Native Colombian Fruits and Their by-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential

 ,
,  , , and
, , and
Abstract
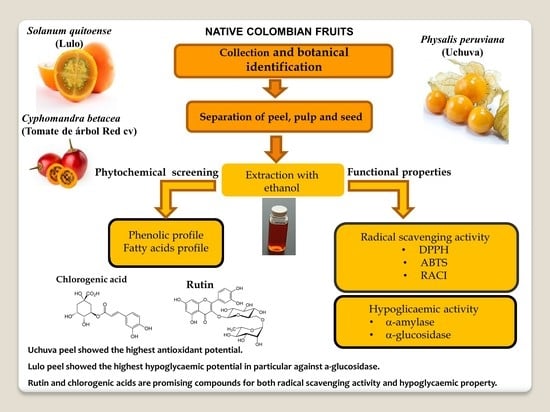
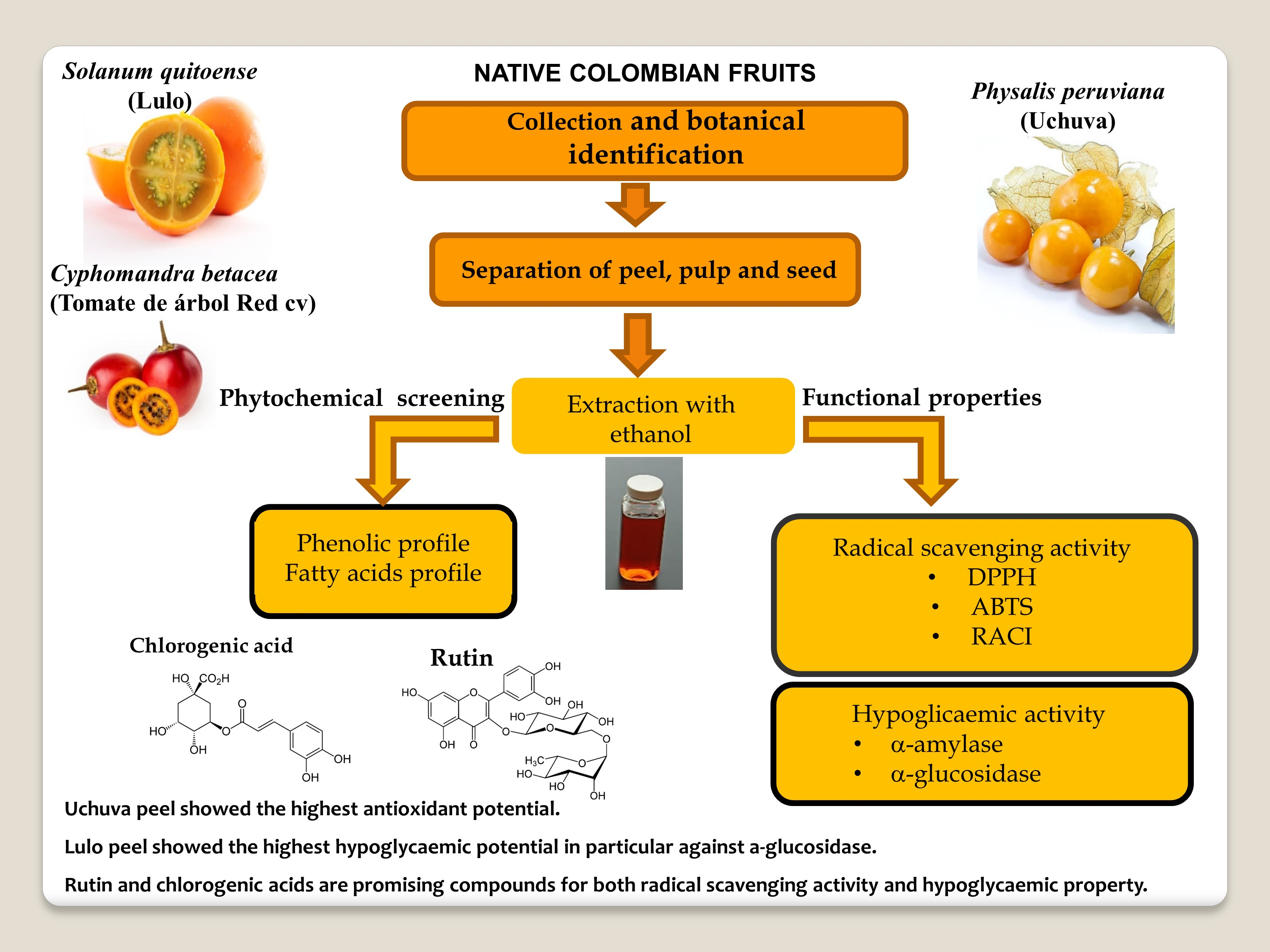
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Material and Extraction Procedure
2.3. Ultra-High Performance Liquid Chromatography (UHPLC)-High Resolution Mass Spectrometry (HRMS). Conditions and Analysis of Phenolic Compounds in Colombian Fruits
2.4. Fatty Acids Profile
2.5. ABTS and DPPH Radical Scavenging Assays
2.6. β-Carotene Bleaching Test
2.7. FRAP (Ferric Reducing Ability Power) Assay
2.8. Relative Antioxidant Capacity Index (RACI) Calculation
2.9. Global Antioxidant Score (GAS)
2.10. a-Amylase and a-Glucosidase Inhibitory Assays
2.11. Statistical Analysis
3. Results and Discussion
3.1. UHPLC-ESI-HRMS Phenolic Profile
3.2. Fatty Acid Composition
3.3. Antioxidant Activity
3.4. Carbohydrate Hydrolysing Enzymes Inhibition by Colombian Fruits Extracts
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federation-Home. Available online: https://www.idf.org/ (accessed on 23 January 2019).
- Montgomery, M.K.; Turner, N. Mitochondrial dysfunction and insulin resistance: An update. Endocr. Connect. 2015, 4, R1–R15. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Bonesi, M.; Nabavi, S.M.; Sobarzo-Sánchez, E.; Rastrelli, L.; Tundis, R. Hypoglycaemic Effects of plants food constituents via inhibition of carbohydrate-hydrolysing enzymes: From chemistry to future applications. Nat. Prod. Target. Clin. Relev. Enzym. 2017, 1, 135–161. [Google Scholar]
- Loizzo, M.R.; Bonesi, M.; Menichini, F.; Tenuta, M.C.; Leporini, M.; Tundis, R. Antioxidant and Carbohydrate-Hydrolysing Enzymes Potential of Sechium edule (Jacq.) Swartz (Cucurbitaceae) Peel, Leaves and Pulp Fresh and Processed. Plant Foods Hum. Nutr. 2016, 71, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Bonesi, M.; Sicari, V.; Pellicanò, T.M.; Tenuta, M.C.; Leporini, M.; Menichini, F.; Loizzo, M.R. Poncirus trifoliata (L.) Raf.: Chemical composition, antioxidant properties and hypoglycaemic activity via the inhibition of alpha-amylase and alpha-glucosidase enzymes. J. Funct. Foods 2016, 25, 477–485. [Google Scholar] [CrossRef]
