

Entropy and Cross-Level Orderliness in Light of the Interconnection between the Neural System and Consciousness


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods of Studying Consciousness in Contemporary Science
2.1. Global Connectivity, Integration of a System, and Cognitive Abilities
2.2. Time, Rhythms, and Sustainability of a System
2.3. The Temporospatial Framework for Studying Neural Systems
3. Results of Considering Subjective Reality within the Evolutionary Framework
3.1. Sustainability of Regular Changes and Linear Evolution of a System
3.2. Increasing Complexity of the System from the Organism and Up to the Society
3.3. Human Society and Transformation of the Reality
4. Discussion of the Research Limitations and New Vistas
- This research was focused on philosophical issues and aimed to present them as a review rather than introducing the same ideas via experiments, modeling, or calculations. Due to the vagueness of the theme and the existence of many theories of consciousness and methods to count entropy, any attempt to consider raised issues more particularly would have been a voluntary choice of some mathematical or research model caused by the author’s preferences and knowledge, with a pretention for some objective inquiry. An effective method to deal with the entropy of the system’s physical and subjective states [22,24] is a challenge that must be resolved by the work of the scientific community rather than by individuals. I suppose that the method and formula must be as simple as possible and connected with the known principles of physical reality. Furthermore, it may be reasonable to consider the entropy of a system in its interconnection with the regularities of the world rather than a closed entity. More profound elaboration of this issue should be based on the philosophical background presented in [33].
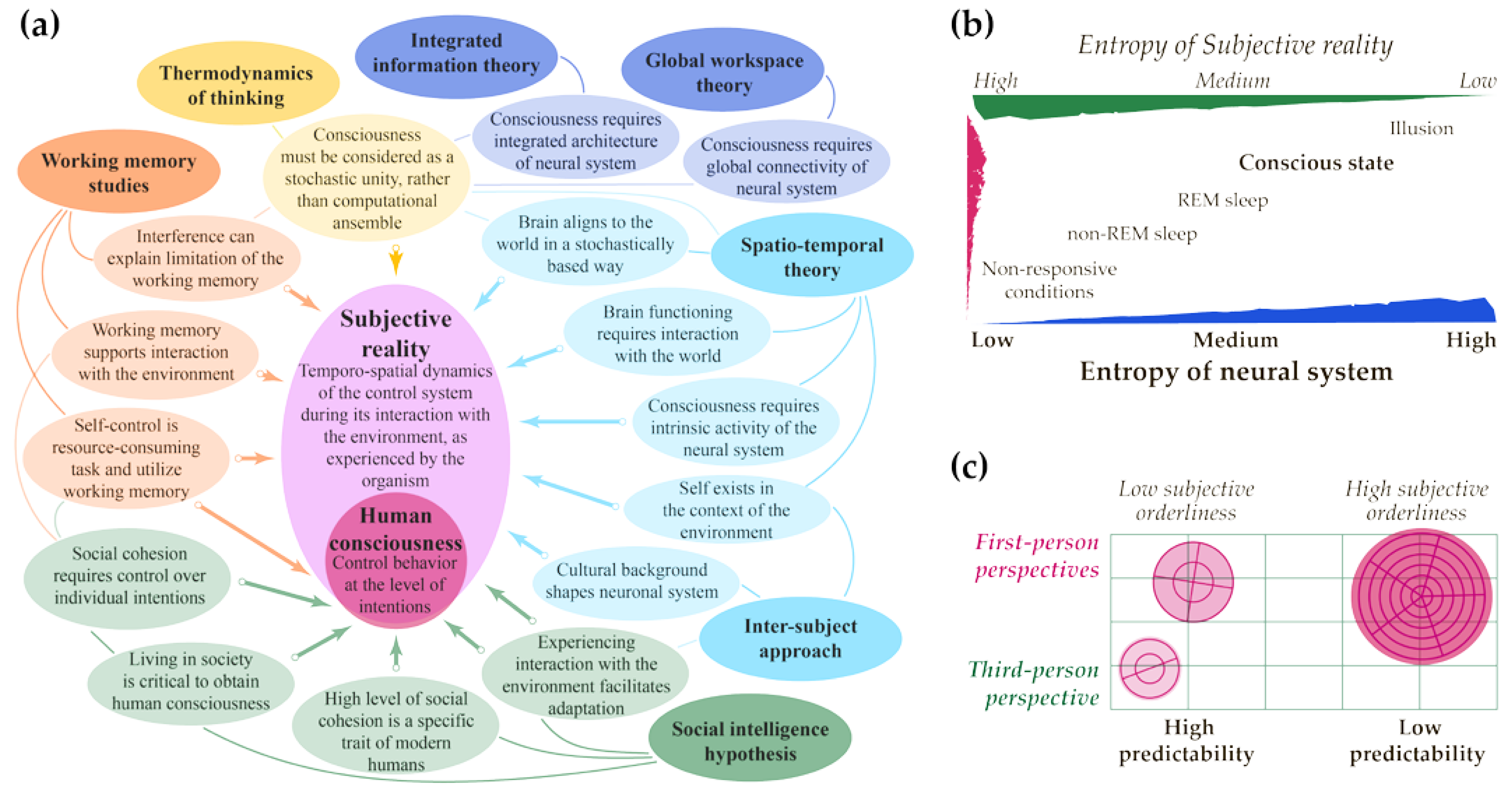
- Considering entropies of the neural system and subjective reality allows us to find new methods of estimating the content of consciousness from the third-person perspective (see Figure 1 above). However, this requires profound empirical research using neuroimaging and other methods to obtain reliable data. To minimize possible bias in the evaluation of subjective reality’s functioning, it is critical to pay great attention to cross-cultural and other diversities, which are relatively poorly studied yet [151,152,153]. As discussed above, the top level of the human orderliness is consciousness. It must be mastered during an individual‘s life by including society and adopting its regularities, which are slower than simple behavioral functioning [84,88]. Thus, particular differences in the neural system across the human population may be best seen in the context of temporal functioning rather than the spatial organization of the brain connectome, see [31]. Hence, the temporospatial theory of consciousness [26,27,28,29,30,31,32,33] seems to be the most appropriate framework for future research on human consciousness, freed from the unintentional bias grounded in considering a part of the population as an etalon.
