

Bioactives in Cocoa: Novel Findings, Health Benefits, and Extraction Techniques
 , and
, and
Abstract
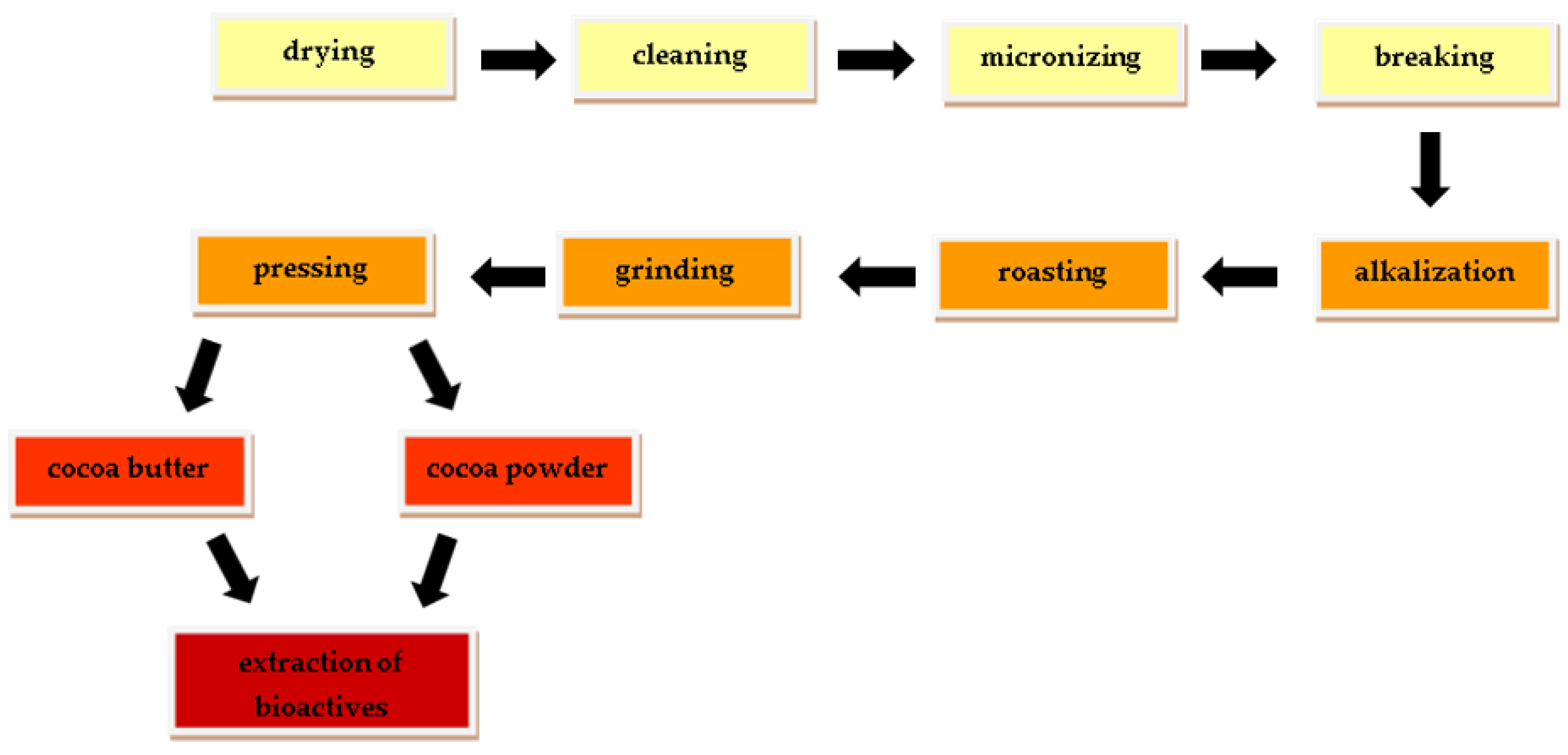
:1. Introduction
2. Bioactives in Cocoa
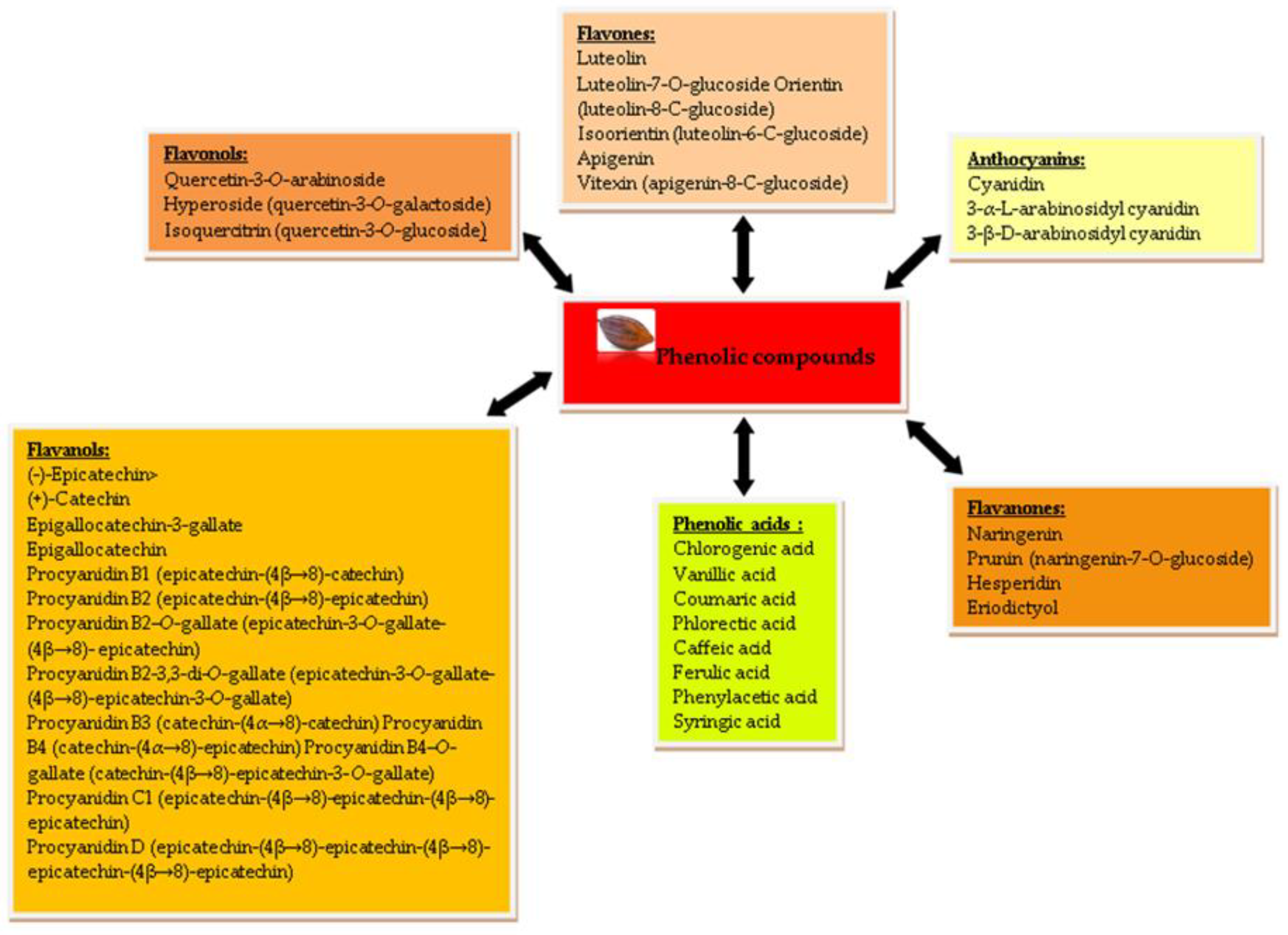
2.1. Polyphenols
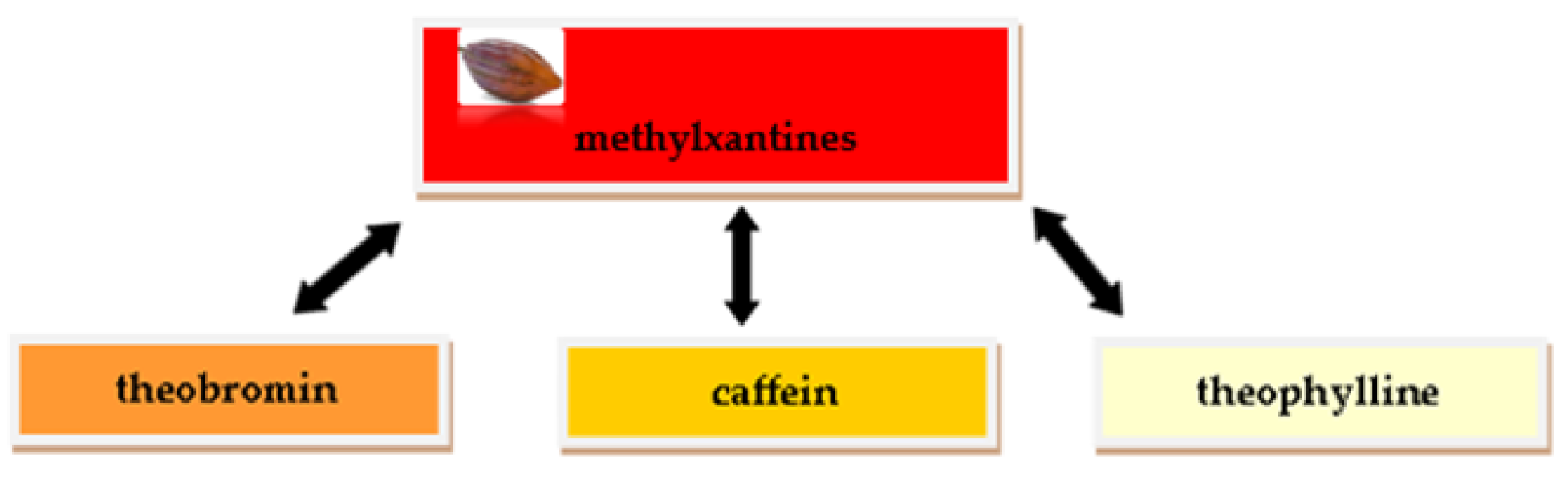
2.2. Methylxanthin
2.3. Phytosterols
2.4. Dietary Fibers
3. Health Benefits of Bioactives from Cocoa
3.1. Effects on Cardiovascular Diseases
3.2. Effects on Cognitive Functions
3.3. Effects on Gut and Gut Microbiota
3.4. Effects on Diabetes
3.5. Effects on Obesity
4. Extraction of Bioactive Compounds from Cocoa
- –
- Ultrasound-assisted extraction (UAE): utilizes ultrasound waves to disrupt plant cell walls, enhancing solvent penetration and bioactive compound release;
- –
- Enzyme-assisted extraction (EAE): employs enzymes to selectively break down plant cell walls, facilitating the release of target compounds;
- –
- Microwave-assisted extraction (MAE): harnesses microwave energy to rapidly heat solvents, promoting faster extraction and potentially improving yield;
- –
- Pulsed electric field-assisted extraction (PEF): applies short, high-voltage electric pulses to create temporary pores in plant cell membranes, allowing for improved solvent access and extraction efficiency;
- –
- Supercritical fluid extraction (SFE): utilizes fluids above their critical point (high temperature and pressure) to act as both solvent and solute carrier, offering high selectivity and purity for extracted compounds;
- –
- Liquid extraction under pressure (PLE): employs heated pressurized solvents to enhance extraction efficiency, often utilizing pressurized static or dynamic extraction processes;
- –
- Ohmic heater-assisted extraction (OHAE): applies an electric current to directly heat the extraction solution, offering rapid and efficient heating for improved extraction yields.
4.1. Ultrasound-Assisted Extraction (UAE)
4.2. Enzyme-Assisted Extraction (EAE)
4.3. Microwave-Assisted Extraction (MAE)
4.4. Pulsed Electric Field-Assisted Extraction (PEF)
4.5. Supercritical Fluid Extraction (SFE)
4.6. Pressurized Liquid Extraction (PLE)
5. Future Research Trends
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jean-Marie, E.; Bereau, D.; Robinson, J.C. Benefits of Polyphenols and Methylxanthines from Cocoa Beans on Dietary Metabolic Disorders. Foods 2021, 10, 2049. [Google Scholar] [CrossRef]
- Goya, L.; Kongor, J.E.; de Pascual-Teresa, S. From Cocoa to Chocolate: Effect of Processing on Flavanols and Methylxanthines and Their Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 14365. [Google Scholar] [CrossRef] [PubMed]
- Verna, R. The History and Science of Chocolate. Malays. J. Pathol. 2013, 35, 111–121. [Google Scholar]
- Kim, J.; Kim, J.; Shim, J.; Lee, C.Y.; Lee, K.W.; Lee, H.J. Cocoa Phytochemicals: Recent Advances in Molecular Mechanisms on Health. Crit. Rev. Food Sci. Nutr. 2014, 54, 1458–1472. [Google Scholar] [CrossRef]
- Martin, M.Á.; Ramos, S. Impact of Cocoa Flavanols on Human Health. Food Chem. Toxicol. 2021, 151, 112121. [Google Scholar] [CrossRef] [PubMed]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and Chocolate in Human Health and Disease. Antioxid. Redox Signal. 2011, 15, 2779. [Google Scholar] [CrossRef] [PubMed]
- Badrie, N.; Bekele, F.; Sikora, E.; Sikora, M. Cocoa Agronomy, Quality, Nutritional, and Health Aspects. Crit. Rev. Food Sci. Nutr. 2015, 55, 620–659. [Google Scholar] [CrossRef] [PubMed]
- Andújar, I.; Recio, M.C.; Giner, R.M.; Ríos, J.L. Cocoa Polyphenols and Their Potential Benefits for Human Health. Oxid. Med. Cell. Longev. 2012, 2012, 906252. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.A.; Goya, L.; Ramos, S. Potential for Preventive Effects of Cocoa and Cocoa Polyphenols in Cancer. Food Chem. Toxicol. 2013, 56, 336–351. [Google Scholar] [CrossRef]
- Martin, M.Á.; Goya, L.; Ramos, S. Antidiabetic Actions of Cocoa Flavanols. Mol. Nutr. Food Res. 2016, 60, 1756–1769. [Google Scholar] [CrossRef]
- Edo, G.I.; Samuel, P.O.; Oloni, G.O.; Ezekiel, G.O.; Onoharigho, F.O.; Oghenegueke, O.; Nwachukwu, S.C.; Rapheal, O.A.; Ajokpaoghene, M.O.; Okolie, M.C.; et al. Review on the Biological and Bioactive Components of Cocoa (Theobroma cacao). Insight on Food, Health and Nutrition. Nat. Resour. Hum. Health 2023, 3, 426–448. [Google Scholar] [CrossRef]
- Guaadaoui, A.; Benaicha, S.; Elmajdoub, N.; Bellaoui, M.; Hamal, A. What Is a Bioactive Compound? A Combined Definition for a Preliminary Consensus. Int. J. Nutr. Food Sci. 2014, 3, 174–179. [Google Scholar] [CrossRef]
- Cooper, K.A.; Campos-Giménez, E.; Alvarez, D.J.; Nagy, K.; Donovan, J.L.; Williamson, G. Rapid Reversed Phase Ultra-Performance Liquid Chromatography Analysis of the Major Cocoa Polyphenols and Inter-Relationships of Their Concentrations in Chocolate. J. Agric. Food Chem. 2007, 55, 2841–2847. [Google Scholar] [CrossRef]