- Heiser Charles, B., Jr. Some Ecuadorian and Colombian Solanums with edible fruits. [Algunos Solanums Colombianos y Ecuatorianos con frutos comestibles. Cienc. Nat. 1968, 11, 3–9. [Google Scholar]
- Facciola, S. Cornucopia II: A Source Book of Edible Plants; Kampong Publications: Vista, CA, USA, 1998. [Google Scholar]
- Suter, B.; Grob, K.; Pacciarelli, B. Determination of fat content and fatty acid composition through 1-min transesterification in the food sample; principles. Z. Für Leb. Forsch. A 1997, 204, 252–258. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pacetti, D.; Lucci, P.; Núñez, O.; Menichini, F.; Frega, N.G.; Tundis, R. Prunus persica var. platycarpa (Tabacchiera Peach): bioactive compounds and antioxidant activity of pulp, peel and seed ethanolic extracts. Plant Foods Hum. Nutr. 2015, 70, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Todorovic, V.; Milenkovic, M.; Vidovic, B.; Todorovic, Z.; Sobajic, S. Correlation between antimicrobial, antioxidant activity, and polyphenols of alkalized/nonalkalized cocoa powders. J. Food Sci. 2017, 82, 1020–1027. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; Tenuta, M.C.; Menichini, F.; Xiao, J.; Tundis, R. Edible flowers: A rich source of phytochemicals with antioxidant and hypoglycemic properties. J. Agric. Food Chem. 2016, 64, 2467–2474. [Google Scholar] [CrossRef] [PubMed]
- Gancel, A.-L.; Alter, P.; Dhuique-Mayer, C.; Ruales, J.; Vaillant, F. Identifying carotenoids and phenolic compounds in naranjilla (Solanum quitoense Lam. var. Puyo hybrid), an Andean fruit. J. Agric. Food Chem. 2008, 56, 11890–11899. [Google Scholar] [CrossRef] [PubMed]
- Mertz, C.; Gancel, A.-L.; Gunata, Z.; Alter, P.; Dhuique-Mayer, C.; Vaillant, F.; Perez, A.M.; Ruales, J.; Brat, P. Phenolic compounds, carotenoids and antioxidant activity of three tropical fruits. J. Food Compos. Anal. 2009, 22, 381–387. [Google Scholar] [CrossRef]
