Performance and Microbial Community Structure of Anaerobic Membrane Bioreactor for Lipids-Rich Kitchen Waste Slurry Treatment: Mesophilic and Thermophilic Processes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Kitchen Waste Slurry Characterization
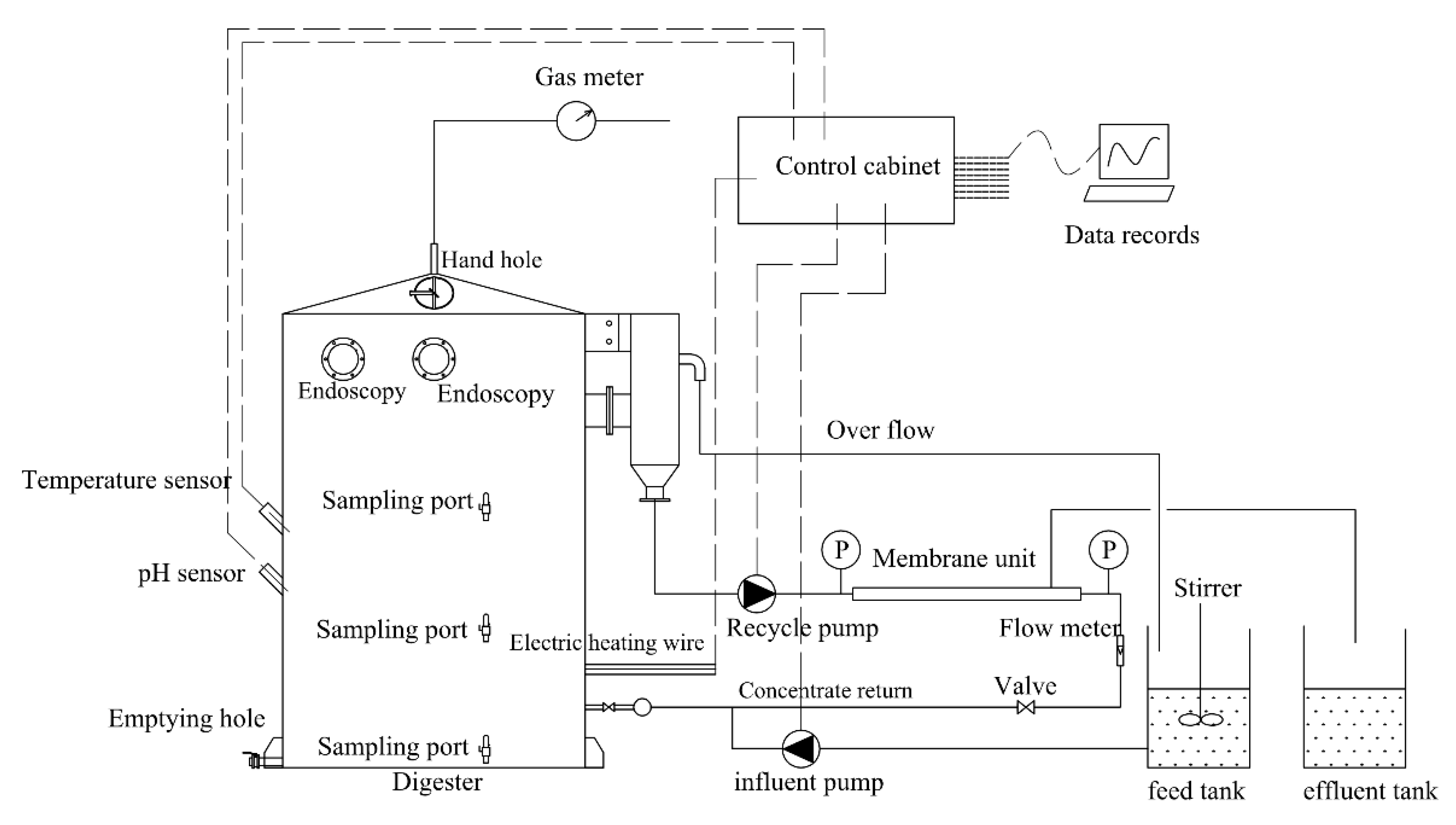
2.2. AnMBR Configuration and Parameters
2.3. Operation Strategy
2.4. Physicochemical Analysis
2.5. Microbial Community Analysis
3. Results and Discussion
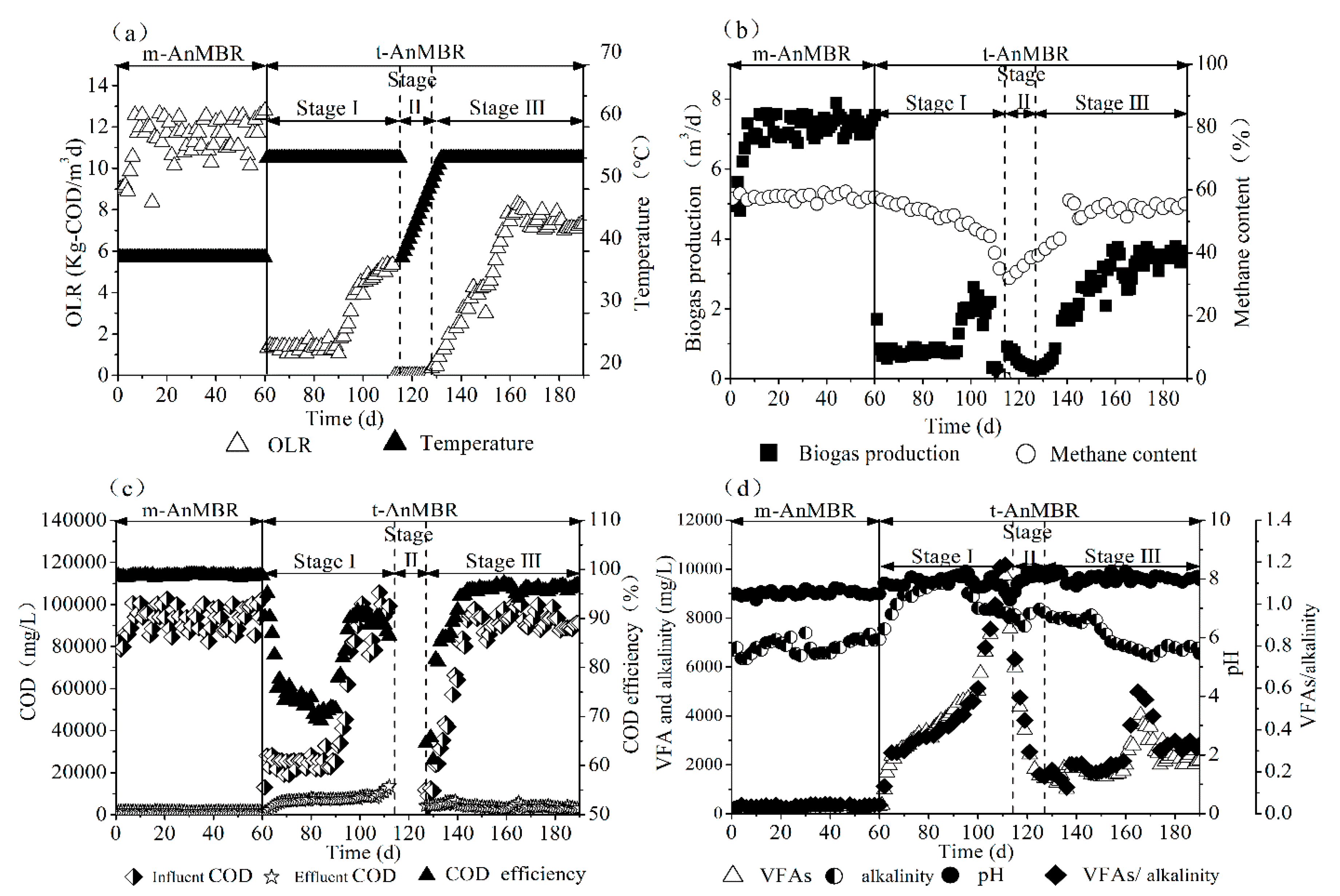
3.1. Digestion Performance
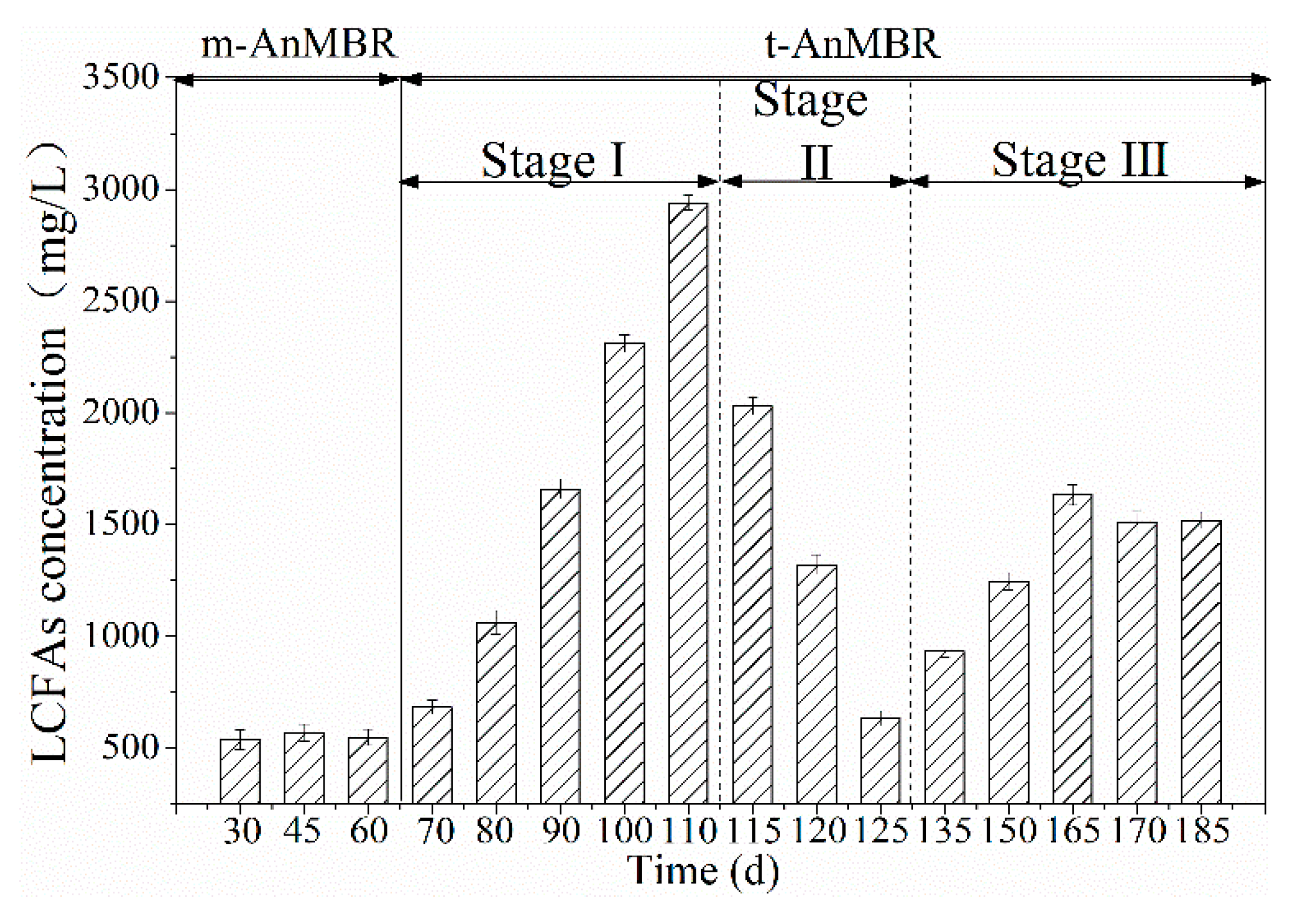
3.2. LCFAs Accumulation
3.3. Microbial Community Structure
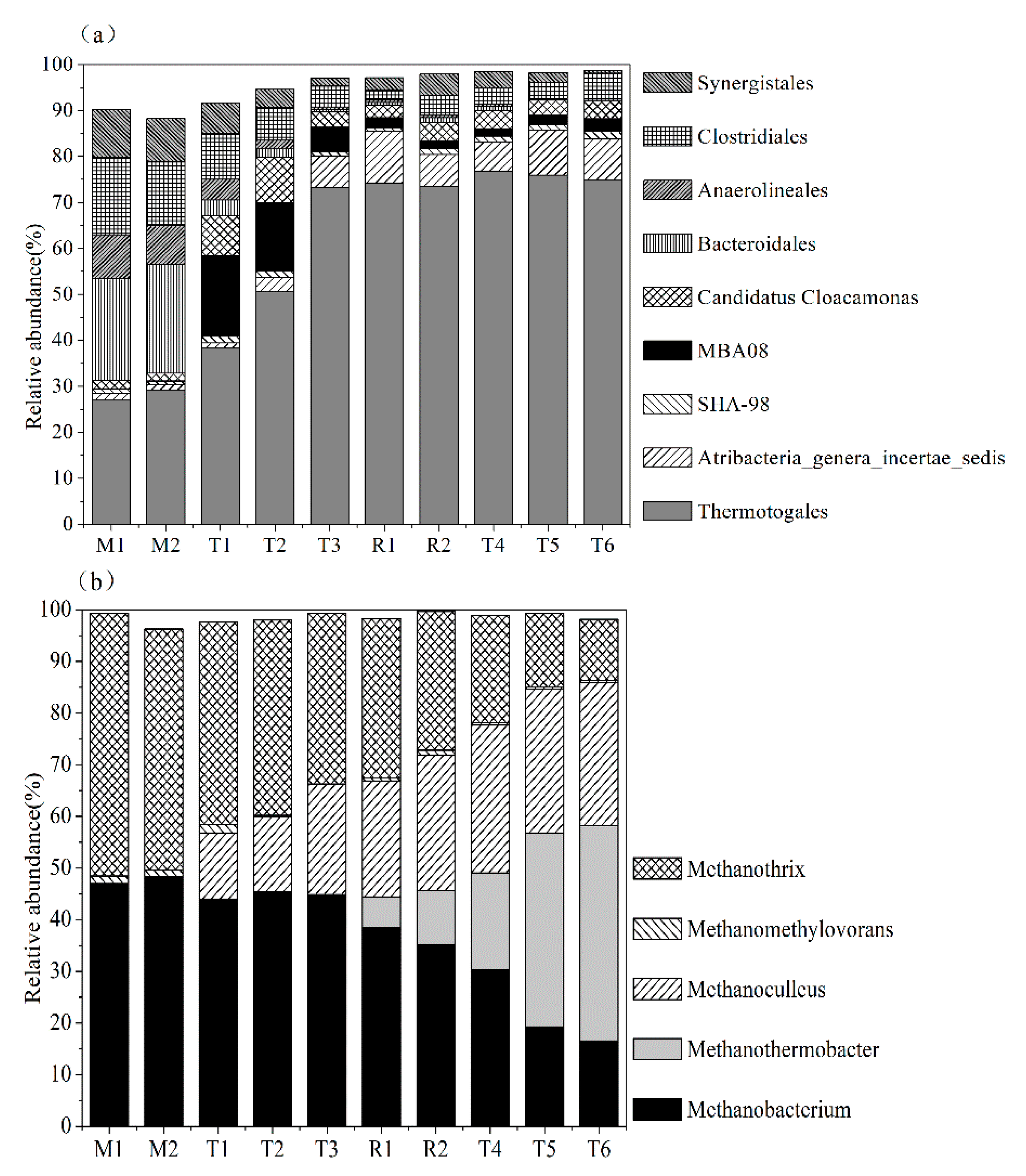
3.3.1. Microbial Diversity
3.3.2. Microbial Relative Abundance
3.4. Membrane Filtration Performance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yin, C.H.; Dong, X.; Lv, L.; Wang, Z.G.; Xu, Q.Q.; Liu, X.L.; Yan, H. Economic production of probiotics from kitchen waste. Food Sci. Biotechenol. 2013, 22, 59–63. [Google Scholar] [CrossRef]
- Li, Y.Y.; Sasaki, H.; Yamashita, K.; Saki, K.; Kamigochi, I. High-rate methane fermentation of lipid-rich food wastes by a high-solids co-digestion process. Water Sci. Technol. 2002, 45, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Cavaleiro, A.J.; Mota, M.; Mota, M.; Alves, M.M. Accumulation of long chain fatty acids onto anaerobic sludge under steady state and shock loading conditions: Effect on acetogenic and methanogenic activity. Water Sci. Technol. 2003, 48, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Hanaki, K.; Matsuo, T.; Nagase, M. Mechanism of inhibition caused by long chain fatty acids in anaerobic digestion process. Biotechnol. Bioeng. 