
Short-Interval, High-Severity Wildfire Depletes Diversity of Both Extant Vegetation and Soil Seed Banks in Fire-Tolerant Eucalypt Forests

1
School of Agriculture, Food and Ecosystem Sciences, Faculty of Science, The University of Melbourne, 500 Yarra Boulevard, Burnley, VIC 3121, Australia
2
School of Agriculture, Food and Ecosystem Sciences, Faculty of Science, The University of Melbourne, Parkville, VIC 3010, Australia
*
Author to whom correspondence should be addressed.
Fire 2024, 7(4), 148; https://doi.org/10.3390/fire7040148
Submission received: 7 February 2024
/
Revised: 16 April 2024
/
Accepted: 18 April 2024
/
Published: 19 April 2024
(This article belongs to the Special Issue Effects of Fires on Forest Ecosystems)
Abstract
:Many plant species are well-adapted to historical fire regimes. An increase in the severity, frequency, and extent of wildfires could compromise the regenerative capacity of species, resulting in permanent shifts in plant diversity. We surveyed extant vegetation and soil seed banks across two forest types with contrasting historical fire regimes—Shrubby Dry Forest (fire return interval: 10–20 years) and Sub-Alpine Woodland (50–100 years). Over the past 20 years, both forests have been subject to repeated, high-severity wildfires at intervals significantly shorter than their historical return intervals. We examined the soil seed bank response to fire-cued germination, and whether the plant diversity in soil seed banks and extant vegetation demonstrated similar responses to short-interval, high-severity wildfires. The soil seed bank demonstrated a positive response to heat in combination with smoke, and for the Sub-Alpine Woodland, this was limited to sites more frequently burnt by fire. With an increase in fire frequency, there was a decline in species richness and Shannon’s Diversity and a shift in species composition in both extant vegetation and the soil seed bank. The fire frequency effects on the relative richness of trait associations were restricted to the Shrubby Dry Forest, and included an increase in short-lived obligate seeders, wind-dispersed species, and ant-dispersed shrubs in burnt relative to long unburnt sites in both extant vegetation and the soil seed bank. Graminoids were the most abundant component of the soil seed banks of Sub-Alpine Woodlands, and this increased with more frequent fire, with a similar trend (p = 0.06) in extant vegetation. Clear shifts in plant diversity in both soil seed banks and extant vegetation in forest types with contrasting historical fire regimes suggest that emerging fire regimes are pushing ecosystems beyond their historical range of variability, including potentially more flammable states and a decline in the buffering capacity of soil seed banks.
1. Introduction
Fire is a globally pervasive and dominant disturbance to terrestrial ecosystems and plays a critical role in shaping plant communities [1]. While adapted to fire, plants are not necessarily adapted to all fire regimes [2,3,4]. The potential for increases in wildfire extent, frequency, and severity under climate change [5,6,7] is being realised on a global scale [8]. In southeastern Australia, wildfires of an unprecedented scale burnt an estimated 7.2 million hectares of temperate forests during the 2019–20 fire season [9]. This megafire followed nearly two decades of an increased occurrence of high-severity fires in the region [10]. This shift in fire regime, particularly to more frequent wildfires, can threaten the persistence of woody species [11] and produce shifts in species composition, including state changes such as a loss of obligate seeder canopy species [12,13,14].
Plant responses to fire are dictated by a combination of fire-adapted traits that allow individuals to survive fire or regenerate following fire and include resprouting, serotiny, and fire-cued germination [3]. Resprouters survive fire by initiating new growth from protected meristems located in above- and/or belowground (i.e., lignotubers) organs [4]. In contrast, obligate seeders are species with adult plants that are killed by fire, with the replacement of individuals reliant on seed dispersal and the germination of canopy- or soil-stored seed [15], while facultative resprouters combine both strategies [16]. The hard, impermeable seed coat of physically dormant seed enables the seed to survive fire temperatures and provides for the accumulation of soil-stored seed during the inter-fire period [3,17]. The heat shock from a fire renders the seed permeable to water and allows for germination [18]. Physically dormant species typically have little response to smoke, although species with other seed dormancy mechanisms can demonstrate increased rates of germination in response to smoke cues [19,20]. These fire-related germination cues provide seedlings with the opportunity to exploit the reduced levels of competition for resources provided by the post-fire environment [21].
Fire-related germination cues may be associated with a parental environment [22], including fire regime [23,24,25,26]; other fire-adapted traits [27]; or other plant traits such as life form [19,27]. For example, Manela et al. [24] reported an increased response to ex situ smoke cues in more frequently burnt sites in a Mediterranean woodland, which was predominantly due to the increased germination of annuals, with similar findings for the in situ application of smoke [28]. This positive response of annual species to smoke cues has been reported across a range of floras, including in southeastern Australia [24,28,29,30,31]. We might also expect obligate seeders to demonstrate a greater response to fire-related germination cues than resprouters, given their greater dependence on post-fire seedling recruitment for population persistence [19,32]. In a broad-scale survey of southeastern Australian species, Carthey et al. [19] demonstrated that obligate seeders are more likely to show positive responses to smoke cues than resprouters. Similarly, many obligate seeding species have seeds with physical dormancy and are more likely to demonstrate heat-promoted germination than resprouters [27,33]. Dispersal mode may also play an important role in post-fire plant assemblages through direct effects on colonisation from surviving vegetation [34], and indirectly through associations with fire-related germination cues and the formation of persistent seed banks. For example, in Australia, many woody species are ant-dispersed species with hard seed coats and may form long-term (≥5 years in [35]), persistent seed banks through physical dormancy or burial [36,37]. Given that soil temperatures decrease sharply with soil depth [38], seed burial by ants may also act to protect physically dormant seed from lethal temperatures and place them at depths where temperatures are conducive to breaking dormancy [36].
The persistence of plant communities in fire-prone areas is primarily related to the interaction of fire-adapted traits with fire regime [39,40], which will result in distinct post-fire assemblages and alternative successional pathways [21,41]. Increases in fire frequency may compromise the regenerative capacity of species, including the depletion of some resprouting species and the potential extinction of obligate seeders if the inter-fire interval is shorter than the time to reproductive maturity. For extant vegetation, recurrent wildfire effects include demographic shifts in fire-tolerant canopy species [42,43], the decline and local extinction of obligate seeding tree [12,44] and shrub species [11,45,46], and a shift to more simplified communities dominated by a short life span, fast growth, and early reproductive life forms [47,48,49]. The concomitant effects of recurrent wildfires on soil seed bank have received less attention, despite its importance as a reservoir of plant diversity, particularly in fire-prone communities [50,51], and work demonstrating marked differences in other fire regime effects—such as prescribed fire [52,53] and time since fire [54,55]—on the composition in the soil seed bank relative to extant vegetation. In the Mediterranean Basin, an increase in wildfire frequency results in an increase in the density of soil seed banks due to a shift from woody to herbaceous species with a short time to reproductive maturity, with, however, no loss in species richness due to the predominance of a soil, rather than canopy seed store [24,31]. This resistance to species loss may be conferred by the bet-hedging strategy used by many species, especially those with physical dormancy, with a reservoir of soil-stored seed buffering the probability of extinction following consecutive fires [17,56,57]. However, a high fire severity can result in the increased germination of physically dormant species and result in a decrease in the size of the residual seed bank, which may, in turn, impact the ability of a species to persist with frequent fires [58,59]. In recent work, Duivenvoorden et al. [60] reported a decline in richness in the soil seed bank following repeated high-severity fires for montane eucalypt forests, which contrasted with a lack of response in the extant vegetation. It is then the functional identities of both the extant vegetation and soil seed bank that are key in predicting post-fire plant assemblages and, in turn, the potential for the re-establishment of ecosystem processes after a fire [61,62].
This study examines the effects of fire-related germination cues and fire frequency on the plant diversity of temperate forest ecosystems via an examination of plant species and functional trait metrics. The study was conducted on two different forest types, Shrubby Dry Forests and Sub-Alpine Woodlands in southeastern Australia. Each forest type is dominated by an overstorey of resprouting eucalypts, and both have been subjected to repeated, high-severity wildfires at intervals significantly shorter than the historical norm [43,63]. The following questions and associated hypotheses were addressed:
- Does the soil seed store respond to fire-related germination cues, and are these cues related to fire history and other plant traits? We hypothesize that the soil seed store will demonstrate a positive response to fire-related germination cues and that these cues will be more prevalent in areas more frequently burnt by fire. We expect seeders, ant-dispersed species, and short-lived species to demonstrate stronger responses to fire-related germination cues than other traits.
- Do the soil seed bank and extant vegetation demonstrate similar responses to fire frequency? We expect a decline in the soil seed bank with more frequent fires and a reduction in the capacity of the soil seed bank to buffer species losses in extant vegetation with an increasing fire frequency. For both the soil seed bank and extant vegetation, we hypothesize that an increase in fire frequency will benefit species with fast reproductive maturity but filter out longer-lived woody species with a greater age to reproductive maturity.
2. Materials and Methods
This study encompassed two areas supporting fire-tolerant eucalypt communities that had been subjected to short-interval, high-severity wildfires. Both areas have been the subject of previous work by Fairman et al. [43,63], who provided detailed descriptions of the study areas and methodology associated with the site selection. The sites chosen for this work are a sub-set of those previously established. In brief, both study areas are in southeastern Australia and support distinct vegetation types: (i) mixed-species Shrubby Dry Forests in the Ben Cruachan and Glenmaggie State Forests in West Gippsland (37.806° S, 146.754° E); and (ii) a Eucalyptus pauciflora Sub-Alpine Woodland on Mount Hotham (37.027°, 147.119° E) (Figures S1 and S2).
2.1. Glenmaggie Study Area and Site Selection
The annual rainfall ranges from 700 to 800 mm, falling mainly in spring (September–November), with mean annual temperatures ranging from 12.4 to 14.5 °C (1981–2010; [64]). The area encompasses elevations of 206–604 m asl, characterised by moderate to steep slopes. The study sites were established in a widespread dry sclerophylly forest, locally classified as a Shrubby Dry Forest (Ecological Vegetation Class, EVC, [65]). This vegetation type is dominated by a diverse mix of eucalypts, including Eucalyptus cypellocarpa, E. globulus, E. globoidea, E. gonicalyx, E. macrorhyncha, E. muelleriana, E. polyanthemos, E. sieberi, and E. tricarpa. Mixed-severity wildfires in 2007 and 2013 culminated in extensive areas of the forest that were either unburnt (since 1939) or burnt once or twice within six years (Figure 1; Table S1). This inter-fire interval is short relative to the historical range (10–20 years in [66]) and below the minimum tolerable fire interval of 15 years required for the longest-lived species to reach a sufficient size or produce a seed store to survive a subsequent high-severity fire [67]. This relatively short inter-fire interval is, however, sufficient for the profuse regeneration of elevated fuels and a return to a very high overall fuel hazard rating [68].
We surveyed five replicate sites in each fire frequency class: unburnt, single burnt in 2013, single burnt in 2007, and double-burned in 2007 and 2013, for a total of 20 sites. At each site, we re-established the original 50 m × 20 m boundary of Fairman et al. [63], and within that, established two 10 m × 10 m plots for sampling. The plots were centred at 15 m and 35 m along the 50 m boundary and at 10 m along the 20 m boundary.
For both Glenmaggie and Mount Hotham, areas of high-severity wildfire were identified by the Normalised Burn Ratio, as estimated from pre- and post-fire SPOT images in combination with Landsat and high-resolution aerial imagery [63].
2.2. Mount Hotham Study Area and Site Selection
The annual rainfall ranges from 1400 to 1700 mm, the majority falling as winter (June–August) snow with a mean annual temperature range from 5.5 to 7.1 °C (1981–2010; [64]). The elevation spans 1477 to 1735 m asl, supporting moderate to steep slopes. Sites were established in forests dominated by Eucalyptus pauciflora and its subspecies and locally classified as Sub-Alpine Woodland (EVC, [65]). These forests lie between 1400 and 1900 m asl at the altitudinal tree limit in Australia [69,70,71,72].
Combinations of large, high-severity wildfires in 2003, 2007, and 2013 created mosaics of E. pauciflora forests that were either unburnt (since 1939), burnt once, twice, or thrice over that decade. This fire frequency is well below the historical norm (one per 50–100 years [73]) and below the minimum tolerable fire interval of 50 years required for the longest-lived species to survive a subsequent high-severity fire [67]. We surveyed four replicate sites in each fire frequency class for a total of 16 study sites (Figure 1, Table S1). The distribution and overlap of the three wildfires constrained the choice of wildfire years in the single- and double-fire history classes, meaning that the fire history varied slightly (Table S1). At each site, we re-established the original 50 m × 10 m boundary of Fairman et al. [43] and established plots following methods for Shrubby Dry Forests.
2.3. Vegetation Survey and Treatment
All sites were sampled in spring 2017, before the onset of flowering and seeding of the extant vegetation. Sampling during this period ensured that the most viable seed found in the soil could be confidently assumed to have been produced in previous seasons, and, therefore, have passed through a minimum of one entire winter and summer period [74,75].
For each plot, the projected foliage cover of all the extant vascular vegetation was estimated to the nearest percentage. For cover less than <1%, cover was assigned at 0.05%. Soil samples were collected at the same time to provide an estimate of the soil seed bank using the seedling emergence method. For this, surface leaf litter was removed [30,76] and 36 soil cores at a 5 cm depth (diameter 6.84 cm) were randomly sampled and bulked to represent a surface area of 0.1323 m2 (equivalent to 0.13% of the plot surface area). The soil samples were air-dried and sieved to 5 mm, concentrating the seed in a smaller volume of soil.