- Di Mattia, C.D.; Sacchetti, G.; Mastrocola, D.; Serafini, M. From Cocoa to Chocolate: The Impact of Processing on in Vitro Antioxidant Activity and the Effects of Chocolate on Antioxidant Markers in Vivo. Front. Immunol. 2017, 8, 1207. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.M.; Jalil, A.M.M.; Ismail, A. Phenolic and Theobromine Contents of Commercial Dark, Milk and White Chocolates on the Malaysian Market. Molecules 2009, 14, 200–209. [Google Scholar] [CrossRef]
- Othman, A.; Ismail, A.; Abdul Ghani, N.; Adenan, I. Antioxidant Capacity and Phenolic Content of Cocoa Beans. Food Chem. 2007, 100, 1523–1530. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Luca, S.V.; Miron, A. Flavor Chemistry of Cocoa and Cocoa Products—An Overview. Compr. Rev. Food Sci. Food Saf. 2016, 15, 73–91. [Google Scholar] [CrossRef]
- Melo, T.S.; Pires, T.C.; Engelmann, J.V.P.; Monteiro, A.L.O.; Maciel, L.F.; Bispo, E. da S. Evaluation of the Content of Bioactive Compounds in Cocoa Beans during the Fermentation Process. J. Food Sci. Technol. 2021, 58, 1947–1957. [Google Scholar] [CrossRef]
- Toker, O.S.; Palabiyik, I.; Pirouzian, H.R.; Aktar, T.; Konar, N. Chocolate Aroma: Factors, Importance and Analysis. Trends Food Sci. Technol. 2020, 99, 580–592. [Google Scholar] [CrossRef]
- Hernández-Hernández, C.; Viera-Alcaide, I.; Morales-Sillero, A.M.; Fernández-Bolaños, J.; Rodríguez-Gutiérrez, G. Bioactive Compounds in Mexican Genotypes of Cocoa Cotyledon and Husk. Food Chem. 2018, 240, 831–839. [Google Scholar] [CrossRef]
- Afoakwa, E.O.; Quao, J.; Takrama, F.S.; Budu, A.S.; Saalia, F.K. Changes in Total Polyphenols, o-Diphenols and Anthocyanin Concentrations during Fermentation of Pulp Pre-Conditioned Cocoa (Theobroma cacao) Beans. Int. Food Res. J. 2012, 19, 1071–1077. [Google Scholar]
- Delgado-Ospina, J.; Di Mattia, C.D.; Paparella, A.; Mastrocola, D.; Martuscelli, M.; Chaves-Lopez, C. Effect of Fermentation, Drying and Roasting on Biogenic Amines and Other Biocompounds in Colombian Criollo Cocoa Beans and Shells. Foods 2020, 9, 520. [Google Scholar] [CrossRef] [PubMed]
- Barišić, V.; Kopjar, M.; Jozinović, A.; Flanjak, I.; Ačkar, Đ.; Miličević, B.; Šubarić, D.; Jokić, S.; Babić, J. The Chemistry behind Chocolate Production. Molecules 2019, 24, 3163. [Google Scholar] [CrossRef] [PubMed]
- De Taeye, C.; Caullet, G.; Eyamo Evina, V.J.; Collin, S. Procyanidin A2 and Its Degradation Products in Raw, Fermented, and Roasted Cocoa. J. Agric. Food Chem. 2017, 65, 1715–1723. [Google Scholar] [CrossRef] [PubMed]
- Oracz, J.; Nebesny, E.; Żyżelewicz, D. Changes in the Flavan-3-Ols, Anthocyanins, and Flavanols Composition of Cocoa Beans of Different Theobroma cacao L. Groups Affected by Roasting Conditions. Eur. Food Res. Technol. 2015, 241, 663–681. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef]
- Fowler, M.S. Cocoa Beans: From Tree to Factory. In Industrial Chocolate Manufacture and Use, 4th ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; pp. 10–47. ISBN 1405139498. [Google Scholar]
- Bravo, L. Polyphenols: Chemistry, Dietary Sources, Metabolism, and Nutritional Significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.A.; Ramos, S. Cocoa Polyphenols in Oxidative Stress: Potential Health Implications. J. Funct. Foods 2016, 27, 570–588. [Google Scholar] [CrossRef]
- Ramiro-Puig, E.; Castell, M. Cocoa: Antioxidant and Immunomodulator. Br. J. Nutr. 2009, 101, 931–940. [Google Scholar] [CrossRef]
- Jalil, A.M.M.; Ismail, A. Polyphenols in Cocoa and Cocoa Products: Is There a Link between Antioxidant Properties and Health? Molecules 2008, 13, 2190. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, L.C.; Londoño-Londoño, J.; Gil, A. Comparison of Polyphenol, Methylxanthines and Antioxidant Activity in Theobroma Cacao Beans from Different Cocoa-Growing Areas in Colombia. Food Res. Int. 2014, 60, 273–280. [Google Scholar] [CrossRef]
- Camu, N.; De Winter, T.; Addo, S.K.; Takrama, J.S.; Bernaert, H.; De Vuyst, L. Fermentation of Cocoa Beans: Influence of Microbial Activities and Polyphenol Concentrations on the Flavour of Chocolate. J. Sci. Food Agric. 2008, 88, 2288–2297. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Borges, G.; Nagai, C.; Jackson, M.C.; Yokota, T.; Crozier, A.; Ashihara, H. Profiles of Phenolic Compounds and Purine Alkaloids during the Development of Seeds of Theobroma Cacao cv. Trinitario. J. Agric. Food Chem. 2013, 61, 427–434. [Google Scholar] [CrossRef]
- Thi, D.; Yen, K.; Vu, N.; Ha, H. Effects of Maturity Stages and Fermentation of Cocoa Beans on Total Phenolic Contents and Antioxidant Capacities in Raw Cocoa Powder. Vietnam J. Biotechnol. 2016, 14, 743–752. [Google Scholar] [CrossRef]