1981, 23, 1590–1610. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, L.; Chi, L.; Long, X.; Mei, Z.; Zhang, Z. Startup and operation of anaerobic EGSB reactor treating palm oil mill effluent. J. Environ. Sci. 2008, 20, 658–663. [Google Scholar] [CrossRef]
- Dereli, R.K.; Ersahin, M.E.; Ozgun, H.; Ozturk, I.; Jeison, D.; van der Zee, F.; van Lier, J.B. Potentials of anaerobic membrane bioreactors to overcome treatment limitations induced by industrial wastewaters. Bioresour. Technol. 2012, 122, 160–170. [Google Scholar] [CrossRef]
- Xiao, X.; Huang, Z.; Ruan, W.; Yan, L.; Miao, H.; Ren, H.; Zhao, M. Evaluation and characterization during the anaerobic digestion of high-strength kitchen waste slurry via a pilot-scale anaerobic membrane bioreactor. Bioresour. Technol. 2015, 193, 234–242. [Google Scholar] [CrossRef]
- Zábranská, J.; Stepova, J.; Wachtl, R.; Jenicek, P.; Dohanyos, M. The activity of anaerobic biomass in thermophilic and mesophilic digesters at differentloading rates. Water Sci. Technol. 2000, 32, 49–56. [Google Scholar] [CrossRef]
- Kim, M.; Ahn, Y.H.; Speece, R.E. Comparative process stability and efficiency of anaerobic digestion; mesophilic vs. Thermophilic. Water Res. 2002, 36, 4369–4385. [Google Scholar] [CrossRef]
- Khanal, S.K.; Surampalli, R.Y.; Zhang, T.C.; Lamsal, B.P.; Tyagi, R.D.; Kao, C.M. Bioenergy and biofuel from biowastes and biomass. Biomass Fuels 2010, 44. [Google Scholar] [CrossRef]