2.4. Soil Seed Bank Treatment
We applied a series of treatments to meet a range of dormancy requirements common to the sampled species [60,77], with applied heat treatments typical of temperatures reached at a 0–5 cm soil depth during wildfires in forests of southeastern Australia [78]. For each plot, each composite soil sample was split into four equal portions and treated separately with high heat and smoke (HHS), low heat and smoke (LHS), smoke only (S), or no treatment (control, C). The samples were placed in aluminium trays at a depth of 2 cm, with thermocouples located at a 1 cm soil depth to measure the temperature. The samples were heated in ovens at 95 °C until the soil temperatures reached 80 °C (high-heat treatment) and 30–50 °C (low-heat treatment) for a duration of 30 min. Temperatures of 80 °C are known to maximise the germination of hard-coated shrub seeds [18], with temperatures of 30–50 °C known to promote sedge and grass species germination [79]. The smoke treatment was applied to the heated soil samples after they had cooled and followed Meers et al. [80]. Control trays with media but without seed bank material were used to monitor for contamination.
The trays were placed randomly on benches in a climate-mediated (~16 °C night and 22 °C day) glasshouse with automatic irrigation and re-randomised fortnightly to reduce position bias. Emerging seedlings were recorded at weekly intervals and removed from the trays once identified. After 45 weeks, there was little further germination, and unidentified seedlings were removed and grown in larger pots until they could be identified. The soils from all treatments were then turned to promote the germination of any remaining seeds within the profile. At this point, gibberellic acid (200 mg L−1) was applied to the seedling trays at a rate of 30 mL per tray for 5 days [81] to account for any underlying physiological dormancy [82]. The seedling trays were monitored for seedling emergence for a further nine weeks. While the soils were not examined for seeds that had not germinated, this technique is deemed appropriate for determining which seeds may germinate in the event of a disturbance [83]. The trial was run from December 2017 to December 2018.
2.5. Functional Types
The functional types associated with resource partitioning, response to disturbance (life form and fire response), and the capacity to re-colonise (dispersal mode) were recorded for all species [84] (Table S2). The life form classification followed Raunkiear [85] and the dispersal mode classification was based on diaspore morphology [84], with a 0.1 mg seed mass used to separate self-dispersed (‘mobile’; [86]) species from heavier seeded barochores. Ballistochores were classified as either myrmecochores or barochores [87]. Those for fire response followed [88,89], recognising that many species lie along a continuum between resprouting and seeding, and a binary view of these traits may lead to an over- or underestimate of regeneration potential [90,91].
2.6. Data Screening
In the soil seed bank treatment trial, the control trays indicated contamination from Cardamine occulata and Oxalis spp., and these species were excluded from analysis. To facilitate comparisons between seed bank germinants and extant vegetation, Epilobium, Galium, Geranium, Juncus, Lepidosperma, Plantago, Rytidosperma, Trifolium, and Wahlenbergia species were aggregated to genus in instances where all recorded species within these genera could not be identified to the species level. An analysis of the soil seed bank treatment effects retained species-level identifications where present. For extant vegetation, all subspecies of Eucalyptus pauciflora (niphophila, pauciflora) were aggregated to the species level, as we could not confirm the sub-species across all sites.
2.7. Statistical Analysis—Soil Seed Bank Treatment and Fire Frequency
We calculated the number of seedlings that germinated in each of the four treatments across three levels of data aggregation (i) individual species; (ii) all species (total, native, and introduced); and (ii) functional types. For functional types, the species richness and germinant density were summed for species within each life form, dispersal mode, and fire response strategy. Species were excluded where trait information was missing (Table S3).
For individual species, the significance of treatment effects was limited to those species recorded in eight or more plots and only included plots in which each species had germinated to avoid zero inflation [92,93]. The analysis followed Kruskal–Wallis and Dunn’s pairwise comparisons (where p ≤ 0.05; SPSS V22.0.0.0, IBM Corp Chicago, IL, USA, 1989, 2013) of standardised data (division by the total). For all subsequent analyses, our experimental unit was at the site level and data were pooled across the two plots.
We assessed the seed bank treatment (four factors: C, S, LHS, and HHS) and fire frequency (four factors: number of wildfires since 2003) effects on species composition via two-way PERMANOA on the Bray–Curtis similarity of transformed (log x + 1) species counts (PERMANOVA Version 1.0.5, PRIMER-E Ltd., Plymouth, UK, [94,95]. Two-way PERMANOVA on the Euclidean distance between sites was used to test effects of the seed bank treatment and its interaction with fire frequency on Shannon’s diversity (H′) [96] and on the richness and germinant density of all species and functional types. In each instance, data were standardised (division by the total) and transformed (log x + 1) [97,98]. For Shrubby Dry Forest, a dummy variable of 1 was added prior to the calculation of the similarity matrix to avoid undefined values. Where differences were significant, the SIMPER procedure of PRIMER (Version 6.1.15, PRIMER-E Ltd., Plymouth, UK) [99] was used to determine the species (or plant attributes) that contributed most to dissimilarity.
The fire frequency classes differed within each forest type: Shrubby Dry Forest (Unburnt, Single 2013, Single 2007, Double) and Sub-Alpine Woodland (Unburnt, Single, Double, and Triple). For Shrubby Dry Forest, the two sets of single-burnt (in 2007 and 2013) sites were not lumped, as we recognised the potential for differences in ‘time since fire’ to affect the species richness and composition of assemblages and functional response types that are independent of fire frequency, given that species with different traits have alternative life histories following a fire [98,100,101].
2.8. Statistical Analysis—Vegetation Type and Fire Frequency
For each site (the experimental unit), data were pooled across the two plots. Given that each soil seed bank treatment produced a unique set of additional species (and that there was an interaction between germination treatment and fire frequency—see results), the comparison with extant vegetation was based on the soil seed bank germinants summed across all treatments, as this approach provides for the most comprehensive account of the germinable soil seed bank and removes the effects of the germination cues [53,80,92]. All species in the extant vegetation (under and overstorey species) and those that had germinated in the soil seed bank were considered in the comparison.
For each of two levels of data aggregation (all species, functional types—as above), we examined the vegetation type (two factors: extant vegetation and soil seed bank) and fire frequency (four factors: number of wildfires since 2003) effects on richness and composition by two-way PERMANOVA, following methods described for the soil seed bank. For each two-way analysis, data were presence–absence transformed to avoid problems commonly associated with measures of abundance, including: (i) apparent discrepancies in the similarity between vegetation types that can be explained by frequently occurring aboveground species that have sparse cover, but produce disproportionally high numbers of seeds relative to cover abundance (e.g., Carex and Luzula spp.); (ii) difficulties with quantifying the abundance of clonal species; and (iii) the influence of fire history on stocking densities [43,63,75,98,102]. We also considered fire frequency effects on H′ in extant vegetation and the soil seed bank using one-way PERMANOVA.
3. Results
3.1. Soil Seed Bank—Individual Species
In Shrubby Dry Forest, a total of 3617 germinants were recorded for 101 vascular plant species from 39 families (Table S3). Introduced species comprised 8.3% of the total germinant pool across 15 species. In total, 40 species were restricted to a single treatment type, with 15 of these belonging to the HHS treatment (Table S3). Of the 35 species for which statistical testing was possible, 8 species, including 2 introduced species, demonstrated a strong germination response to the LHS and/or HHS treatments, with marked increases in densities over the control and/or smoke treatments (Table S3). Several species of herbs, sedges, grasses, annuals, and shrubs did not respond to the treatments (Table S3). Acacia species were limited to the LHS or HHS treatments, as were other shrubs belonging to Fabaceae, although the statistical testing of individual species was not possible.
In Sub-Alpine Woodland, a total of 3024 germinants were recorded for 86 vascular plant species from 32 families (Table S3). Introduced species comprised 15.4% of the total germinant pool across 21 species (Table S3). A total of 5 of the 18 species for which statistical testing was possible demonstrated significant increases in LHS and/or HHS treatments over the smoke and/or control treatments. Several species of herbs and sedges did not respond to the treatments (Table S3). Consistent with trends for the same genera within the Shrubby Dry Forest, Acacia alpina and Pelargonium helmsii were limited to the LHS or HHS treatments, although the statistical testing of treatment effects was not possible.
3.2. Soil Seed Bank—All Species
In Shrubby Dry Forest, heat plus smoke cues increased the native species richness, with effects on the total species richness limited to sites that had been burnt (Figure 2 and Figure 3; Table S4). For both forest types, the combined heat and smoke treatments produced a greater density of germinants relative to the control and smoke only treatments, and in Sub-Alpine Woodland, these effects were limited to sites that had been burnt (Figure 2 and Figure 3; Table S4). For both forest types, the species richness and density decreased with an increasing fire frequency (Figure 2, Table S4).
The soil seed bank treatment did not affect the Shannon’s diversity (H′) or species composition in either forest type, with also no interaction with fire frequency (Tables S5 and S6, Figure S3). The species composition and H′ did, however, differ according to fire frequency, with declines in H′ and species similarity with an increase in fire frequency and the greatest differences between the unburnt and most frequently burnt sites in both forest types (Tables S5 and S6, Figure S3). SIMPER analyses indicated that differences in species composition were the result of additive contributions from a few dominant species in both forest types (Table S6).
3.3. Soil Seed Bank—Functional Types
In Shrubby Dry Forest, the relative proportion (both richness and density) of species within different life forms, dispersal mechanisms, and fire response strategies varied with the soil seed bank treatment and fire frequency, with responses to fire-related germination cues being independent of fire frequency (Table 1). The treatment effects included a greater proportion (both richness and density) of phanerophytes and barochores, and an increased richness of myrmecochores and resprouters in the HHS-treated samples relative to the smoke and control (Table 1). The fire frequency effects included decreases in the proportion of proto-hemicryptophytes, species with mobile seed and weak seeders (Sr), and in the richness of phanerophytes with an increasing fire frequency (Table 1). In Sub-Alpine Woodland, the relative proportion of functional types did not change in response to fire-related germination cues (Table 2), with, however, significant fire frequency effects that included an increase in the proportion of erect rosettes, a decrease in the proportion of flat and partial rosettes in double-burnt sites, and an overall decrease in phanerophytes, ant-dispersed species, and weak seeders with an increasing fire frequency (Table 2).
3.4. Vegetation Type and Fire Frequency Effects—All Species
A total of 156 species were recorded in the soil seed bank plus extant vegetation in the Shrubby Dry Forest and 157 in the Sub-Alpine Woodland (Table S3). Phanerophytes were the dominant life form, while barochores and resprouters were the dominant modes of dispersal and fire response strategies, with each of these trends being consistent across forest types (Figure S4). For Shrubby Dry Forest, 55 species were unique to extant vegetation, 43 species were unique to the soil seed bank, and 58 species were shared between the soil seed bank and extant vegetation. Corresponding species’ numbers for Sub-Alpine Woodland included 71 for extant vegetation, 30 for the soil seed bank, and 56 that were shared.
For both forest types, the species richness differed significantly with vegetation type and fire frequency, with, however, no interaction effects (Figure 4, Table S7). The native species richness in extant vegetation was consistently greater than that in the soil seed bank for both forest types, with contrasting trends for introduced species in Shrubby Dry Forest (Figure 4). Species richness declined with an increasing fire frequency (Figure 4). In Shrubby Dry Forest, the richness within unburnt and single-burnt sites was significantly greater than that in double-burnt sites, while in Sub-Alpine Woodland, both richness (Figure 4) and H′ (Figure S5, Table S8) in unburnt sites were significantly greater than those in double- and triple-burnt sites (Figure 4). In Shrubby Dry Forest, there was no change in H′ with fire frequency for extant vegetation, with trends in the soil seed bank matching those for species richness (Figure S5, Table S8).
The similarity in species composition between extant vegetation and the soil seed bank was low (Shrubby Dry Forest 15%; and Sub-Alpine Woodland 25%) and independent of fire frequency (Table S9). There were significant changes in species composition with fire frequency, with the greatest differences being between double-burnt and unburnt sites for both forest types and differences also occurring between double and single (2013) burns in Shrubby Dry Forest, while triple burns were relative to all other fire frequencies in Sub-Alpine Woodland (Figure 5, Table S9). SIMPER indicated that, for both forest types, the species most responsible for the differences between vegetation types included Eucalyptus spp. and shrubs which were more frequent in the extant vegetation, while the fire frequency effects were largely explained by Stypandra glauca, Wahlenbergia spp., and Daviesia buxifolia in Shrubby Dry Forest, and Cerastium vulgare, Oreomyrrhis eriopoda, Luzula meridionalis subsp. Flaccida, Poa hothamensis var. hothamensis, and Senecio gunnii in Sub-Alpine Woodland (Table S9).
3.5. Vegetation Type and Fire Frequency Effects—Functional Types
The relative proportion of species with different life forms and dispersal mechanisms varied significantly with vegetation type, with consistently greater proportions of phanerophytes in the extant vegetation and anemochores in the soil seed bank (Tables S10 and S11). For Shrubby Dry Forest, the fire response strategies also differed with vegetation type; resprouters were more frequent in the extant vegetation and seeders were more frequent in the soil seed bank (Table S10).
The relative proportion of species with different life forms and dispersal modes also varied significantly with fire frequency in Shrubby Dry Forest, with that in double-burnt sites being significantly different to that in unburnt and single-burnt (2013) sites, but not single-burnt (2007) sites with a longer time since fire (Table S10). Specifically, there was an increase in the proportions of phanerophytes, therophytes, erect rosettes, and anemochores with an increasing fire frequency, while there was a decrease in the proportion of barochores (Table S10). In Sub-Alpine Woodland, there were no significant differences in the relative proportions of functional trait associations with fire frequency outside a trend for near-significant (p = 0.06) changes in life forms (Table S11).
