
Age and Growth of Japanese Anchovy (Engraulis japonicus, Temminck & Schlegel, 1846) in Coastal Waters around Shandong Peninsula, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey and Sample Processing
2.2. Data Analysis
2.2.1. Assessment of Anchovy Biomass
2.2.2. Size Distribution and Length–Weight Relationship
2.2.3. Relative Condition Factor
2.2.4. Age Estimation
2.2.5. Growth Parameters
3. Results
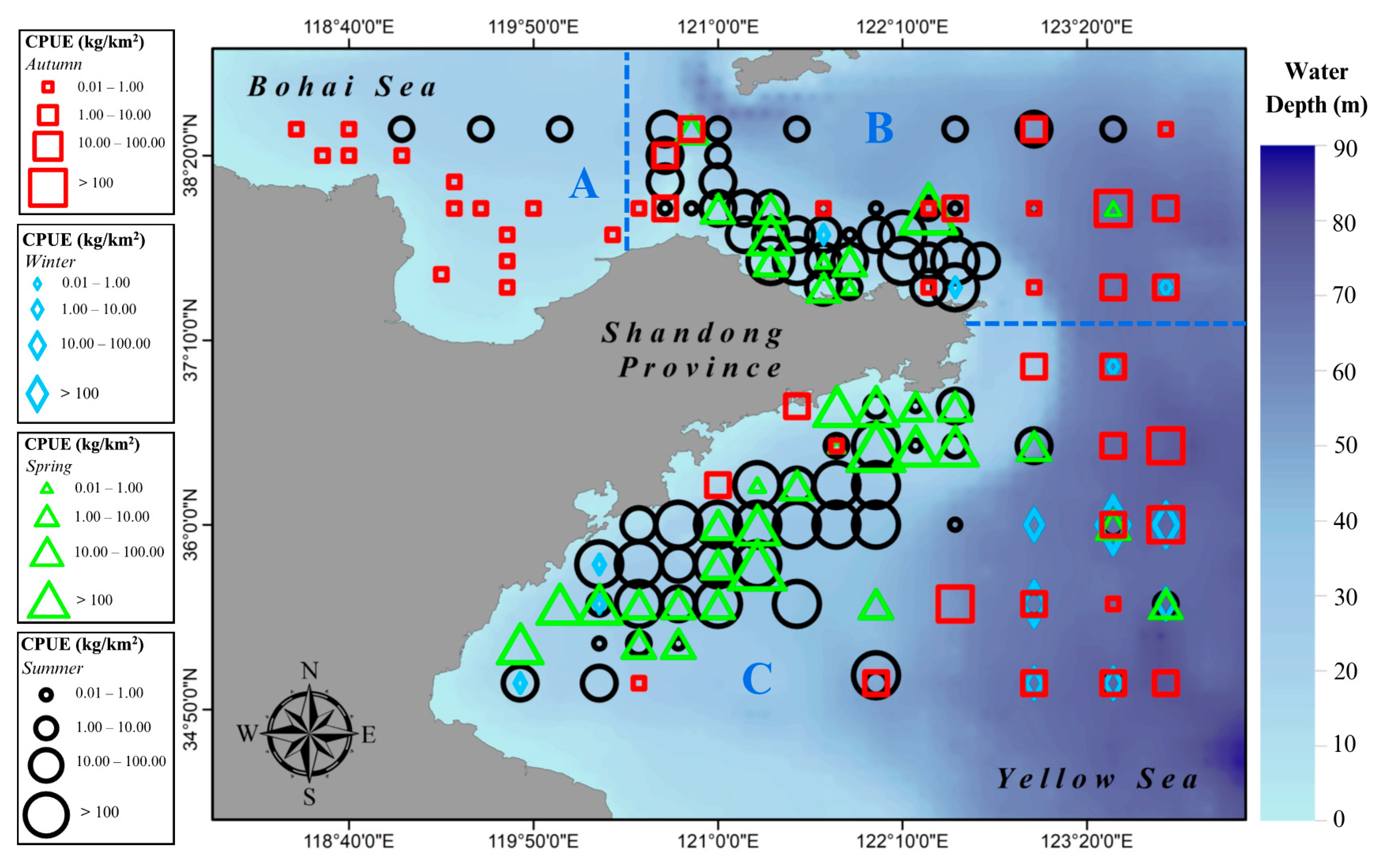
3.1. Spatial–Temporal Distribution
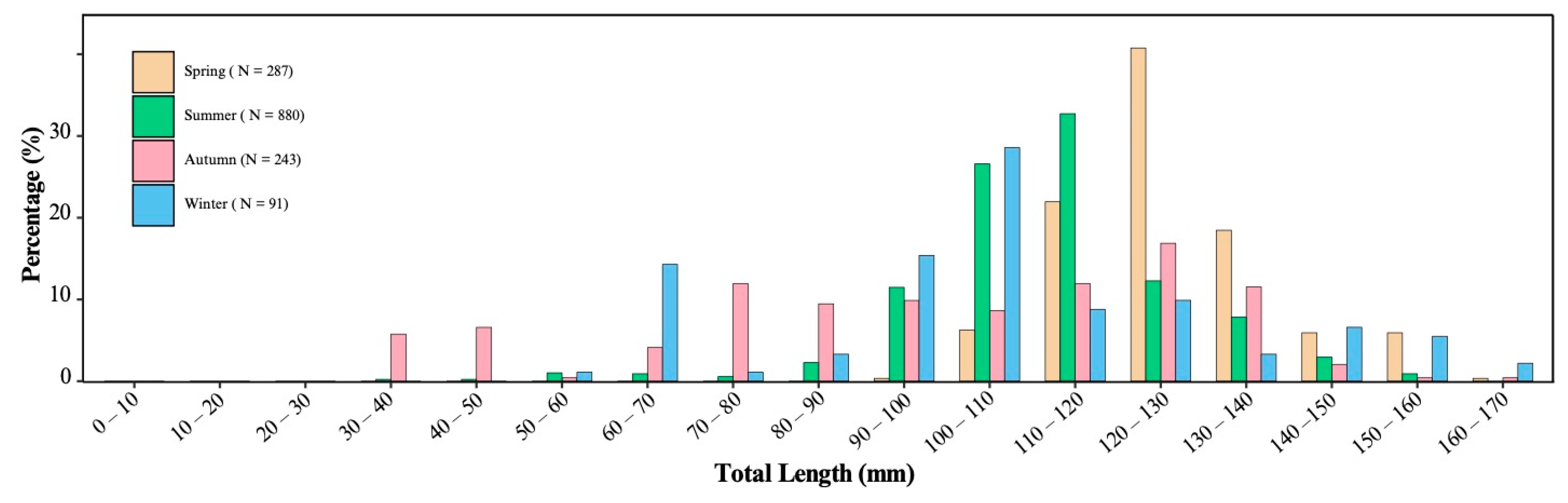
3.2. Seasonal Variation Pattern of Length and Length–Weight Relationship
3.3. Age Estimation and Growth Parameter
4. Discussion
4.1. Spatial–Temporal Distribution of Japanese Anchovy
4.2. Growth Pattern
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albo-Puigserver, M.; Pennino, M.G.; Bellido, J.M.; Colmenero, A.I.; Giráldez, A.; Hidalgo, M.; Gabriel Ramírez, J.; Steenbeek, J.; Torres, P.; Cousido-Rocha, M.; et al. Changes in Life History Traits of Small Pelagic Fish in the Western Mediterranean Sea. Front. Mar. Sci. 2021, 8, 570354. [Google Scholar] [CrossRef]
- Ningrum, E.W.N.; Patria, M.P. Microplastic contamination in Indonesian anchovies from fourteen locations. Biodiversitas J. Biol. Divers. 2021, 23, 125–134. [Google Scholar] [CrossRef]
- Xing, Q.; Yu, H.; Ito, S.-i.; Ma, S.; Yu, H.; Wang, H.; Tian, Y.; Sun, P.; Liu, Y.; Li, J.; et al. Using a larval growth index to detect the environment-recruitment relationships and its linkage with basin-scale climate variability: A case study for Japanese anchovy (Engraulis japonicus) in the Yellow Sea. Ecol. Indic. 2021, 122, 107301. [Google Scholar] [CrossRef]
- Ma, S.; Cheng, J.; Li, J.; Liu, Y.; Wan, R.; Tian, Y. Interannual to decadal variability in the catches of small pelagic fishes from China Seas and its responses to climatic regime shifts. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 159, 112–129. [Google Scholar] [CrossRef]