- Labatut, R.A.; Angenent, L.T.; Scott, N.R. Conventional mesophilic vs. thermophilic anaerobic digestion: A trade-off between performance and stability. Water Res. 2014, 53, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Bayr, S.; Rantanen, M.; Kaparaju, P.; Rintala, J. Mesophilic and thermophilic anaerobic co-digestion of rendering plant and slaughterhouse wastes. Bioresour. Technol. 2012, 104, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Broughton, M.J. Anaerobic batch digestion of sheep tallow. Water Res. 1998, 32, 1423–1428. [Google Scholar] [CrossRef]
- Xiao, X.; Shi, W.; Huang, Z.; Ruan, W.; Miao, H.; Ren, H.; Zhao, M. Process stability and microbial response of anaerobic membrane bioreactor treating high-strength kitchen waste slurry under different organic loading rates. Int. Biodeterior. Biodegrad. 2017, 121, 35–43. [Google Scholar] [CrossRef]
- APHA; AWWA; WEF. Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Huang, Z.; Yu, X.; Miao, H.; Ren, H.; Zhao, M.; Ruan, W. Enzymatic dynamics of microbial acid tolerance response (ATR) during the enhanced biohydrogen production process via anaerobic digestion. Int. J. Hydrog. Energy 2012, 37, 10655–10662. [Google Scholar] [CrossRef]
- Yang, L.; Huang, Y.; Zhao, M.; Huang, Z.; Miao, H.; Xu, Z.; Ruan, W. Enhancing biogas generation performance from food wastes by high-solids thermophilic anaerobic digestion: Effect of pH adjustment. Int. Biodeterior. Biodegrad. 2015, 105, 153–159. [Google Scholar] [CrossRef]
- Thiex, N.J.; Anderson, S.; Gildemeister, B. Crude fat, diethyl ether extraction, in feed, cereal grain, and forage (Randall/Soxtec/submersion method): Collaborative study. J. AOAC Int. 2003, 86, 888–898. [Google Scholar]
