
Cymbopogon citratus Water Extract and Methyl Jasmonate Improve Polyunsaturated Fatty Acid Metabolism in Taiwanofungus camphoratus Mycelia
Abstract
:1. Introduction
2. Materials and Methods
2.1. LWE Preparation and Monosaccharide Composition Analysis
2.2. Growth of T. camphoratus Mycelia
2.3. LA and OA Contents
2.4. Transcriptome Analysis of T. camphoratus Mycelia
2.5. Quantitative RT-PCR Analysis of Differential Genes
2.6. Bioinformatics Analysis of Full−Length Genes
2.7. Data Analysis
3. Results
3.1. Monosaccharide Composition Analysis of LWE
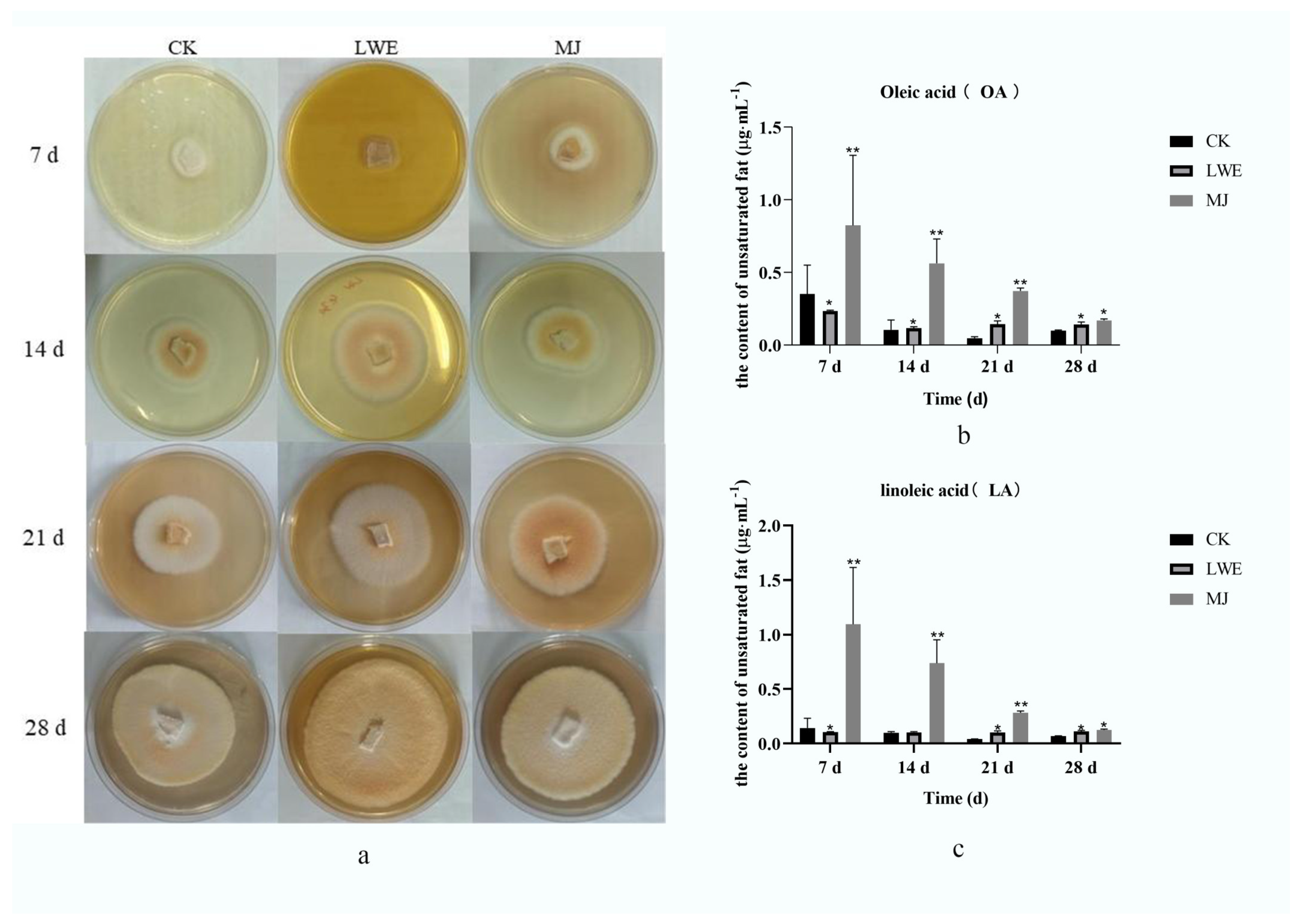
3.2. T. camphoratus Mycelia Growth
3.3. Oleic and Linoleic Acid Determination
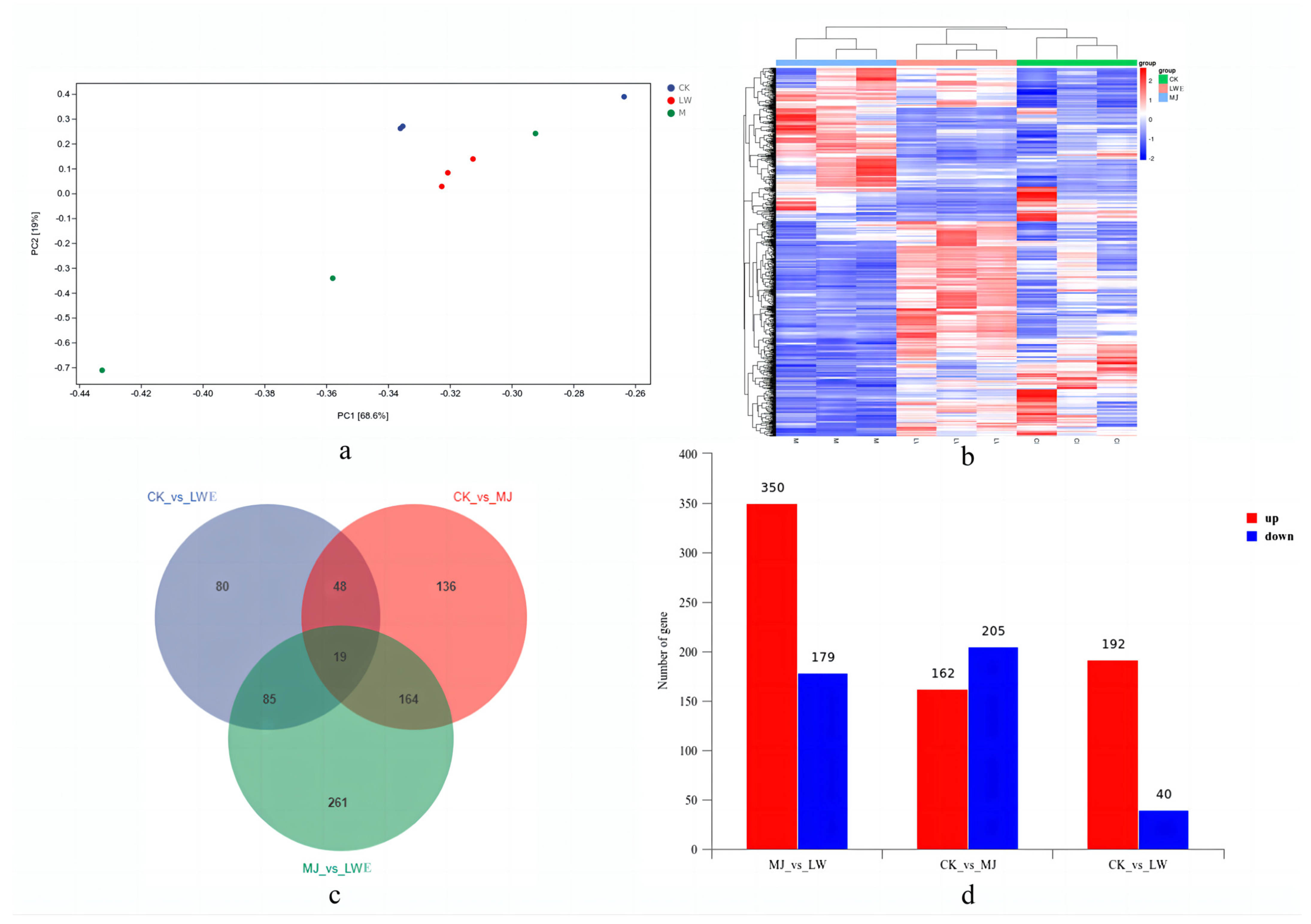
3.4. T. camphoratus Mycelia Transcriptome Sequencing and Analysis
3.5. Functional Annotation of T. camphoratus Mycelia
3.6. Functional Enrichment Annotation of Differentially Expressed Genes
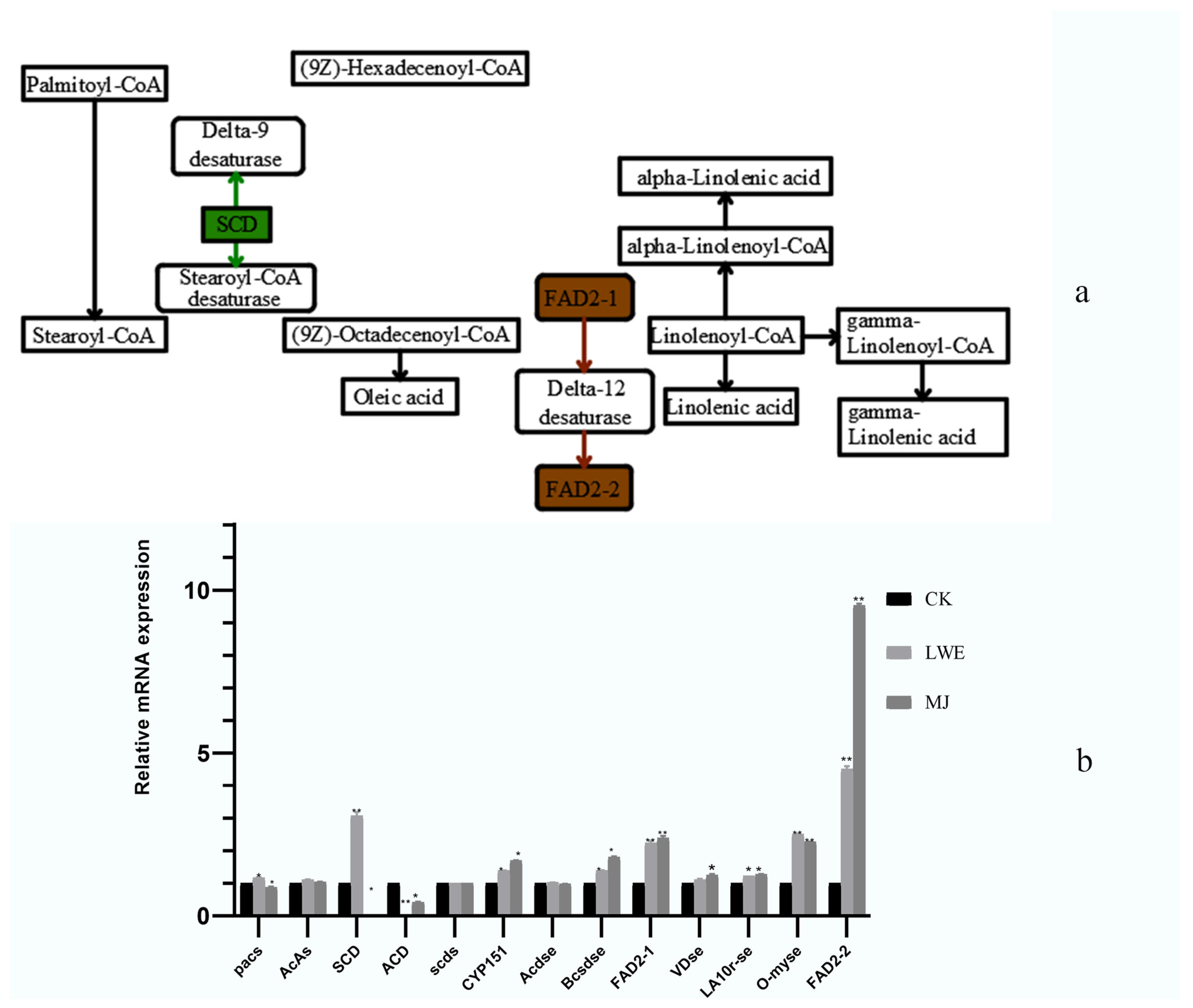
3.7. Quantitative Genetic Analysis
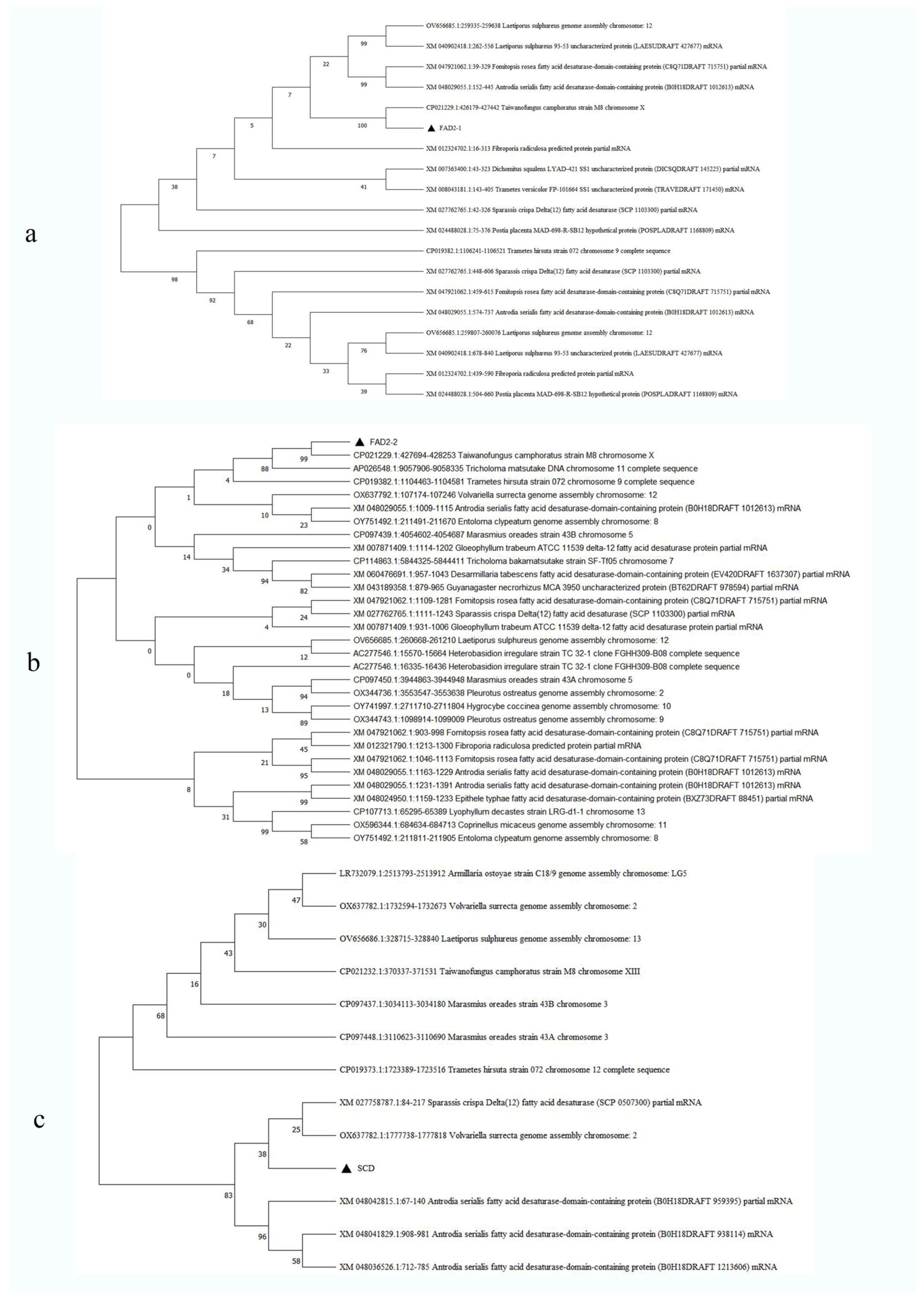
3.8. Bioinformatics Analysis of Three Genes
4. Discussion
4.1. Exogenous Additives in the Artificial Cultivation of T. camphoratus
4.2. PUFAs in Edible Fungi
4.3. Profiles of Genes Associated with Polyunsaturated Fatty Acids
4.4. Study Limitations and Future Research Possibilities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Chen, R.; Li, L.; Dong, R.; Yin, H.; Wang, Y. The triterpenoids-enriched extracts from Antrodia cinnamomea mycelia attenuate alcohol-induced chronic liver injury via suppression lipid accumulation in c57bl/6 mice. Food Sci. Hum. Wellness 2021, 10, 497–507. [Google Scholar] [CrossRef]
- Qiao, X.; Wang, Q.; Ji, S. Metabolites identification and multi−component pharmacokinetics of ergostane and lanostane triterpenoids in the anticancer mushroom Antrodia cinnamomea. J. Pharm. Biomed. Anal. 2015, 111, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.H.; Lu, M.K.; Lo, H.C.; Chang, C.C.; Tseng, A.J.; Chao, C.H.; Lin, T.Y. ZnF3, a sulfated polysaccharide from Antrodia cinnamomea, inhibits lung cancer cells via induction of apoptosis and activation of M1-like macrophage−induced cell death. Int. J. Biol. Macromol. 2023, 238, 124–144. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.K.; Cheng, J.J.; Lai, W.L.; Lin, Y.R.; Huang, N.K. Adenosine as an active component of Antrodia cinnamomea that prevents rat PC12 cells from serum deprivation−induced apoptosis through the activation of adenosine A2A receptors. Life Sci. 2006, 3, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.H.; Lai, K.S.; Huang, Y.Y.; Chen, H.Y.; Xiong, L.Q.; Guo, H.K.; Yang, Q.Q.; Zhang, B.B. Efficient production of Antrodin C by microparticle−enhanced cultivation of medicinal mushroom Antrodia cinnamomea. J. Biosci. Bioeng. 2023, 3, 232–237. [Google Scholar] [CrossRef]
- Lu, M.K.; Chao, C.H.; Chang, T.Y. A branched 2-O sulfated 1,3-/1,4-galactoglucan from Antrodia cinnamomea exhibits moderate antiproliferative and anti−inflammatory activities. Int. J. Biol. Macromol. 2023, 241, 124559. [Google Scholar] [CrossRef]
