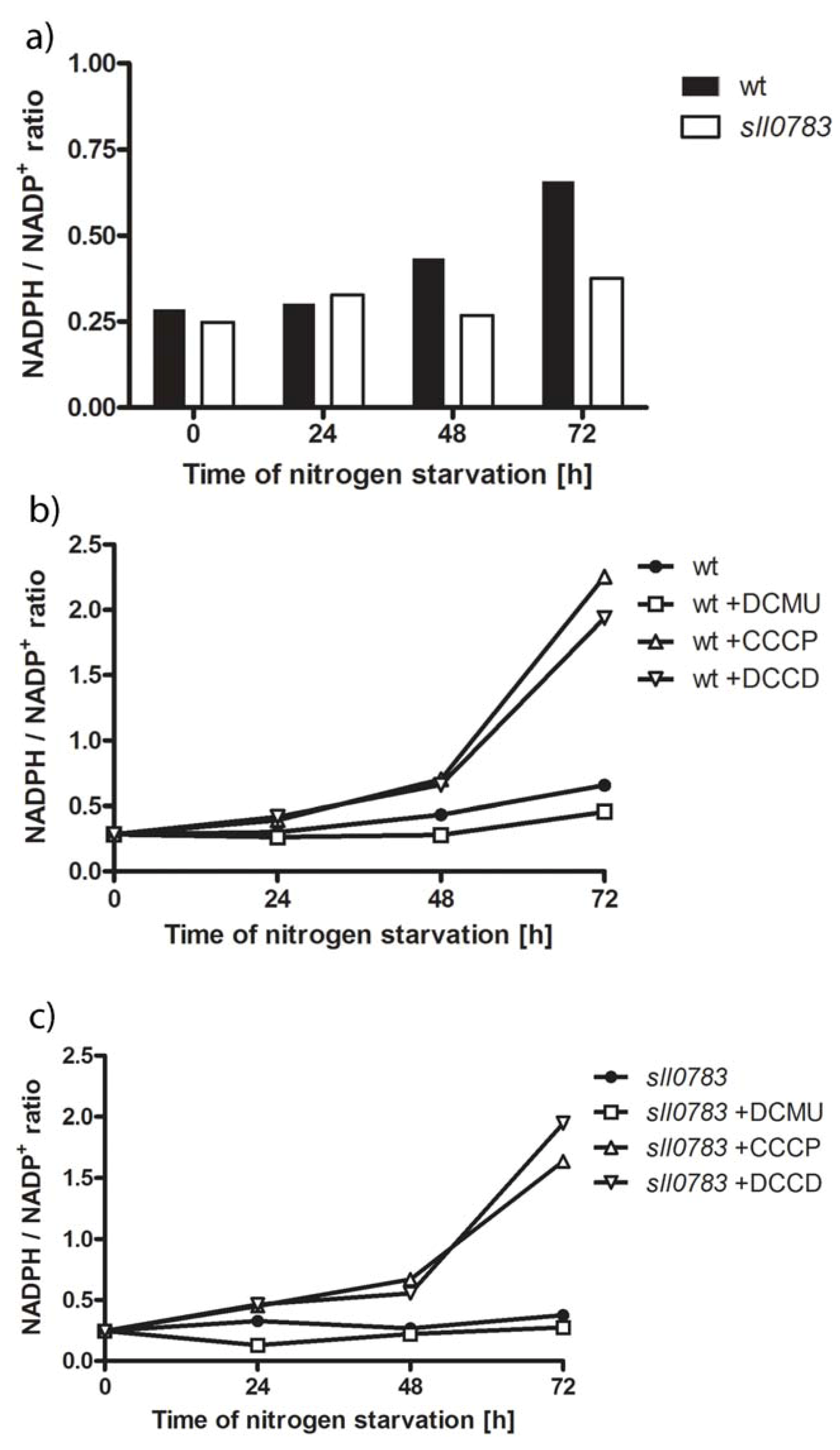
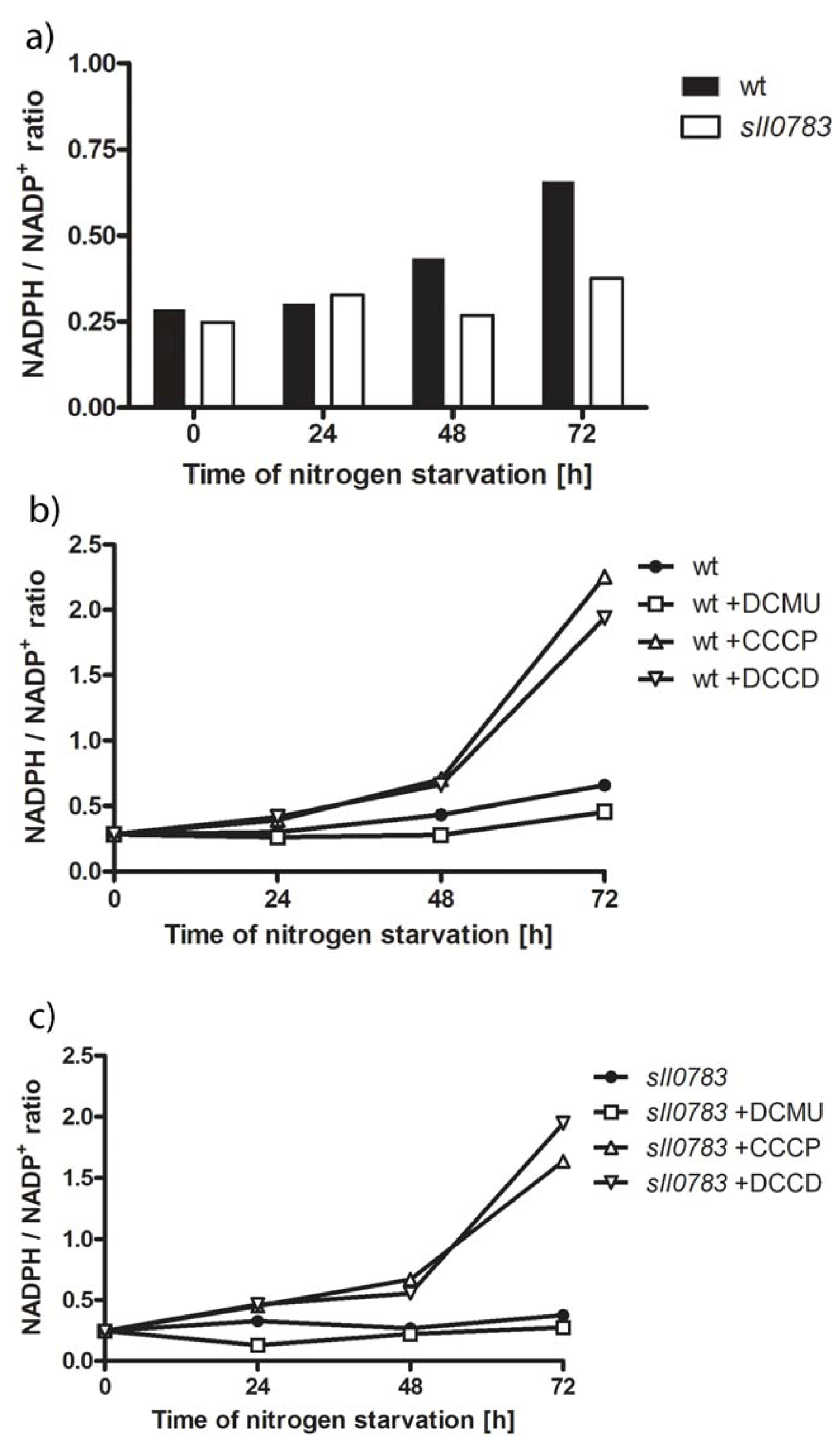
2.3. Nitrogen Starvation Shifts the Intracellular Redox Balance towards a More Reduced State
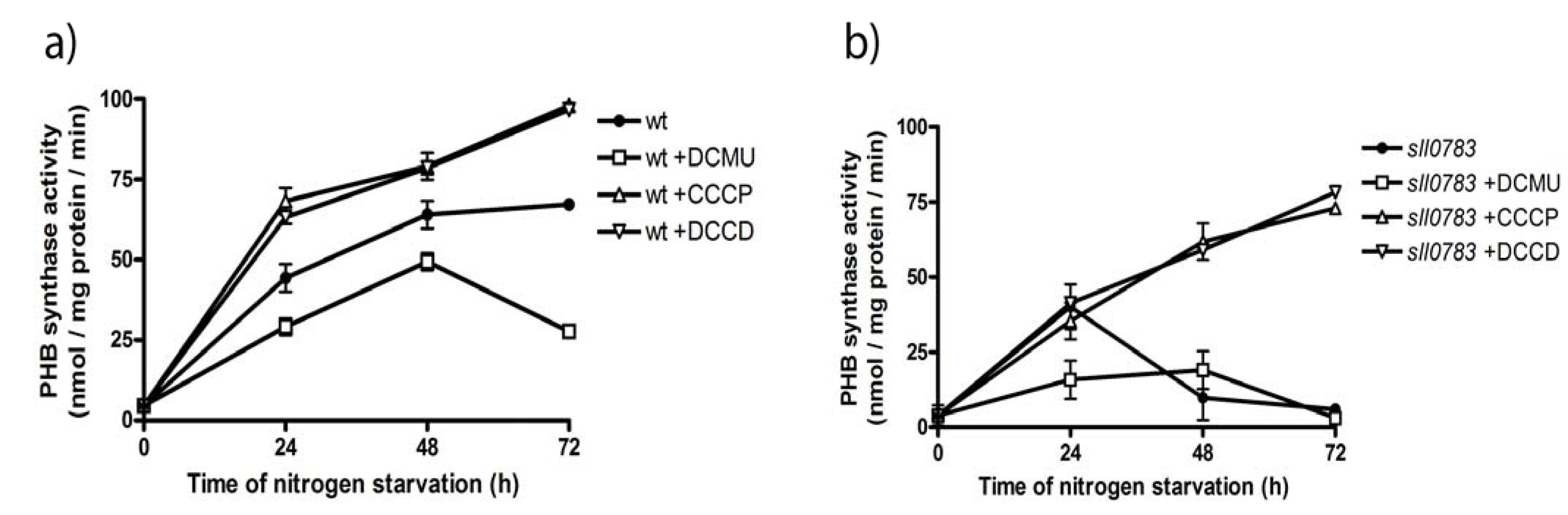
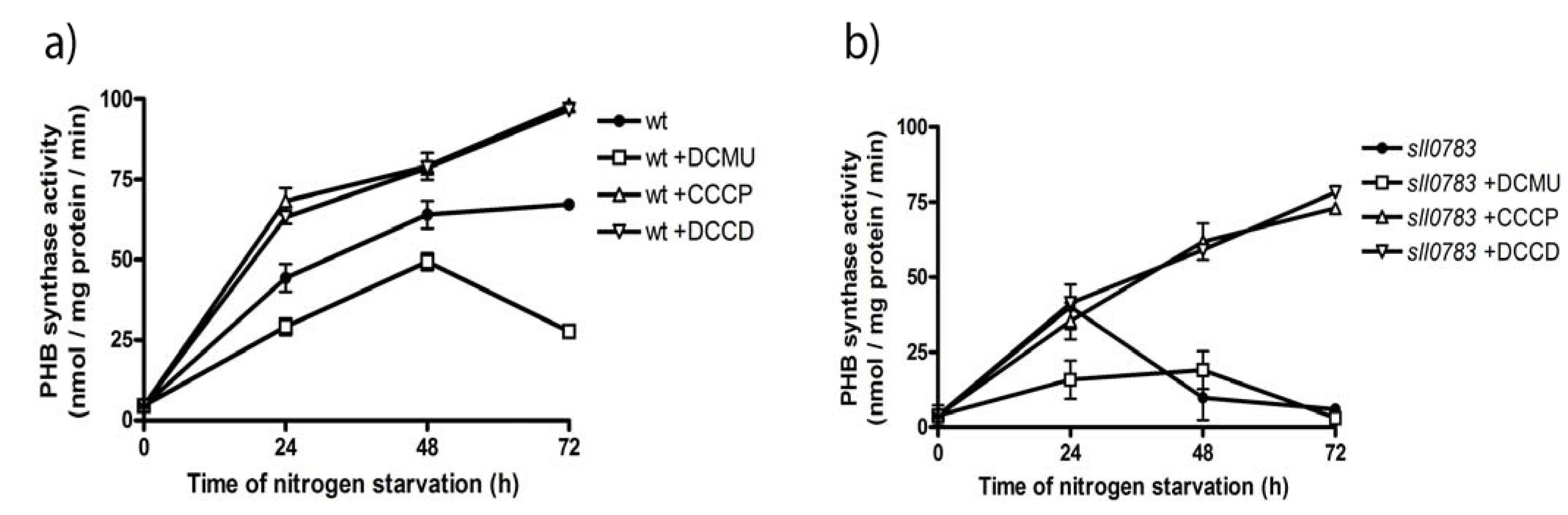
2.4. PHB Synthase in the sll0783 Mutant is Impaired Due to an Altered Redox Balance
3. Experimental Section
3.1. Culture Conditions
3.2. Metabolite Extraction
3.3. Measurements of the NADPH Pool
3.4. PHB Synthase Assay
3.5. Microscopy
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Stanier, R.Y.; Cohen-Bazire, G. Phototrophic prokaryotes: the cyanobacteria. Annu. Rev. Microbiol. 1977, 31, 225–274. [Google Scholar] [CrossRef]
- Harder, W.; Dijkhuizen, L. Physiological responses to nutrient limitation. Annu. Rev. Microbiol. 1983, 37, 1–23. [Google Scholar] [CrossRef]
- Gorl, M.; Sauer, J.; Baier, T.; Forchhammer, K. Nitrogen-starvation-induced chlorosis in Synechococcus PCC 7942: Adaptation to long-term survival. Microbiology 1998, 144, 2449–2458. [Google Scholar] [CrossRef]
- Schwarz, R.; Forchhammer, K. Acclimation of unicellular cyanobacteria to macronutrient deficiency: Emergence of a complex network of cellular responses. Microbiology 2005, 151, 2503–2514. [Google Scholar] [CrossRef]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen control in cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar]
- Fadi Aldehni, M.; Sauer, J.; Spielhaupter, C.; Schmid, R.; Forchhammer, K. Signal transduction protein P(II) is required for NtcA-regulated gene expression during nitrogen deprivation in the cyanobacterium Synechococcus elongatus strain PCC 7942. J. Bacteriol. 2003, 185, 2582–2591. [Google Scholar] [CrossRef]