- do Carmo Brito, B.D.N.; Campos Chisté, R.; da Silva Pena, R.; Abreu Gloria, M.B.; Santos Lopes, A. Bioactive Amines and Phenolic Compounds in Cocoa Beans Are Affected by Fermentation. Food Chem. 2017, 228, 484–490. [Google Scholar] [CrossRef] [PubMed]
- EFRAIM, P.; Barreto ALVES, A.; Calil Pereira JARDIM, D. Revisão: Polifenóis Em Cacau e Derivados: Teores, Fatores de Variação e Efeitos Na Saúde. Braz. J. Food Technol. 2011, 14, 181–201. [Google Scholar] [CrossRef]
- Sorrenti, V.; Ali, S.; Mancin, L.; Davinelli, S.; Paoli, A.; Scapagnini, G. Cocoa Polyphenols and Gut Microbiota Interplay: Bioavailability, Prebiotic Effect, and Impact on Human Health. Nutrients 2020, 12, 1908. [Google Scholar] [CrossRef]
- D’Archivio, M.; Filesi, C.; Varì, R.; Scazzocchio, B.; Masella, R. Bioavailability of the Polyphenols: Status and Controversies. Int. J. Mol. Sci. 2010, 11, 1321. [Google Scholar] [CrossRef]
- Rechner, A.R.; Smith, M.A.; Kuhnle, G.; Gibson, G.R.; Debnam, E.S.; Srai, S.K.S.; Moore, K.P.; Rice-Evans, C.A. Colonic Metabolism of Dietary Polyphenols: Influence of Structure on Microbial Fermentation Products. Free Radic. Biol. Med. 2004, 36, 212–225. [Google Scholar] [CrossRef]
- Metere, A.; Giacomelli, L. Absorption, Metabolism and Protective Role of Fruits and Vegetables Polyphenols against Gastric Cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5820–5858. [Google Scholar] [CrossRef]
- Oracz, J.; Nebesny, E.; Zyzelewicz, D.; Budryn, G.; Luzak, B. Bioavailability and Metabolism of Selected Cocoa Bioactive Compounds: A Comprehensive Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1947–1985. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Fortinguerra, S.; Caudullo, G.; Buriani, A. Deciphering the Role of Polyphenols in Sports Performance: From Nutritional Genomics to the Gut Microbiota toward Phytonutritional Epigenomics. Nutrients 2020, 12, 1265. [Google Scholar] [CrossRef] [PubMed]
- D’Archivio, M.; Filesi, C.; Di Benedetto, R.; Gargiulo, R.; Giovannini, C.; Masella, R. Polyphenols, Dietary Sources and Bioavailability. Ann. Ist. Super. Sanita 2007, 43, 348–361. [Google Scholar] [PubMed]
- Llerena, W.; Samaniego, I.; Vallejo, C.; Arreaga, A.; Zhunio, B.; Coronel, Z.; Quiroz, J.; Angós, I.; Carrillo, W. Profile of Bioactive Components of Cocoa (Theobroma cacao L.) By-Products from Ecuador and Evaluation of Their Antioxidant Activity. Foods 2023, 12, 2583. [Google Scholar] [CrossRef] [PubMed]
- Martínez, R.; Torres, P.; Meneses, M.A.; Figueroa, J.G.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical, Technological and in Vitro Antioxidant Properties of Cocoa (Theobroma cacao L.) Co-Products. Food Res. Int. 2012, 49, 39–45. [Google Scholar] [CrossRef]
- Okiyama, D.C.G.; Navarro, S.L.B.; Rodrigues, C.E.C. Cocoa Shell and Its Compounds: Applications in the Food Industry. Trends Food Sci. Technol. 2017, 63, 103–112. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Zhu, F. Changes in the Composition of Methylxanthines, Polyphenols, and Volatiles and Sensory Profiles of Cocoa Beans from the Sul 1 Genotype Affected by Fermentation. J. Agric. Food Chem. 2020, 68, 8658–8675. [Google Scholar] [CrossRef] [PubMed]
- Febrianto, N.A.; Zhu, F. Composition of Methylxanthines, Polyphenols, Key Odorant Volatiles and Minerals in 22 Cocoa Beans Obtained from Different Geographic Origins. LWT 2022, 153, 112395. [Google Scholar] [CrossRef]
- Dang, Y.K.T.; Nguyen, H.V.H. Effects of Maturity at Harvest and Fermentation Conditions on Bioactive Compounds of Cocoa Beans. Plant Foods Hum. Nutr. 2019, 74, 54–60. [Google Scholar] [CrossRef]
- Neufingerl, N.; Zebregs, Y.E.M.P.; Schuring, E.A.H.; Trautwein, E.A. Effect of Cocoa and Theobromine Consumption on Serum HDL-Cholesterol Concentrations: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2013, 97, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pinilla, E.; Oñatibia-Astibia, A.; Franco, R. The Relevance of Theobromine for the Beneficial Effects of Cocoa Consumption. Front. Pharmacol. 2015, 6, 126866. [Google Scholar] [CrossRef] [PubMed]
- Aneja, M.; Gianfagna, T. Induction and Accumulation of Caffeine in Young, Actively Growing Leaves of Cocoa (Theobroma cacao L.) by Wounding or Infection with Crinipellis Perniciosa. Physiol. Mol. Plant Pathol. 2001, 59, 13–16. [Google Scholar] [CrossRef]
- Ahmed, S.; Griffin, T.S.; Kraner, D.; Schaffner, M.K.; Sharma, D.; Hazel, M.; Leitch, A.R.; Orians, C.M.; Han, W.; Stepp, J.R.; et al. Environmental Factors Variably Impact Tea Secondary Metabolites in the Context of Climate Change. Front. Plant Sci. 2019, 10, 939. [Google Scholar] [CrossRef] [PubMed]
- Cabral, C.E.; Klein, M.R.S.T. Phytosterols in the Treatment of Hypercholesterolemia and Preventionof Cardiovascular Diseases. Arq. Bras. Cardiol. 2017, 109, 475. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Escrig, A.; Santos-Hidalgo, A.B.; Saura-Calixto, F. Common Sources and Estimated Intake of Plant Sterols in the Spanish Diet. J. Agric. Food Chem. 2006, 54, 3462–3471. [Google Scholar] [CrossRef]