- Dakpa, G.; Kumar, K.J.S.; Nelen, J. Antcin-B, a phytosterol−like compound from Taiwanofungus camphoratus inhibits SARS-CoV-2 3-chymotrypsin−like protease (3CLPro) activity in silico and in vitro. Sci. Rep. 2023, 13, 17106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lv, P.; Ma, J.; Chen, N.; Guo, H.; Chen, Y.; Gan, X.; Wang, R.; Liu, X.; Fan, S.; et al. Antrodia cinnamomea exerts an anti-hepatoma effect by targeting PI3K/AKT−mediated cell cycle progression in vitro and in vivo. Acta Pharm. Sin. B 2022, 12, 890–906. [Google Scholar] [CrossRef]
- Chang, C.C.; Lu, Y.C.; Wang, C.C.; Ko, T.L.; Chen, J.R.; Wang, W.; Chen, Y.L.; Wang, Y.W.; Chang, T.H.; Hsu, H.F.; et al. . Antrodia cinnamomea extraction waste supplementation promotes thermal stress tolerance and tissue regeneration ability of zebrafish. Molecules 2020, 25, 4213. [Google Scholar] [CrossRef]
- Cheng, J.J.; Chao, C.H.; Lu, M.K. Large-scale preparation of sulfated polysaccharides with anti−angionenic and anti-inflammatory properties from Antrodia cinnamomia. Int. J. Biol. Macromol. 2018, 113, 1198–1205. [Google Scholar] [CrossRef]
- Lu, M.K.; Chao, C.H.; Hsu, Y.C.; Chang, C.C. Structural sequencing and anti−inflammatory, anti-lung cancer activities of 1,4-α/β−sulfomalonoglucan in Antrodia cinnamomea. Int. J. Biol. Macromol. 2021, 170, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.T.; Huang, K.W.; Chen, W.J.; Lai, T.H. A Botanical Drug Extracted from Antrodia cinnamomea: A first−in−human phase i study in healthy volunteers. J. Am. Coll. Nutr. 2023, 42, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, S.; Zhang, Y.; Zhang, W.; Zhong, H.; Lu, X.; Guan, R. A review on the protective effect of active components in Antrodia camphorata against alcoholic liver injury. J. Ethnopharmacol. 2023, 300, 115740. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Gai, S.P.; Li, X.H.; Zhang, J.S.; Wang, W.H. Transcriptional analysis of Antrodin C synthesis in Taiwanofungus camphoratus (Syn. Antrodia camphorate, Antrodia cinnamomea) to understand its biosynthetic mechanism. Fermentation 2024, 10, 28. [Google Scholar] [CrossRef]
- Lin, T.Y.; Tseng, A.J.; Chao, C.H.; Lu, M.K. Microelements induce changes in characterization of sulfated polysaccharides from Antrodia cinnamomea. Int. J. Biol. Macromol. 2018, 120, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Lu, M.K.; Tseng, A.J.; Chao, C.H. Effects of sterol−type elicitors on biochemical characterization of polysaccharides from Antrodia cinnamomea. Int. J. Biol. Macromol. 2020, 162, 1476–1483. [Google Scholar] [CrossRef]
- Schisler, L.C. Stimulation of yield in the cultivated mushroom by vegetable oils. Appl. Microbiol. Biotechnol. 1967, 15, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, B.; Balázs, V.L.; Molnár, S.; Szögi−Tatár, B.; Böszörményi, A.; Palkovics, T.; Horváth, G.; Schneider, G. Antibacterial effect of lemongrass (Cymbopogon citratus) against the aetiological agents of pitted keratolyis. Molecules 2022, 27, 1423. [Google Scholar] [CrossRef] [PubMed]
- Lorenzetti, B.B.; Souza, G.E.; Sarti, S.J.; Santos, F.D.; Ferreira, S.H. Myrcene mimics the peripheral analgesic activity of lemongrass tea. J. Ethnopharmacol. 1991, 34, 43–48. [Google Scholar] [CrossRef]
- Sforcin, J.M.; Amaral, J.T.; Fernandes, A.J.; Sousa, J.P.; Bastos, J.K. Lemongrass effects on IL−1beta and IL−6 production by macrophages. Nat. Prod. Res. 2009, 23, 1151–1159. [Google Scholar] [CrossRef]