- Llacer, J.L.; Espinosa, J.; Castells, M.A.; Contreras, A.; Forchhammer, K.; Rubio, V. Structural basis for the regulation of NtcA-dependent transcription by proteins PipX and PII. Proc. Natl. Acad. Sci. USA 2010, 107, 15397–15402. [Google Scholar]
- Panda, B.; Jain, P.; Sharma, L.; Mallick, N. Optimization of cultural and nutritional conditions for accumulation of poly-beta-hydroxybutyrate in Synechocystis sp. PCC 6803. Bioresource Technol. 2006, 97, 1296–1301. [Google Scholar] [CrossRef]
- Taroncher-Oldenburg, G.; Nishina, K.; Stephanopoulos, G. Identification and analysis of the polyhydroxyalkanoate-specific beta-ketothiolase and acetoacetyl coenzyme A reductase genes in the cyanobacterium Synechocystis sp. strain PCC6803. Appl. Environ. Microbiol. 2000, 66, 4440–4448. [Google Scholar] [CrossRef]
- Hein, S.; Tran, H.; Steinbuchel, A. Synechocystis sp. PCC6803 possesses a two-component polyhydroxyalkanoic acid synthase similar to that of anoxygenic purple sulfur bacteria. Arch. Microbiol. 1998, 170, 162–170. [Google Scholar] [CrossRef]
- Schlebusch, M.; Forchhammer, K. Requirement of the nitrogen starvation-induced protein Sll0783 for polyhydroxybutyrate accumulation in Synechocystis sp. strain PCC 6803. Appl. Environ. Microbiol. 2010, 76, 6101–6107. [Google Scholar] [CrossRef]
- Miyake, M.; Kataoka, K.; Shirai, M.; Asada, Y. Control of poly-beta-hydroxybutyrate synthase mediated by acetyl phosphate in cyanobacteria. J. Bacteriol. 1997, 179, 5009–5013. [Google Scholar]