4. Discussion
Fire regime shifts in the form of short-interval, high-severity wildfires modify both the soil seed stores and extant vegetation in Shrubby Dry Forest and Sub-Alpine Woodland communities in southeastern Australia. Our results were largely consistent with our first hypothesis that seedling emergence was promoted by fire-related germination cues, and that these cues were related to the post-fire regeneration strategy, mode of seed dispersal, and life form and were more prevalent in burnt than long-unburnt sites. Consistent with our expectations, we found that the soil seed bank and extant vegetation demonstrated similar responses to fire frequency, which included an increase in species with a fast reproductive maturity and a reduced capacity of the soil seed bank to buffer against species loss in extant vegetation following double and triple high-severity fires.
4.1. Fire-Related Germination Cues
Fire cues promoted the germination of the soil seed store in both forest types, and for Sub-Alpine Woodland, was limited to areas more frequently burnt by fire, where the heat plus smoke treatments increased the density of germinants relative to untreated samples. For Shrubby Dry Forest, the heat plus smoke treatments increased the species richness and germinant density relative to the control and smoke-only treatments and were concentrated in phanerophytes, barochores, resprouters, and ant-dispersed species with hard seed coats. For both forest types, the germination of several shrub species with hard seed coats was limited to the heat plus smoke treatments. There was no effect of smoke in isolation and, contrary to our hypothesis, fire cues did not enhance the germination of short-lived species outside of positive effects on the introduced annual Erigeron bonariense, with positive effects also for other introduced species, including the perennials Cerastium vulgare and Acetocella vulgaris.
Several species showed no response to fire cues, including 27 of 35 individual species in Shrubby Dry Forest and 13 of 18 species in Sub-Alpine Woodland. These were often non-woody species or early successional shrubs (e.g., Cassinia spp.) with persistent seed banks. The results for individual species were consistent for some species common to both forest types (e.g., Luzula meridionalis subsp. flaccida, Laphangium luteoalbum, and Wahlenbergia multicaulis) and with other studies (e.g., Cassinia spp. [34,92]), but not consistent for other species (e.g., Cerastium vlugare and Poranthera microphylla [34,92]), providing support for regional variations in germination cues [26]. The low incidence of fire-cued germination in non-woody species agrees with work in other southeastern Australian forest types [34,103], including alpine treeline ecotones [104], and arguments for a greater reliance on resprouting or gap-phase recruitment in species lacking post-fire seedling recruitment [104,105,106,107]. This non-fire-released dormancy provides a bet-hedging strategy, where long-lived seed can germinate once suitable (non-fire) conditions emerge [108].
The requirement for fire cues in hard-seeded species, including many from Fabaceae, is consistent with the requirement for high temperatures to break physical dormancy [18]. Each of the 54 hard-seeded species across both forest types were ant-dispersed and consistent with our hypothesis, suggesting that seed burial by ants serves to protect physically dormant seeds from lethal temperatures at the soil surface [36,108]. Seed burial may also serve to increase the post-fire residual seed bank if the soil temperatures do not reach the thresholds needed to break physical dormancy [58]. The concentration of myrmecochores in Shrubby Dry Forest (38 species), with a historically shorter fire-return interval than Sub-Alpine Woodland (18 species), also supports arguments by Pausas and Lamont [108] that myrmecochory should be expressed more often in fire-prone habitats if its primary purpose is to break dormancy and stimulate germination.
Across both forest types, the lack of response to smoke independent of heat is consistent with a lower incidence of smoke-cued germination in southeastern Australian flora relative to fire-prone areas elsewhere [19], despite broad global trends of increases in both seed bank richness and abundance in response to simulated smoke [109]. For obligate seeders, the lack of response to smoke in the absence of heat contrasts broad trends in southeastern Australian flora [19], although the requirement for both smoke and heat is more consistent with the conditions encountered during the passage of a wildfire [109]. This lack of response to smoke may have been related to the method of application, with evidence for a greater effectiveness with aerosol smoke [19] or the application of the active smoke compound karrakinolide [110], although our recent work examining the method of smoke application suggests this is unlikely [77].
Across both forest types, several species present in extant vegetation were absent from the soil seed bank, including 18 of 34 hard-seeded ant-dispersed species (e.g., Acacia spp., Bossiaea spp., and Grevillea spp.) and 17 of 19 endozoochorous phanerophytes (e.g., Acrothamnus spp., Coprosma spp., Persoonia spp., and Tasmannia spp.). The absence of a number of these species is consistent with other work [34,60,75] and may be associated with rarity (e.g., the endangered Grevillea victoriae subsp. victoriae), an insufficient fire return interval to replenish the soil seed bank stores of obligate seeding species (e.g., 5–10 year juvenile period for Acacia obliquinervia and Tasmania xerophila subsp. xerophila [111]), or transient or short-term soil seed banks [108]. For example, Coprosma quadrifida can establish from seed post-fire [112], but likely has a short-term (<2 years) soil seed bank, with previous work demonstrating frequent germination from fresh seed collected in seed rain but absent from complementary samples of the soil seed bank (S. Kasel, unpublished data). Short-lived seed is notable among obligate resprouters with fleshy fruit (e.g., Acotriche serrulata)—these species take advantage of the long-distance dispersal provided by vertebrates rather than post-fire conditions for recruitment [108]. Seed may also be present in the soil seed bank but ungerminated due to a lack of suitable germination cues; for example, Persoonia spp. are known for their complex seed germination requirements [113]. The germination of Pelargonium helmsii in the smoke-only and LHS treatments and absence in extant vegetation are consistent with previous work, suggesting that the species is fire-dependent but short-lived and rare in extant vegetation [114], with a persistent soil seed bank providing for its partial germination following fire [111].
Differences in responses to fire cues between the two forest types may have been related to differences in fire history. Shrubby Dry Forest has a shorter historical fire return interval (10–20 years) than Sub-Alpine Woodland (50–100 years). Similar results have been reported for Mediterranean woodlands, where the response of soil seed bank communities to the application of smoke was more pronounced in sites with a history of more frequent fire [24]. Elsewhere, the time since fire effects on the application of smoke and/or heat to soil seed banks of heathland communities included both positive and negative effects according to species and functional groups [115]. Our results add to the body of evidence demonstrating fire effects, and here, germination response to fire cues is more common in more fire-prone areas that favour species stimulated by fire [116].
4.2. Fire Frequency Effects on the Soil Seed Bank and Extant Vegetation
Short-interval, high-severity wildfires produce unequivocal shifts in understorey diversity. There was a decline in the species richness in both the soil seed bank and extant vegetation with more frequent, severe wildfires. This was mirrored by a decline in H′ in the soil seed bank, with changes in H′ in the extant vegetation limited to Sub-Alpine Woodland, although there was a trend of declines in H′ in double-burnt sites relative to other fire histories in Shrubby Dry Forest. For the soil seed bank, depletions in richness and H′ are consistent with the demonstrated promotion of germination via fire cues and conditions of increased resource availability after fire [117]. For example, Fairman et al. [43,63] demonstrated that short-interval wildfires reduced seedling regeneration and increased mortality in the fire-tolerant eucalypt overstorey of our study sites, providing for structurally mediated effects on resource availability and understorey diversity [34,118,119,120].
The decline in richness and H′ in both the soil seed bank and extant vegetation suggests that the soil seed bank has lost its capacity to buffer species’ loss in the extant vegetation [57,121], and that this change is more apparent in Sub-Alpine Woodland with a historically longer fire return interval than Shrubby Dry Forest. Duivenvoorden et al. [60] reported similar declines in richness and H′ in the soil seed bank following double and triple short-interval wildfires in both obligate seeder and fire-tolerant montane eucalypt forests, although this decline in alpha diversity was not reflected in extant vegetation, despite similar historical fire return intervals to the Sub-Alpine Woodland. This maintenance in alpha diversity in the extant vegetation was purported by Duivenvoorden et al. [60] to be an example of a “resilience debt” (sensu [122]). For the eucalypt-dominated communities in this study, the concomitant decline in alpha diversity in tje extant vegetation and the soil seed bank suggests that this resilience debt has been realized.
An increased fire frequency also shifted the plant community composition (beta diversity), with the greatest differences occurring between the most frequently burnt and unburnt sites, and these differences were consistent for both forest types and between extant vegetation and the soil seed bank, despite a low similarity between species’ pools. These findings are consistent with those for soil seed banks and extant vegetation in other forest types in the region, including montane obligate seeding and resprouting eucalypt forests [60] and coastal woodlands [54]. For lodgepole pine (Pinus contorta var. contorta)-dominated forests in Yellowstone National Park, the dissimilarity between short- and long-interval fires was context-dependent and diminished with time [120], consistent with our results for Shrubby Dry Forest, where differences between double- and single-burnt sites were limited to the most recently burnt sites. For both forest types, species that demonstrated strong positive responses to more frequent fire and made the strongest contributions to differences in community composition demonstrated multiple adaptations to fire, including fire-cued germination, the capacity to resprout and form long-term persistent soil seed banks, and a relative short time (3–5 years) to seed production (e.g., Daviesia buxifolia and Poa hothamensis var. hothamensis).
The fire frequency effects on the relative richness of trait associations were restricted to Shrubby Dry Forest, and, consistent with our hypothesis, included increased proportions of predominantly obligate seeding therophytes and wind-dispersed species in burnt relative to long-unburnt sites in both extant vegetation and the soil seed bank. These results echo those in other studies [60] and can be explained by the conditions of increased resource availability, reduced herbivory and seed predation, and greater exposure of mineral soil following severe fires, which can favour recolonizing species or those emerging from the seed bank [123,124,125]. There was also an increase in ant-dispersed phanerophytes in double-burnt sites that was driven by Daviesia buxifolia, a resprouting shrub with a short juvenile period. This contrasts the reduced recruitment and increased mortality in the resprouting eucalypt canopy with canopy-stored seed [63]. Duivenvoorden et al. [60] also reported an increase in understorey shrubs with persistent soil seed stores and a short time to reproductive maturity following more frequent fires for both obligate seeding and resprouting montane eucalypt forests. Similarly, in Californian shrublands, a higher fire frequency promotes the recruitment of resprouting shrubs and annual grasses but leads to a decline in obligate seeding shrubs with a longer time to reproductive maturity [49,59]. Consistent with the predictions of Pausas and Keeley [117], our reported increase in obligate seeding therophytes suggests that, for temperate climates, frequent fires can favour obligate seeders where immaturity risk is low or absent.
There was an absence of fire frequency effects on the relative frequency of traits in Sub-Alpine Woodland in both extant vegetation and the soil seed bank, despite shifts in alpha and beta diversity and previous work on the extant vegetation of these forests reporting a decrease in shrub prominence and increase in grass frequency with more frequent high-severity fires [43]. For the soil seed bank, there was, however, a decline in the abundance of ant-dispersed species and an increase in the abundance in erect rosettes with more frequent fires, and a near-significant trend (p = 0.06) for increases in the relative frequency of erect rosettes and declines in myrmecochores in both the soil seed bank and extant vegetation. Increases in the abundance of graminoids have been reported in subalpine forests elsewhere [120] and in other montane forests in southeastern Australia, including those dominated by Eucalyptus delegatensis [60,126] and E. regnans [127], as well as mixed-species forests [60]. Like other montane forests [60,75,104], erect rosettes were the most abundant functional group in the soil seed bank in Sub-Alpine Woodland, driven by the high seed production of a few species (Poa hothamensis var. hothamensis, Carex breviculmis, and Luzula meridionalis subsp. flaccida; see also [75]). Abundant soil seed banks, combined with conditions of increased resource availability and other previously mentioned mechanisms, favour the recruitment of these species.
For Sub-Alpine Woodland, the decline in myrmecochores reflects poor representation in the soil seed bank (7 of 18 species). The absence of obligate seeding species capable of producing persistent seed banks such as Acacia obliquinervia and A. alpina points to the combined effect of fire-cue germination and short-interval fires in eroding the residual seed store [58,60]. We cannot discount likely residual seed stores at greater soil depths (>5 cm) given the decline in fire-related germination cues—including lethal temperatures—with depth, although there is a lower abundance of seed with depth and a reduced capacity for seeds to emerge successfully [17]. More broadly, the poor representation of phanerophytes in the soil seed bank is likely to produce further changes in the occurrence and abundance of trees and shrubs in the extant vegetation, given the predicted intensification of fire regimes across southeastern Australia [13]. These changes are likely more severe for Sub-Alpine Woodland than Shrubby Dry Forest, given the poorer representation of phanerophytes in the soil seed bank (9 of 38 species vs. 31 of 64, respectively).
For both Shrubby Dry Forest and Sub-Alpine Woodland, there was no change in the relative frequency of fire response strategies in extant vegetation and the soil seed bank. This contrasts broader relationships for an increase in frequency in resprouting species with a shorter fire return interval, although such relationships are based on longer (>10 years) fire return intervals [128]. During high-severity fires, resprouters may be killed outright [123], and repeated, short-interval, high-severity fires may erode resprouting capacity due to a depletion in carbon reserves [63], as well as reducing benefits associated with carbon allocation to regrowth and meristem formation over seed production [128,129]. Duivenvoorden et al. [60] reported such a decline in clonal resprouters with an increase in the frequency of high-severity wildfires with a short (<10 years) return interval. However, vegetation recovery is not a sole consequence of regeneration strategy, and for these study sites, may be related to post-fire environmental conditions (e.g., drought, heat stress, and insect outbreaks) that can be just as important for vegetation recovery [130,131]. For example, two of the wildfires took place during the ‘Millennium Drought’, a period of extended drought in southeastern Australia between 1996 and 2010 that led to widespread failure in eucalypt regeneration with flow-on effects on understorey diversity [132,133].
