
Biotechnological Processing of Sugarcane Bagasse through Solid-State Fermentation with White Rot Fungi into Nutritionally Rich and Digestible Ruminant Feed
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. White-Rot Fungi Species and Grain Spawn Preparation
2.2. Substrate Preparation, Pasteurization and Mycobag Processing
2.3. Chemical Analysis
2.4. In Vitro Dry Matter Digestibility
2.5. In Vitro Gas Production
2.6. Statistical Analysis
3. Results
3.1. Changes in Chemical Composition of Sugarcane Bagasse Bioprocessed with WRF Species for Different Treatment Periods
3.2. Dry Matter and Nutrient Losses during Bioprocessing of Sugarcane Bagasse with Four WRF Species
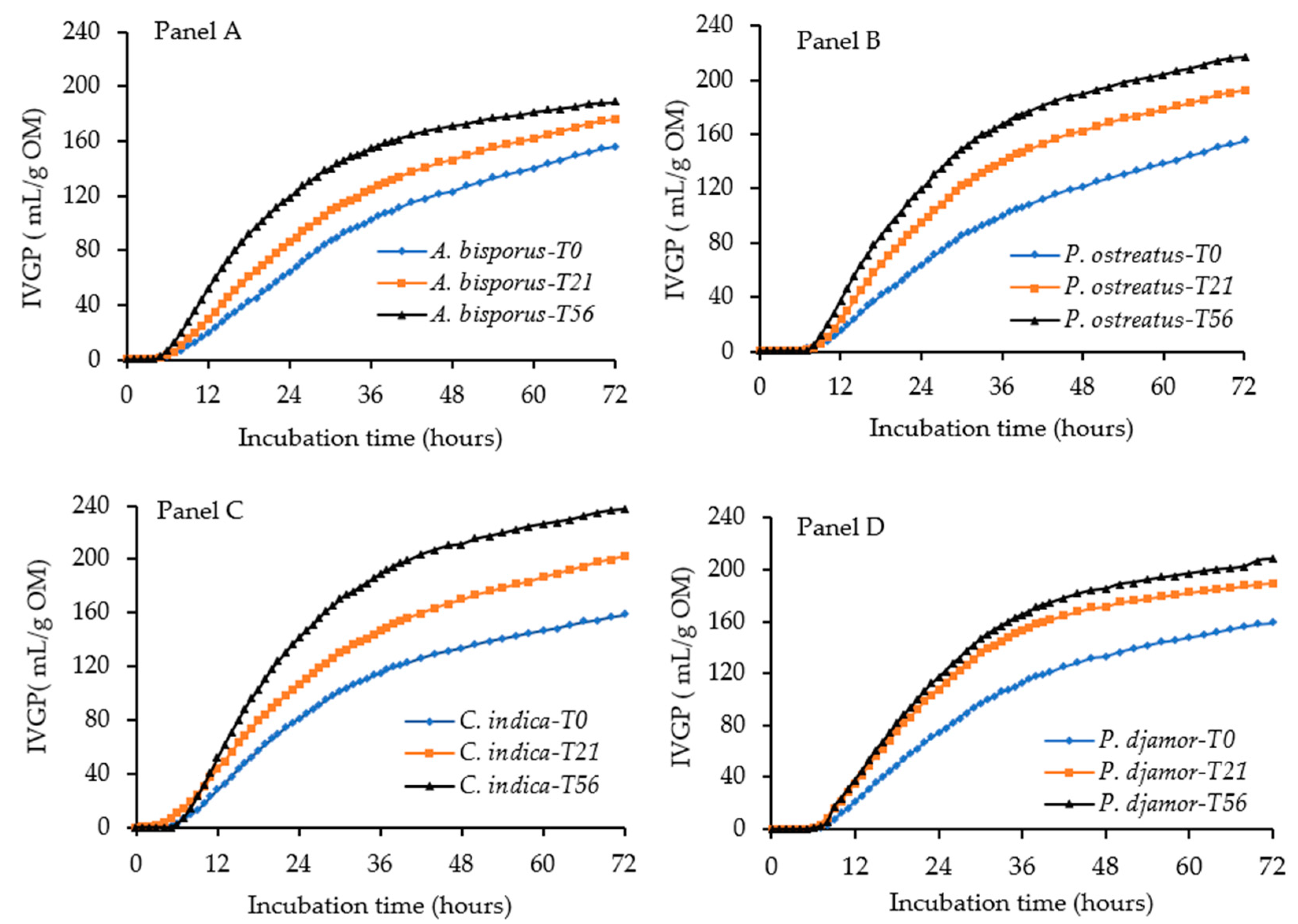