- Ramakrishnan, Y.; Khoddami, A.; Gannasin, S.P.; Muhammad, K. Tamarillo (Cyphomandra betacea) seed oils as a potential source of essential fatty acid for food, cosmetic and pharmacuetical industries. Acta Hortic. 2013, 1012, 1415–1421. [Google Scholar] [CrossRef]
- Hassan, A.; Hawa, S.; Bakar, A.; Fadzelly, M. Antioxidative and anticholinesterase activity of Cyphomandra betacea fruit. Sci. World J. 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Lin, S.; Jiang, Y.; Ashraf, M. Variation in contents of total phenolics and flavonoids and antioxidant activities in the leaves of 11 Eriobotrya species. Plant Foods Hum. Nutr. 2008, 63, 200. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademosun, A.O.; Ayeni, P.O.; Omojokun, O.S.; Bello, F. Comparative effect of quercetin and rutin on α-amylase, α-glucosidase, and some pro-oxidant-induced lipid peroxidation in rat pancreas. Comp. Clin. Pathol. 2015, 24, 1103–1110. [Google Scholar] [CrossRef]
- Oboh, G.; Agunloye, O.M.; Adefegha, S.A.; Akinyemi, A.J.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-H.; Hsu, C.-H.; Ng, L.-T. Inhibitory potential of fatty acids on key enzymes related to type 2 diabetes. Biofactors 2013, 39, 415–421. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| (+)-Catechin | Chlorogenic Acid | trans-Cinnamic Acid | p-Coumaric Acid | (−)-Epicatechin | Ferulic Acid | Gallic Acid | Homogentisic Acid | Polydatin | Rutin Hydrate | Sinapic Acid | Syringaldehyde | Taxifolin | Vanillic Acid | Vanillin | Veratric Acid | ⅀Phenolic | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Solanaceae | |||||||||||||||||
| S. quitoense | |||||||||||||||||
| Peel | n.d. | 98.6 ± 2.9 | n.d. | 0.4 ± 0.0 | n.d. | n.d. | 1.3 ± 0.0 | n.d. | n.d. | 51.1 ± 3.6 | n.d. | n.d. | 0.3 ± 0.0 | n.d. | n.d. | n.d. | 151.6 ± 2.5 |
| Pulp + Seeds | n.d. | 19.1 ± 1.0 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.1 ± 0.0 | n.d. | n.d. | n.d. | 20.0 ± 1.0 |
| P. peruviana | |||||||||||||||||