- Cho, M.; Brigham, C.J.; Sinskey, A.J.; Stubbe, J. Purification of polyhydroxybutyrate synthase from its native organism, Ralstonia eutropha: implications for the initiation and elongation of polymer formation in vivo. Biochemistry 2012, 51, 2276–2288. [Google Scholar] [CrossRef]
- Jendrossek, D. Polyhydroxyalkanoate granules are complex subcellular organelles (carbonosomes). J. Bacteriol. 2009, 191, 3195–3202. [Google Scholar]
- Krasikov, V.; Aguirre von Wobeser, E.; Dekker, H.L.; Huisman, J.; Matthijs, H.C. Time-series resolution of gradual nitrogen starvation and its impact on photosynthesis in the cyanobacterium Synechocystis PCC 6803. Physiol. Plantarum 2012, 145, 426–439. [Google Scholar] [CrossRef]
- Muro-Pastor, A.M.; Olmedo-Verd, E.; Flores, E. All4312, an NtcA-regulated two-component response regulator in Anabaena sp. strain PCC 7120. FEMS Microbiol. Lett. 2006, 256, 171–177. [Google Scholar] [CrossRef]
- Zinchenko, V.V.; Piven, I.V.; Melnik, V.A.; Shestakov, S.V. Vectors for the complementation analysis of cyanobacterial mutants. Russ. J. Genet. 1999, 35, 228–232. [Google Scholar]
- Wolk, C.P.; Vonshak, A.; Kehoe, P.; Elhai, J. Construction of shuttle vectors capable of conjugative transfer from Escherichia coli to nitrogen-fixing filamentous cyanobacteria. Proc. Natl. Acad. Sci. USA 1984, 81, 1561–1565. [Google Scholar] [CrossRef]
- Eisenhut, M.; Huege, J.; Schwarz, D.; Bauwe, H.; Kopka, J.; Hagemann, M. Metabolome phenotyping of inorganic carbon limitation in cells of the wild type and photorespiratory mutants of the cyanobacterium Synechocystis sp. strain PCC 6803. Plant. Physiol. 2008, 148, 2109–2120. [Google Scholar] [CrossRef]
- Zhang, S.; Bryant, D.A. The tricarboxylic acid cycle in cyanobacteria. Science 2011, 334, 1551–1553. [Google Scholar] [CrossRef]