- Otero, J.; Hidalgo, M.; Secor, D. Life-history traits and environment shape small pelagic fish demography and responses to fishing and climate across European Atlantic seas. ICES J. Mar. Sci. 2023, 80, 1447–1461. [Google Scholar] [CrossRef]
- Peck, M.A.; Alheit, J.; Bertrand, A.; Catalán, I.A.; Garrido, S.; Moyano, M.; Rykaczewski, R.R.; Takasuka, A.; van der Lingen, C.D. Small pelagic fish in the new millennium: A bottom-up view of global research effort. Prog. Oceanogr. 2021, 191, 102494. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Liu, S.; Liang, C.; Xian, W. Assessing the Distribution and Sustainable Exploitation of Lophius litulon in Marine Areas Off Shandong, China. Front. Mar. Sci. 2022, 9, 759591. [Google Scholar] [CrossRef]
- Lefort, S.; Aumont, O.; Bopp, L.; Arsouze, T.; Gehlen, M.; Maury, O. Spatial and body-size dependent response of marine pelagic communities to projected global climate change. Glob. Chang. Biol. 2015, 21, 154–164. [Google Scholar] [CrossRef]
- Montero-Serra, I.; Edwards, M.; Genner, M.J. Warming shelf seas drive the subtropicalization of European pelagic fish communities. Glob. Chang. Biol. 2015, 21, 144–153. [Google Scholar] [CrossRef]
- Huang, C.-H.; Chen, C.-S.; Chiu, T.-S. Growth of Buccaneer Anchovy (Encrasicholina punctifer) During Juvenile Stage in the Waters off Southwestern Taiwan. J. Taiwan Fish. Res. 2018, 26, 53–61. [Google Scholar]
- Yedier, S. Otolith shape analysis and relationships between total length and otolith dimensions of European barracuda, Sphyraena sphyraena in the Mediterranean Sea. Iran. J. Fish. Sci. 2021, 20, 1080–1096. [Google Scholar] [CrossRef]
- Chen, Z.; Lu, H.; Liu, W.; Liu, K.; Chen, X. Beak Microstructure Estimates of the Age, Growth, and Population Structure of Purpleback Flying Squid (Sthenoteuthis oualaniensis) in the Xisha Islands Waters of the South China Sea. Fishes 2022, 7, 187. [Google Scholar] [CrossRef]
- Hayashi, A.; Zhang, K.; Saruwatari, T.; Kawamura, T.; Watanabe, Y. Distribution of eggs and larvae of Japanese anchovy Engraulis japonicus in the Pacific waters off northern Japan in summer. Fish. Sci. 2016, 82, 311–319. [Google Scholar] [CrossRef]