3.3. Effect of Bioprocessing of Sugarcane Bagasse with Different WRF Species on In Vitro Dry Matter Digestibility, In Vitro Total Gas and CH4 Production
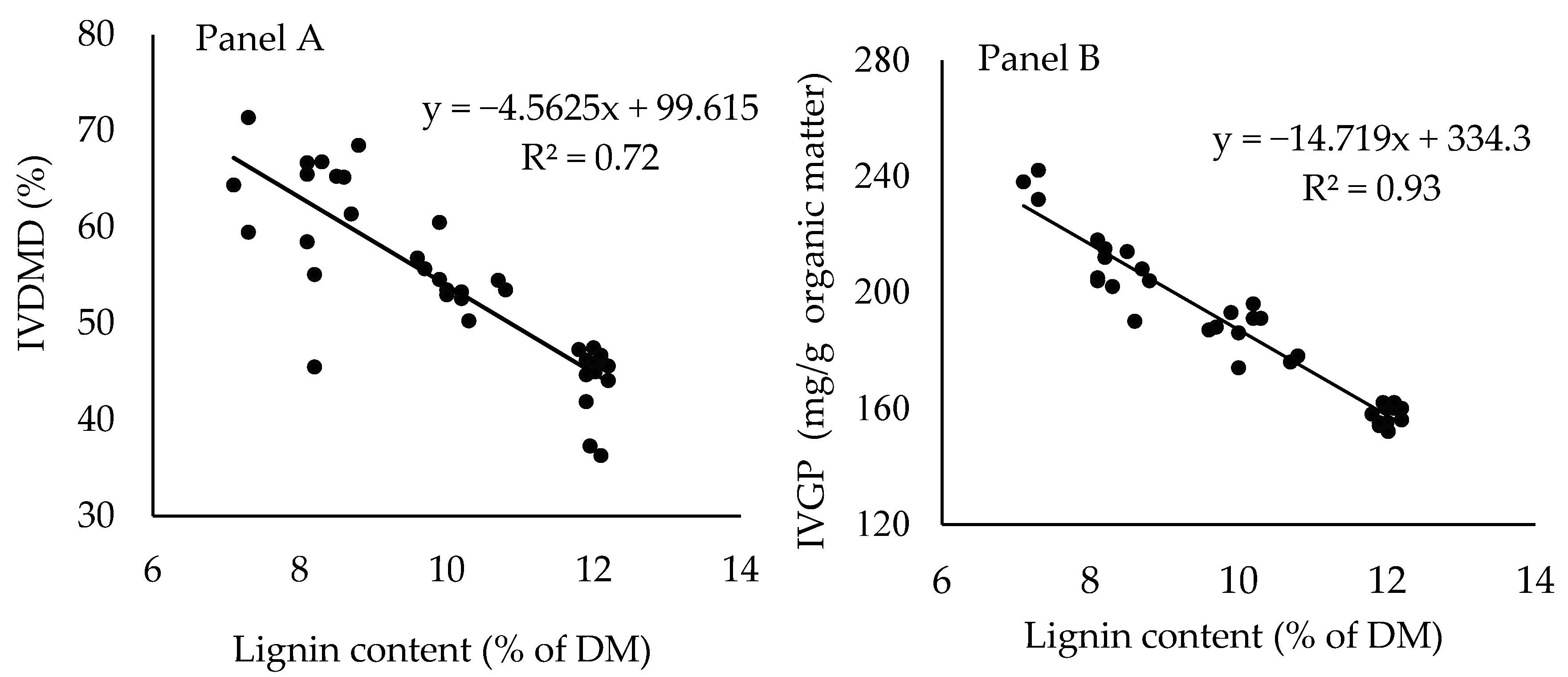
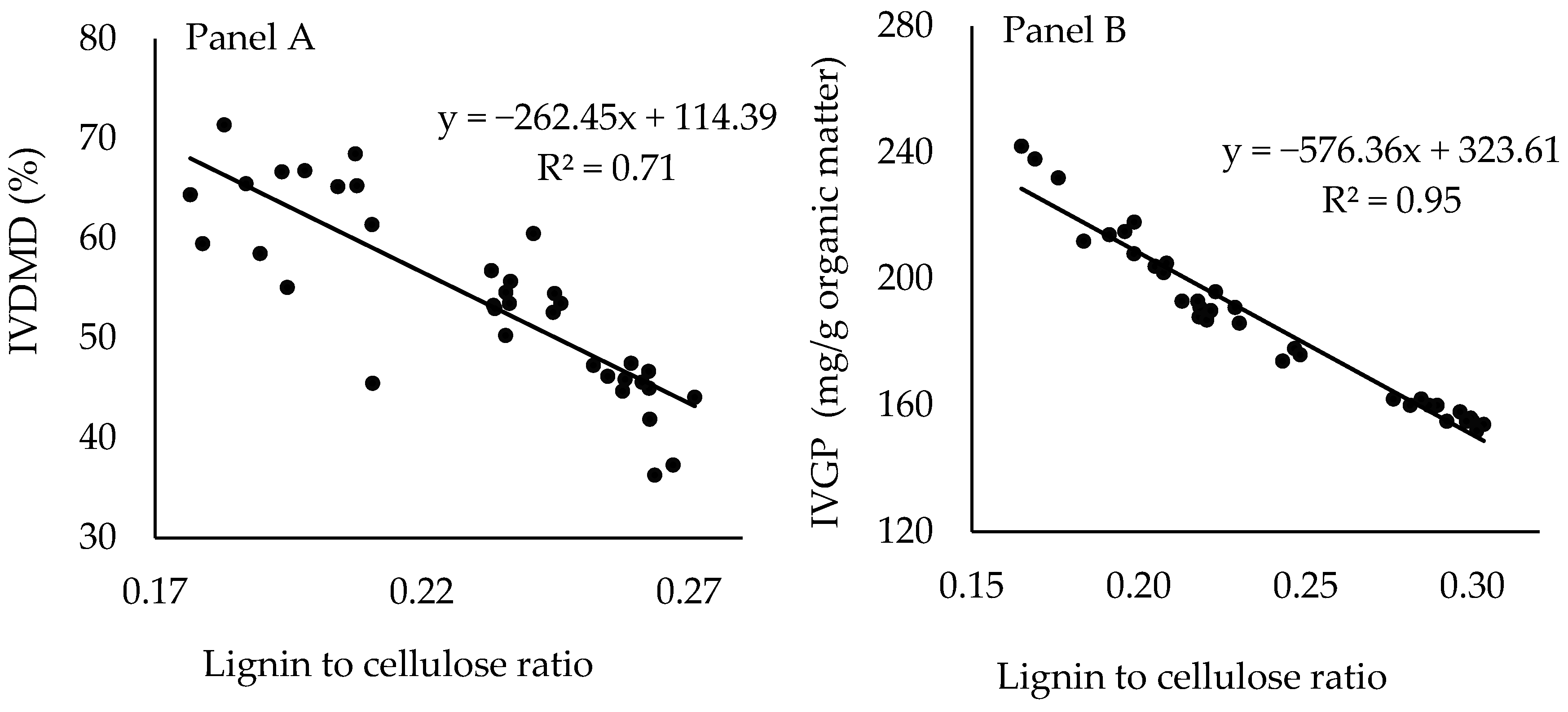
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liang, J.; Nabi, M.; Zhang, P.; Zhang, G.; Cai, Y.; Wang, Q.; Zhou, Z.; Ding, Y. Promising biological conversion of lignocellulosic biomass to renewable energy with rumen microorganisms: A comprehensive review. Renew. Sust. Energ. Rev. 2020, 134, 110335. [Google Scholar] [CrossRef]
- Kainthola, J.; Podder, A.; Fechner, M.; Goel, R. An overview of fungal pretreatment processes for anaerobic digestion: Applications, bottlenecks and future needs. Bioresour. Technol. 2021, 321, 124397. [Google Scholar] [CrossRef]
- Sufyan, A.; Khan, N.A.; AbuGhazaleh, A.; Ahmad, N.; Tang, S.; Tan, Z. Novel techniques for the mass production of nutritionally improved fungus-treated lignocellulosic biomass for ruminant nutrition. J. Sci. Food Agric. 2024, 104, 2215–2224. [Google Scholar] [CrossRef]
- Van Zanten, H.H.E.; Van Ittersum, M.K.; De Boer, I.J.M. The role of farm animals in a circular food system. Glob. Food Secur. 2019, 21, 18–22. [Google Scholar] [CrossRef]
- FAOSTAT. Crops and Livestock Products. Food and Agricluture Organization of the United Nations, Rome. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 24 February 2024).