5. Conclusions
Clear shifts in plant diversity in extant vegetation and soil seed banks, including a decline in alpha diversity and changes in species composition and functional trait associations, provide strong evidence for changes in plant diversity with emerging fire regimes. The largely consistent changes in plant diversity in both forest types, despite contrasting historical fire regimes, suggests that severe wildfires with short fire return intervals are pushing ecosystems beyond their historical range of variability, with the realization of a previously hidden resilience debt. Shifts in species composition, including the potential for an increased flammability of these forests and positive feedback between past fires and future fires [120,134,135], point to a likely increase in extent in areas burnt at even higher frequencies, with some areas in southeastern Australia already being burnt four to five times since 1995 [136]. The erosion in the buffering capacity of soil seed banks points to an increase in the reliance on post-fire resprouting, clonal spread, or long-distance dispersal from fire refugia for maintaining plant diversity, particularly in extensively burnt areas such as those in southeastern Australia.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fire7040148/s1, Table S1: Fire history for each site sampled as part of this study. Table S2: Description of plant traits and their defining attributes. Table S3: Total seedling density (germinants m−2) per treatment (C, control; S, smoke only; LHS, low heat plus smoke; and HHS, high heat plus smoke) for all species in soils from Shrubby Dry Forest and Sub-Alpine Woodland. Table S4: Results of the PERMANOVA, testing for the effect of germination treatment (T: C, control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke) and fire frequency (F, number of fires since 2003) on the richness and density of the germinable soil seed bank in Shrubby Dry Forest and Sub-Alpine Woodland communities. Table S5: Results of the PERMANOVA, testing for the effect of germination treatment (T, control, smoke, low heat plus smoke, and high heat plus smoke) and fire frequency (F, number of fires since 2003) on Shannon’s diversity (H′) of the germinable soil seed bank in Shrubby Dry Forest and Sub-Alpine Woodland communities. Table S6: Results of the PERMANOVA and SIMPER analysis, testing for the effect of germination treatment (T, control, smoke, low heat plus smoke, and high heat plus smoke) and fire frequency (F, number of fires since 2003) on species composition of the germinable soil seed bank in Shrubby Dry Forest and Sub-Alpine Woodland communities. Table S7: Results of the PERMANOVA, testing for the effect of vegetation type (V: Veg, extant vegetation; SSB, soil seed bank) and fire frequency (F, number of fires since 2003) on species richness in Shrubby Dry Forest and Sub-Alpine Woodland communities. Table S8: Results of the PERMANOVA, testing for the effect of fire frequency (F, number of fires since 2003) on H′ in Shrubby Dry Forest and Sub-Alpine Woodland communities. Table S9: Results of the PERMANOVA and SIMPER analysis, testing for the effect of vegetation type (V; Veg, Extant Vegetation; SSB, Soil Seed Bank) and fire frequency (F, number of fires since 2003) on species composition Shrubby Dry Forest and Sub-Alpine Woodland communities. Table S10: Significance of Vegetation Type and Fire Frequency effects on trait associations for Shrubby Dry Forest. For each trait, there was no significant (p > 0.05) Vegetation Type × Fire Frequency interaction. Table S11: Significance of Vegetation Type and Fire Frequency effects on trait associations for Sub-Alpine Woodland. Figure S1: Map of the study area showing the distribution of the two forest types and location of the study sites. Figure S2: Examples of the study plots of the two forest types burnt at different frequencies. Figure S3: Shannon’s Diversity (H′) of all species (a), native species (b), and introduced species (c) in the soil seed bank according to Fire Frequency for Shrubby Dry Forest and Sub-Alpine Woodland. Figure S4: Total aggregated species richness (across all sites) for traits within each plant functional type (dispersal mode, life form, fire response strategy). Figure S5: Shannon’s Diversity (H′) of all species (a), native species (b), and introduced species (c) in the extant vegetation and soil seed bank (combined pool across all treatments) according to Fire Frequency for (a) Shrubby Dry Forest and (b) Sub-Alpine Woodland.
Author Contributions
Conceptualization, S.K., T.A.F. and C.R.N.; methodology, S.K., T.A.F. and C.R.N.; formal analysis, S.K.; data curation, S.K.; investigation, S.K. and T.A.F.; writing—original draft preparation, S.K.; writing—review and editing, S.K., T.A.F. and C.R.N.; visualization, S.K.; funding acquisition, S.K. and C.R.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Victorian Government Department of Energy, Environment and Climate Action Integrated Forest Ecosystem Research Program, agreement numbers 37536 and 301103.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting the findings of this study are available from the corresponding author upon reasonable request. The data are not publicly available due to privacy restrictions.
Acknowledgments
This research was undertaken with approval by the Victorian Department of Energy, Environment and Climate Action (DEECA) under research permit number 10008476. We thank Ben Smith and David Lockwood for field assistance and David Lockwood for providing expertise in plant identification. We thank Rowan Berry, Sascha Andrusiak and Brett Hough for maintaining the glasshouse facilities.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bowman, D.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P.; et al. Fire in the Earth System. Science 2009, 324, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Gill, A.M. Fire and The Australian Flora: A Review. Aust. For. 1975, 38, 4–25. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. A Burning Story: The Role of Fire in the History of Life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef]
- Bradstock, R.A. A biogeographic model of fire regimes in Australia: Current and future implications. Glob. Ecol. Biogeogr. 2010, 19, 145–158. [Google Scholar] [CrossRef]
- Clarke, H.; Evans, J.P. Exploring the future change space for fire weather in southeast Australia. Theor. Appl. Climatol. 2019, 136, 513–527. [Google Scholar] [CrossRef]
- Flannigan, M.; Cantin, A.S.; de Groot, W.J.; Wotton, M.; Newbery, A.; Gowman, L.M. Global wildland fire season severity in the 21st century. For. Ecol. Manag. 2013, 294, 54–61. [Google Scholar] [CrossRef]
- Stephens, S.L.; Burrows, N.; Buyantuyev, A.; Gray, R.W.; Keane, R.E.; Kubian, R.; Liu, S.; Seijo, F.; Shu, L.; Tolhurst, K.G.; et al. Temperate and boreal forest mega-fires: Characteristics and challenges. Front. Ecol. Environ. 2014, 12, 115–122. [Google Scholar] [CrossRef]
- Nolan, R.H.; Bowman, D.M.J.S.; Clarke, H.; Haynes, K.; Ooi, M.K.J.; Price, O.F.; Williamson, G.J.; Whittaker, J.; Bedward, M.; Boer, M.M.; et al. What Do the Australian Black Summer Fires Signify for the Global Fire Crisis? Fire 2021, 4, 97. [Google Scholar] [CrossRef]
- Collins, L.; Clarke, H.; Clarke, M.F.; McColl Gausden, S.C.; Nolan, R.H.; Penman, T.; Bradstock, R. Warmer and drier conditions have increased the potential for large and severe fire seasons across south-eastern Australia. Glob. Ecol. Biogeogr. 2022, 31, 1933–1948. [Google Scholar] [CrossRef]
- Enright, N.J.; Fontaine, J.B.; Bowman, D.; Bradstock, R.A.; Williams, R.J. Interval squeeze: Altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 2015, 13, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Fairman, T.A.; Nitschke, C.R.; Bennett, L.T. Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. Int. J. Wildland Fire 2016, 25, 831–848. [Google Scholar] [CrossRef]
- McColl-Gausden, S.C.; Bennett, L.T.; Ababei, D.A.; Clarke, H.G.; Penman, T.D. Future fire regimes increase risks to obligate-seeder forests. Divers. Distrib. 2022, 18, 542–558. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Whelan, R.J. The Ecology of Fire; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Keeley, J.F. Resilience of Mediterranean shrub communities to fires. In Resilience in Mediterranean-Type Ecosystems; Dell, B., Hopkins, J.J.M., Lamont, B.B., Eds.; Dr. W. Junk Publishers: Dordrecht, The Netherlands, 1986; pp. 95–112. [Google Scholar]
- Auld, T.D.; Denham, A.J. How much seed remains in the soil after a fire? Plant Ecol. 2006, 187, 15–24. [Google Scholar] [CrossRef]
- Auld, T.D.; O’Connell, M.A. Predicting patterns of post-fire germination in 35 eastern Australian Fabaceae. Aust. J. Ecol. 1991, 16, 53–70. [Google Scholar] [CrossRef]
- Carthey, A.J.R.; Tims, A.; Geedicke, I.; Leishman, M.R. Broad-scale patterns in smoke-responsive germination from the south-eastern Australian flora. J. Veg. Sci. 2018, 29, 737–745. [Google Scholar] [CrossRef]
- Enright, N.J.; Kintrup, A. Effects of smoke, heat and charred wood on the germination of dormant soil-stored seeds from a Eucalyptus baxteri heathy-woodland in Victoria, SE Australia. Austral Ecol. 2001, 26, 132–141. [Google Scholar] [CrossRef]
- Noble, I.R.; Slatyer, R.O. The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 1980, 43, 5–21. [Google Scholar] [CrossRef]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Bradbury, D.; Tapper, S.L.; Coates, D.; McArthur, S.; Hankinson, M.; Byrne, M. The role of fire and a long-lived soil seed bank in maintaining persistence, genetic diversity and connectivity in a fire-prone landscape. J. Biogeogr. 2016, 43, 70–84. [Google Scholar] [CrossRef]
- Manela, N.; Dagon, E.; Semesh, H.; Ovadia, O. Smoke interacts with fire history to stimulate soil seed bank germination in Mediterranean woodlands. J. Plant Ecol. 2019, 12, 419–427. [Google Scholar] [CrossRef]
- Zaki, E.; Abedi, M.; Naqinezhad, A. How fire history affects germination cues of three perennial grasses from the mountain steppes of Golestan National Park. Flora 2021, 280, 151835. [Google Scholar] [CrossRef]
- Maikano, G.N.; Cohn, J.; Di Stefano, J. Are germination cues for soil-stored seed banks different in structurally different fire-prone communities? Austral Ecol. 2018, 43, 89–101. [Google Scholar] [CrossRef]
- Paula, S.; Pausas, J.G. Burning seeds: Germinative response to heat treatments in relation to resprouting ability. J. Ecol. 2008, 96, 543–552. [Google Scholar] [CrossRef]
- Tormo, J.; Moreira, B.; Pausas, J.G.; Vandvik, V. Field evidence of smoke-stimulated seedling emergence and establishment in Mediterranean Basin flora. J. Veg. Sci. 2014, 25, 771–777. [Google Scholar] [CrossRef]
- Keeley, J.E.; Bond, W.J. Convergent seed germination in South African fynbos and Californian chaparral. Plant Ecol. 1997, 133, 153–167. [Google Scholar] [CrossRef]
- Read, T.R.; Bellairs, S.M.; Mulligan, D.R.; Lamb, D. Smoke and heat effects on soil seed bank germination for the re-establishment of a native forest community in New South Wales. Austral Ecol. 2000, 25, 48–57. [Google Scholar] [CrossRef]