- Hwang, S.D.; Song, M.H.; Lee, T.W.; McFarlane, G.A.; King, J.R. Growth of larval Pacific anchovy Engraulis japonicus in the Yellow Sea as indicated by otolith microstructure analysis. J. Fish Biol. 2006, 69, 1756–1769. [Google Scholar] [CrossRef]
- Zhao, X.; Hamre, J.; Li, F.; Jin, X.; Tang, Q. Recruitment, sustainable yield and possible ecological consequences of the sharp decline of the anchovy (Engraulis japonicus) stock in the Yellow Sea in the 1990s. Fish. Oceanogr. 2003, 12, 495–501. [Google Scholar] [CrossRef]
- Wan, R.; Bian, X. Size variability and natural mortality dynamics of anchovy Engraulis japonicus eggs under high fishing pressure. Mar. Ecol. Prog. Ser. 2012, 465, 243–251. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, H.; Ye, Z.; Tian, Y.; Liu, Y.; Li, J.; Xing, Q.; Jiang, Y. Spawning strategy of Japanese anchovy Engraulis japonicus in the coastal Yellow Sea: Choice and dynamics. Fish. Oceanogr. 2020, 30, 366–381. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Q.; Ye, Z. A Bayesian analysis on the anchovy stock (Engraulis japonicus) in the Yellow Sea. Fish. Res. 2006, 82, 87–94. [Google Scholar] [CrossRef]
- Zheng, F.; Liu, Q.; Wang, Y. Study of impacts of environmental factors on stock and recruitment relationship of the anchovy stock in the Yellow Sea. South China FIsheries Sci. 2008, 4, 15–20. [Google Scholar]
- Li, X.; Wang, K.; Xu, B.; Xue, Y.; Ren, Y.; Zhang, C. Annual variation of species composition and spatial structure of fish community in Shandong offshore. J. Fish. China 2021, 45, 552–562. [Google Scholar] [CrossRef]
- Zhao, K.; Gaines, S.D.; García Molinos, J.; Zhang, M.; Xu, J. Climate change and fishing are pulling the functional diversity of the world’s largest marine fisheries to opposite extremes. Glob. Ecol. Biogeogr. 2022, 31, 1616–1629. [Google Scholar] [CrossRef]
- Bureau of Fisheries of the Ministry of Agriculture of the People’s Republic of China. China Fishery Statistical Yearbook; China Agriculture Pree: Beijing, China, 2021; pp. 1979–2021.