- Neves, L.; Pereira, M.A.; Mota, M.; Alves, M.M. Detection and quantification of long chain fatty acids in liquid and solid samples and its relevance to understand anaerobic digestion of lipids. Bioresour. Technol. 2009, 100, 91–96. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 2049–2618. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Fang, Z. Research Advances in the Inhibition of Long Chain Fatty Acid to Methanogenic Activity in Anaerobic Digestion System. J. Anhui Agric. Sci. 2012, 13, 2389–2393. [Google Scholar]
- Lin, H.J.; Xie, K.; Mahendran, B.; Bagley, D.M.; Leung, K.T.; Liss, S.N.; Liao, B.Q. Sludge properties and their effects on membrane fouling in submerged anaerobic membrane bioreactors (SAnMBRs). Water Res. 2009, 43, 3827–3837. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, F.J.; Wase, D.A.J.; Thayanithy, K.; Forster, C.F. Continuous co-digestion of cattle slurry with fruit and vegetable wastes and chicken manure. Biomass Bioenergy 2002, 27, 71–77. [Google Scholar] [CrossRef]
- Palatsi, J.; Laureni, M.; Andrés, M.V.; Flotats, X.; Nielsen, H.B.; Angelidaki, I. Strategies for recovering inhibition caused by long-chain fatty acids on anaerobic thermophilic biogas reactors. Bioresour. Technol. 2009, 100, 4588–4596. [Google Scholar] [CrossRef]
- Silvestre, G.; Illa, J.; Fernández, B.; Bonmatí, A. Thermophilic anaerobic co-digestion of sewage sludge with grease waste: Effect of long chain fatty acids in the methane yield and its dewatering properties. Appl. Energy 2014, 117, 87–94. [Google Scholar] [CrossRef]