- Martianto, D.; Bararah, A.; Andarwulan, N.; Średnicka-Tober, D. Cross-Sectional Study of Plant Sterols Intake as a Basis for Designing Appropriate Plant Sterol-Enriched Food in Indonesia. Nutrients 2021, 13, 452. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, A.M.; Waśkiewicz, A.; Zujko, M.E.; Mirończuk-Chodakowska, I.; Cicha-Mikołajczyk, A.; Drygas, W. Assessment of Plant Sterols in the Diet of Adult Polish Population with the Use of a Newly Developed Database. Nutrients 2021, 13, 2722. [Google Scholar] [CrossRef]
- Lesma, G.; Luraghi, A.; Bavaro, T.; Bortolozzi, R.; Rainoldi, G.; Roda, G.; Viola, G.; Ubiali, D.; Silvani, A. Phytosterol and γ-Oryzanol Conjugates: Synthesis and Evaluation of Their Antioxidant, Antiproliferative, and Anticholesterol Activities. J. Nat. Prod. 2018, 81, 2212–2221. [Google Scholar] [CrossRef]
- Vilahur, G.; Ben-Aicha, S.; Diaz-Riera, E.; Badimon, L.; Padró, T. Phytosterols and Inflammation. Curr. Med. Chem. 2019, 26, 6724–6734. [Google Scholar] [CrossRef]
- Li, Q.; Xing, B. A Phytosterol-Enriched Spread Improves Lipid Profile and Insulin Resistance of Women with Gestational Diabetes Mellitus: A Randomized, Placebo-Controlled Double-Blind Clinical Trial. Diabetes Technol. Ther. 2016, 18, 499–504. [Google Scholar] [CrossRef]
- Guo, X.X.; Zeng, Z.; Qian, Y.Z.; Qiu, J.; Wang, K.; Wang, Y.; Ji, B.P.; Zhou, F. Wheat Flour, Enriched with γ-Oryzanol, Phytosterol, and Ferulic Acid, Alleviates Lipid and Glucose Metabolism in High-Fat-Fructose-Fed Rats. Nutrients 2019, 11, 1697. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xu, M.; Fang, Y.J.; Lu, M.S.; Pan, Z.Z.; Huang, W.Q.; Chen, Y.M.; Zhang, C.X. Association between Phytosterol Intake and Colorectal Cancer Risk: A Case-Control Study. Br. J. Nutr. 2017, 117, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Luo, Z.H.; Shi, H.W.; Zhang, L.X.; Ma, X.J. Research Advance of Functional Plant Pharmaceutical Cycloartenol about Pharmacological and Physiological Activity. Zhongguo Zhong Yao Za Zhi 2017, 42, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Hansel, B.; Courie, R.; Bayet, Y.; Delestre, F.; Bruckert, E. Phytostérols et Athérosclérose. La Rev. Méd. Interne 2011, 32, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Oracz, J.; Nebesny, E.; Zyzelewicz, D. Effect of Roasting Conditions on the Fat, Tocopherol, and Phytosterol Content and Antioxidant Capacity of the Lipid Fraction from Cocoa Beans of Different Theobroma cacao L. Cultivars. Eur. J. Lipid Sci. Technol. 2014, 116, 1002–1014. [Google Scholar] [CrossRef]
- Zarabadipour, F.; Piravi-Vanak, Z.; Aminifar, M. Evaluation of Sterol Composition in Different Formulations of Cocoa Milk as Milk Fat Purity Indicator. Food Sci. Technol. 2020, 41, 519–523. [Google Scholar] [CrossRef]
- Trowell, H. Definition of Dietary Fiber and Hypotheses That It Is a Protective Factor in Certain Diseases. Am. J. Clin. Nutr. 1976, 29, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Qian, Y.; Pan, Y.; Li, P.; Yang, J.; Ye, X.; Xu, G. Association between Dietary Fiber Intake and Risk of Coronary Heart Disease: A Meta-Analysis. Clin. Nutr. 2015, 34, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, G.; Liu, D.; Zhu, W.; Fan, X.; Liu, X. Dietary Fiber Consumption and Risk of Stroke. Eur. J. Epidemiol. 2013, 28, 119–130. [Google Scholar] [CrossRef]
- Reynolds, A.; Mann, J.; Cummings, J.; Winter, N.; Mete, E.; Te Morenga, L. Carbohydrate Quality and Human Health: A Series of Systematic Reviews and Meta-Analyses. Lancet 2019, 393, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary Fiber and Health Outcomes: An Umbrella Review of Systematic Reviews and Meta-Analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef]
- Nsor-Atindana, J.; Zhong, F.; Mothibe, K.J.; Bangoura, M.L.; Lagnika, C. Quantification of Total Polyphenolic Content and Antimicrobial Activity of Cocoa (Theobroma cacao L.) Bean Shells. Pak. J. Nutr. 2012, 11, 574–579. [Google Scholar] [CrossRef]
- Braojos, C.; Benitez, V.; Rebollo-Hernanz, M.; Cañas, S.; Aguilera, Y.; Arribas, S.M.; Martin-Cabrejas, M.A. Evaluation of the Hypolipidemic Properties of Cocoa Shell after Simulated Digestion Using in Vitro Techniques and a Cell Culture Model of Non-Alcoholic Fatty Liver Disease. Proceedings 2020, 70, 58. [Google Scholar] [CrossRef]
- Belwal, T.; Cravotto, C.; Ramola, S.; Thakur, M.; Chemat, F.; Cravotto, G. Bioactive Compounds from Cocoa Husk: Extraction, Analysis and Applications in Food Production Chain. Foods 2022, 11, 798. [Google Scholar] [CrossRef] [PubMed]
- Barišić, V.; Stokanović, M.C.; Flanjak, I.; Doko, K.; Jozinović, A.; Babić, J.; Šubarić, D.; Miličević, B.; Cindrić, I.; Ačkar, Đ. Cocoa Shell as a Step Forward to Functional Chocolates—Bioactive Components in Chocolates with Different Composition. Molecules 2020, 25, 5470. [Google Scholar] [CrossRef] [PubMed]