- Wan, R.-J.; Wei, H.; Sun, S.; Zhao, X.-Y. Spawning ecology of the anchovy Engraulis japonicus in the spawning ground of the Southern Shandong PeninsulaI. Abundance and distribution characters of anchovy eggs and larvae. Acta Zool. Sin. (Chin. Abstr. Engl.) 2008, 54, 785–797. [Google Scholar]
- Yu, H.; Yu, H.; Ito, S.-i.; Tian, Y.; Wang, H.; Liu, Y.; Xing, Q.; Bakun, A.; Kelly, R.M. Potential environmental drivers of Japanese anchovy (Engraulis japonicus) recruitment in the Yellow Sea. J. Mar. Syst. 2020, 212, 103431. [Google Scholar] [CrossRef]
- Iversen, S.A.; Zhu, D.; Johannessen, A.; Toresen, R. Stock size, distribution and biology of anchovy in the Yellow Sea and East China Sea. Fish. Res. 1993, 16, 147–163. [Google Scholar] [CrossRef]
- Chiu, T.S.; Chen, C.S. Growth and temporal variation of two Japanese anchovy cohorts during their recruitment to the East China Sea. Fish. Res. 2001, 53, 1–15. [Google Scholar]
- Zhu, J.c.; Zhao, X.y.; Li, F.G. Growth characters of the anchovy stock in the Yellow Sea with its annual and seasonal variations. Mar. Fish. Res. 2007, 28, 64–72. [Google Scholar]
- Liu, C.; Xian, W.; Liu, S.; Chen, Y. Variations in early life history traits of Japanese anchovy Engraulis japonicus in the Yangtze River Estuary. PeerJ 2018, 6, e4789. [Google Scholar] [CrossRef]
- Campana, S.E. Otolith science entering the 21st century. Mar. Freshw. Res. 2005, 56, 485–495. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, J.; Ning, Y.; Feng, J. Methodology Advances in Vertebrate Age Estimation. Animals 2024, 14, 343. [Google Scholar] [CrossRef]
- Bostanci, D.; Yalcinkaya, S.K.; Yedier, S.; Kurucu, G.; Polat, N. Otolith morphometry and scanning electron microscopy analysis of three fish species from the Black Sea. Acta Biol. Turc. 2024, 37, J3:1-7. [Google Scholar]
- D’Iglio, C.; Albano, M.; Famulari, S.; Savoca, S.; Panarello, G.; Di Paola, D.; Perdichizzi, A.; Rinelli, P.; Lanteri, G.; Spano, N.; et al. Intra- and interspecific variability among congeneric Pagellus otoliths. Sci. Rep. 2021, 11, 16315. [Google Scholar] [CrossRef]
- Popper, A.N.; Ramcharitar, J.; Campana, S.E. Why otoliths? Insights from inner ear physiology and fisheries biology. Mar. Freshw. Res. 2005, 56, 497–504. [Google Scholar] [CrossRef]
- Vieira, A.R. Assessment of Age and Growth in Fishes. Fishes 2023, 8, 479. [Google Scholar] [CrossRef]
- Li, P.; Liu, J.; Liu, Y.; Wang, T.; Liu, K.; Wang, J. A comparative study on the age, growth, and mortality of Gobio huanghensis (Luo, Le & Chen, 1977) in the Gansu and Ningxia sections of the upper Yellow River, China. BMC Ecol. Evol. 2024, 24, 30. [Google Scholar] [CrossRef]
- Bojesen, T.A.; Denechaud, C.; Malde, K.; Andrews, A. Annotating otoliths with a deep generative model. ICES J. Mar. Sci. 2024, 81, 55–65. [Google Scholar] [CrossRef]
- Cayetano, A.; Stransky, C.; Birk, A.; Brey, T.; Juanes, F. Fish age reading using deep learning methods for object-detection and segmentation. ICES J. Mar. Sci. 2024. [Google Scholar] [CrossRef]
- Aldanondo, N.; Cotano, U.; Etxebeste, E. Growth of young-of-the-year European anchovy (Engraulis encrasicolus) in the Bay of Biscay. Sci. Mar. 2011, 75, 227–235. [Google Scholar] [CrossRef]
- Zhu, Q.; Wu, R.; Masuda, Y.; Takahashi, Y.; Okabe, K.; Koizumi, K.; Iida, A.; Katayama, S. Spawning Phenology and Early Growth of Japanese Anchovy (Engraulis japonicus) off the Pacific Coast of Japan. Fishes 2023, 8, 11. [Google Scholar] [CrossRef]