- Cuomo, F.; Cofelice, M.; Lopez, F. Rheological characterization of hydrogels from alginate−based nanodispersion. Polymers 2019, 11, 259. [Google Scholar] [CrossRef] [PubMed]
- Somparn, N.; Saenthaweeuk, S.; Naowaboot, J.; Thaeomor, A.; Kukongviriyapan, V. Effect of lemongrass water extract supplementation on atherogenic index and antioxidant status in rats. Acta Pharm. Sin. 2018, 68, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Park, J.; Lee, H.; Hwang, D.Y.; Park, S.H. Evaluation of the EtOAc extract of lemongrass (Cymbopogon citratus) as a potential skincare cosmetic material for acne vulgaris. J. Biotechnol. 2022, 32, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Gilling, D.H.; Ravishankar, S.; Bright, K.R. Antimicrobial efficacy of plant essential oils and extracts against Escherichia Coli. J. Environ. Sci. Health 2019, 54, 608–616. [Google Scholar] [CrossRef]
- Ramar, T.; Veeraperumal, S.; Kannan, S. Optimized extraction of polysaccharides from Cymbopogon citratus and its biological activities. Int. J. Biol. Macromol. 2014, 65, 415–423. [Google Scholar]
- Baek, M.W.; Choi, H.R.; Solomon, T.; Jeong, C.S.; Lee, O.-H.; Tilahun, S. Preharvest, methyl jasmonate treatment increased the antioxidant activity and glucosinolate contents of hydroponically grown Pak Choi. Antioxidants 2021, 10, 131. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, B.; Xu, H.; Wu, J.; Xu, Z.; Wang, Y. The phytophthora effector Avh94 manipulates host jasmonic acid signaling to promote infection. J. Integr. Plant Biol. 2022, 64, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Zhang, Z.; Siddiqui, M.; Ayala Zavala, J.; Hwang, C.A. Postharvest management approaches for maintaining quality of fresh produce. Springer Int. Publ. Switz. 2016, 1226, 97–111. [Google Scholar]
- Gomez, H.A.; Niederauer, G.F.; Minatel, I.O.; Antunes, E.R.M.; Carneiro, M.J.; Frankland Sawaya, A.C.H.; Zanus, M.C.; Ritschel, P.S.; Quecini, V.; Pereira Lima, G.P.; et al. Metabolite profiling reveals the influence of grapevine genetic distance on the chemical signature of juices. J. Sci. Food Agric. 2023, 1002, 13124. [Google Scholar] [CrossRef]
- Reyes-Díaz, M.; Lobos, T.; Cardemil, L.; Nunes-Nesi, A.; Retamales, J.; Jaakola, L.; Alberdi, M.; Ribera−Fonseca, A. Methyl Jasmonate: An alternative for improving the quality and health properties of fresh fruits. Molecules 2016, 21, 567. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Niknejad, Y.; Fallah, H.; Tari, D.B. Methyl jasmonate alleviates arsenic toxicity in rice. Plant Cell Rep. 2020, 39, 1041–1060. [Google Scholar] [CrossRef] [PubMed]
- He, W.W.; Luo, H.; Xu, H.; Zhou, Z.Y.; Li, D.J.; Bao, Y.H.; Fu, Q.; Song, J.F.; Jiao, Y.; Zhang, Z.Y. Effect of exogenous methyl jasmonate on physiological and carotenoid composition of yellow maize sprouts under NaCl stress. Food Chem. 2021, 361, 130177. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.L.; Liu, Y.N.; Liu, R.; Ren, A.; Ma, H.Y.; Shu, L.B.; Shi, L.; Zhu, J.; Zhao, M.W. Integrated proteomics and metabolomics analysis provides insights into ganoderic acid biosynthesis in response to Methyl Jasmonate in Ganoderma lucidum. Int. J. Mol. Med. 2019, 20, 6116. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.M.; Wang, H.D.; Zhang, Y.X.; Xi, Z.A.; Yang, R.; Sheng, J.P.; Zhang, X.H.; Ding, Y.; Wang, J.P.; Fan, Z.C. Ornithine decarboxylase is involved in methyl jasmonate−regulated postharvest quality retention in button mushrooms (Agaricus bisporus). J. Sci. Food Agric. 2019, 99, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Wiktorowska-Owczarek, A.; Berezińska, M.; Nowak, J.Z. PUFAs: Structures, Metabolism and Functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Bartella, L.; Mazzotti, F.; Talarico, I.R.; Santoro, I.; Di Donna, L. Paper spray tandem mass spectrometry for assessing oleic, linoleic and linolenic acid content in edible vegetable oils. Separations 2023, 10, 26. [Google Scholar] [CrossRef]
- Nava Lauson, C.B.; Tiberti, S.; Corsetto, P.A.; Conte, F.; Tyagi, P.; Machwirth, M.; Ebert, S.; Loffreda, A.; Scheller, L.; Sheta, D.; et al. Linoleic acid potentiates cytotoxic T lymphocytes (CD8+ T cell) metabolic fitness and antitumor immunity. Cell Metab. 2023, 35, 633–650. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.S.; Klett, E.L. Linoleic acid and the regulation of glucose homeostasis: A review of the evidence. Prostaglandins Leukot Essent Fat. Acids 2021, 175, 102366. [Google Scholar] [CrossRef] [PubMed]
- Belury, M.A. Linoleic acid, an omega−6 fatty acid that reduces risk for cardiometabolic diseases: Premise, promise and practical implications. Curr. Opin. Clin. Nutr. Metab. Care 2023, 26, 288–292. [Google Scholar] [CrossRef]
- Leon−Aparicio, D.; Sánchez-Solano, A.; Arreola, J.; Perez−Cornejo, P. Oleic acid blocks the calcium-activated chloride channel TMEM16A/ANO. Biophys. Acta 2022, 1867, 159134. [Google Scholar]
- Lou, H.; Li, H.; Wei, T.; Chen, Q. Stimulatory Effects of Oleci Acid and Fungal Elicitor on Betulinic Acid Production by Submerged Cultivation of Medicinal Mushroom Inonotus obliquus. J. Fungi 2021, 7, 266. [Google Scholar] [CrossRef] [PubMed]
- Sande, D.; Oliveira, G.P.; Moura, M.A.F.E.; Martins, B.A.; Lima, M.T.N.S.; Takahashi, J.A. Edible mushrooms as a ubiquitous source of essential fatty acids. Food Res. Int. 2019, 125, 108524. [Google Scholar] [CrossRef] [PubMed]
- Koh, G.Y.; Chou, G.; Liu, Z. Purification of a water extract of Chinese sweet tea plant (Rubus suavissimus S. Lee) by alcohol precipitation. J. Agric. Food Chem. 2009, 57, 5000–5006. [Google Scholar] [CrossRef] [PubMed]
- Weitzhandler, M.; Barreto, V.; Pohl, C.; Jandik, P.; Cheng, J.; Avdalovic, N. CarboPac PA20: A new monosaccharide separator column with electrochemical detection with disposable gold electrodes. J. Biochem. Biophys. Methods 2004, 60, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Kamran, S.H.; Ahmad, M.; Ishtiaq, S.; Ajaib, M.; Razashah, S.H.; Shahwar, D.E. Metabolite profiling and biochemical investigation of the antidiabetic potential of Loranthus pulverulentus wall n-butanol fraction in diabetic animal models. J. Ethnopharmacol. 2024, 318, 116963. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, F.A.; Tajik, H.; Jafari, K.; Hoseinzadeh, E.; Mirahmadi, S.S.; Conti, G.O.; Miri, M. Evaluation of dioxin-like polychlorinated biphenyls in fish of the Caspian Sea. MethodsX 2020, 7, 100803. [Google Scholar]