- Wolk, C.P.; Thomas, J.; Shaffer, P.W.; Austin, S.M.; Galonsky, A. Pathway of nitrogen metabolism after fixation of 13N-labeled nitrogen gas by the cyanobacterium, Anabaena cylindrica. J. Biol. Chem. 1976, 251, 5027–5034. [Google Scholar]
- Collier, J.L.; Grossman, A.R. A small polypeptide triggers complete degradation of light-harvesting phycobiliproteins in nutrient-deprived cyanobacteria. EMBO J. 1994, 13, 1039–1047. [Google Scholar]
- Sauer, J.; Gorl, M.; Forchhammer, K. Nitrogen starvation in synechococcus PCC 7942: involvement of glutamine synthetase and NtcA in phycobiliprotein degradation and survival. Arch. Microbiol. 1999, 172, 247–255. [Google Scholar] [CrossRef]
- Senior, P.J.; Dawes, E.A. Poly-β-hydroxybutyrate biosynthesis and the regulation of glucose metabolism in Azotobacter beijerinckii. Biochem. J. 1971, 125, 55–66. [Google Scholar]
- England, P.J.; Denton, R.M.; Randle, P.J. The influence of magnesium ions and other bivalent metal ions on the aconitase equilibrium and its bearing on the binding of magnesium ions by citrate in rat heart. Biochem. J. 1967, 105, 32C–33C. [Google Scholar]
- Huege, J.; Goetze, J.; Schwarz, D.; Bauwe, H.; Hagemann, M.; Kopka, J. Modulation of the major paths of carbon in photorespiratory mutants of synechocystis. PloS One 2011, 6, e16278. [Google Scholar]
- Grundel, M.; Scheunemann, R.; Lockau, W.; Zilliges, Y. Impaired glycogen synthesis causes metabolic overflow reactions and affects stress responses in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 2012, 158, 3032–3043. [Google Scholar] [CrossRef]