- Cerna, F.; Gómez, M.; Moyano, G.; Plaza, G.; Morales-Nin, B. Spatial and inter-annual changes in the growth patterns of young-of-year anchovy in a high productive ecosystem. Fish. Res. 2022, 249, 106236. [Google Scholar] [CrossRef]
- Uriarte, A.; Rico, I.; Villamor, B.; Duhamel, E.; Dueñas, C.; Aldanondo, N.; Cotano, U. Validation of age determination using otoliths of the European anchovy (Engraulis encrasicolus L.) in the Bay of Biscay. Mar. Freshw. Res. 2016, 67, 951–966. [Google Scholar] [CrossRef]
- Yimer, M.A.; Cao, L.; Shen, J.Z.; Zhang, E. Age, growth, maturity and mortality of the tapetail anchovy Coilia brachygnathus (Engraulidae) in Lake Honghu, China. J. Fish Biol. 2023, 104, 410–421. [Google Scholar] [CrossRef]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State University Press/Ames: Ames, IA, USA, 1989. [Google Scholar]
- Le Cren, E.D. The Length-Weight Relationship and Seasonal Cycle in Gonad Weight and Condition in the Perch (Perca fluviatilis). J. Anim. Ecol. 1951, 20, 201–219. [Google Scholar]
- Li, P.; Qin, Y.; Chen, J. Age and growth of Japanese anchovies in the northern Yellow Sea. Fisheries Sci. 1982, 1, 1–5. [Google Scholar] [CrossRef]
- Soeth, M.; Fávaro, L.F.; Spach, H.L.; Daros, F.A.; Woltrich, A.E.; Correia, A.T. Age, growth, and reproductive biology of the Atlantic spadefish Chaetodipterus faber in southern Brazil. Ichthyol. Res. 2019, 66, 140–154. [Google Scholar] [CrossRef]
- Carvalho, M.G.; Moreira, C.; Queiroga, H.; Santos, P.T.; Correia, A.T. Age, growth and sex of the shanny, Lipophrys pholis (Linnaeus, 1758) (Teleostei, Blenniidae), from the NW coast of Portugal. J. Appl. Ichthyol. 2017, 33, 242–251. [Google Scholar] [CrossRef]
- Correia, A.T.; Manso, S.; Coimbra, J. Age, growth and reproductive biology of the European conger eel (Conger conger) from the Atlantic Iberian waters. Fish. Res. 2009, 99, 196–202. [Google Scholar] [CrossRef]
- Giannetti, G.; Donato, F. Age Determination Manual; AdriaMed Occasional Papers: Termoli, Italy, 2003. [Google Scholar]
- Sun, Y.; Zhang, C.; Tian, Y.; Watanabe, Y. Age, growth, and mortality rate of the yellow goosefish Lophius litulon (Jordan, 1902) in the Yellow Sea. J. Oceanol. Limnol. 2020, 39, 732–740. [Google Scholar] [CrossRef]
- Mu, X.; Zhang, C.; Zhang, C.; Yang, J.; Ren, Y. Age-structured otolith chemistry profiles revealing the migration of Conger myriaster in China Seas. Fish. Res. 2021, 239, 105938. [Google Scholar] [CrossRef]
- Xu, S.; Sun, P.; Zhang, C.; Li, J.; Xi, X.; Ma, S.; Zhang, W.; Tian, Y. Age and Feeding Habits of Caml Grenadier Macrourus caml in Cosmonauts Sea. Fishes 2023, 8, 56. [Google Scholar] [CrossRef]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations; Chapman & Hall: Boca Raton, FL, USA, 1957. [Google Scholar]
- Sparre, P.; Venema, S. Introduction to Tropical Fish Stock Assessment. Part 1: Manual. I, 2nd ed.; FAO Fisheries Technical Paper; FAO: Rome, Italy, 1998; Volume 306, 407p. [Google Scholar]
- Ogle, D.; Doll, J.; Wheeler, P.; Dinno, A. Simple Fisheries Stock Assessment Methods: Package “FSA”. FSA version 0.9.5, the R software version is 4.3.2, 2023. Available online: https://cran.r-project.org/web/packages/FSA/FSA.pdf (accessed on 27 February 2024).