- Wang, L.; Liu, Q.; Wang, N.; Li, S.; Bian, W.; Sun, Z.; Wang, L.; Wang, L.; Liu, C.; Song, C.; et al. Oleic Acid Dissolves cGAS-DNA Phase Separation to Inhibit Immune Surveillance. Adv. Sci. 2023, 10, e2206820. [Google Scholar] [CrossRef]
- Chini, V.; Foka, A.; Dimitracopoulos, G.; Spiliopoulou, I. Absolute and relative real-time PCR in the quantification of tst gene expression among methicillin−resistant Staphylococcus aureus: Evaluation by two mathematical models. Lett. Appl. Microbiol. 2007, 45, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Fayed, B.; Kohder, G.; Soliman, S.S.M. Gas Chromatography−Mass Spectrometry (GC−MS) Analysis of Candida auris Metabolites. Methods Mol. Biol. 2022, 2517, 165–172. [Google Scholar]
- Wang, F. Semi-Quantitative RT-PCR: An Effective Method to Explore the Regulation of Gene Transcription Level Affected by Environmental Pollutants. Methods Mol. Biol. 2021, 2326, 95–103. [Google Scholar]
- Chuang, L.Y.; Cheng, Y.H.; Yang, C.H. Specific primer design for the polymerase chain reaction. Biotechnol. Lett. 2013, 35, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Guo, B.; Sun, H.; Zhang, J.; Liu, S.; Hexige, S.; Yu, X.; Wang, X. Identification of the key genes implicated in the transformation of OLP to OSCC using RNA−sequencing. Oncol. Rep. 2017, 37, 2355–2365. [Google Scholar] [CrossRef] [PubMed]
- Patuel, S.J.; English, C.; Lopez-Scarim, V.; Konig, I.; Ivantsova, E.; Martyniuk, C.J. Dataset for diseases associated with exposure to broflanilide, a novel pesticide, in larval zebrafish (Danio rerio). Data Brief 2023, 50, 109534. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Guo, Z.; Wang, P.; Liu, Z.; Wang, Z. Transcriptomic analysis reveals the potential crosstalk genes and immune relationship between IgA nephropathy and periodontitis. Front. Immunol. 2023, 30, 1062590. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Segura-Alabart, N.; Serratosa, F. The multi-furcating Neighbor−Joining algorithm for reconstructing polytomic phylogenetic trees. J. Mol. Evol. 2023, 91, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 10, 1093. [Google Scholar]
- Rosa, S.B.A.; Csordas, B.G.; do Valle Leone de Oliveira, S.M.; Ribeiro dos Santos, A.; Paniago, A.M.M.; Venturini, J. Prediction of conserved peptides of paracoccidioides for interferon−γ release assay: The first step in the development of a lab-based approach for immunological assessment during antifungal therapy. J. Fungi 2020, 6, 379. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Zhou, Z.; Huayu, M. A multi-epitope vaccine GILE against echinococcus multilocularis infection in mice. Front. Immunol. 2023, 13, 1091004. [Google Scholar] [CrossRef]
- Meng, L.; Luo, B.; Yang, Y.; Faruque, M.O.; Zhang, J.; Li, X.; Hu, X. Addition of vegetable oil to improve triterpenoids production in liquid fermentation of medicinal fungus Antrodia cinnamomea. J. Fungi 2021, 7, 926. [Google Scholar] [CrossRef]
- Cao, C.; Zhong, H.; Chen, Z.; Song, Z.; Hu, B.; Wang, X. Triterpene acid fromnalysis Version 11. Mol. Biol. Evol. 2023, 91, 773–777. [Google Scholar]