- Santana, V.M.; Alday, J.G.; Baeza, M.J. Effects of fire regime shift in Mediterranean Basin ecosystems: Changes in soil seed bank composition among functional types. Plant Ecol. 2014, 215, 555–566. [Google Scholar] [CrossRef]
- Ne’eman, G.; Ne’eman, R.; Keith, D.A.; Whelan, R.J. Does post-fire plant regeneration mode affect the germination response to fire-related cues? Oecologia 2009, 159, 483–492. [Google Scholar] [CrossRef]
- Merritt, D.J.; Turner, S.R.; Clarke, S.; Dixon, K.W. Seed dormancy and germination stimulation syndromes for Australian temperate species. Aust. J. Bot. 2007, 55, 336–344. [Google Scholar] [CrossRef]
- Kasel, S.; Nitschke, C.R.; Baker, S.C.; Pryde, E.C. Concurrent assessment of functional types in extant vegetation and soil seed banks informs environmental constraints and mechanisms of plant community turnover in temperate forests of south-eastern Australia. For. Ecol. Manag. 2022, 519, 120321. [Google Scholar] [CrossRef]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C.; Hidayati, S.N. Defining transient and persistent seed banks in species with pronounced seasonal dormancy and germination patterns. Seed Sci. Res. 2005, 15, 189–196. [Google Scholar] [CrossRef]
- Berg, R.Y. Myrmecochorous plants in Australia and their dispersal by ants. Aust. J. Bot. 1975, 23, 475–508. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Bradstock, R.A.; Auld, T.D. Soil Temperatures During Experimental Bushfires in Relation to Fire Intensity: Consequences for Legume Germination and Fire Management in South-Eastern Australia. J. Appl. Ecol. 1995, 32, 76–84. [Google Scholar] [CrossRef]
- Bradstock, R.A.; Tozer, M.G.; Keith, D.A. Effects of high frequency fire on floristic composition and abundance in a fire-prone heathland near Sydney. Aust. J. Bot. 1997, 45, 641–655. [Google Scholar] [CrossRef]
- Pausas, J.G. Response of plant functional types to changes in the fire regime in Mediterranean ecosystems: A simulation approach. J. Veg. Sci. 1999, 10, 717–722. [Google Scholar] [CrossRef]
- Bond, W.J.; van Wilgen, B.W. Fire and Plants: Populationand Community Biology Series 14; Chapman and Hall: London, UK, 1996. [Google Scholar]
- Collins, L. Eucalypt forests dominated by epicormic resprouters are resilient to repeated canopy fires. J. Ecol. 2020, 108, 310–324. [Google Scholar] [CrossRef]
- Fairman, T.A.; Bennett, L.T.; Tupper, S.; Nitschke, C.R. Frequent wildfires erode tree persistence and alter stand structure and initial composition of a fire-tolerant sub-alpine forest. J. Veg. Sci. 2017, 28, 1151–1165. [Google Scholar] [CrossRef]
- Buma, B.; Brown, C.D.; Donato, D.C.; Fontaine, J.B.; Johnstone, J.F. The Impacts of Changing Disturbance Regimes on Serotinous Plant Populations and Communities. Bioscience 2013, 63, 866–876. [Google Scholar] [CrossRef]
- Vila-Cabrera, A.; Saura-Mas, S.; Lloret, F. Effects of fire frequency on species composition in a Mediterranean shrubland. Ecoscience 2008, 15, 519–528. [Google Scholar] [CrossRef]
- Zedler, P.H.; Gautier, C.R.; McMaster, G.S. Vegetation Change in Response to Extreme Events: The Effect of a Short Interval between Fires in California Chaparral and Coastal Scrub. Ecology 1983, 64, 809–818. [Google Scholar] [CrossRef]
- Keeley, J.E.; Brennan, T.J. Fire-driven alien invasion in a fire-adapted ecosystem. Oecologia 2012, 169, 1043–1052. [Google Scholar] [CrossRef]
- Santana, V.M.; Baeza, M.J.; Marrs, R.H.; Vallejo, V.R. Old-field secondary succession in SE Spain: Can fire divert it? Plant Ecol. 2010, 211, 337–349. [Google Scholar] [CrossRef]
- Syphard, A.D.; Brennan, T.J.; Keeley, J.E. Drivers of chaparral type conversion to herbaceous vegetation in coastal Southern California. Divers. Distrib. 2019, 25, 90–101. [Google Scholar] [CrossRef]
- Chick, M.P.; Cohn, J.S.; Nitschke, C.R.; York, A. Lack of soil seedbank change with time since fire: Relevance to seed supply after prescribed burns. Int. J. Wildland Fire 2016, 25, 849–860. [Google Scholar] [CrossRef]
- Paula, S.; Arianoutsou, M.; Kazanis, D.; Tavsanoglu, Ç.; Lloret, F.; Buhk, C.; Ojeda, F.; Luna, B.; Moreno, J.M.; Rodrigo, A.; et al. Fire-related traits for plant species of the Mediterranean Basin. Ecology 2009, 90, 1420. [Google Scholar] [CrossRef]
- Lewis, T.; Reif, M.; Prendergast, E.; Tran, C. The effect of long-term repeated burning and fire exclusion on above- and below-ground Blackbutt (Eucalyptus pilularis) forest vegetation assemblages. Austral Ecol. 2012, 37, 767–778. [Google Scholar] [CrossRef]
- Penman, T.D.; Binns, D.; Shiels, R.; Allen, R.; Penman, S. Hidden effects of forest management practices: Responses of a soil stored seed bank to logging and repeated prescribed fire. Austral Ecol. 2011, 36, 571–580. [Google Scholar] [CrossRef]
- Chick, M.P.; Nitschke, C.R.; Cohn, J.S.; Penman, T.D.; York, A. Factors influencing above-ground and soil seed bank vegetation diversity at different scales in a quasi-Mediterranean ecosystem. J. Veg. Sci. 2018, 29, 684–694. [Google Scholar] [CrossRef]
- Chick, M.P.; York, A.; Sitters, H.; Di Stefano, J.; Nitschke, C.R. Combining optimization and simulation modelling to measure the cumulative impacts of prescribed fire and wildfire on vegetation species diversity. J. Appl. Ecol. 2019, 56, 722–732. [Google Scholar] [CrossRef]
- Van der Veken, S.; Bellemare, J.; Verheyen, K.; Hermy, M. Life-history traits are correlated with geographical distribution patterns of western European forest herb species. J. Biogeogr. 2007, 34, 1723–1735. [Google Scholar] [CrossRef]
- Vandvik, V.; Klanderud, K.; Meineri, E.; Måren, I.E.; Töpper, J. Seed banks are biodiversity reservoirs: Species–area relationships above versus below ground. Oikos 2016, 125, 218–228. [Google Scholar] [CrossRef]
- Palmer, H.D.; Denham, A.J.; Ooi, M.K.J. Fire severity drives variation in post-fire recruitment and residual seed bank size of Acacia species. Plant Ecol. 2018, 219, 527–537. [Google Scholar] [CrossRef]
- Syphard, A.D.; Franklin, J.; Keeley, J.E. Simulating the effects of frequent fire on southern California coastal shrublands. Ecol. Appl. 2006, 16, 1744–1756. [Google Scholar] [CrossRef] [PubMed]
- Duivenvoorden, E.; Wagner, B.; Nitschke, C.R.; Kasel, S. Short-interval, high-severity wildfires cause declines in soil seed bank diversity in montane forests of south-eastern Australia. For. Ecol. Manag. 2024, 553, 121627. [Google Scholar] [CrossRef]
- Díaz, S.; Cabido, M. Plant Functional Types and Ecosystem Function in Relation to Global Change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Lipoma, M.L.; Funes, G.; Diaz, S. Fire effects on the soil seed bank and post-fire resilience of a semi-arid shrubland in central Argentina. Austral Ecol. 2018, 43, 46–55. [Google Scholar] [CrossRef]
- Fairman, T.A.; Bennett, L.T.; Nitschke, C.R. Short-interval wildfires increase likelihood of resprouting failure in fire-tolerant trees. J. Environ. Manag. 2019, 231, 59–65. [Google Scholar] [CrossRef]
- Stewart, S.B.; Nitschke, C.R. Improving temperature interpolation using MODIS LST and local topography: A comparison of methods in south east Australia. Int. J. Climatol. 2017, 37, 3098–3110. [Google Scholar] [CrossRef]
- DEECA. Victorian Ecological Vegetation Communities. Available online: https://www.environment.vic.gov.au/biodiversity/bioregions-and-evc-benchmarks (accessed on 31 May 2023).
- Murphy, B.P.; Bradstock, R.A.; Boer, M.M.; Carter, J.; Cary, G.J.; Cochrane, M.A.; Fensham, R.J.; Russell-Smith, J.; Williamson, G.J.; Bowman, D. Fire regimes of Australia: A pyrogeographic model system. J. Biogeogr. 2013, 40, 1048–1058. [Google Scholar] [CrossRef]
- Cheal, D. Growth Stages and Tolerable Fire Intervals for Victoria’s Native Vegetation Data Sets. Fire and Adaptive Management Report No. 84; Department of Sustainability and Environment: East Melbourne, VIC, Australia, 2010. [Google Scholar]
- Volkova, L.; Weiss Aparicio, A.G.; Weston, C.J. Fire intensity effects on post-fire fuel recovery in Eucalyptus open forests of south-eastern Australia. Sci. Total Environ. 2019, 670, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Costermans, L. Native Trees and Shrubs of South-Eastern Australia; New Holland: Melbourne, VIC, Australia, 2009. [Google Scholar]
- Ferrar, P.J.; Cochrane, P.M.; Slatyer, R.O. Factors influencing germination and establishment of Eucalyptus pauciflora near the alpine tree line. Tree Physiol. 1988, 4, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Green, K. Causes of stability in the alpine treeline in the Snowy Mountains of Australia a natural experiment. Aust. J. Bot. 2009, 57, 171–179. [Google Scholar] [CrossRef]
- Slatyer, R.O.; Noble, I.R. Dynamics of montane treelines. In Landscape Boundaries; Hansen, A.J., Di Castri, F., Eds.; Springer: New York, NY, USA, 1992. [Google Scholar]
- Williams, R.J.; Wahren, C.-H.; Tolsma, A.D.; Sanecki, G.M.; Papst, W.A.; Myers, B.A.; McDougall, K.L.; Heinze, D.A.; Green, K. Large fires in Australian alpine landscapes: Their part in the historical fire regime and their impacts on alpine biodiversity. Int. J. Wildland Fire 2009, 17, 793–808. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Cavieres, L.A.; Carmen, C.; Humaña, A.M. Persistent Soil Seed Bank and Standing Vegetation at a High Alpine Site in the Central Chilean Andes. Oecologia 1999, 119, 126–132. [Google Scholar]
- Venn, S.E.; Morgan, J.W. Soil seedbank composition and dynamics across alpine summits in south-eastern Australia. Aust. J. Bot. 2010, 58, 349–362. [Google Scholar] [CrossRef]
- Lipoma, M.L.; Fortunato, V.; Enrico, L.; Díaz, S. Where does the forest come back from? Soil and litter seed banks and the juvenile bank as sources of vegetation resilience in a semiarid Neotropical forest. J. Veg. Sci. 2020, 31, 1017–1027. [Google Scholar] [CrossRef]
- Younis, S.; Kasel, S. Do Fire Cues Enhance Germination of Soil Seed Stores across an Ecotone of Wet Eucalypt Forest to Cool Temperate Rainforest in the Central Highlands of South-Eastern Australia? Fire 2023, 6, 138. [Google Scholar] [CrossRef]
- Beadle, N.C.W. Soil Temperatures during Forest Fires and Their Effect on the Survival of Vegetation. J. Ecol. 1940, 28, 180–192. [Google Scholar] [CrossRef]
- Warcup, J. Effect of heat treatment of forest soil on germination of buried seed. Aust. J. Bot. 1980, 28, 567–571. [Google Scholar] [CrossRef]
- Meers, T.L.; Enright, N.J.; Bell, T.L.; Kasel, S. Deforestation strongly affects soil seed banks in eucalypt forests: Generalisations in functional traits and implications for restoration. For. Ecol. Manag. 2012, 266, 94–107. [Google Scholar] [CrossRef]
- Hoyle, G.L.; Venn, S.E.; Steadman, K.J.; Good, R.B.; McAuliffe, E.J.; Williams, E.R.; Nicotra, A.B. Soil warming increases plant species richness but decreases germination from the alpine soil seed bank. Glob. Chang. Biol. 2013, 19, 1549–1561. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Brown, D. Estimating the composition of a forest seed bank: A comparison of the seed extraction and seedling emergence methods. Can. J. Bot. 1992, 70, 1603–1612. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: Oxford, UK, 1934. [Google Scholar]