- Niu, M.; Jin, X.; Li, X.; Wang, J. Effects of spatio-temporal and environmental factors on distribution and abundance of wintering anchovy Engraulis japonicus in central and southern Yellow Sea. Chin. J. Oceanol. Limnol. 2014, 32, 565–575. [Google Scholar] [CrossRef]
- Huang, J.; Sun, Y.; Jia, H.; Yang, Q.; Tang, Q. Spatial distribution and reconstruction potential of Japanese anchovy (Engraulis japonicus) based on scale deposition records in recent anaerobic sediment of the Yellow Sea and East China Sea. Acta Oceanol. Sin. 2014, 33, 138–144. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Alabia, I.D.; Tian, Y.; Ye, Z.; Yu, H.; Li, J.; Cheng, J. Impact of Climate Change on Wintering Ground of Japanese Anchovy (Engraulis japonicus) Using Marine Geospatial Statistics. Front. Mar. Sci. 2020, 7, 604. [Google Scholar] [CrossRef]
- Hongchao, R. Studies on The Eggs and larvae of Engraulis japonicus. Stud. Mar. Sin. 1984, 22, 1–32. [Google Scholar]
- Li, X.S.; Zhao, X.Y.; Li, F.; Li, F.G.; Dai, F.Q.; Zhu, J.C. Structure and its variation of the anchovy (Engraulis japonicus) spawning stock in the Southern waters to Shandong Peninsula. Mar. Fish. Res. 2006, 27, 46–53. [Google Scholar]
- Xiao, H.; Zhang, C.; Xu, B.; Xue, Y.; Liu, H.; Li, Z.; Ren, Y. Spatial Pattern of Ichthyoplankton assemblage in the coastal waters of Central and Southern Yellow Sea in Spring. Haiyang Xuebao (Chin. Abstr. Engl.) 2017, 39, 34–47. [Google Scholar]
- Aoki, I.; Miyashita, K. Dispersal of larvae and juveniles of Japanese anchovy Engraulis japonicus in the Kuroshio Extension and Kuroshio±Oyashio transition regions, western North Pacific Ocean. Fish. Res. 2000, 49, 155–164. [Google Scholar]
- Sasmita, S.; Pebruwanti, N.; Fitrani, I. Distribution of Anchovy Size in Purse Seine Fishing at Pulolampes Waters, Brebes Regency, Central Java. J. Fish. Mar. Sci. 2018, 2, 96–102. [Google Scholar]
- Liang, Z.; Sun, P.; Yan, W.; Huang, L.; Tang, Y. Significant effects of fishing gear selectivity on fish life history. J. Ocean Univ. China 2014, 13, 467–471. [Google Scholar] [CrossRef]
- Audzijonyte, A.; Richards, S.A.; Stuart-Smith, R.D.; Pecl, G.; Edgar, G.J.; Barrett, N.S.; Payne, N.; Blanchard, J.L. Fish body sizes change with temperature but not all species shrink with warming. Nat. Ecol. Evol. 2020, 4, 809–814. [Google Scholar] [CrossRef]
- Zhao, X. The Acoustic Survey of Anchovy in the Yellow Sea in February 1999, with Emphasis on the Estimation of the Size Structure of The Anchovy Population. Mar. Fish. Res. 2001, 22, 40–44. [Google Scholar]
- Lee, H.W.; Hwang, S.D.; Kim, H. Age and Growth Characteristic of Pacific Anchovy, Engraulis japonicus, in the Southern Waters of Korea Based on the Year Ring of Otolith and Collection Date. Korean J. Ichthyol. 2021, 33, 31–36. [Google Scholar] [CrossRef]
- Yukami, R.; Aoki, I.; Mitani, I. Daily age of adult Japanese anchovy Engraulis japonicus off eastern Honshu, Japan by otolith daily increment. Fish. Sci. 2008, 74, 1348–1350. [Google Scholar] [CrossRef]
- Singh, M.; Serajuddin, M. Length-weight, length- length relationship and condition factor of Channa punctatus collected from three different rivers of India. J. Entomol. Zool. Stud. 2017, 5, 191–197. [Google Scholar]