- Ungureanu, N.; Vladut, V.; Biris, S.S. Sustainable valorization of waste and by-products from sugarcane processing. Sustainability 2022, 14, 11089. [Google Scholar] [CrossRef]
- Parameswaran, B. Sugarcane bagasse. In Biotechnology for Agro-Industrial Residues Utilisation: Utilisation of Agro-Residues; Nigam, P.S., Pandey, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 239–252. [Google Scholar]
- da Costa, D.A.; de Souza, C.L.; Saliba, E.; Carneiro, J. By-products of sugar cane industry in ruminant nutrition. Int. J. Adv. Agric. Res. 2015, 3, 1–95. [Google Scholar]
- Gunun, N.; Wanapat, M.; Gunun, P.; Cherdthong, A.; Khejornsart, P.; Kang, S. Effect of treating sugarcane bagasse with urea and calcium hydroxide on feed intake, digestibility, and rumen fermentation in beef cattle. Trop. Anim. Health Prod. 2016, 48, 1123–1128. [Google Scholar] [CrossRef]
- Ding, G.; Chang, Y.; Zhou, Z.; Ren, L.; Meng, Q. Effect of Saccharomyces cerevisiae on rumen fermentation characteristics, nutrient degradation and cellulase activity of steers fed diets with different concentrate to forage ratios. World J. Agric. Res. 2014, 2, 303–308. [Google Scholar] [CrossRef]
- Sufyan, A.; Ahmad, N.; Shahzad, F.; Embaby, M.G.; AbuGhazaleh, A.; Khan, N.A. Improving the nutritional value and digestibility of wheat straw, rice straw and corn cob through solid state fermentation using different Pleurotus species. J. Sci. Food Agric. 2022, 102, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Van Kuijk, S.; Sonnenberg, A.; Baars, J.; Hendriks, W.; Cone, J. Fungal treated lignocellulosic biomass as ruminant feed ingredient: A review. Biotechnol. Adv. 2015, 33, 191–202. [Google Scholar] [CrossRef]
- Van Kuijk, S.J.A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; Cone, J.W. Fungal treatment of lignocellulosic biomass: Importance of fungal species, colonization and time on chemical composition and in vitro rumen degradability. Anim. Feed Sci. Technol. 2015, 209, 40–50. [Google Scholar] [CrossRef]
- Nayan, N.; Sonnenberg, A.S.; Hendriks, W.H.; Cone, J.W. Prospects and feasibility of fungal pretreatment of agricultural biomass for ruminant feeding. Anim. Feed Sci. Technol. 2020, 268, 114577. [Google Scholar] [CrossRef]
- Tuyen, D.V.; Phuong, H.N.; Cone, J.W.; Baars, J.J.; Sonnenberg, A.S.; Hendriks, W.H. Effect of fungal treatments of fibrous agricultural by-products on chemical composition and in vitro rumen fermentation and methane production. Bioresour. Technol. 2013, 129, 256–263. [Google Scholar] [CrossRef]
- Khan, N.A.; Hussain, S.; Ahmad, N.; Alam, S.; Bezabhi, M.; Hendriks, W.H.; Yu, P.; Cone, J.W. Improving the feeding value of straws with Pleurotus ostreatus. Anim. Prod. Sci. 2015, 55, 241–245. [Google Scholar] [CrossRef]
- Nayan, N.; van Erven, G.; Kabel, M.A.; Sonnenberg, A.S.; Hendriks, W.H.; Cone, J.W. Improving ruminal digestibility of various wheat straw types by white-rot fungi. J. Sci. Food Agric. 2019, 99, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Zuo, S.; Ren, J.; Li, C.; Zheng, M.; Jiang, D.; Xu, C. Effect of wheat straw types on biological delignification and in vitro rumen degradability of wheat straws during treatment with Irpex lacteus. Anim. Feed Sci. Technol. 2020, 267, 114558. [Google Scholar] [CrossRef]
- Datsomor, O.; Yan, Q.; Wang, K.; Mohamed, S.; Opoku-Mensah, L.; Zhao, G.; Miao, L. Effect of ammoniated and/or basidiomycete white-rot fungi treatment on rice straw proximate composition, cell wall component, and in vitro rumen fermentation characteristics. Fermentation 2022, 8, 228. [Google Scholar] [CrossRef]