- Kuang, Y.; Li, B.; Wang, Z.; Qiao, X.; Ye, M. Terpenoids from the medicinal mushroom Antrodia camphorata: Chemistry and medicinal potential. Nat. Prod. Rep. 2021, 38, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Menon, M.P.; Chien, Y.H.; Thomas, J.; Yu, Y.H.; Chang, C.T.; Hua, K.F. Nano Modification of Antrodia cinnamomea Exhibits Anti-Inflammatory Action and Improves the Migratory Potential of Myogenic Progenitors. Cells 2022, 11, 2512. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.D.; Zhu, L.P.; Guo, Y.T.; Zhao, Y.M.; Betchem, G.B.; Yolandani, Y.; Ma, H.L. Enhancing submerged fermentation of Antrodia camphorata by low−frequency alternating magnetic field. Innov. Food Sci. Emerg. Technol. 2023, 86, 103382. [Google Scholar] [CrossRef]
- Suzuki, D.; Sato, Y.; Kamasaka, H.; Kuriki, T. Effects of phosphoryl oligosaccharides of calcium (POs−Ca) on mycelial growth and fruiting body development of the edible mushroom, Pleurotus ostreatus. J. Appl. Glycosci. 2020, 67, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Nargotra, P.; Kuo, C.H.; Liu, Y.C. High−molecular−weight exopolysaccharides production from Tuber borchii cultivated by submerged fermentation. Int. J. Mol. Sci. 2023, 24, 4875. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.T.; Li, D.Y.; Wang, Z.; Zang, W.T.; Rao, P.; Liang, Y.X.; Mei, Y.X. Alpha-terpineol affects synthesis and antitumor activity of triterpenoids from Antrodia cinnamomea mycelia in solid-state culture. Food Funct. 2018, 12, 6517–6525. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Xu, Y.H.; Xiong, B.; Lin, D.; Huang, S.J.; Dong, T.T.; Sun, G.C.; Liao, L.; Deng, Q.X.; Wang, X.; et al. Effects of exogenous methyl jasmonate on the synthesis of endogenous jasmonates and the regulation of photosynthesis in citrus. Photobiol. Photosynth. 2020, 20, 15. [Google Scholar]
- Felipe, M.R.; Martin, R.; Tomas, C.; Ondrej, K. Analysis of fatty acid esters of hydroxy fatty acids in edible mushrooms. Food Sci. Technol. 2023, 173, 114311. [Google Scholar]
- Turk, A.; Lee, S.; Lee, H.H.; Yeon, S.W.; Ryu, S.H.; Seo, G.H.; Chang, H.Y.; Hwang, B.Y.; Lee, M.K. α−Glucosidase inhibitory fatty acids from Morchella fluvialis mushroom. Acta Pharm. 2023, 17, 369–379. [Google Scholar] [CrossRef]
- Dimitrijevic, M.V.; Mitic, V.D.; Nikolic, J.S. First report about mineral content, fatty acids composition and biological activities of four wild edible mushrooms. Chem. Biodivers. 2019, 16, e1800492. [Google Scholar] [CrossRef]
- Zhou, T.T.; Hu, W.K.; Yang, Z.B.; Li, J.M.; Zeng, X.F. Study on nutrients, non-volatile compounds, volatile compounds and antioxidant capacity of oyster mushroom cultivated with corn distillers’ grains. Food Sci. Technol. 2023, 183, 114967. [Google Scholar] [CrossRef]
- Cao, S.; Zhang, J.; Cheng, H. Identification and evolutionary analysis of FAD2 gene family in green plants. Trop. Plant Biol. 2021, 14, 239–250. [Google Scholar] [CrossRef]