- Rojo-Poveda, O.; Barbosa-Pereira, L.; El Khattabi, C.; Youl, E.N.H.; Bertolino, M.; Delporte, C.; Pochet, S.; Stévigny, C. Polyphenolic and Methylxanthine Bioaccessibility of Cocoa Bean Shell Functional Biscuits: Metabolomics Approach and Intestinal Permeability through Caco-2 Cell Models. Antioxidants 2020, 9, 1164. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, E.; Ova, G. Evaluation of Cocoa Bean Hulls as a Fat Replacer On Functional Cake Production. Turkish J. Agric. Food Sci. Technol. 2018, 6, 1043–1050. [Google Scholar] [CrossRef]
- Choi, J.; Kim, N.; Choi, H.Y.; Han, Y.S. Effect of Cacao Bean Husk Powder on the Quality Properties of Pork. Food Sci. Anim. Resour. 2019, 39, 742. [Google Scholar] [CrossRef]
- Scapagnini, G.; Davinelli, S.; Drago, F.; De Lorenzo, A.; Oriani, G. Antioxidants as Antidepressants: Fact or Fiction? CNS Drugs 2012, 26, 477–490. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, Y.; Sun, X.Z.; Wang, B.Y.; Zhao, Y.; Liu, D.C.; Zhang, D.D.; Liu, X.J.; Zhang, R.Y.; Sun, H.H.; et al. Chocolate Consumption and Risk of Cardiovascular Diseases: A Meta-Analysis of Prospective Studies. Heart 2019, 105, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, S.; Salek, M.; Clark, C.C.T. Cocoa Consumption and Blood Pressure in Middle-Aged and Elderly Subjects: A Meta-Analysis. Curr. Hypertens. Rep. 2020, 22, 1. [Google Scholar] [CrossRef]
- Sun, Y.; Zimmermann, D.; De Castro, C.A.; Actis-Goretta, L. Dose-Response Relationship between Cocoa Flavanols and Human Endothelial Function: A Systematic Review and Meta-Analysis of Randomized Trials. Food Funct. 2019, 10, 6322–6330. [Google Scholar] [CrossRef] [PubMed]
- Martiniuk, A.L.; Lee, C.M.; Lawes, C.M.M.; Ueshima, H.; Suh, I.; Lam, T.H.; Gu, D.; Feigin, V.; Jamrozik, K.; Ohkubo, T.; et al. Hypertension: Its Prevalence and Population-Attributable Fraction for Mortality from Cardiovascular Disease in the Asia-Pacific Region. J. Hypertens. 2007, 25, 73–79. [Google Scholar] [CrossRef]
- Grassi, D.; Ferri, C.; Desideri, G. Brain Protection and Cognitive Function: Cocoa Flavonoids as Nutraceuticals. Curr. Pharm. Des. 2016, 22, 145–151. [Google Scholar] [CrossRef]
- Lamport, D.J.; Christodoulou, E.; Achilleos, C. Beneficial Effects of Dark Chocolate for Episodic Memory in Healthy Young Adults: A Parallel-Groups Acute Intervention with a White Chocolate Control. Nutrients 2020, 12, 483. [Google Scholar] [CrossRef]
- Carrillo, J.Á.; Pilar Zafrilla, M.; Marhuenda, J. Cognitive Function and Consumption of Fruit and Vegetable Polyphenols in a Young Population: Is There a Relationship? Foods 2019, 8, 507. [Google Scholar] [CrossRef]
- Haskell-Ramsay, C.F.; Schmitt, J.; Actis-Goretta, L. The Impact of Epicatechin on Human Cognition: The Role of Cerebral Blood Flow. Nutrients 2018, 10, 986. [Google Scholar] [CrossRef]
- Ezra-Nevo, G.; Henriques, S.F.; Ribeiro, C. The Diet-Microbiome Tango: How Nutrients Lead the Gut Brain Axis. Curr. Opin. Neurobiol. 2020, 62, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Sarriá, B.; Martínez-López, S.; Sierra-Cinos, J.L.; García-Diz, L.; Mateos, R.; Bravo, L. Regular Consumption of a Cocoa Product Improves the Cardiometabolic Profile in Healthy and Moderately Hypercholesterolaemic Adults. Br. J. Nutr. 2014, 111, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.; Meyer-Gerspach, A.C.; Wendebourg, M.J.; Gruber, M.; Heinrich, H.; Sauter, M.; Woelnerhanssen, B.; Koeberle, D.; Juengling, F. Effect of Cocoa on the Brain and Gut in Healthy Subjects: A Randomised Controlled Trial. Br. J. Nutr. 2019, 121, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; Turner, J.R. Cell Biology of Tight Junction Barrier Regulation and Mucosal Disease. Cold Spring Harb. Perspect. Biol. 2018, 10, a029314. [Google Scholar] [CrossRef] [PubMed]
- Rossin, D.; Barbosa-Pereira, L.; Iaia, N.; Sottero, B.; Danzero, A.C.; Poli, G.; Zeppa, G.; Biasi, F. Protective Effect of Cocoa Bean Shell against Intestinal Damage: An Example of Byproduct Valorization. Antioxidants 2021, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.; Yeboah-Awudzi, M.; Magazine, N.; King, J.M.; Xu, Z.; Losso, J.N. Procyanidin B2 Rich Cocoa Extracts Inhibit Inflammation in Caco-2 Cell Model of in Vitro Celiac Disease by down-Regulating Interferon-Gamma- or Gliadin Peptide 31-43-Induced Transglutaminase-2 and Interleukin-15. J. Funct. Foods 2019, 57, 112–120. [Google Scholar] [CrossRef]