- Despite attempts to highlight the critical role of time, its understanding is a great challenge that comes from one of the most foundational qualities of subjective reality: its exclusiveness [66]. Complicating the current state and quantification of time is a natural tendency for the extensive development of the current level of orderliness. At the same time, any attempt to comprehend regularities of the higher order requires dealing with slower frequencies, which are difficult to reach by means of the current tools of perception and action. The development of any system, from organism to culture and knowledge, follows the same process as natural selection with the available regularities of the environment [132]. In turn, the latter requires that the existing system be sufficiently rich in meeting them and warrants sustainable development for future development. Thus, the concept of temporal nestedness suggested in temporospatial theory may have a much wider application that describes the functioning of the neural system [26,27,28,29,30,31,32,41,42].
- Last but not least, considering the creative abilities of humans to modify their living world raises ethical and philosophical questions [3,4]. Any complex system is a product of aligning with the world; however, it is also affected by its functioning. From a physical perspective, preserving a system’s orderliness facilitates its environment’s entropy. However, the basic needs of the organism are the product of the natural selection of the species and, thus, tend to preserve the balance with reality. As the product of social selection, human consciousness reflects more rapid changes and can introduce meanings above natural needs, which tend to modify the environment much more dramatically [90]. Thus, the ability for intentional activity in a cohesive collective allows humans to control entropy not only within the group but also outside it. Recent social disturbances have highlighted these dangers [154,155]. The outcome of our actions depends on the adopted intentions and goals, which may be both within the system and the world. Significantly, the borders between them are determined by one’s consciousness: it can be an individual body, family, society, or the whole world perceived as a unity. Determining which goal will be chosen is a matter of current choice.
5. Conclusions
- In a regular environment, any occasional assemblage of things is subject to natural selection for coherence with the world by obtaining surrounding orderliness. Thus, the latter’s effect allows the maintenance of the entropy of the former below a particular threshold and causes the emergence of a system that can preserve homeostasis, anticipate upcoming events, and reproduce under suitable circumstances. This demonstrates the scale-free nature of the world’s orderliness.