| Peel | n.d. | 1.5 ± 0.1 | n.d. | n.d. | n.d. | n.d. | 0.9 ± 0.0 | n.d. | 0.6 ± 0.03 | 0.1 ± 0.0 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 3.2 ± 0.2 |
| Pulp + Seeds | n.d. | 0.7 ± 0.1 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.6 ± 0.07 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.4 ± 0.2 |
| C. betacea | |||||||||||||||||
| Peel | n.d. | 253.8 ± 3.8 | n.d. | 0.2 ± 0.0 | n.d. | 8.7 ± 0.2 | n.d. | n.d. | n.d. | 9.5 ± 1.4 | 10.3 ± 0.7 | 0.7 ± 0.0 | 0.2 ± 0.0 | n.d. | n.d. | n.d. | 284.1 ± 1.2 |
| Pulp | n.d. | 125.5 ± 1.2 | 28.7 ± 2.0 | 0.2 ± 0.0 | n.d. | 7.6 ± 0.1 | n.d. | n.d. | n.d. | n.d. | 1.6 ± 0.2 | n.d. | 0.1 ± 0.0 | n.d. | n.d. | n.d. | 165.1 ± 2.2 |
| Seeds | 0.8 ± 0.2 | 37.7 ± 1.3 | n.d. | n.d. | 2.5 ± 0.1 | n.d. | n.d. | n.d. | n.d. | 0.7 ± 0.0 | n.d. | n.d. | 0.1 ± 0.0 | n.d. | n.d. | n.d. | 42.1 ± 1.2 |
| Passifloracea | |||||||||||||||||
| P. pinnatistipula | |||||||||||||||||
| Peel | n.d. | n.d. | 132.4 ± 5.1 | 1.5 ± 0.0 | 2.4 ± 0.2 | 4.6 ± 0.5 | 8.1 ± 0.2 | n.d. | 1.1 ± 0.03 | 4.7 ± 0.2 | 9.4 ± 0.1 | n.d. | 1.2 ± 0.0 | n.d. | n.d. | n.d. | 166.2 ± 3.2 |
| Pulp | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 0.5 ± 0.02 | 0.4 ± 0.1 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | 1.0 ± 0.1 |
| Seeds | n.d. | n.d. | n.d. | 0.3 ± 0.0 | 8.1 ± 0.8 | n.d. | n.d. | n.d. | 3.4 ± 0.22 | n.d. | n.d. | n.d. | 0.2 ± 0.0 | n.d. | n.d. | n.d. | 12.4 ± 0.3 |
| P. tripartita | |||||||||||||||||
| Peel | 40.1 ± 2.0 | n.d. | n.d. | 2.0 ± 0.1 | n.d. | 47.8 ± 1.8 | n.d. | n.d. | 0.7 ± 0.05 | n.d. | 10.2 ± 0.9 | n.d. | n.d. | 0.5 ± 0.0 | n.d. | n.d. | 101.9 ± 1.3 |
| Pulp | 140.9 ± 8.2 | n.d. | n.d. | 1.8 ± 0.2 | 7.9 ± 0.5 | 16.3 ± 1.4 | n.d. | n.d. | 1.9 ± 0.27 | n.d. | 211.4 ± 1.5 | 0.8 ± 0.0 | n.d. | 1.3 ± 0.0 | n.d. | n.d. | 383.3 ± 1.3 |
| Seeds | 208.8 ± 3.6 | n.d. | n.d. | n.d. | 81.5 ± 3.5 | n.d. | n.d. | n.d. | 0.7 ± 0.03 | n.d. | 55.3 ± 4.1 | 2.2 ± 0.1 | 2.1 ± 0.1 | n.d. | n.d. | 42.6 ± 1.7 | 394.2 ± 2.6 |
| P. ligularis | |||||||||||||||||