- Singh, A.K.; Cabral, C.; Kumar, R.; Ganguly, R.; Rana, H.K.; Gupta, A.; Lauro, M.R.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial Effects of Dietary Polyphenols on Gut Microbiota and Strategies to Improve Delivery Efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [PubMed]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P.E. Prebiotic Evaluation of Cocoa-Derived Flavanols in Healthy Humans by Using a Randomized, Controlled, Double-Blind, Crossover Intervention Study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of Polyphenols on Gut Microbiota and Implications in Human Health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef]
- Ma, G.; Chen, Y. Polyphenol Supplementation Benefits Human Health via Gut Microbiota: A Systematic Review via Meta-Analysis. J. Funct. Foods 2020, 66, 103829. [Google Scholar] [CrossRef]
- Martín-Peláez, S.; Camps-Bossacoma, M.; Massot-Cladera, M.; Rigo-Adrover, M.; Franch, À.; Pérez-Cano, F.J.; Castell, M. Effect of Cocoa’s Theobromine on Intestinal Microbiota of Rats. Mol. Nutr. Food Res. 2017, 61, 1700238. [Google Scholar] [CrossRef] [PubMed]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry Polyphenols Metabolism and Impact on Human Gut Microbiota and Health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; Bashmakov, Y.; Chalyk, N.; Nielsen, D.S.; Krych, Ł.; Kot, W.; Klochkov, V.; Pristensky, D.; Bandaletova, T.; Chernyshova, M.; et al. Prebiotic Effect of Lycopene and Dark Chocolate on Gut Microbiome with Systemic Changes in Liver Metabolism, Skeletal Muscles and Skin in Moderately Obese Persons. BioMed Res. Int. 2019, 2019, 4625279. [Google Scholar] [CrossRef] [PubMed]
- Crichton, G.E.; Elias, M.F.; Dearborn, P.; Robbins, M. Habitual Chocolate Intake and Type 2 Diabetes Mellitus in the Maine-Syracuse Longitudinal Study: (1975–2010): Prospective Observations. Appetite 2017, 108, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.A. Chocolate Intake and Diabetes Risk. Clin. Nutr. 2015, 34, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, C.; Petrone, A.B.; Sesso, H.D.; Gaziano, J.M.; Djoussé, L. Chocolate Consumption and Risk of Diabetes Mellitus in the Physicians’ Health Study. Am. J. Clin. Nutr. 2015, 101, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Oba, S.; Nagata, C.; Nakamura, K.; Fujii, K.; Kawachi, T.; Takatsuka, N.; Shimizu, H. Consumption of Coffee, Green Tea, Oolong Tea, Black Tea, Chocolate Snacks and the Caffeine Content in Relation to Risk of Diabetes in Japanese Men and Women. Br. J. Nutr. 2010, 103, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Maskarinec, G.; Jacobs, S.; Shvetsov, Y.; Boushey, C.J.; Setiawan, V.W.; Kolonel, L.N.; Haiman, C.A.; Le Marchand, L. Intake of Cocoa Products and Risk of Type 2 Diabetes: The Multiethnic Cohort. Eur. J. Clin. Nutr. 2019, 73, 671. [Google Scholar] [CrossRef] [PubMed]
- Jafarirad, S.; Ayoobi, N.; Karandish, M.; Jalali, M.T.; Haghighizadeh, M.; Jahanshahi, A. Dark Chocolate Effect on Serum Adiponectin, Biochemical and Inflammatory Parameters in Diabetic Patients: A Randomized Clinical Trial. Int. J. Prev. Med. 2018, 9, 86. [Google Scholar] [CrossRef]
- Dicks, L.; Kirch, N.; Gronwald, D.; Wernken, K.; Zimmermann, B.F.; Helfrich, H.P.; Ellinger, S. Regular Intake of a Usual Serving Size of Flavanol-Rich Cocoa Powder Does Not Affect Cardiometabolic Parameters in Stably Treated Patients with Type 2 Diabetes and Hypertension—A Double-Blinded, Randomized, Placebo-Controlled Trial. Nutrients 2018, 10, 1435. [Google Scholar] [CrossRef]
- Rynarzewski, J.; Dicks, L.; Zimmermann, B.F.; Stoffel-Wagner, B.; Ludwig, N.; Helfrich, H.P.; Ellinger, S. Impact of a Usual Serving Size of Flavanol-Rich Cocoa Powder Ingested with a Diabetic-Suitable Meal on Postprandial Cardiometabolic Parameters in Type 2 Diabetics—A Randomized, Placebo-Controlled, Double-Blind Crossover Study. Nutrients 2019, 11, 417. [Google Scholar] [CrossRef]
- Davis, D.W.; Tallent, R.; Navalta, J.W.; Salazar, A.; Lyons, T.J.; Basu, A. Effects of Acute Cocoa Supplementation on Postprandial Apolipoproteins, Lipoprotein Subclasses, and Inflammatory Biomarkers in Adults with Type 2 Diabetes after a High-Fat Meal. Nutrients 2020, 12, 1902. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Cilleros, D.; Ramos, S.; Goya, L.; Martín, M.Á. Colonic Metabolites from Flavanols Stimulate Nitric Oxide Production in Human Endothelial Cells and Protect against Oxidative Stress-Induced Toxicity and Endothelial Dysfunction. Food Chem. Toxicol. 2018, 115, 88–97. [Google Scholar] [CrossRef]
- Martin, M.A.; Goya, L.; Ramos, S. Protective Effects of Tea, Red Wine and Cocoa in Diabetes. Evidences from Human Studies. Food Chem. Toxicol. 2017, 109, 302–314. [Google Scholar] [CrossRef]