- Interaction with the world and natural behavior involves the entire system, requiring all its components to be united in a cohesive motion that constitutes a specific subjective state. Therefore, each anticipative system can be considered to have a particular subjective reality with its cycles of activity and level of entropy that is coherent with its living world.
- The evolution of systems involves facing new regularities and starting a new round of adaptation, either introducing competitive subsystems of the same level or mounting a higher level of orderliness. The former complicates the behavior and increases the entropy of the system. The latter is linked to obtaining new abilities, which usually invoke changes in the organism’s structure. This decreases the entropy of the system and can even lead to the emergence of society.
- The rhythms of the higher level must outperform and guide the activities of the subsystems. This assumes that the higher level constantly performs its intrinsic activity, in which frequencies are slower but involve more units than any sub-system. The human neural system demonstrates all of these qualities and grants exclusive access to the contents of subjective reality. The extraordinary evolutionary history of the human neural system allows one to master the ability of conscious action. This is an achievement that must be gained during the life history of an individual, and the duty of society is to provide opportunities for this.
- The framework of the temporospatial theory allows us to understand human consciousness in the context of natural world evolution. This demonstrates that alignment with the living world is necessary for achieving coherence between the environment’s regularity and the system’s entropy. However, the emergence of human consciousness may be a new coil that allows for intentional change in entropy within the system as well as in the environment. This increases the responsibility of the species to master conscious action during their lives.
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Schneider, S.; Velmans, M. (Eds.) The Blackwell Companion to Consciousness, 2nd ed.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2017. [Google Scholar]
- Dehaene, S.; Lau, H.; Kouider, S. What is consciousness, and could machines have it? Science 2017, 358, 486–492. [Google Scholar] [CrossRef] [Green Version]
- Yuste, R.; Goering, S.; Arcas, B.A.Y.; Bi, G.; Carmena, J.M.; Carter, A.; Fins, J.J.; Friesen, P.; Gallant, J.; Huggins, J.E.; et al. Four ethical priorities for neurotechnologies and AI. Nature 2017, 551, 159–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reardon, S. Can lab-grown brains become conscious? Nature 2020, 586, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Chalmers, D.J. Is the Hard Problem of Consciousness Universal? J. Conscious. Stud. 2020, 27, 227–257. [Google Scholar]
- Dennett, D.C. Facing up to the hard question of consciousness. Philos. Trans. R. Soc. B 2018, 373, 20170342. [Google Scholar] [CrossRef] [Green Version]
- Boly, M.; Seth, A.K.; Wilke, M.; Ingmundson, P.; Baars, B.; Laureys, S.; Edelman, D.B.; Tsuchiya, N. Consciousness in humans and non-human animals: Recent advances and future directions. Front. Psychol. 2013, 4, 625. [Google Scholar] [CrossRef] [Green Version]
- Gutfreund, Y. The Mind-Evolution Problem: The Difficulty of Fitting Consciousness in an Evolutionary Framework. Front. Psychol. 2018, 9, 1537. [Google Scholar] [CrossRef] [PubMed]
- Rudrauf, D.; Sergeant-Perthuis, G.; Belli, O.; Tisserand, Y.; Serugendo, G.D.M. Modeling the subjective perspective of consciousness and its role in the control of behaviours. J. Theoret. Biol. 2022, 534, 110957. [Google Scholar] [CrossRef] [PubMed]
- Krakauer, J.W.; Ghazanfar, A.A.; Gomez-Marin, A.; MacIver, M.A.; Poeppel, D. Neuroscience Needs Behavior: Correcting a Reductionist Bias. Neuron 2017, 93, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Pessoa, L.; Medina, L.; Desfilis, E. Refocusing neuroscience: Moving away from mental categories and towards complex behaviours. Philos. Trans. R. Soc. B 2022, 377, 20200534. [Google Scholar] [CrossRef] [PubMed]