| Peel | 257.6 ± 4.2 | 0.2 ± 0.0 | n.d. | 1.6 ± 0.1 | n.d. | 11.9 ± 1.9 | 2.4 ± 0.3 | 8.95 ± 1.1 | 2.1 ± 0.08 | n.d. | 4.0 ± 0.2 | 2.6 ± 0.1 | n.d. | n.d. | 5.2 ± 0.1 | n.d. | 297.5 ± 1.4 |
| Pulp + Seeds | n.d. | n.d. | n.d. | 0.7 ± 0.1 | n.d. | n.d. | n.d. | n.d. | 1.7 ± 0.04 | 0.3 ± 0.0 | n.d. | n.d. | n.d. | n.d. | 1.8 ± 0.0 | n.d. | 4.4 ± 0.9 |
| Fatty Acids | S. Quitoense | P. Peruviana | C. Betacea | P. Pinnatistipula | P. Tripartita | P. Ligularis | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Peel | Pulp + Seed | Peel | Pulp + Seed | Peel | Pulp | Seed | Peel | Pulp | Seed | Peel | Pulp | Seed | Peel | Pulp + Seed | |
| C16:0 | 22.6 ± 0.1 | 25.6 ± 0.4 | 20.6 ± 2.6 | 19.5 ± 0.7 | 22.9 ± 0.4 | 19.8 ± 0.1 | 13.6 ± 0.0 | 15.4 ± 0.1 | 15.6 ± 0.1 | 11.8 ± 0.0 | 24.0 ± 0.2 | 19.1 ± 0.0 | 9.6 ± 0.0 | 34.0 ± 0.2 | 7.8 ± 0.0 |
| C18:0 | 6.3 ± 0.2 | 4.7 ± 0.2 | 2.7 ± 0.1 | 2.4 ± 0.1 | 2.6 ± 0.2 | 1.4 ± 0.1 | 3.3 ± 0.1 | 2.2 ± 0.0 | 2.9 ± 0.1 | 1.9 ± 0.0 | n.d. | 25.0 ± 0.1 | 1.9 ± 0.0 | 2.1 ± 0.0 | 2.5 ± 0.0 |
| C20:0 | n.d. | n.d. | 5.4 ± 0.5 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| SFA | 28.9 ± 0.1 | 30.3 ± 0.6 | 28.8 ± 2.1 | 22.0 ± 0.5 | 25.4 ± 0.2 | 21.5 ± 0.2 | 16.9 ± 0.1 | 17.6 ± 0.1 | 18.5 ± 0.2 | 13.6 ± 0.0 | 24.0 ± 0.2 | 44.1 ± 0.1 | 11.5 ± 0.0 | 36.2 ± 0.2 | 10.3 ± 0.1 |
| C16:1 | 1.6 ± 0.1 | n.d. | 0.6 ± 0.0 | 1.4 ± 0.1 | n.d. | 1.8 ± 0.1 | 0.7 ± 0.0 | 0.4 ± 0.0 | 4.7 ± 0.1 | 0.2 ± 0.0 | n.d. | n.d. | n.d. | n.d. | 0.2 ± 0.0 |
| C18:1 9 | 12.7 ± 0.3 | 17.0 ± 0.1 | 26.5 ± 0.5 | 20.5 ± 0.5 | 41.4 ± 0.3 | 39.1 ± 0.2 | 17.2 ± 0.6 | 11.3 ± 0.0 | 7.5 ± 0.1 | 10.0 ± 0.0 | 5.7 ± 0.3 | n.d. | 10.8 ± 0.1 | 3.3 ± 0.0 | 15.1 ± 0.3 |
| C18:1 11 | 3.6 ± 0.3 | n.d. | 6.4 ± 0.1 | 8.4 ± 0.4 | 2.3 ± 0.1 | 1.2 ± 0.1 | 1.4 ± 0.1 | 1.2 ± 0.0 | 8.1 ± 0.1 | 0.2 ± 0.0 | n.d. | n.d. | n.d. | 0.9 ± 0.1 | 0.3 ± 0.0 |
| MUFA | 18.0 ± 0.7 | 17.0 ± 0.1 | 33.5 ± 0.6 | 30.3 ± 1.0 | 43.7 ± 0.4 | 41.7 ± 0.4 | 19.3 ± 0.8 | 13.0 ± 0.1 | 20.3 ± 0.2 | 10.4 ± 0.0 | 5.7 ± 0.3 | n.d. | 10.8 ± 0.1 | 4.2 ± 0.1 | 15.6 ± 0.4 |