- Ibero-Baraibar, I.; Perez-Cornago, A.; Ramirez, M.J.; Martínez, J.A.; Zulet, M.A. An Increase in Plasma Homovanillic Acid with Cocoa Extract Consumption Is Associated with the Alleviation of Depressive Symptoms in Overweight or Obese Adults on an Energy Restricted Diet in a Randomized Controlled Trial. J. Nutr. 2015, 146, 897S–904S. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Ismail, A.; Kersten, S. Molecular Mechanisms Underlying the Potential Antiobesity-Related Diseases Effect of Cocoa Polyphenols. Mol. Nutr. Food Res. 2014, 58, 33–48. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Segura-Carretero, A.; del Mar Contreras, M. Phenolic Compounds as Natural and Multifunctional Anti-Obesity Agents: A Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1212–1229. [Google Scholar] [CrossRef]
- Gu, Y.; Yu, S.; Park, J.Y.; Harvatine, K.; Lambert, J.D. Dietary Cocoa Reduces Metabolic Endotoxemia and Adipose Tissue Inflammation in High-Fat Fed Mice. J. Nutr. Biochem. 2014, 25, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Jurinjak Tušek, A.; Šamec, D.; LastNameLastNameLastNameLastNameLastNameŠalić, A. Modern Techniques for Flavonoid Extraction—To Optimize or Not to Optimize? Appl. Sci. 2022, 12, 11865. [Google Scholar] [CrossRef]
- Insang, S.; Kijpatanasilp, I.; Jafari, S.; Assatarakul, K. Ultrasound-Assisted Extraction of Functional Compound from Mulberry (Morus alba L.) Leaf Using Response Surface Methodology and Effect of Microencapsulation by Spray Drying on Quality of Optimized Extract. Ultrason. Sonochem. 2022, 82, 105806. [Google Scholar] [CrossRef]
- Kobus-Cisowska, J.; Szczepaniak, O.; Szymanowska-Powałowska, D.; Piechocka, J.; Szulc, P.; Dziedziński, M. Antioxidant Potential of Various Solvent Extract from Morus Alba Fruits and Its Major Polyphenols Composition Potencial Antioxidante de Vários Extratos de Frutos de Morus Alba e de Sua Composição Majoritária de Polifenóis. Cienc. Rural 2019, 50, 2020. [Google Scholar] [CrossRef]
- Kumar, K.; Srivastav, S.; Sharanagat, V.S. Ultrasound Assisted Extraction (UAE) of Bioactive Compounds from Fruit and Vegetable Processing by-Products: A Review. Ultrason. Sonochem. 2021, 70, 105325. [Google Scholar] [CrossRef] [PubMed]
- Yi Ling, Y.; Sook Fun, P.; Yeop, A.; Yusoff, M.M.; Gimbun, J. Assessment of Maceration, Ultrasonic and Microwave Assisted Extraction for Total Phenolic Content, Total Flavonoid Content and Kaempferol Yield from Cassia Alata via Microstructures Analysis. Mater. Today Proc. 2019, 19, 1273–1279. [Google Scholar] [CrossRef]
- Charpe, T.W.; Rathod, V.K. Effect of Ethanol Concentration in Ultrasound Assisted Extraction of Glycyrrhizic Acid from Licorice Root. Iran. J. Chem. Eng. 2014, 11, 21–30. [Google Scholar]
- Jafari, S.; Karami, Z.; Shiekh, K.A.; Kijpatanasilp, I.; Worobo, R.W.; Assatarakul, K. Ultrasound-Assisted Extraction of Bioactive Compounds from Cocoa Shell and Their Encapsulation in Gum Arabic and Maltodextrin: A Technology to Produce Functional Food Ingredients. Foods 2023, 12, 412. [Google Scholar] [CrossRef]
- Md Yusof, A.H.; Abd Gani, S.S.; Zaidan, U.H.; Halmi, M.I.E.; Zainudin, B.H. Optimization of an Ultrasound-Assisted Extraction Condition for Flavonoid Compounds from Cocoa Shells (Theobroma cacao) Using Response Surface Methodology. Molecules 2019, 24, 711. [Google Scholar] [CrossRef] [PubMed]
- Streimikyte, P.; Viskelis, P.; Viskelis, J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. Int. J. Mol. Sci. 2022, 23, 2359. [Google Scholar] [CrossRef] [PubMed]
- Huynh, G.H.; Van Pham, H.; Hong Nguyen, H.V. Effects of Enzymatic and Ultrasonic-Assisted Extraction of Bioactive Compounds from Cocoa Bean Shells. J. Food Meas. Charact. 2023, 17, 4650–4660. [Google Scholar] [CrossRef]
- Routray, W.; Orsat, V. Microwave Assisted Extraction of Flavonoids: A Comprehensive Overview. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar] [CrossRef]
- Bagade, S.B.; Patil, M. Recent Advances in Microwave Assisted Extraction of Bioactive Compounds from Complex Herbal Samples: A Review. Crit. Rev. Anal. Chem. 2021, 51, 138–149. [Google Scholar] [CrossRef]
- Mellinas, A.C.; Jiménez, A.; Garrigós, M.C. Optimization of Microwave-Assisted Extraction of Cocoa Bean Shell Waste and Evaluation of Its Antioxidant, Physicochemical and Functional Properties. LWT 2020, 127, 109361. [Google Scholar] [CrossRef]
- Chatzimitakos, T.; Athanasiadis, V.; Kalompatsios, D.; Mantiniotou, M.; Bozinou, E.; Lalas, S.I. Pulsed Electric Field Applications for the Extraction of Bioactive Compounds from Food Waste and by-Products: A Critical Review. Biomass 2023, 3, 367–401. [Google Scholar] [CrossRef]
- Gilbert-López, B.; Barranco, A.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Development of New Green Processes for the Recovery of Bioactives from Phaeodactylum Tricornutum. Food Res. Int. 2017, 99, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Pereira, L.; Guglielmetti, A.; Zeppa, G. Pulsed Electric Field Assisted Extraction of Bioactive Compounds from Cocoa Bean Shell and Coffee Silverskin. Food Bioprocess Technol. 2018, 11, 818–835. [Google Scholar] [CrossRef]