- Koksharova, O.; Schubert, M.; Shestakov, S.; Cerff, R. Genetic and biochemical evidence for distinct key functions of two highly divergent GAPDH genes in catabolic and anabolic carbon flow of the cyanobacterium Synechocystis sp. PCC 6803. Plant. Mol. Biol. 1998, 36, 183–194. [Google Scholar] [CrossRef]
- Metz, J.G.; Pakrasi, H.B.; Seibert, M.; Arntzen, C.J. Evidence for a Dual Function of the Herbicide-Binding D1-Protein in Photosystem-Ii. FEBS Lett. 1986, 205, 269–274. [Google Scholar] [CrossRef]
- Zürrer, H.; Snozzi, M.; Bachofen, R. Specific binding of DCCD to reaction centers of the photosynthetic bacterium Rhodospirillum rubrum and its effect of certain photosynthetic reactions. FEBS Lett. 1983, 153, 151–155. [Google Scholar] [CrossRef]
- Sauer, J.; Schreiber, U.; Schmid, R.; Volker, U.; Forchhammer, K. Nitrogen starvation-induced chlorosis in Synechococcus PCC 7942. Low-level photosynthesis as a mechanism of long-term survival. Plant. Physiol. 2001, 126, 233–243. [Google Scholar] [CrossRef]
- Mallick, N.; Sharma, L.; Singh, A.K. Poly-beta-hydroxybutyrate accumulation in Nostoc muscorum: Effects of metabolic inhibitors. J. Plant. Physiol. 2007, 164, 312–317. [Google Scholar] [CrossRef]
- Dephilippis, R.; Ena, A.; Guastini, M.; Sili, C.; Vincenzini, M. Factors Affecting Poly-Beta-Hydroxybutyrate Accumulation in Cyanobacteria and in Purple Nonsulfur Bacteria. FEMS Microbiol. Rev. 1992, 9, 187–194. [Google Scholar]
- Rippka, R. Isolation and Purification of Cyanobacteria. Method Enzymol. 1988, 167, 3–27. [Google Scholar]
- Valentin, H.; Steinbüchel, A. Application of enzymatically synthesized short-chain-length hydroxy fatty acid coenzyme A thioesters for assay of polyhydroxyalkanoic acid synthases. Appl. Microbiol. Biotechnol. 1994, 40, 699–709. [Google Scholar] [CrossRef]