- Funamoto, T.; Aoki, I.; Wada, Y. Reproductive characteristics of Japanese anchovy, Engraulis japonicus, in two bays of Japan. Fish. Res. 2004, 70, 71–81. [Google Scholar] [CrossRef]
- Mehanna, S.F.; Osman, Y.A.A.; Khalil, M.T.; Hassan, A. Age and growth, mortality and exploitation ratio of Epinephelus summana (Forsskål, 1775) and Cephalopholis argus (Schneider, 1801) from the Egyptian Red Sea coast, Hurghada fishing area. Egypt. J. Aquat. Biol. Fish. 2019, 23, 65–75. [Google Scholar]
- El-Aiatt, A.O.; Shalloof, K.A.S.; El-Betar, T.A. Some Biological Aspect of 9 Fish Species from The Mediterranean Coast, North Sinai, Egypt, with Species Reference to Grey mullet, Mugil cephalus (Linnaeus, 1758). Egypt. J. Aquat. Biol. Fish. 2022, 26, 45–62. [Google Scholar]
- Richter, T.J. Development and Evaluation of Standard Weight Equations for Bridgelip Suckers and Largescale Suckers. North Am. J. Fish. Manag. 2011, 27, 936–939. [Google Scholar] [CrossRef]
- Joshi, K.K.; Sreeram, M.P.; Zacharia, P.U.; Abdussamad, E.M.; Varghese, M.; Mohamed Habeeb, O.M.M.J.; Jayabalan, K.; Kanthan, K.P.; Kannan, K.; Sreekumar, K.M.; et al. Check list of fishes of the Gulf of Mannar ecosystem, Tamil Nadu, India. J. Mar. Biol. Assoc. India 2016, 58, 34–54. [Google Scholar] [CrossRef]
- Mughul, W.G.; Rajput, S.; Laghari, S.; Hussain, I.; Khan, P.; Bilal, Z.; Sheikh, M.; Gachal, G.S.; Laghari, M.Y. Length-weight relationship and condition factor of Labeo bata (Hamilton) (Cypriniformes: Cyprinidae) found in Ranikot stream, Sindh-Pakistan. J. Surv. Fish. Sci. 2022, 8, 91–102. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, X.; Dai, F. Monthly variation in the fat content of anchovy (Engraulis japonicus) in the Yellow Sea: Implications for acoustic abundance estimation. Chin. J. Oceanol. Limnol. 2011, 29, 556–563. [Google Scholar] [CrossRef]
- McBride, R.S.; Somarakis, S.; Fitzhugh, G.R.; Albert, A.; Yaragina, N.A.; Wuenschel, M.J.; Alonso-Fernández, A.; Basilone, G. Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish Fish. 2015, 16, 23–57. [Google Scholar] [CrossRef]
- Basilone, G.; Ferreri, R.; Barra, M.; Bonanno, A.; Pulizzi, M.; Gargano, A.; Fontana, I.; Giacalone, G.; Rumolo, P.; Mazzola, S.; et al. Spawning ecology of the European anchovy (Engraulis encrasicolus) in the Strait of Sicily: Linking variations of zooplankton prey, fish density, growth, and reproduction in an upwelling system. Prog. Oceanogr. 2020, 184, 102330. [Google Scholar] [CrossRef]
- Morato, T.; Afonso, P.; Lourinho, P.; Barreiros, J.P.; Santos, R.S.; Nash, R.D.M. Length—weight relationships for 21 coastal fish species of the Azores, north-eastern Atlantic. Fish. Res. 2001, 50, 297–302. [Google Scholar]
- Effendie, I. Fisheries Bilogy (Biologi Perikanan); Yayasan Pustaka Nusatama (in Indonesia): Yogyakarta, Indonesia, 2002. [Google Scholar]
- Pauly, D.; Morgan, G.R. Length Based Methods in Fisheries Research. In ICLARM Conference Proceedings 13; International Center for Living Aquatic Resources Management: Manila, Philippines; Kuwait Institute for Scientific Research: Safat, Kuwait, 1987; p. 468. [Google Scholar]