- Liu, X.F.; Xia, Y.J.; Lai, P.F.H. An increase in cell membrane permeability in the in situ extractive fermentation improves the production of antroquinonol from Antrodia camphorata S−29. J. Ind. Microbiol. Biotechnol. 2020, 47, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wang, W.; Wei, L.; Chen, P.; Peng, L.; Qin, Z.; Yuan, F.; Wang, Z.; Ying, X. The evolution and biocatalysis of FAD2 indicate its correlation to the content of seed oil in plants. Int. J. Mol. Sci. 2019, 20, 849. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.C.; Nakamura, Y.; Kanehara, K. Membrane lipid polyunsaturation mediated by fatty acid desaturase 2 (FAD2) is involved in endoplasmic reticulum stress tolerance in Arabidopsis thaliana. Plant J. 2019, 99, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Pan, W.S.; Peng, Q.; Chen, Y.C.; Zhou, T.; Wu, C.; Hartley, H.; Li, J.; Xu, M.H.; Liu, C.W. Characteristics of Metabolites by Seed−Specific Inhibition of FAD2 in Brassica napus L. Journal of Agricultural and Food Chemistry. 2021, 69, 5452–5462. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.K.; Mishra, G. Functional characterization and expression profile of microsomal FAD2 and FAD3 genes involved in linoleic and α−linolenic acid production in Leucas cephalotes. Physiol. Mol. Biol. Plants 2021, 27, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Mauvoisin, D.; Mounier, C. Hormonal and nutritional regulation of SCD1 gene expression. Biochimie 2011, 93, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Tsukamoto, H. Stearoyl−CoA desaturase and tumorigenesis. Chem. -Biol. Interact. 2020, 316, 108917. [Google Scholar] [CrossRef]
- Tracz-Gaszewska, Z.; Dobrzyn, P. Stearoyl−CoA Desaturase 1 as a therapeutic target for the treatment of cancer. Cancers 2019, 11, 948. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) |
|---|---|
| SCD F | CTTGATTGGCGTTTGGATCT |
| SCD R | ATGCCAGAAGAAGCGACCTA |
| Cytochrome P450 monooxygenase (CYP 151) F | GGATGTCGTTGCGCAGATTC |
| Cytochrome P450 monooxygenase (CYP 151) R | CGACCACCACTGCGATCATA |
| Putative aryl−alcohol dehydrogenase (pacs) F | ATCAGTTCGTCGGCCATTAC |
| Putative aryl−alcohol dehydrogenase (pacs) R | TGCAGTGAGTGTGACGTTGA |
| FAD2−1 F | ACAATGCGCACCATAAAACA |
| FAD2−1 R | CCCATCGCGTTAGTCGTTAT |
| O-methyltransferase gedA (O−myse) F | AGTGCCTCCCTAGTGCAGAA |
| O-methyltransferase gedA (O−myse) R | ATCAGGCTGCTCAATTTGCT |
| Acyl-coenzyme A thioesterase (AcAs) F | GAAACTGTGGGACCAGGAGA |
| Acyl-coenzyme A thioesterase (AcAs) R | CGTGTATTGTCTGCCCATTG |
| Acyl-CoA dehydrogenase (ACD) F | GGGCATTCAAATCGCTAAAA |
| Acyl-CoA dehydrogenase (ACD) R | GATACGCCCTTCATTCAGGA |
| Short acyl−CoA dehydrogenase scds F | AAGGTGATGTGCTTGCACTG |
| Short acyl−CoA dehydrogenase (scds) R | CTTGCCATATTTGAGCAGCA |
| Acyl-coenzyme A thioesterase (Acdse) F | GCCAAGGAACTGCAGAAGAC |
| Acyl-coenzyme A thioesterase (Acdse) R | GATCTCGTGTAAGCGCATGA |
| Linoleate 10R−lipoxygenase (LA10r−se) F | AGCAGCTGCACAAGACTTCA |
| Linoleate 10R−lipoxygenase (LA10r−se) R | CTCCCGCATAACCTTGACAT |
| Vanillin dehydrogenase (VDse) F | TCAGCAACTTCAACCACGAG |
| Vanillin dehydrogenase (VDse) R | CACTTCAGGCTCTCCTCCAC |
| Branched chain-specific acyl−CoA dehydrogenase (Bcsdse) F | TAGCCTTTGTGCTGTCATCG |
| Branched chain-specific acyl−CoA dehydrogenase (Bcsdse) R | CCTCTTCTTCGCACGACTTC |
| FAD2−2 F | TGCTGACTTATCTCCATCATTCC |
| FAD2−2 R | CTTGGACCCCATTGTCTTG |
| Actin F | ATTCTCGTCTCATGGCCTTTC |
| Actin R | GTCGCAGCTGGAGTTGATTA |
| Sample | Raw Read Number (bp) | Raw Bases (bp) | Raw Q30 Number (bp) | Raw Nrate | Raw Q20 Rate (%) | Raw Q30 Rate (%) |
|---|---|---|---|---|---|---|
| CK1 | 43,988,436 | 6,598,265,400 | 6,005,976,301 | 0.000586 | 96.82 | 91.02 |