- Marin, K.; Stirnberg, M.; Eisenhut, M.; Kramer, R.; Hagemann, M. Osmotic stress in Synechocystis sp. PCC 6803: Low tolerance towards nonionic osmotic stress results from lacking activation of glucosylglycerol accumulation. Microbiology 2006, 152, 2023–2030. [Google Scholar] [CrossRef]
- Estepa, J.; Luque-Almagro, V.M.; Manso, I.; Escribano, M.P.; Martinez-Luque, M.; Castillo, F.; Moreno-Vivian, C.; Roldan, M.D. The nit1C gene cluster of Pseudomonas pseudoalcaligenes CECT5344 involved in assimilation of nitriles is essential for growth on cyanide. Env. Microbiol. Rep. 2012, 4, 326–334. [Google Scholar] [CrossRef]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hauf, W.; Schlebusch, M.; Hüge, J.; Kopka, J.; Hagemann, M.; Forchhammer, K. Metabolic Changes in Synechocystis PCC6803 upon Nitrogen-Starvation: Excess NADPH Sustains Polyhydroxybutyrate Accumulation. Metabolites 2013, 3, 101-118. https://doi.org/10.3390/metabo3010101
Hauf W, Schlebusch M, Hüge J, Kopka J, Hagemann M, Forchhammer K. Metabolic Changes in Synechocystis PCC6803 upon Nitrogen-Starvation: Excess NADPH Sustains Polyhydroxybutyrate Accumulation. Metabolites. 2013; 3(1):101-118. https://doi.org/10.3390/metabo3010101
Chicago/Turabian StyleHauf, Waldemar, Maximilian Schlebusch, Jan Hüge, Joachim Kopka, Martin Hagemann, and Karl Forchhammer. 2013. "Metabolic Changes in Synechocystis PCC6803 upon Nitrogen-Starvation: Excess NADPH Sustains Polyhydroxybutyrate Accumulation" Metabolites 3, no. 1: 101-118. https://doi.org/10.3390/metabo3010101