- Liang, C.; Pauly, D. Growth and mortality of exploited fishes in China’s coastal seas and their uses for yield-per-recruit analyses. J. Appl. Ichthyol. 2017, 33, 746–756. [Google Scholar] [CrossRef]
- Kutsyn, D.N.; Chesnokova, I.I.; Danilyuk, O.N.; Statkevich, S.V.; Ablyazov, E.R.; Belogurova, R.E. Age, Growth, Maturation, and Mortality of Grass Goby Zosterisessor ophiocephalus (Gobiidae) of the Karkinitsky Gulf, the Black Sea. J. Ichthyol. 2022, 62, 109–116. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season (Time Collection) | Total Number of Survey Stations | Anchovy Occurrence Station (Anchovy Measurement Station) | Number of Anchovies (ind.) | Total Length (Length Range) (mm) | Wet Weight (Weight Range) (gram) | Frequency of Occurrence (%) | Mean CPUE (kg/km2) |
|---|---|---|---|---|---|---|---|
| Autumn (October) | 160 | 54 (19) | 243 | 98.63 ± 30.18 (35–165) | 9.01 ± 6.19 (0.27–24.89) | 33.75 | 2020.29 ± 113.28 |
| Winter (late December to early January) | 166 | 14 (11) | 91 | 106.98 ± 26.09 (60–162) | 7.71 ± 5.97 (0.79–25.51) | 8.43 | 248.03 ± 46.14 |
| Spring (May) | 161 | 37 (33) | 287 | 126.75 ± 13.66 (26–161) | 12.01 ± 4.56 (5.56–30.25) | 22.98 | 1356.37 ± 99.54 |
| Summer (August) | 164 | 83 (74) | 880 | 113.06 ± 16.55 (39–159) | 7.08 ± 3.60 (0.35–22.06) | 50.61 | 31,436.93 ± 1469.03 |
| Total | 651 | 188 (137) | 1501 | 112.97 ± 21.34 (26–165) | 8.37 ± 4.84 (0.27–30.25) | 115.77 | 35,061.62 ± 1727.99 |
| Season | Length–Weight Relationship (LWR) Parameters | Kn | ||
|---|---|---|---|---|
| a | b | R2 | ||
| Autumn | 0.005153 | 3.066 | 0.953 | 1.052 ± 0.287 |
| Winter | 0.003171 | 3.197 | 0.908 | 1.016 ± 0.155 |
| Spring | 0.002445 | 3.372 | 0.971 | 1.018 ± 0.151 |
| Summer | 0.003516 | 3.188 | 0.802 | 1.015 ± 0.165 |
| Location | L∞ (mm) | K (/Years) | t0 (Years) | References |
|---|---|---|---|---|
| Coastal waters of Shandong Peninsula (summer stock) | 154.40 | 0.604 | 0.965 | This study |
| Chinese waters | 184 | 0.51 | [83] | |
| Yellow Sea (wintering stock in 2001) | 190.1 | 0.47 | 0.06 | [27] |
| Yellow Sea (wintering stock in 2004) | 172.7 | 0.61 | 0.17 | |
| East China Sea (spring stock) | 53.22 | 0.019 | 2.80 | [26] |
| East China Sea (late summer stock) | 42.17 | 0.024 | 3.10 | |
| East China Sea and Yellow Sea | 155 | 0.6 | 1 | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fellatami, K.; Zhang, W.; Zhang, C.; Liu, S.; Tian, Y. Age and Growth of Japanese Anchovy (Engraulis japonicus, Temminck & Schlegel, 1846) in Coastal Waters around Shandong Peninsula, China. Fishes 2024, 9, 124. https://doi.org/10.3390/fishes9040124
Fellatami K, Zhang W, Zhang C, Liu S, Tian Y. Age and Growth of Japanese Anchovy (Engraulis japonicus, Temminck & Schlegel, 1846) in Coastal Waters around Shandong Peninsula, China. Fishes. 2024; 9(4):124. https://doi.org/10.3390/fishes9040124
Chicago/Turabian StyleFellatami, Kalysta, Wenchao Zhang, Chi Zhang, Shude Liu, and Yongjun Tian. 2024. "Age and Growth of Japanese Anchovy (Engraulis japonicus, Temminck & Schlegel, 1846) in Coastal Waters around Shandong Peninsula, China" Fishes 9, no. 4: 124. https://doi.org/10.3390/fishes9040124