- Khan, N.A.; Sufyan, A.; Nazar, M.; Yu, P.; Tang, S.; Liu, Y.; Tan, Z. Promising biological conversion of lignocellulosic biomass to digestible ruminant feed with white rot fungi: A comprehensive review. Biotech. Advan. 2024; in press. [Google Scholar]
- Subbiah, K.A.; Balan, V. A comprehensive review of tropical milky white mushroom (Calocybe indica P&C). Mycobiology 2015, 43, 184–194. [Google Scholar] [PubMed]
- Khalkho, S.; Koreti, D.; Kosre, A.; Jadhav, S.; Chandrawanshi, N.K. Review on production technique and nutritional status of Calocybe indica (P&C). NewBioWorld 2021, 3, 184–194. [Google Scholar]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef]
- Cohen, R.; Persky, L.; Hadar, Y. Biotechnological applications and potential of wood-degrading mushrooms of the genus Pleurotus. Appl. Microbiol. Biotechnol. 2002, 58, 582–594. [Google Scholar] [CrossRef]
- Nayan, N.; Sonnenberg, A.S.M.; Hendriks, W.H.; Cone, J.W. Screening of white-rot fungi for bioprocessing of wheat straw into ruminant feed. Appl. Microbiol. 2018, 125, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Khonkhaeng, B.; Cherdthong, A. Improving nutritive value of purple field corn residue and rice straw by culturing with white-rot fungi. J. Fungi 2020, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.; Robertson, J.; Lewis, B. Symposium: Carbohydrate methodology, metabolism, and nutritional implications in dairy cattle. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Tang, S.; Jin, Y.; Yang, C.; He, L.; Wang, J.; Chen, Y. Multiplex SERS-based lateral flow immunosensor for the detection of major mycotoxins in maize utilizing dual Raman labels and triple test lines. J. Hazard. Mater. 2020, 393, 122348. [Google Scholar] [CrossRef] [PubMed]
- Günal, M.; Pinski, B.; AbuGhazaleh, A.A. Evaluating the effects of essential oils on methane production and fermentation under in vitro conditions. Ital. J. Anim. Sci. 2017, 16, 500–506. [Google Scholar] [CrossRef]
- Hatakka, A.; Hammel, K.E. Fungal biodegradation of lignocelluloses. In Industrial Applications; Hofrichter, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 319–340. [Google Scholar]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-degrading enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Li, Y. Fungal pretreatment of lignocellulosic biomass. Biotechnol. Adv. 2012, 30, 1447–1457. [Google Scholar] [CrossRef] [PubMed]
- Tuyen, V.D.; Cone, J.W.; Baars, J.J.; Sonnenberg, A.S.; Hendriks, W.H. Fungal strain and incubation period affect chemical composition and nutrient availability of wheat straw for rumen fermentation. Bioresour. Technol. 2012, 111, 336–342. [Google Scholar] [CrossRef]
- Van Kuijk, S.J.; Sonnenberg, A.S.; Baars, J.J.; Hendriks, W.H.; Cone, J.W. The effect of particle size and amount of inoculum on fungal treatment of wheat straw and wood chips. J. Anim. Sci. Biotechnol. 2016, 7, 39. [Google Scholar] [CrossRef]
- Van Kuijk, S.J.A.; José, C.; Rencoret, J.; Gutiérrez, A.; Sonnenberg, A.S.; Baars, J.J.; Hendriks, W.H.; Cone, J.W. Selective ligninolysis of wheat straw and wood chips by the white-rot fungus Lentinula edodes and its influence on in vitro rumen degradability. J. Anim. Sci. Biotechnol. 2016, 7, 55. [Google Scholar] [CrossRef]
- Datta, S.; Banerjee, G.; Mitra, A.K. Effect of different agricultural waste substrates on the biological efficiency and therapeutic value of Calocybe Indica. Int. J. Pharm. Biol. Sci. 2019, 9, 1157–1168. [Google Scholar]
- Nayan, N.; van Erven, G.; Kabel, M.A.; Sonnenberg, A.S.; Hendriks, W.H.; Cone, J.W. Evaluation of fungal degradation of wheat straw cell wall using different analytical methods from ruminant nutrition perspective. J. Sci. Food Agric. 2019, 99, 4054–4062. [Google Scholar] [CrossRef] [PubMed]