- McIntyre, S.; Lavorel, S.; Tremont, R.M. Plant life-history attributes: Their relationships to disturbance response in herbaceous vegetation. J. Ecol. 1995, 83, 31–44. [Google Scholar] [CrossRef]
- Leishman, M.R.; Westoby, M.; Jurado, E. Correlates of Seed Size Variation: A Comparison among Five Temperate Floras. J. Ecol. 1995, 83, 517–529. [Google Scholar] [CrossRef]
- NPWS. NSW Flora Fires Response Database, Version 1.3a; NSW National Parks and Wildlife Service: Hurstville, NSW, Australia, 2002. [Google Scholar]
- Vivian, L.M.; Doherty, M.D.; Cary, G.J. Classifying the fire-response traits of plants: How reliable are species-level classifications? Austral Ecol. 2010, 35, 264–273. [Google Scholar] [CrossRef]
- Pausas, J.G.; Pratt, R.B.; Keeley, J.E.; Jacobsen, A.L.; Ramirez, A.R.; Vilagrosa, A.; Paula, S.; Kaneakua-Pia, I.N.; Davis, S.D. Towards understanding resprouting at the global scale. New Phytol. 2016, 209, 945–954. [Google Scholar] [CrossRef]
- Vesk, P.A.; Westoby, M. Sprouting ability across diverse disturbances and vegetation types worldwide. J. Ecol. 2004, 92, 310–320. [Google Scholar] [CrossRef]
- Penman, T.D.; Binns, D.; Allen, R.; Shiels, R.; Plummer, S. Germination responses of a dry sclerophyll forest soil-stored seedbank to fire related cues. Cunninghamia 2008, 10, 547–555. [Google Scholar]
- Trezise, J.E.; Facelli, J.M.; Paton, D.C.; Davies, R.J.-P. The effect of heat and smoke on the soil seed banks of heathlands on permanent freshwater swamps. Austral Ecol. 2021, 46, 39–51. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, MA, USA, 2008. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Anderson, M.J.; Connell, S.D.; Gillanders, B.M.; Diebel, C.E.; Blom, W.M.; Saunders, J.E.; Landers, T.J. Relationships between taxonomic resolution and spatial scales of multivariate variation. J. Anim. Ecol. 2005, 74, 636–646. [Google Scholar] [CrossRef]
- Burrows, N.; Ward, B.; Wills, A.; Williams, M.; Cranfield, R. Fine-scale temporal turnover of jarrah forest understory vegetation assemblages is independent of fire regime. Fire Ecol. 2019, 15, 18. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth Marine Laboratory, UK, 2001. [Google Scholar]
- Keith, D.A. Functional traits: Thier roles in undertanding and predicting biotic responses to fire regimes from individuals to landscapes. In Flammable Australia: Fire Regimes, Biodiversity and Ecosystems in a Changing World, 2nd ed.; Bradstock, R.A., Gill, A.M., Williams, R.J., Eds.; CSIRO: Melbourne, VIC, Australia, 2012; pp. 97–125. [Google Scholar]
- Keith, D.A.; Holman, L.; Rodoreda, S.; Lemmon, J.; Bedward, M. Plant functional types can predict decade-scale changes in fire-prone vegetation. J. Ecol. 2007, 95, 1324–1337. [Google Scholar] [CrossRef]
- Anderson, B.J.; Chiarucci, A.; Williamson, M. How differences in plant abundance measures produce different species-abundance distributions. Methods Ecol. Evol. 2012, 3, 783–786. [Google Scholar] [CrossRef]
- Clarke, P.J.; Dorji, K. Are trade-offs in plant resprouting manifested in community seed banks. Ecology 2008, 89, 1850–1858. [Google Scholar] [CrossRef]
- Vázquez-Ramírez, J.; Venn, S.E. Snow, fire and drought: How alpine and treeline soil seed banks are affected by simulated climate change. Ann. Bot. 2023, mcad184. [Google Scholar] [CrossRef] [PubMed]
- Ashton, D.H. The Big Ash forest, Wallaby Creek, Victoria; changes during one lifetime. Aust. J. Bot. 2000, 48, 1–26. [Google Scholar] [CrossRef]
- Campbell, M.L.; Clarke, P.J. Response of montane wet sclerophyll forest understorey species to fire: Evidence from high and low intensity fires. Proc. Linn. Soc. New South Wales 2006, 127, 63. [Google Scholar]
- Williams, R.J. Gap Dynamics in Subalpine Heathland and Grassland Vegetation in South- Eastern Australia. J. Ecol. 1992, 80, 343–352. [Google Scholar] [CrossRef]
- Pausas, J.G.; Lamont, B.B. Fire-released seed dormancy—A global synthesis. Biol. Rev. 2022, 97, 1612–1639. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.-F.; Shi, S.-H.; Jiang, Y.-S.; Liu, J. A global synthesis of fire effects on soil seed banks. Glob. Ecol. Conserv. 2022, 36, e02132. [Google Scholar] [CrossRef]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A Compound from Smoke That Promotes Seed Germination. Science 2004, 305, 977. [Google Scholar] [CrossRef]
- Falster, D.; Gallagher, R.; Wenk, E.H.; Wright, I.J.; Indiarto, D.; Andrew, S.C.; Baxter, C.; Lawson, J.; Allen, S.; Fuchs, A.; et al. AusTraits, a curated plant trait database for the Australian flora. Sci. Data 2021, 8, 254. [Google Scholar] [CrossRef]
- Taranto, M.; Downe, J.; Coates, F.; Oates, A. Recovery of Montane Swamp Complex after Bushfires in North East Victoria 2003. Arthur Rylah Institute for Environmental Research Technical Report Series No. 152; Victorian Government Department of Sustainability and Environment: Melbourne, VIC, Australia, 2004; p. 44. [Google Scholar]
- Emery, N.J.; Offord, C.A. Managing Persoonia (Proteaceae) species in the landscape through a better understanding of their seed biology and ecology. Cunninghamia J. Plant Ecol. East. Aust. 2018, 18, 89–107. [Google Scholar]
- Walsh, N.G.; McDougall, K.L. Progress in the recovery of the flora of treeless subalpine vegetation in Kosciuszko National Park after the 2003 fires. Cunninghamia 2004, 8, 439–452. [Google Scholar]
- Bargmann, T.; Måren, I.E.; Vandvik, V.; Fraser, L. Life after fire: Smoke and ash as germination cues in ericads, herbs and graminoids of northern heathlands. Appl. Veg. Sci. 2014, 17, 670–679. [Google Scholar] [CrossRef]
- Keeley, J.E. Seed-germination patterns in fire-prone Mediterranean-climate regions. In Ecology and Biogeography of Mediterranean Ecosystems in Chile, California, and Australia; Arroyo, M., Zedler, P.H., Fox, M.D., Eds.; Springer: New York, NY, USA, 1995; pp. 239–273. [Google Scholar]
- Pausas, J.G.; Keeley, J.E. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Kasel, S.; Bennett, L.; Aponte, C.; Fedrigo, M.; Nitschke, C. Environmental heterogeneity promotes floristic turnover in temperate forests of south-eastern Australia more than dispersal limitation and disturbance. Landsc. Ecol. 2017, 32, 1613–1629. [Google Scholar] [CrossRef]
- Kiel, N.G.; Braziunas, K.H.; Turner, M.G. Peeking under the canopy: Anomalously short fire-return intervals alter subalpine forest understory plant communities. New Phytol 2023, 239, 1225–1238. [Google Scholar] [CrossRef] [PubMed]
- Chesson, P. Mechanisms of Maintenance of Species Diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.W.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Halpern, C.B.; Antos, J.A. Burn severity and pre-fire seral state interact to shape vegetation responses to fire in a young, western Cascade Range forest. For. Ecol. Manag. 2022, 507, 18. [Google Scholar] [CrossRef]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Wright, B.R.; Clarke, P.J. Fire, aridity and seed banks. What does seed bank composition reveal about community processes in fire-prone desert? J. Veg. Sci. 2009, 20, 663–674. [Google Scholar] [CrossRef]
- Bowd, E.J.; Blair, D.P.; Lindenmayer, D.B. Prior disturbance legacy effects on plant recovery post-high-severity wildfire. Ecosphere 2021, 12, e03480. [Google Scholar] [CrossRef]
- Blair, D.P.; McBurney, L.M.; Blanchard, W.; Banks, S.C.; Lindenmayer, D.B. Disturbance gradient shows logging affects plant functional groups more than fire. Ecol. Appl. 2016, 26, 2280–2301. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Cai, W.; Prentice, I.C.; Harrison, S.P. Community Abundance of Resprouting in Woody Plants Reflects Fire Return Time, Intensity, and Type. Forests 2023, 14, 878. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef] [PubMed]
- McDougall, K.L.; Walsh, N.G.; Wright, G.T. Recovery of treeless subalpine vegetation in Kosciuszko National Park after the landscape-scale fire of 2003. Aust. J. Bot. 2015, 63, 597–607. [Google Scholar] [CrossRef]
- Nolan, R.H.; Collins, L.; Leigh, A.; Ooi, M.K.J.; Curran, T.J.; Fairman, T.A.; de Dios, V.R.; Bradstock, R. Limits to post-fire vegetation recovery under climate change. Plant Cell Environ. 2021, 44, 3471–3489. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Baker, P.J.; Kasel, S.; Trouvé, R.; Stewart, S.B.; Nitschke, C.R. The role of climatic variability on Eucalyptus regeneration in southeastern Australia. Glob. Ecol. Conserv. 2021, 32, e01929. [Google Scholar] [CrossRef]
- Singh, A.; Kasel, S.; Hui, F.K.C.; Trouvé, R.; Baker, P.J.; Nitschke, C.R. Acacia Density, Edaphic, and Climatic Factors Shape Plant Assemblages in Regrowth Montane Forests in Southeastern Australia. Forests 2023, 14, 1166. [Google Scholar] [CrossRef]
- Barker, J.W.; Price, O.F.; Jenkins, M.E. High severity fire promotes a more flammable eucalypt forest structure. Austral Ecol. 2022, 47, 519–529. [Google Scholar] [CrossRef]
- Zylstra, P.J. Flammability dynamics in the Australian Alps. Austral Ecol. 2018, 43, 578–591. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Taylor, C. New spatial analyses of Australian wildfires highlight the need for new fire, resource, and conservation policies. Proc. Natl. Acad. Sci. USA 2020, 117, 12481–12485. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the fire history in Shrubby Dry Forest (top) and Sub-Alpine Woodland (bottom). Values in the grey bars represent the fire year, with values above the grey bars indicating the inter-fire interval. Adapted from [31].
Figure 1.
Schematic representation of the fire history in Shrubby Dry Forest (top) and Sub-Alpine Woodland (bottom). Values in the grey bars represent the fire year, with values above the grey bars indicating the inter-fire interval. Adapted from [31].
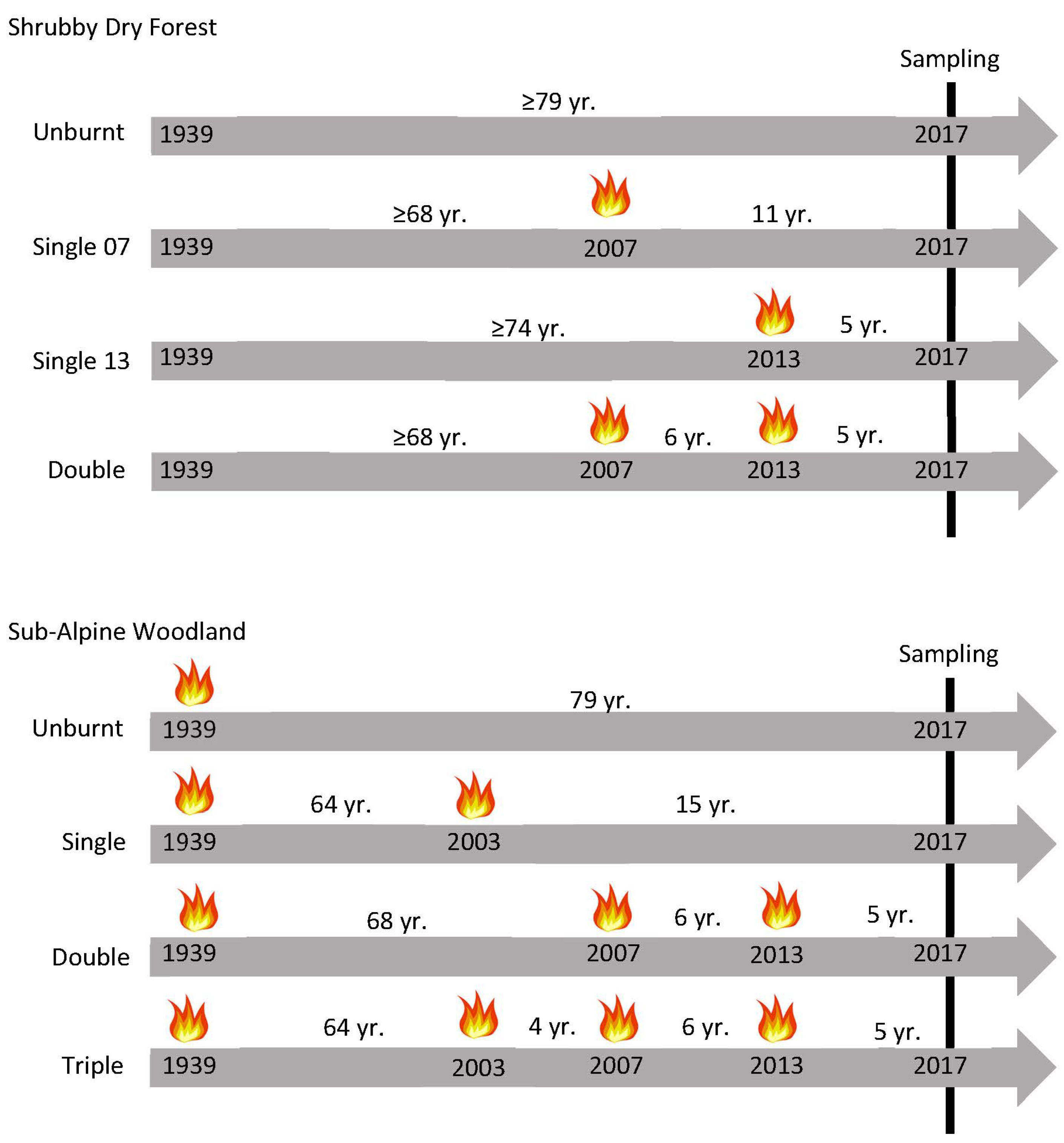
Figure 2.
Richness and density of soil seed bank germinants according to Treatment (C, control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke) and Fire Frequency for Shrubby Dry Forest (top) and Sub-Alpine Woodland (bottom). Bars represent means with one standard error. Significance of effects were determined via two-way PERMANOVA (on Euclidean distance) of Treatment (T), Fire Frequency (F), and their interaction (T × F). For each species pool (total, native, and introduced), significant T and FF effects are indicated by superscript letters (p ≤ 0.05). Significant interaction effects are provided in Figure 3 and statistical analyses in Table S4. (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001, and NS not significant).
Figure 2.
Richness and density of soil seed bank germinants according to Treatment (C, control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke) and Fire Frequency for Shrubby Dry Forest (top) and Sub-Alpine Woodland (bottom). Bars represent means with one standard error. Significance of effects were determined via two-way PERMANOVA (on Euclidean distance) of Treatment (T), Fire Frequency (F), and their interaction (T × F). For each species pool (total, native, and introduced), significant T and FF effects are indicated by superscript letters (p ≤ 0.05). Significant interaction effects are provided in Figure 3 and statistical analyses in Table S4. (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001, and NS not significant).