| CK2 | 42,045,370 | 6,306,805,500 | 5,705,731,106 | 0.000577 | 96.57 | 90.46 |
| CK3 | 42,579,874 | 6,386,981,100 | 5,799,211,776 | 0.000554 | 96.79 | 90.79 |
| LWE1 | 41,970,376 | 6,295,556,400 | 5,680,465,685 | 0.00064 | 96.45 | 90.22 |
| LWE2 | 42,804,348 | 6,420,652,200 | 5,779,971,506 | 0.000638 | 96.34 | 90.02 |
| LWE3 | 42,313,142 | 6,346,971,300 | 5,780,384,737 | 0.000617 | 96.8 | 91.07 |
| MJ1 | 45,274,450 | 6,791,167,500 | 6,160,571,236 | 0.000567 | 96.73 | 90.71 |
| MJ2 | 41,820,156 | 6,273,023,400 | 5,706,592,462 | 0.00056 | 96.83 | 90.97 |
| MJ3 | 43,813,866 | 6,572,079,900 | 5,991,770,058 | 0.000618 | 96.86 | 91.17 |
| Sample | Trimmed Read Number (bp) | Trimmed Bases (bp) | Useful Read (%) | Useful Bases (%) |
|---|---|---|---|---|
| CK1 | 41,367,030 | 6,205,054,500 | 94.04 | 94.04 |
| CK2 | 39,768,216 | 5,965,232,400 | 94.58 | 94.58 |
| CK3 | 40,409,158 | 6,061,373,700 | 94.9 | 94.9 |
| LWE1 | 39,689,780 | 5,953,467,000 | 94.56 | 94.56 |
| LWE2 | 40,401,392 | 6,060,208,800 | 94.38 | 94.38 |
| LWE3 | 39,986,370 | 5,997,955,500 | 94.5 | 94.5 |
| MJ1 | 42,780,564 | 6,417,084,600 | 94.49 | 94.49 |
| MJ2 | 39,540,216 | 5,931,032,400 | 94.54 | 94.54 |
| MJ3 | 41,449,692 | 6,217,453,800 | 94.6 | 94.6 |
| Sample | Total Mapped | Uniquely Mapped | Map Events | Mapped to Gene | Mapped to Exon |
|---|---|---|---|---|---|
| CK1 | 39,170,219 (94.69%) | 38,449,806 (98.16%) | 38,449,806 | 36,413,793 (94.70%) | 36,302,523 (99.69%) |
| CK2 | 37,451,028 (94.17%) | 36,756,202 (98.14%) | 36,756,202 | 34,864,214 (94.85%) | 34,773,751 (99.74%) |
| CK3 | 37,713,189 (93.33%) | 37,083,669 (98.33%) | 37,083,669 | 35,015,905 (94.42%) | 34,881,302 (99.62%) |
| LWE1 | 37,624,754 (94.80%) | 36,909,466 (98.10%) | 36,909,466 | 34,927,870 (94.63%) | 34,836,929 (99.74%) |
| LWE2 | 38,302,378 (94.80%) | 37,531,270 (97.99%) | 37,531,270 | 35,399,289 (94.32%) | 35,288,391 (99.69%) |
| LWE3 | 37,636,026 (94.12%) | 36,900,546 (98.05%) | 36,900,546 | 34,847,007 (94.43%) | 34,745,804 (99.71%) |
| MJ1 | 40,113,869 (93.77%) | 39,269,479 (97.90%) | 39,269,479 | 37,244,117 (94.84%) | 37,144,889 (99.73%) |
| MJ2 | 37,641,204 (95.20%) | 36,754,340 (97.64%) | 36,754,340 | 34,869,880 (94.87%) | 34,782,747 (99.75%) |
| MJ3 | 38,788,515 (93.58%) | 38,066,007 (98.14%) | 38,066,007 | 36,089,745 (94.81%) | 35,984,942 (99.71%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, Y.; Lin, Z.; Lin, D.; Lin, B.; Chen, G.; Lin, Z.; Li, J. Cymbopogon citratus Water Extract and Methyl Jasmonate Improve Polyunsaturated Fatty Acid Metabolism in Taiwanofungus camphoratus Mycelia. Separations 2024, 11, 127. https://doi.org/10.3390/separations11040127
Wen Y, Lin Z, Lin D, Lin B, Chen G, Lin Z, Li J. Cymbopogon citratus Water Extract and Methyl Jasmonate Improve Polyunsaturated Fatty Acid Metabolism in Taiwanofungus camphoratus Mycelia. Separations. 2024; 11(4):127. https://doi.org/10.3390/separations11040127
Chicago/Turabian StyleWen, Yeyan, Zixuan Lin, Dongmei Lin, Biaosheng Lin, Gexin Chen, Zhanxi Lin, and Jing Li. 2024. "Cymbopogon citratus Water Extract and Methyl Jasmonate Improve Polyunsaturated Fatty Acid Metabolism in Taiwanofungus camphoratus Mycelia" Separations 11, no. 4: 127. https://doi.org/10.3390/separations11040127