- Angelidaki, I.; Sanders, W. Assessment of the anaerobic biodegradability of macropollutants. Rev. Environ. Sci. Bio/Technol. 2004, 3, 117–129. [Google Scholar] [CrossRef]
- Kim, S.; Han, S.; Shin, H. Kinetics of LCFA Inhibition on Acetoclastic Methanogenesis, Propionate Degradation and β-Oxidation. J. Environ. Sci. Health Part A 2004, 39, 1025. [Google Scholar] [CrossRef]
- Neves, L.; Oliveira, R.; Alves, M.M. Fate of LCFA in the co-digestion of cow manure, food waste and discontinuous addition of oil. Water Res. 2009, 43, 5142–5150. [Google Scholar] [CrossRef]
- Speece, R.E.; Boonyakitsombut, S.; Kim, M.; Azbar, N.; Ursillo, P. Overview of anaerobic treatment: Thermophilic and propionate implications. Water Sci. Technol. 2006, 78, 460–473. [Google Scholar]
- Wilson, C.A.; Murthy, S.M.; Fang, Y. The effect of temperature on the performance and stability of thermophilic anaerobic digestion. Water Sci. Technol. 2008, 57, 297. [Google Scholar] [CrossRef] [PubMed]
- Kabouris, J.C.; Tezel, U.; Pavlostathis, S.G.; Engelmann, M.; Dulaney, J.; Gillette, R.A. Methane recovery from the anaerobic codigestion of municipal sludge and FOG. Bioresour. Technol. 2009, 100, 3701–3705. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.T.; Kraus, D.L. Degradation of long chain fatty acids by activated sludge. Water Res. 1973, 7, 843–851. [Google Scholar] [CrossRef]