- Van Kuijk, S.J.A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; del Río, J.C.; Rencoret, J.; Gutiérrez, A.; de Ruijter, N.C.A.; Cone, J.W. Chemical changes and increased degradability of wheat straw and oak wood chips treated with the white rot fungi Ceriporiopsis subvermispora and Lentinula edodes. Biomass Bioenergy 2017, 105, 381–391. [Google Scholar] [CrossRef]
- Mao, L.; van Arkel, J.; Hendriks, W.H.; Cone, J.W.; de Vos, R.C.; Sonnenberg, A.S. Assessing the nutritional quality of fungal treated wheat straw: Compounds formed after treatment with Ceriporiopsis subvermispora and Lentinula edodes. Anim. Feed Sci. Technol. 2021, 276, 114924. [Google Scholar] [CrossRef]
- Tirado-González, D.N.; Jáuregui-Rincón, J.; Tirado-Estrada, G.G.; Martínez-Hernández, P.A.; Guevara-Lara, F.; Miranda-Romero, L.A. Production of cellulases and xylanases by white-rot fungi cultured in corn stover media for ruminant feed applications. Anim. Feed Sci. Technol. 2016, 221, 147–156. [Google Scholar] [CrossRef]
- Davis, M.A.; Wong, K.H. Nitrogen metabolism in filamentous fungi. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K.A., Ebbole, D.J., Eds.; American Society for Microbiology Press: Washington, DC, USA, 2010; pp. 325–338. [Google Scholar]
- Periasamy, K.; Natarajan, K. Role of lignocellulosic enzymes during basidiomata production by Pleurotus djamor var. roseus. Indian J. Biotechnol. 2004, 3, 577–583. [Google Scholar]
- Kaur, M.; Sharma, S.; Sodhi, H. White rot fungus Calocybe indica: Incredulous factory of lignocellulolytic enzymes and their potential applicability for mushroom cultivation. Acta Aliment. 2022, 51, 424–436. [Google Scholar] [CrossRef]
- Sun, X.; Cheng, L.; Jonker, A.; Munidasa, S.; Pacheco, D. A review: Plant carbohydrate types—The potential impact on ruminant methane emissions. Front. Vet. Sci. 2022, 9, 880115. [Google Scholar] [CrossRef]
- Huyen, N.T.; Le, N.T.T.; Tuan, B.Q. Fermented sugarcane bagasse with the fungus Pleurotus eryngii reduced in vitro methane production. Livest. Res. Rural Dev. 2021, 33, 156–157. [Google Scholar]
- Alberts, J.F.; Gelderblom, W.; Botha, A.; Van Zyl, W. Degradation of aflatoxin B1 by fungal laccase enzymes. Int. J. Food Microbiol. 2009, 135, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, W.; Shen, Y.; Zhao, Y.; Wu, D.; Yin, H.; Yang, S.; Yuan, Q.; Liang, W.; Wang, J. Enhancing the degradation of Aflatoxin B1 by co-cultivation of two fungi strains with the improved production of detoxifying enzymes. Food Chem. 2022, 371, 131092. [Google Scholar] [CrossRef] [PubMed]
- Nazar, M.; Xu, Q.; Ullah, M.W.; Khan, N.A.; Iqbal, B.; Zhu, D. Integrated laccase delignification with improved lignocellulose recalcitrance for enhancing enzymatic saccharification of ensiled rice straw. Ind. Crops Prod. 2023, 202, 116987. [Google Scholar] [CrossRef]
- Liu, E.Y.; Wang, S.; Wang, S.; Khan, N.A.; Zhou, X.; Tang, S.; Zhou, C.; Tan, Z.; Liu, Y. Bacterial inoculants and enzymes based silage cocktails boost the ensiling quality of biomasses from reed, corn and rice straw. Chem. Biolog. Technol. Agricult. 2024, 11, 29. [Google Scholar] [CrossRef]
- Sufyan, A.; Khan, N.A.; Akbar, A.; Tang, S.; Tan, Z. Scaling-up fungal pretreatment of lignocellulose biomass: Impact on nutritional value, ruminal degradability, methane production, and performance of lactating dairy cows. Livest. Sci. 2024; in press. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Treatment Period (Days) | WRF Species | DM (% FM) | Concentration (% DM) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Ash | CP | NDF | ADF | Lignin | HC | CEL | |||