- Carpentieri, S.; Režek Jambrak, A.; Ferrari, G.; Pataro, G. Pulsed Electric Field-Assisted Extraction of Aroma and Bioactive Compounds from Aromatic Plants and Food By-Products. Front. Nutr. 2022, 8, 792203. [Google Scholar] [CrossRef] [PubMed]
- Uwineza, P.A.; Waśkiewicz, A. Recent Advances in Supercritical Fluid Extraction of Natural Bioactive Compounds from Natural Plant Materials. Molecules 2020, 25, 3847. [Google Scholar] [CrossRef] [PubMed]
- Wrona, O.; Rafińska, K.; Możeński, C.; Buszewski, B. Supercritical Fluid Extraction of Bioactive Compounds from Plant Materials. J. AOAC Int. 2017, 100, 1624–1635. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Dwivedi, A.; Upadhyayula, S. Supercritical Fluids as Green Solvents. In Handbook of Greener Synthesis of Nanomaterials and Compounds: Volume 1: Fundamental Principles and Methods; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, pp. 891–916. ISBN 9780128219386. [Google Scholar]
- Williams, J.R.; Clifford, A.A.; Al-Saidi, S.H.R. Supercritical Fluids and Their Applications in Biotechnology and Related Areas. Mol. Biotechnol. 2002, 22, 263–286. [Google Scholar] [CrossRef] [PubMed]
- Tripodo, G.; Ibáñez, E.; Cifuentes, A.; Gilbert-López, B.; Fanali, C. Optimization of Pressurized Liquid Extraction by Response Surface Methodology of Goji Berry (Lycium barbarum L.) Phenolic Bioactive Compounds. Electrophoresis 2018, 39, 1673–1682. [Google Scholar] [CrossRef]
- Putra, N.R.; Wibobo, A.G.; Machmudah, S.; Winardi, S. Recovery of Valuable Compounds from Palm-Pressed Fiber by Using Supercritical CO2 Assisted by Ethanol: Modeling and Optimization. Sep. Sci. Technol. 2020, 55, 3126–3139. [Google Scholar] [CrossRef]
- He, J.Z.; Shao, P.; Liu, J.H.; Ru, Q.M. Supercritical Carbon Dioxide Extraction of Flavonoids from Pomelo (Citrus grandis (L.) Osbeck) Peel and Their Antioxidant Activity. Int. J. Mol. Sci. 2012, 13, 13065. [Google Scholar] [CrossRef]
- Melloul, S.; Zehioua, R.; Meniai, A.H. Supercritical CO2 Extraction of Bioactive Compounds from Local Peganum Harmala Plant Seeds and Optimization of the Extraction Yield and the Antioxidant Activities. Sustain. Chem. Pharm. 2022, 28, 100729. [Google Scholar] [CrossRef]
- Pico Hernández, S.M.; Jaimes Estévez, J.; López Giraldo, L.J.; Murillo Méndez, C.J.; Pico Hernández, S.M.; Jaimes Estévez, J.; López Giraldo, L.J.; Murillo Méndez, C.J. Supercritical Extraction of Bioactive Compounds from Cocoa Husk: Study of the Main Parameters. Rev. Fac. Ing. Univ. Antioq. 2019, 91, 95–105. [Google Scholar] [CrossRef]
- Alvarez-Rivera, G.; Bueno, M.; Ballesteros-Vivas, D.; Mendiola, J.A.; Ibañez, E. Pressurized Liquid Extraction. In Liquid-Phase Extraction; Poole, C.F., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 1, pp. 375–398. ISBN 9780128169117. [Google Scholar]
- Raynie, D.E. Extraction. In Encyclopedia of Separation Science; Wilson, I.D., Ed.; Academic Press: Cambridge, UK, 2000; Volume 1, pp. 118–128. ISBN 9780122267703. [Google Scholar]
- Carabias-Martínez, R.; Rodríguez-Gonzalo, E.; Revilla-Ruiz, P.; Hernández-Méndez, J. Pressurized Liquid Extraction in the Analysis of Food and Biological Samples. J. Chromatogr. A 2005, 1089, 1–17. [Google Scholar] [CrossRef]
- Pagliari, S.; Celano, R.; Rastrelli, L.; Sacco, E.; Arlati, F.; Labra, M.; Campone, L. Extraction of Methylxanthines by Pressurized Hot Water Extraction from Cocoa Shell by-Product as Natural Source of Functional Ingredient. LWT 2022, 170, 114115. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Macronutrients per 100 g | Percentage |
|---|---|
| Protein | 15–20% |
| Carbohydrates | ~15% |
| Lipids | 10–25% |
| Fiber | 25–40% |
| Micronutrients per 100 g | Per 100 g |
| Vitamin A (Retinol) | <0.2 mg |
| Vitamin E (Tocopherol) | 2.5 mg |
| Vitamin B1 (Thiamine) | 0.3 mg |
| Vitamin B1 (Riboflavin) | 0.4 mg |
| Vitamin B3 (Niacin) | 0.7 mg |
| Sodium (Na) | 0.03 g |
| Potassium (K) | 4.3 g |
| Calcium (Ca) | 151 mg |
| Phosphorus (P) | 700 mg |
| Iron (Fe) | 26 mg |
| Magnesium (mg) | 555 mg |
| Copper (Cu) | 5 mg |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tušek, K.; Valinger, D.; Jurina, T.; Sokač Cvetnić, T.; Gajdoš Kljusurić, J.; Benković, M. Bioactives in Cocoa: Novel Findings, Health Benefits, and Extraction Techniques. Separations 2024, 11, 128. https://doi.org/10.3390/separations11040128
Tušek K, Valinger D, Jurina T, Sokač Cvetnić T, Gajdoš Kljusurić J, Benković M. Bioactives in Cocoa: Novel Findings, Health Benefits, and Extraction Techniques. Separations. 2024; 11(4):128. https://doi.org/10.3390/separations11040128
Chicago/Turabian StyleTušek, Kristina, Davor Valinger, Tamara Jurina, Tea Sokač Cvetnić, Jasenka Gajdoš Kljusurić, and Maja Benković. 2024. "Bioactives in Cocoa: Novel Findings, Health Benefits, and Extraction Techniques" Separations 11, no. 4: 128. https://doi.org/10.3390/separations11040128