- Pereira, M.A.; Pires, O.C.; Mota, M.; Alves, M.M. Anaerobic biodegradation of oleic and palmitic acids: Evidence of mass transfer limitations caused by long chain fatty acid accumulation onto the anaerobic sludge. Biotechnol. Bioeng. 2005, 92, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.M.; Zhao, M.X.; Chen, Y.; Yu, M.J.; Ruan, W.Q. Tolerance response to in situ ammonia stress in a pilot-scale anaerobic digestion reactor for alleviating ammonia inhibition. Bioresour. Technol. 2015, 198, 372–379. [Google Scholar] [CrossRef]
- Gannoun, H.; Khelifi, E.; Omri, I.; Jabari, L.; Fardeau, M.L.; Bouallagui, H.; Godon, J.J.; Hamdia, M. Microbial monitoring by molecular tools of an upflow anaerobic filter treating abattoir wastewaters. Bioresour. Technol. 2013, 142, 269–277. [Google Scholar] [CrossRef]
- Deng, Y.; Huang, Z.; Zhao, M.; Ruan, W.; Ren, H. Effects of co-inoculating rice straw with ruminal microbiota and anaerobic sludge: Digestion performance and spatial distribution of microbial communities. Appl. Environ. Microbiol. 2017, 101, 5937–5948. [Google Scholar] [CrossRef]
- Regueiro, L.; Veiga, P.; Figueroa, M.; Alonso-Gutierrez, J.; Stams, A.J.M.; Lema, J.M.; Carballa, M. Relationship between microbial activity and microbial community structure in six full-scale anaerobic digesters. Microbiol. Res. 2012, 167, 581–589. [Google Scholar] [CrossRef]
- Sundberg, C.; Al-Soud, W.A.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626. [Google Scholar] [CrossRef] [Green Version]
- Town, J.R.; Links, M.G.; Fonstad, T.A.; Dumonceaux, T.J. Molecular characterization of anaerobic digester microbial communities identifies microorganisms that correlate to reactor performance. Bioresour. Technol. 2014, 151, 249–257. [Google Scholar] [CrossRef]