| 0 | A. bisporus | 24.9 | 5.09 a | 1.99 a | 86.3 g | 58.5 | 12.9 e | 27.8 c | 46.5 |
| P. djamor | 25.4 | 5.04 a | 2.02 a | 85.1 g | 57.3 | 12.0 e | 27.8 c | 45.3 | |
| C. indica | 25.8 | 5.08 a | 2.08 a | 85.6 g | 57.8 | 12.1 e | 27.8 c | 45.8 | |
| P. ostreatus | 25.2 | 5.09 a | 1.98 a | 86.5 g | 58.4 | 12.0 e | 28.0 c | 46.4 | |
| 21 | A. bisporus | 23.2 | 6.23 b | 2.86 b | 79.4 f | 53.8 | 10.5 d | 25.6 bc | 43.3 |
| P. djamor | 24.6 | 6.27 b | 3.60 c | 73.7 d | 52.2 | 10.0 cd | 21.5 a | 42.2 | |
| C. indica | 22.9 | 6.34 b | 4.17 d | 72.0 c | 50.7 | 8.40 b | 21.6 a | 42.3 | |
| P. ostreatus | 22.9 | 6.73 bc | 3.77 c | 73.9 d | 53.0 | 10.1 cd | 20.9 a | 42.8 | |
| 56 | A. bisporus | 22.8 | 7.11 cd | 3.98 d | 75.6 e | 50.7 | 9.30 c | 24.9 b | 41.4 |
| P. djamor | 24.1 | 7.75 e | 4.24 d | 69.1 b | 49.0 | 8.33 b | 20.1 a | 40.7 | |
| C. indica | 21.8 | 7.45 de | 5.02 e | 67.2 a | 47.5 | 7.23 a | 19.7 a | 40.3 | |
| P. ostreatus | 21.4 | 7.39 de | 4.30 d | 69.6 b | 50.4 | 8.27 b | 19.2 a | 42.1 | |
| SEM | 0.48 | 0.19 | 0.077 | 0.85 | 0.53 | 0.17 | 0.51 | 0.50 | |
| Overall mean fungal species | |||||||||
| A. bisporus | 23.6 a | 6.14 | 2.94 a | 80.4 a | 54.3 c | 10.6 c | 26.1 b | 43.7 b | |
| P. djamor | 24.7 b | 6.35 | 3.28 b | 76.0 c | 52.8 ab | 10.1 b | 23.1 a | 42.7 a | |
| C. indica | 23.5 a | 6.29 | 3.76 c | 75.1 d | 52.0 a | 9.20 a | 23.0 a | 42.8 a | |
| P. ostreatus | 23.2 a | 6.40 | 3.35 b | 76.9 b | 53.9 bc | 10.1 b | 22.7 a | 43.8 b | |
| SEM | 0.38 | 0.16 | 0.044 | 0.41 | 0.31 | 0.09 | 0.30 | 0.29 | |
| Overall mean treatment period | |||||||||
| 0 days | 25.3 c | 5.07 a | 2.02 a | 85.9 c | 58.0 c | 12.0 c | 27.9 c | 46.0 c | |
| 21 days | 23.4 b | 6.39 b | 3.60 b | 74.8 b | 52.4 b | 9.77 b | 22.4 b | 42.7 b | |
| 56 days | 22.5 a | 7.43 c | 4.39 c | 70.4 a | 49.4 a | 8.28 a | 21.0 a | 41.1 a | |
| SEM | 0.38 | 0.052 | 0.038 | 0.13 | 0.26 | 0.083 | 0.26 | 0.25 | |
| Significance | |||||||||
| WRF species | *** | NS | *** | *** | *** | *** | *** | * | |
| Treatment periods | ** | *** | *** | *** | *** | *** | *** | *** | |
| WRF species × incubation time | NS | ** | *** | *** | NS | *** | *** | NS | |
| Treatment Period (Days) | WRF Species | Percentage Losses of Nutrients in Comparison to Day 0 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| DM | Ash | CP | NDF | ADF | Lignin | HC | CEL | ||
| 21 | A. bisporus | 6.69 abc | −14.3 cd | −33.0 d | 14.2 a | 14.1 ab | 18.1 a | 14.2 a | 13.1 ab |
| P. djamor | 3.27 a | −20.5 bcd | −72.4 c | 16.2 b | 11.9 a | 19.0 a | 25.0 bc | 10.0 a | |
| C. indica | 11.2 d | −10.9 d | −77.9 bc | 25.0 e | 22.2 cd | 38.2 e | 30.8 c | 17.9 bc | |
| P. ostreatus | 9.11 cd | −20.2 bcd | −72.9 bc | 22.3 d | 17.6 bc | 22.9 b | 32.1 cd | 16.1 ab | |
| 56 | A. bisporus | 8.4 bcd | −27.9 b | −83.6 bc | 19.7 c | 20.5 c | 28.8 c | 18.1 ab | 18.4 bc |
| P. djamor | 5.4 ab | −45.7 a | −98.9 bc | 23.2 de | 19.1 bc | 34.2 d | 31.7 cd | 15.1 ab | |
| C. indica | 15.5 e | −24.0 bc | −104.1 a | 33.6 g | 30.5 e | 49.3 f | 40.0 de | 25.6 d | |
| P. ostreatus | 15.1 e | −23.3 bcd | −84.3 b | 31.7 f | 26.5 d | 41.5 e | 41.8 e | 23.0 cd | |
| SEM | 0.74 | 2.58 | 2.41 | 0.39 | 1.06 | 0.71 | 1.67 | 1.03 | |
| Overall mean fungal species | |||||||||
| A. bisporus | 7.56 b | −21.1 b | −58.2 c | 17.0 a | 17.3 a | 23.5 a | 16.1 a | 15.8 a | |
| P. djamor | 4.31 a | −33.1 a | −85.6 a | 19.8 b | 15.5 a | 26.6 b | 28.3 b | 12.5 a | |
| C. indica | 13.4 c | −17.4 c | −90.9 a | 29.3 d | 26.3 c | 43.8 d | 35.4 c | 21.7 b | |
| P. ostreatus | 12.1 c | −21.7 b | −78.6 b | 27.0 c | 22.2 b | 32.2 c | 36.9 c | 19.8 b | |