| C18:2 n-6 | 11.5 ± 0.1 | 23.5 ± 0.9 | 3.6 ± 0.1 | 11.9 ± 0.1 | 10.4 ± 0.1 | 13.1 ± 0.2 | 58.3 ± 1.0 | 63.9 ± 0.3 | 41.0 ± 0.4 | 75.5 ± 1.4 | 39.9 ± 0.3 | 16.6 ± 0.0 | 76.6 ± 0.3 | 36.0 ± 0.1 | 73.3 ± 0.7 |
| C18:3 n-6 | n.d. | n.d. | 4.3 ± 0.1 | 4.4 ± 0.7 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| C18:3 n-3 | 41.6 ± 0.9 | 29.2 ± 0.5 | 29.8 ± 1.3 | 31.4 ± 1.0 | 20.5 ± 0.4 | 23.6 ± 0.7 | 5.5 ± 0.1 | 5.5 ± 0.0 | 20.2 ± 0.0 | 0.5 ± 0.0 | 30.4 ± 0.2 | 39.3 ± 0.1 | 1.0 ± 0.0 | 23.6 ± 0.0 | 0.8 ± 0.0 |
| PUFA | 53.1 ± 0.8 | 52.7 ± 0.5 | 37.7 ± 1.5 | 47.7 ± 0.4 | 30.9 ± 0.3 | 36.8 ± 0.5 | 63.8 ± 0.9 | 69.4 ± 0.2 | 61.2 ± 0.4 | 76.0 ± 0.0 | 70.4 ± 0.1 | 55.9 ± 0.1 | 77.7 ± 0.1 | 59.6 ± 0.1 | 74.1 ± 1.4 |
| Title | Sample | DPPH (IC50 μg/mL) | ABTS (IC50 μg/mL) | β-Carotene Bleacing Test (IC50 μg/mL) | FRAP (μM Fe(II)/g) | RACI | GAS | α-Amylase (IC50 μg/mL) | α-Glucosidase (IC50 μg/mL) |
|---|---|---|---|---|---|---|---|---|---|
| Solanaceae | |||||||||
| S. quitoense | Peel | 38.8 ± 2.1 **** | 167.6 ± 3.7 **** | 11.1 ± 1.3 **** | 49.4 ± 1.5 **** | −0.43 | 0.81 | 31.8 ± 1.2 **** | 27.9 ± 1.0 |
| Pulp + Seed | 61.3 ± 2.0 **** | 576.8 ± 7.5 **** | 6.9 ± 0.5 | 16.2 ± 2.5 **** | −0.38 | 0.87 | 54.9 ± 2.4 | 57.1 ± 2.0 **** | |
| P. peruviana | Peel | 117.9 ± 5.1 **** | 843.3 ± 3.9 **** | 10.2 ± 1.1 *** | 13.0 ± 0.8 **** | 0.20 | 1.72 | 34.1 ± 2.2 **** | 37.6 ± 2.5 |
| Pulp + Seed | 65.3 ± 2.1 **** | > 1000 | 19.3 ± 1.6 **** | 7.3 ± 0.6 **** | −0.03 | 1.40 | 64.3 ± 3.0 **** | 45.0 ± 1.1 | |
| C. betacea | Peel | 74.7 ± 3.5 **** | 149.8 ± 3.6 **** | 21.9 ± 1.5 **** | 63.9 ± 3.5 | 0.05 | 1.52 | 77.1 ± 3.3 **** | 32.9 ± 2.9 ** |
| Pulp | 141.3 ± 5.5 **** | 463.8 ± 4.7 **** | 93.8 ± 4.5 **** | 9.8 ± 0.6 **** | 0.68 | 2.42 | 92.7 ± 3.8 **** | 95.1 ± 4.6 **** | |
| Seed | 57.9 ± 1.5 **** | 329.8 ± 3.3 **** | 58.2 ± 4.0 **** | 25.4 ± 2.5 **** | −0.09 | 1.30 | 102.9 ± 4.0 **** | 195.1 ± 4.7 **** | |
| Passifloraceae | |||||||||
| P. pinnatistipula | Peel | 207.9 ± 2.5 **** | 125.3 ± 1.5 **** | >1000 | 40.5 ± 1.4 **** | 0.23 | 1.47 | 46.4 ± 2.2 | 37.7 ± 2.2 |