- Wang, T.X.; Ma, X.Y.; Wang, M.M.; Chu, H.J.; Zuo, J.E.; Yang, Y.F. A comparative study of microbial community compositions in thermophilic and mesophilic sludge anaerobic digestion systems. Microbiol. China 2016, 43, 26–35. [Google Scholar]
- Huber, R.; Hannig, M. Thermotogales. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; pp. 899–922. [Google Scholar]
- Nesb, C.L.; Kumaraswamy, R.; Dlutek, M.; Doolittle, W.F.; Foght, J. Searching for mesophilic Thermotogales bacteria: “mesotogas” in the wild. Appl. Environ. Microbiol. 2010, 76, 4896–4900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meabe, E.; Déléris, S.; Soroa, S.; Sancho, L. Performance of anaerobic membrane bioreactor for sewage sludge treatment: Mesophilic and thermophilic processes. J. Membr. Sci. 2013, 446, 26–33. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Value |
|---|---|---|
| Total Chemical Oxygen Demand | (g/L) | 90.2 ± 6.9 |
| Total Suspended Solids | (g/L) | 18.5 ± 3.2 |
| Total Volatile Suspended Solids | (g/L) | 16.2 ± 2.6 |
| lipids | 5.95 ± 0.28 | |
| pH | --- | 3.88 ± 0.32 |
| Ammonia Nitrogen | (mg/L) | 325 ± 45 |
| Total Kjeldahl Nitrogen | (mg/L) | 1848 ± 205 |
| Total Phosphorus | (mg/L) | 83.5 ± 5.4 |
| Conductivity | (ms/cm) | 10.64 ± 2.58 |
| LCFA | m-AnMBR | t-AnMBR |
|---|---|---|
| mg/L | mg/L | |
| Lauric (C12:0) | 12.5 ± 0.5 | 17.7 ± 0.3 |
| Myristic (C14:0) | 19.6 ± 1.2 | 38.3 ± 3.4 |
| Palmitic (C16:0) | 179.4 ± 10.6 | 417.5 ± 18.8 |
| Palmitoleic (C16:1) | 9.7 ± 0.9 | 28.4 ± 2.5 |
| Stearic (C18:0) | 56.5 ± 5.3 | 162.3 ± 15.8 |
| Oleic (C18:1) | 163.2 ± 18.1 | 535.7 ± 23.6 |