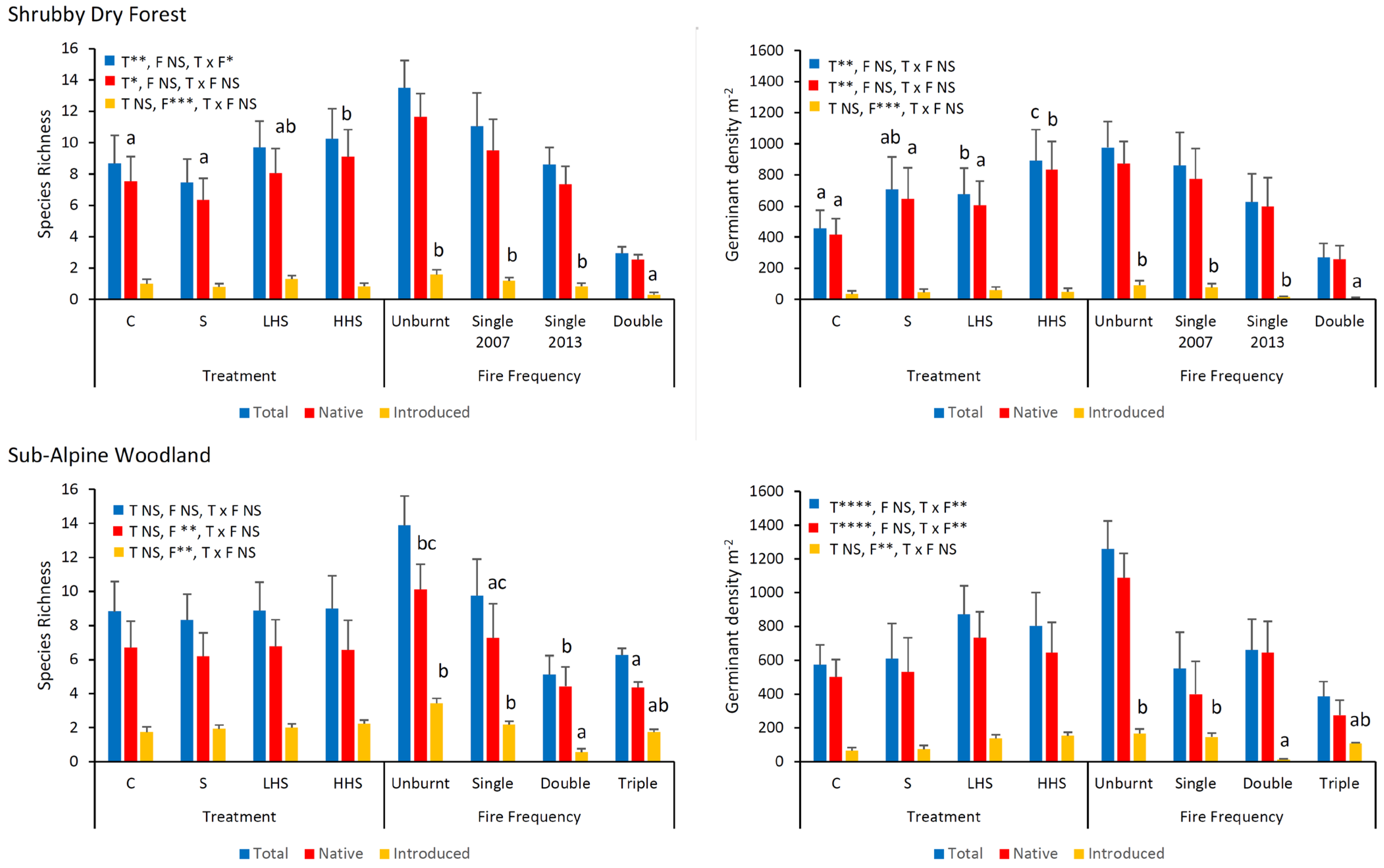
Figure 3.
Richness and density of the soil seed bank as a function of Treatment (C, control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke) within Fire Frequency. For Sub-Alpine Woodland, results for native species germinant density mirror those for total species (see Table S4). Bars represent means with one standard error. Significance of effects were determined via one-way PERMANOVA (on Euclidean distance) of Treatment (T) within Fire Frequency (F). Significant effects are indicated by superscript letters (p ≤ 0.05).
Figure 3.
Richness and density of the soil seed bank as a function of Treatment (C, control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke) within Fire Frequency. For Sub-Alpine Woodland, results for native species germinant density mirror those for total species (see Table S4). Bars represent means with one standard error. Significance of effects were determined via one-way PERMANOVA (on Euclidean distance) of Treatment (T) within Fire Frequency (F). Significant effects are indicated by superscript letters (p ≤ 0.05).
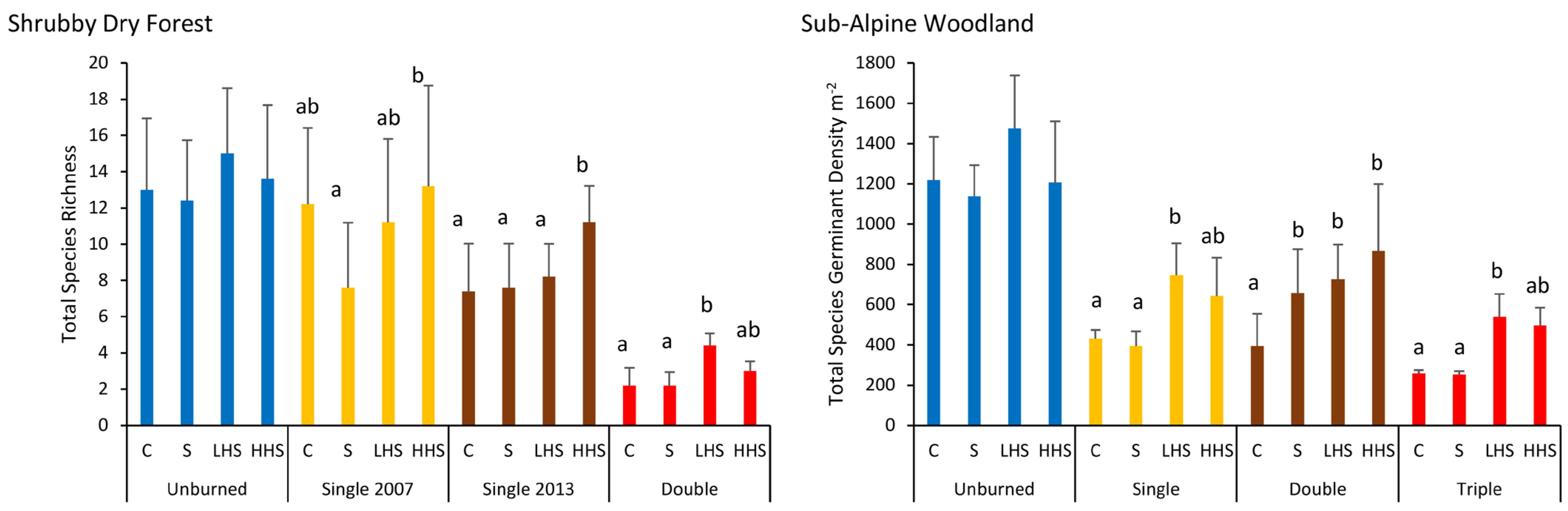
Figure 4.
Species richness within Vegetation Type and Fire Frequency for Shrubby Dry Forest (left) and Sub-Alpine Woodland (right). Bars represent means with one standard error. Significance of effects were determined via two-way PERMANOVA (on Euclidean distance) of Vegetation Type (V), Fire Frequency (F), and their interaction (V × F). For each species pool (total, native, and introduced) significant effects are indicated by superscript letters (p ≤ 0.05) and are across Fire Frequency (i.e., inclusive of extant vegetation and soil seed bank) and Vegetation Type (i.e., inclusive of all fire frequencies), as there were no significant interaction effects for either Shrubby Dry Forest or Sub-Alpine Woodland (Table S7). (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001, and NS not significant).
Figure 4.
Species richness within Vegetation Type and Fire Frequency for Shrubby Dry Forest (left) and Sub-Alpine Woodland (right). Bars represent means with one standard error. Significance of effects were determined via two-way PERMANOVA (on Euclidean distance) of Vegetation Type (V), Fire Frequency (F), and their interaction (V × F). For each species pool (total, native, and introduced) significant effects are indicated by superscript letters (p ≤ 0.05) and are across Fire Frequency (i.e., inclusive of extant vegetation and soil seed bank) and Vegetation Type (i.e., inclusive of all fire frequencies), as there were no significant interaction effects for either Shrubby Dry Forest or Sub-Alpine Woodland (Table S7). (* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001, and NS not significant).
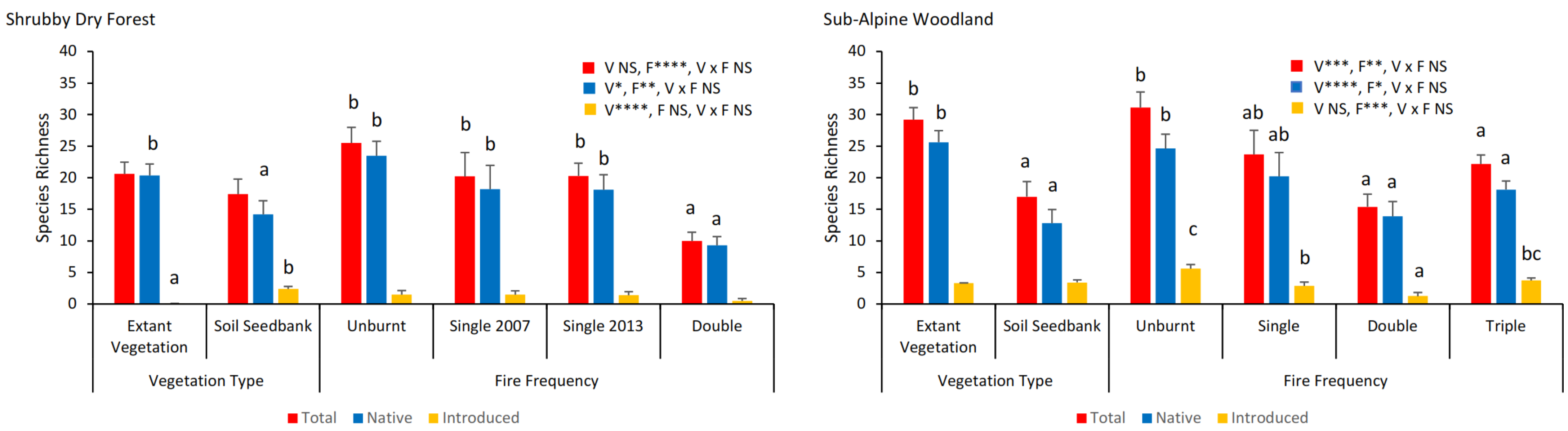
Figure 5.
NMDS ordination of species in the germinable soil seed bank (SSB) and extant vegetation (Veg): (a) Shrubby Dry Forest and (b) Sub-Alpine Woodland. Symbols represent individual sites. The vector overlay represents the strength and direction of the relationship for underlying species with r ≥ 0.65. The vector insert represents relationships with all underlying plant functional types (abbreviations follow Table 1). PERMANOVA tested for differences in Euclidean distance between Vegetation Type (V), Fire Frequency (F), and their interaction. Significant pairwise comparisons are indicated (V, vegetation type; F, fire frequency; UB, unburnt; S, single; D, double; T, triple; *** p ≤ 0.001, **** p ≤ 0.0001; and NS, not significant).
Figure 5.
NMDS ordination of species in the germinable soil seed bank (SSB) and extant vegetation (Veg): (a) Shrubby Dry Forest and (b) Sub-Alpine Woodland. Symbols represent individual sites. The vector overlay represents the strength and direction of the relationship for underlying species with r ≥ 0.65. The vector insert represents relationships with all underlying plant functional types (abbreviations follow Table 1). PERMANOVA tested for differences in Euclidean distance between Vegetation Type (V), Fire Frequency (F), and their interaction. Significant pairwise comparisons are indicated (V, vegetation type; F, fire frequency; UB, unburnt; S, single; D, double; T, triple; *** p ≤ 0.001, **** p ≤ 0.0001; and NS, not significant).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Significance of Seedbank Treatment and Fire Frequency effects on trait associations for Shrubby Dry Forest (based on counts and richness). For each trait, there was no significant (p > 0.05) Seedbank Treatment × Fire Frequency interaction. Values represent the mean proportion of species (based on density or richness) with each attribute according to Seedbank Treatment and Fire Frequency. Bold values indicate the two attributes that contributed most strongly to dissimilarity for each pairwise comparison (via SIMPER).
Table 1.