| Pulp | 671.9 ± 5.7 **** | 151.7 ± 1.7 **** | 50.6 ± 2.5 **** | 3.2 ± 0.7 **** | 0.11 | 1.15 | 78.2 ± 3.7 **** | 44.1 ± 2.3 | |
| Seed | 372.2 ± 4.0 **** | >1000 | 133.8 ± 2.7 **** | 28.7 ± 2.3 **** | 0.59 | 1.74 | 54.1 ± 2.5 | 40.6 ± 2.8 | |
| P. tripartita | Peel | 3.9 ± 0.8 | 177.8 ± 3.2 **** | >1000 | 22.7 ± 2.2 **** | 0.03 | 1.18 | 86.3 ± 3.0 **** | 56.1 ± 3.1 **** |
| Pulp | 3.8 ± 0.5 | 50.1 ± 2.5 **** | 3.8 ± 0.3 **** | 64.1 ± 3.4 | −0.59 | 0.62 | 67.5 ± 3.7 **** | 78.4 ± 3.9 **** | |
| Seed | 3.2 ± 0.2 | 96.2 ± 3.7 **** | 14.8 ± 0.7 **** | 92.0 ± 3.7 **** | −0.49 | 0.27 | 52.7 ± 3.5 | 54.6 ± 3.2 **** | |
| P. ligularis | Peel | 61.3 ± 2.2 **** | 282.1 ± 4.0 **** | 265.1 ± 4.0 **** | 39.9 ± 4.3 **** | −0.22 | 0.69 | 122.7 ± 4.5 **** | 154.9 ± 3.9 **** |
| Pulp + Seed | 73.9 ± 2.7 **** | 223.9 ± 3.7 **** | 116.0 ± 3.1 **** | 42.9 ± 3.8 **** | 0.35 | 1.40 | 22.6 ± 3.7 **** | 24.8 ± 3.9 ** | |
| Positive controls | Propyl gallate a | 1.0 ± 0.01 | |||||||
| Ascorbic acid a | 2.0 ± 0.01 | 1.7 ± 0.8 | |||||||
| BHT a | 63.2 ± 4.5 | ||||||||
| Acarbose a | 50.0 ± 0.9 | 35.5 ± 1.2 | |||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loizzo, M.R.; Lucci, P.; Núñez, O.; Tundis, R.; Balzano, M.; Frega, N.G.; Conte, L.; Moret, S.; Filatova, D.; Moyano, E.; et al. Native Colombian Fruits and Their by-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential. Foods 2019, 8, 89. https://doi.org/10.3390/foods8030089
Loizzo MR, Lucci P, Núñez O, Tundis R, Balzano M, Frega NG, Conte L, Moret S, Filatova D, Moyano E, et al. Native Colombian Fruits and Their by-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential. Foods. 2019; 8(3):89. https://doi.org/10.3390/foods8030089
Chicago/Turabian StyleLoizzo, Monica Rosa, Paolo Lucci, Oscar Núñez, Rosa Tundis, Michele Balzano, Natale Giuseppe Frega, Lanfranco Conte, Sabrina Moret, Daria Filatova, Encarnación Moyano, and et al. 2019. "Native Colombian Fruits and Their by-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential" Foods 8, no. 3: 89. https://doi.org/10.3390/foods8030089