| Linoleic (C18:2) | 38.6 ± 1.3 | 98.6 ± 5.3 |
| Linolenic (C18:3) | 11.7 ± 0.7 | 24.6 ± 2.7 |
| Arachidic (C20:0) | 8.3 ± 0.4 | 27.2 ± 1.9 |
| Arachidonic (C20:1) | 15.3 ± 1.1 | 32.9 ± 2.2 |
| Behenic (C22:0) | 10.6 ± 0.9 | 28.4 ± 1.8 |
| Docosahexaenoic (C22:5) | 9.4 ± 0.5 | 16.6 ± 0.8 |
| Others | 15.2 ± 1.8 | 72.4 ± 6.8 |
| Total | 550 | 1500 |
| Sampling Point | Shannon | Sampling Point | Shannon | ||
|---|---|---|---|---|---|
| Bacteria | M1 | 13.13 | Archaea | M1 | 8.73 |
| M2 | 13.65 | M2 | 8.31 | ||
| T1 | 12.52 | T1 | 7.28 | ||
| T2 | 11.93 | T2 | 6.96 | ||
| T3 | 10.62 | T3 | 5.57 | ||
| R1 | 10.64 | R1 | 5.68 | ||
| R2 | 10.87 | R2 | 6.28 | ||
| T4 | 11.32 | T4 | 6.68 | ||
| T5 | 11.87 | T5 | 6.93 | ||
| T6 | 12. 02 | T6 | 7.20 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, X.; Shi, W.; Ruan, W. Performance and Microbial Community Structure of Anaerobic Membrane Bioreactor for Lipids-Rich Kitchen Waste Slurry Treatment: Mesophilic and Thermophilic Processes. Processes 2019, 7, 879. https://doi.org/10.3390/pr7120879
Xiao X, Shi W, Ruan W. Performance and Microbial Community Structure of Anaerobic Membrane Bioreactor for Lipids-Rich Kitchen Waste Slurry Treatment: Mesophilic and Thermophilic Processes. Processes. 2019; 7(12):879. https://doi.org/10.3390/pr7120879
Chicago/Turabian StyleXiao, Xiaolan, Wansheng Shi, and Wenquan Ruan. 2019. "Performance and Microbial Community Structure of Anaerobic Membrane Bioreactor for Lipids-Rich Kitchen Waste Slurry Treatment: Mesophilic and Thermophilic Processes" Processes 7, no. 12: 879. https://doi.org/10.3390/pr7120879