Significance of Seedbank Treatment and Fire Frequency effects on trait associations for Shrubby Dry Forest (based on counts and richness). For each trait, there was no significant (p > 0.05) Seedbank Treatment × Fire Frequency interaction. Values represent the mean proportion of species (based on density or richness) with each attribute according to Seedbank Treatment and Fire Frequency. Bold values indicate the two attributes that contributed most strongly to dissimilarity for each pairwise comparison (via SIMPER).
| Density | Richness | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait and Attribute | Soil Seedbank Treatment A | Fire Frequency B | Soil Seedbank Treatment | Fire Frequency | ||||||||||||
| C | S | LHS | HHS | UB | S07 | S13 | D | C | S | LHS | HHS | UB | S07 | S13 | D | |
| Life Form C | F(3, 64) = 1.72, p = 0.0379 | F(3,64) = 4.09, p = 0.0001 | F(3, 64) = 2.06, p = 0.0062 | F(3, 64) = 3.45, p = 0.0003 | ||||||||||||
| ab | a | ab | b | a | a | a | b | a | a | ab | b | a | a | a | b | |
| HE | 7.6 | 4 | 3.8 | 7.1 | 3.7 | 3.7 | 8.9 | 6.1 | 9.7 | 7 | 5 | 8.5 | 8.8 | 8 | 8 | 5.4 |
| T | 35.9 | 57.3 | 40.1 | 34.2 | 22.8 | 45.6 | 36.8 | 62.4 | 33.7 | 50 | 36.9 | 23.9 | 23.5 | 40 | 33.6 | 47.4 |
| C | 3.5 | 3.2 | 7.6 | 6.4 | 6.5 | 5.1 | 5.4 | 3.7 | 4 | 8.8 | 10.9 | 8.1 | 8.4 | 7.3 | 7.6 | 7.5 |
| HPr | 29.4 | 16.3 | 23.5 | 25.8 | 39.8 | 24 | 29.3 | 2 | 19.9 | 13.2 | 17.4 | 19.2 | 24.5 | 17.9 | 21.5 | 5.9 |
| HF | 3.1 | 1.3 | 1.8 | 1.1 | 4.3 | 1.9 | 0.9 | 0.2 | 6.1 | 3.1 | 1.6 | 1.5 | 2.8 | 5.6 | 2.6 | 1.2 |
| G | 3.1 | 4.9 | 1.5 | 1.3 | 0.8 | 0.4 | 3.8 | 5.8 | 4 | 5.8 | 1.8 | 2.7 | 2.5 | 1.3 | 7 | 3.7 |
| P | 10.4 | 11.6 | 21.1 | 20.9 | 18.7 | 19 | 11.5 | 14.8 | 14.9 | 10.9 | 24.9 | 31.6 | 23.5 | 18.7 | 16.2 | 23.8 |
| E | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| HP | 2 | 1.4 | 0.5 | 3.1 | 3.3 | 0.3 | 3.5 | 0 | 2.6 | 2.2 | 1.6 | 4.4 | 5.9 | 1.2 | 3.6 | 0 |
| Dispersal Mode D | F(3, 64) = 2.94, p = 0.0010 | F(3, 64) = 5.51, p = 0.0001 | F(3, 64) = 3.01, p = 0.0002 | F(3, 64) = 5.29, p = 0.0001 | ||||||||||||
| a | a | b | b | a | b | a | b | a | a | b | b | a | b | c | d | |
| bar | 11.8 | 9.7 | 22.7 | 23.4 | 27.5 | 14.4 | 16.2 | 9.5 | 12.4 | 18.8 | 20.1 | 23.4 | 25.1 | 17.3 | 17.9 | 14.3 |
| ane | 34.8 | 56.1 | 38.9 | 38.3 | 20.2 | 51.3 | 37.5 | 59 | 38.3 | 43.7 | 39.6 | 26.5 | 23.9 | 43.8 | 34.1 | 46.4 |
| myr | 4.5 | 1.9 | 16 | 14.7 | 8.9 | 9.9 | 5.6 | 12.6 | 7.7 | 4.7 | 18 | 22 | 13.5 | 12.2 | 7.1 | 19.7 |
| end | 1.8 | 1.8 | 2.2 | 1 | 2 | 2.6 | 1.5 | 0.6 | 2.9 | 1.8 | 3.9 | 3.3 | 3.6 | 4.7 | 1.6 | 2.1 |
| mob | 38.7 | 30.2 | 19.7 | 21.8 | 40.1 | 21.1 | 37.8 | 11.4 | 28.4 | 30.1 | 17.3 | 21.9 | 31.5 | 20.1 | 36.3 | 10 |
| epi | 3.4 | 0.3 | 0.5 | 0.9 | 1.3 | 0.6 | 1.3 | 1.8 | 5.2 | 1 | 1 | 2.8 | 2.5 | 2 | 3.1 | 2.5 |
| Fire Response E | F(3, 64) = 1.35, p = 0.1861 | F(3, 64) = 3.71, p = 0.0002 | F(3, 64) = 2.02, p = 0.0198 | F(3, 64) = 3.78, p = 0.0001 | ||||||||||||
| a | a | a | b | a | a | a | b | a | a | a | b | |||||
| R | 14.5 | 12.1 | 22 | 27.9 | 21.8 | 14.6 | 18.3 | 21.8 | 20.9 | 17.3 | 20.2 | 34.1 | 27.2 | 18.3 | 20.7 | 26.3 |
| S | 50.7 | 67.4 | 50.3 | 46.4 | 40.5 | 57.4 | 49.7 | 67.2 | 49.8 | 62.4 | 53.3 | 37.9 | 44.5 | 51.1 | 50.3 | 57.4 |
| Rs | 8.2 | 4.4 | 5.5 | 8.5 | 6.4 | 8.8 | 10.8 | 0.6 | 9 | 8.8 | 6.4 | 6.9 | 7.7 | 7.7 | 13.8 | 2 |
| SR | 2.4 | 2.5 | 4.3 | 3.8 | 1.9 | 5.3 | 3.7 | 2.2 | 3.2 | 3.6 | 4.7 | 9.1 | 3.8 | 7.6 | 5.6 | 3.5 |
| Sr | 19.2 | 13.6 | 17.8 | 13.3 | 29.4 | 13.8 | 17.5 | 3.2 | 12 | 7.9 | 15.4 | 12 | 16.8 | 15.3 | 9.5 | 5.7 |
PERMANOVA tested differences in Euclidean distance between Soil Seedbank Treatment, Fire Frequency, and their interaction. Significant pairwise comparisons are indicated by letters. A Soil Seedbank Treatment (C, Control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke). B Fire Frequency (UB, Unburnt; S07, Single2007; S13, Single 2013; and D, Double). C C, chamaephyte; G, geophyte; HE, erect rosette; HF, flat or versatile rosette; HP, partial rosette; HPr, proto-hemicryptophyte; P, phanerophyte; and T, therophyte. D ane, anemochory; bar, barochory; end, endozoochory; epi, epizoochory; mob, mobile; myr, myrmechcory. E R, obligate resprouters; S, obligate seeders; SR, facultative resprouters; Sr, weak seeders; and Rs, weak resprouters. B,C,D Trait definitions are provided in Table S2.
Table 2.
Significance of Seedbank Treatment and Fire Frequency effects on trait associations for Sub-Alpine Woodland (based on counts and richness). For each trait, there was no significant (p > 0.05) Seedbank Treatment × Fire Frequency interaction. Values represent the mean proportion (based on density or richness) of species with each attribute according to Seedbank Treatment and Fire Frequency. Bold values indicate the attributes that contributed most strongly to dissimilarity for each pairwise comparison (via SIMPER).
Table 2.
Significance of Seedbank Treatment and Fire Frequency effects on trait associations for Sub-Alpine Woodland (based on counts and richness). For each trait, there was no significant (p > 0.05) Seedbank Treatment × Fire Frequency interaction. Values represent the mean proportion (based on density or richness) of species with each attribute according to Seedbank Treatment and Fire Frequency. Bold values indicate the attributes that contributed most strongly to dissimilarity for each pairwise comparison (via SIMPER).
| Density | Richness | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait and Attribute C | Soil Seedbank Treatment A | Fire Frequency B | Soil Seedbank Treatment | Fire Frequency | ||||||||||||
| C | S | LHS | HHS | UB | S | D | T | C | S | LHS | HHS | UB | S | D | T | |
| Life Form | F(3, 48) = 0.39, p = 0.9883 | F(3, 48) = 4.06, p = 0.0001 | F(3, 48) = 0.67, p = 0.8468 | F(3, 48) = 2.89, p = 0.0008 | ||||||||||||
| a | a | b | a | a | a | b | ab | |||||||||
| HE | 61.9 | 59.7 | 60.3 | 58 | 66 | 49.9 | 86 | 37.9 | 44.7 | 41.6 | 38.2 | 40.2 | 36 | 35.5 | 56.5 | 36.7 |
| T | 3 | 4.6 | 2 | 3.4 | 2.9 | 3.3 | 1.8 | 4.9 | 8.5 | 11.9 | 7.1 | 9.9 | 9 | 9.5 | 7.4 | 11.5 |
| C | 0.9 | 0.6 | 0.8 | 0.4 | 1.3 | 0.3 | 0.7 | 0.4 | 3.4 | 1.4 | 1.1 | 1.2 | 3.4 | 0.8 | 1.4 | 1.6 |
| HPr | 14.2 | 13.8 | 10.5 | 11 | 10 | 17.4 | 7.4 | 14.7 | 17.7 | 20.6 | 18.8 | 20.6 | 21.3 | 21.7 | 19.4 | 15.4 |
| HF | 6.8 | 6.3 | 6.7 | 7.5 | 10.5 | 9.9 | 1.6 | 5.2 | 8.5 | 8.8 | 9.6 | 9.5 | 14.2 | 10.6 | 4.5 | 7.2 |
| G | 4.6 | 5.7 | 4.6 | 4.7 | 0.1 | 0.7 | 0.3 | 18.5 | 2.9 | 2.6 | 2.7 | 2.1 | 0.6 | 1.2 | 1.6 | 7 |
| P | 0.5 | 1.6 | 3.4 | 2.1 | 0.8 | 5.3 | 1.1 | 0.3 | 1.5 | 3.1 | 8.9 | 5.3 | 4.6 | 9.4 | 4.2 | 0.6 |
| E | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| HP | 8.1 | 7.8 | 11.7 | 12.9 | 8.4 | 13.1 | 1 | 18 | 12.8 | 9.9 | 13.5 | 11.1 | 10.8 | 11.4 | 5.1 | 20.1 |
| Dispersal Mode | F(3, 48) = 0.52, p = 0.8875 | F(3, 48) = 4.03, p = 0.0001 | F(3, 48) = 0.31, p = 0.8998 | F(3, 48) = 4.19, p = 0.0008 | ||||||||||||
| a | ac | b | c | a | a | b | ab | |||||||||
| Bar | 76.3 | 73.6 | 76.4 | 76.6 | 74.4 | 73.3 | 90.5 | 64.6 | 63.8 | 59.3 | 62.1 | 65.6 | 60 | 54.7 | 74.6 | 61.5 |
| Ane | 11 | 11.8 | 7.4 | 8.2 | 6 | 16.5 | 3.5 | 12.4 | 20.9 | 23 | 17.1 | 15.9 | 15.6 | 25.3 | 13.3 | 22.6 |
| Myr | 4.6 | 5.9 | 8.2 | 8.1 | 12.8 | 6.8 | 5.2 | 2 | 6.3 | 8.6 | 10.3 | 10.2 | 12 | 10.6 | 8.9 | 3.9 |
| End | 0 | 0.1 | 0.5 | 0.3 | 0.6 | 0 | 0 | 0.4 | 0 | 0.5 | 1.4 | 0.3 | 1.5 | 0 | 0 | 0.7 |
| Mob | 7.8 | 8 | 7.2 | 6.3 | 5.3 | 3.2 | 0.7 | 20.1 | 7.9 | 7.2 | 7.7 | 6.1 | 7.1 | 9 | 2.6 | 10.1 |
| Epi | 0.3 | 0.6 | 0.3 | 0.5 | 0.9 | 0.2 | 0.1 | 0.5 | 1.1 | 1.5 | 1.5 | 1.9 | 3.8 | 0.4 | 0.6 | 1.3 |
| Fire Response | F(3, 48) = 0.56, p = 0.8481 | F(3, 48) = 4.76, p = 0.0002 | F(3, 48) = 0.40, p = 0.9636 | F(3, 48) = 3.53, p = 0.0009 | ||||||||||||
| a | b | b | a | a | abc | b | c | |||||||||
| R | 66.5 | 65.3 | 65.2 | 60.7 | 62 | 52.2 | 75.2 | 68.2 | 54 | 50.6 | 47.3 | 48.8 | 40.3 | 43.4 | 64.9 | 52.1 |
| S | 16.4 | 17.1 | 14.1 | 13.1 | 8.9 | 22.9 | 20.6 | 8.4 | 20.7 | 22.5 | 26.1 | 21.4 | 23.3 | 28.1 | 20.5 | 18.7 |
| Rs | 7.2 | 6.2 | 4.1 | 4.9 | 6.8 | 8.8 | 1.4 | 5.3 | 7.9 | 11.4 | 7.8 | 7.4 | 10.5 | 11.1 | 4.8 | 8.2 |
| SR | 6.9 | 7.8 | 11.7 | 16.5 | 11.8 | 14.1 | 1.3 | 15.6 | 13.1 | 11.3 | 14.2 | 15 | 18.5 | 11.6 | 6.3 | 17.2 |
| Sr | 3.1 | 3.6 | 4.9 | 4.8 | 10.5 | 2 | 1.5 | 2.5 | 4.2 | 4.2 | 4.6 | 7.4 | 7.4 | 5.7 | 3.5 | 3.7 |
PERMANOVA tested differences in Euclidean distance between Soil Seedbank Treatment, Fire Frequency, and their interaction. Significant pairwise comparisons are indicated by letters. A Soil Seedbank Treatment (C, Control; S, smoke; LHS, low heat plus smoke; and HHS, high heat plus smoke). B Fire Frequency (UB, Unburnt; S, Single; D, Double; and T, Triple). C Abbreviations follow Table 1, with trait definitions provided in Table S2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kasel, S.; Fairman, T.A.; Nitschke, C.R. Short-Interval, High-Severity Wildfire Depletes Diversity of Both Extant Vegetation and Soil Seed Banks in Fire-Tolerant Eucalypt Forests. Fire 2024, 7, 148. https://doi.org/10.3390/fire7040148
AMA Style
Kasel S, Fairman TA, Nitschke CR. Short-Interval, High-Severity Wildfire Depletes Diversity of Both Extant Vegetation and Soil Seed Banks in Fire-Tolerant Eucalypt Forests. Fire. 2024; 7(4):148. https://doi.org/10.3390/fire7040148
Chicago/Turabian StyleKasel, Sabine, Thomas A. Fairman, and Craig R. Nitschke. 2024. "Short-Interval, High-Severity Wildfire Depletes Diversity of Both Extant Vegetation and Soil Seed Banks in Fire-Tolerant Eucalypt Forests" Fire 7, no. 4: 148. https://doi.org/10.3390/fire7040148