| SEM | 0.52 | 1.83 | 1.70 | 0.28 | 0.78 | 0.50 | 1.23 | 0.89 | |
| Overall mean treatment period | |||||||||
| 21 days | 7.58 a | −16.5 b | −64.3 b | 19.4 a | 16.4 a | 24.6 a | 25.5 a | 21.0 a | |
| 56 days | 11.1 b | −30.2 a | −92.7 a | 27.1 b | 24.2 b | 38.5 b | 32.9 b | 49.1 b | |
| SEM | 0.37 | 1.29 | 1.20 | 0.21 | 0.75 | 0.70 | 0.87 | 0.63 | |
| Significance | |||||||||
| WRF species | *** | *** | *** | *** | *** | *** | *** | *** | |
| Treatment periods | *** | *** | *** | *** | *** | *** | *** | *** | |
| WRF species × treatment period | * | ** | *** | *** | ** | *** | ** | ** | |
| Treatment Period (Days) | Fungal Species | IVDMD (g/100 g) | IVGP (mL/g OM) | CH4 (% IVGP) | AFB1, µg kg−1 | ||
|---|---|---|---|---|---|---|---|
| Total | CO2 | CH4 | |||||
| 0 | A. bisporus | 46.0 a | 156.3 a | 130.5 a | 22.8 ab | 14.6 ef | <5.0 |
| P. djamor | 46.8 a | 159.0 a | 133.7 a | 22.4 a | 14.1 ef | <5.0 | |
| C. indica | 46.9 a | 159.0 a | 132.2 a | 22.6 a | 14.2 ef | <5.0 | |
| P. ostreatus | 46.1 a | 155.3 a | 130.2 a | 22.9 ab | 14.7 f | <5.0 | |
| 21 | A. bisporus | 53.8 c | 176.0 e | 148.2 e | 24.4 bcdef | 13.4 bcd | <5.0 |
| P. djamor | 53.4 c | 190.0 d | 165.1 d | 23.8 cdef | 12.6 de | <5.0 | |
| C. indica | 64.6 a | 203.7 c | 177.2 c | 23.7 def | 11.6 ef | <5.0 | |
| P. ostreatus | 55.7 bc | 193.3 d | 165.9 d | 24.6 bcde | 12.7 cde | <5.0 | |
| 56 | A. bisporus | 59.2 c | 188.3 c | 160.2 c | 25.2 cdef | 13.1 cd | <5.0 |
| P. djamor | 57.9 c | 208.0 de | 180.5 e | 25.7 def | 12.4 bc | <5.0 | |
| C. indica | 65.1 d | 237.3 f | 205.9 f | 25.9 ef | 10.9 a | <5.0 | |
| P. ostreatus | 60.5 c | 215.7 e | 185.8 e | 26.8 f | 12.4 bc | <5.0 | |
| SEM | 1.98 | 1.82 | 1.90 | 0.40 | 0.25 | ||
| Overall mean of WRF species | |||||||
| A. bisporus | 53.2 ab | 173.6 a | 148.2 a | 24.1b | 13.9 c | <5.0 | |
| P. djamor | 51.3 a | 185.7 b | 158.3 b | 24.0b | 13.0 b | <5.0 | |
| C. indica | 58.3 c | 200.0 c | 174.2 d | 24.0b | 12.2 a | <5.0 | |
| P. ostreatus | 54.2 ab | 188.1 b | 161.1 c | 24.8a | 13.2 b | <5.0 | |
| SEM | 1.05 | 0.23 | 0.35 | 0.14 | 0.21 | ||
| Overall mean treatment period | |||||||
| 0 days | 45.0 a | 157.4 a | 132.1 a | 22.7 a | 14.4 c | <5.0 | |
| 21 days | 56.6 b | 190.8 b | 164.3 b | 24.1 b | 12.7 b | <5.0 | |
| 56 days | 61.0 c | 212.3 c | 186.4 c | 25.9 c | 12.3 a | <5.0 | |
| SEM | 1.29 | 0.91 | 0.88 | 0.20 | 0.12 | ||
| Significance | |||||||
| WRF species | * | *** | *** | NS | *** | ||
| Treatment period | *** | *** | *** | *** | *** | ||
| WRF species × treatment period | * | ** | ** | NS | * | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, N.A.; Khan, M.; Sufyan, A.; Saeed, A.; Sun, L.; Wang, S.; Nazar, M.; Tan, Z.; Liu, Y.; Tang, S. Biotechnological Processing of Sugarcane Bagasse through Solid-State Fermentation with White Rot Fungi into Nutritionally Rich and Digestible Ruminant Feed. Fermentation 2024, 10, 181. https://doi.org/10.3390/fermentation10040181
Khan NA, Khan M, Sufyan A, Saeed A, Sun L, Wang S, Nazar M, Tan Z, Liu Y, Tang S. Biotechnological Processing of Sugarcane Bagasse through Solid-State Fermentation with White Rot Fungi into Nutritionally Rich and Digestible Ruminant Feed. Fermentation. 2024; 10(4):181. https://doi.org/10.3390/fermentation10040181
Chicago/Turabian StyleKhan, Nazir Ahmad, Mussayyab Khan, Abubakar Sufyan, Ashmal Saeed, Lin Sun, Siran Wang, Mudasir Nazar, Zhiliang Tan, Yong Liu, and Shaoxun Tang. 2024. "Biotechnological Processing of Sugarcane Bagasse through Solid-State Fermentation with White Rot Fungi into Nutritionally Rich and Digestible Ruminant Feed" Fermentation 10, no. 4: 181. https://doi.org/10.3390/fermentation10040181