- Sidorova, A.; Levashova, N.; Garaeva, A.; Tverdislov, V. A percolation model of natural selection. Biosystems 2020, 193–194, 104120. [Google Scholar] [CrossRef]
- Papale, F.; Saget, J.; Bapteste, E. Networks Consolidate the Core Concepts of Evolution by Natural Selection. Trends Microbiol. 2020, 28, 254–265. [Google Scholar] [CrossRef] [Green Version]
- Cisek, P.; Hayden, B.Y. Neuroscience needs evolution. Philos. Trans. R. Soc. B 2022, 377, 20200518. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, F. Does the Explanatory Gap Rest on a Fallacy? Rev. Philos. Psychol. 2019, 10, 649–667. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Shultz, S. Evolution in the social brain. Science 2007, 317, 1344–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gopnik, A.; Frankenhuis, W.E.; Tomasello, M. Introduction to special issue: ‘Life history and learning: How childhood, caregiving and old age shape cognition and culture in humans and other animals’. Philos. Trans. R. Soc. B 2020, 375, 20190489. [Google Scholar] [CrossRef] [PubMed]
- Pettitt, P. The Palaeolithic Origins Of Human Burial; Routledge: Milton Park, UK, 2011. [Google Scholar]
- Searle, J. Theory of mind and Darwin‘s legacy. Proc. Natl. Acad. Sci. USA 2013, 110, 10343–10348. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.J.; Wiese, W.; Hobson, J.A. Sentience and the Origins of Consciousness: From Cartesian Duality to Markovian Monism. Entropy 2020, 22, 516. [Google Scholar] [CrossRef]
- Mason, J.W.D. From Learning to Consciousness: An Example Using Expected Float Entropy Minimisation. Entropy 2019, 21, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buxton, R.B. The thermodynamics of thinking: Connections between neural activity, energy metabolism and blood flow. Philos. Trans. R. Soc. B 2021, 376, 20190624. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef]
- Carhart-Harris, R.L. The entropic brain—Revisited. Neuropharmacology 2018, 142, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.V.; Gutiérrez, A.M.L. (Eds.) Temporal Points of View; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Northoff, G.; Wainio-Theberge, S.; Evers, K. Is temporo-spatial dynamics the "common currency" of brain and mind? In Quest of “Spatiotemporal Neuroscience”. Phys. Life Rev. 2020, 33, 34–54. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Tumati, S. “Average is good, extremes are bad”—Non-linear inverted U-shaped relationship between neural mechanisms and functionality of mental features. Neurosci. Biobehav. Rev. 2019, 104, 11–25. [Google Scholar] [CrossRef]
- Northoff, G. The Spontaneous Brain: From the Mind–Body to the World–Brain Problem; The MIT Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Northoff, G.; Huang, Z.R. How do the brain‘s time and space mediate consciousness and its different dimensions? Temporo-spatial theory of consciousness (TTC). Neurosci. Biobehav. Rev. 2017, 80, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G. Is the self a higher-order or fundamental function of the brain? The “basis model of self-specificity” and its encoding by the brain‘s spontaneous activity. Cogn. Neurosci. 2016, 7, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Zilio, F. From Shorter to Longer Timescales: Converging Integrated Information Theory (IIT) with the Temporo-Spatial Theory of Consciousness (TTC). Entropy 2022, 24, 270. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Zilio, F. Temporo-spatial Theory of Consciousness (TTC)—Bridging the gap of neuronal activity and phenomenal states. Behav. Brain Res. 2022, 424, 113788. [Google Scholar] [CrossRef] [PubMed]
- Northoff, G.; Smith, D. The subjectivity of self and its ontology: From the world-brain relation to the point of view in the world. Theory Psychol. 2022, 8, 120. [Google Scholar] [CrossRef]
- Barack, D.L.; Krakauer, J.W. Two views on the cognitive brain. Nat. Rev. Neurosci. 2021, 6, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.K.; Bayne, T. Theories of consciousness. Nat. Rev. Neurosci. 2022, 23, 439–452. [Google Scholar] [CrossRef]
- Gallagher, S. The Extended Mind: State of the Question. South. J. Philos. 2018, 56, 421–447. [Google Scholar] [CrossRef]
- Hameroff, S. ‘Orch OR’ is the most complete, and most easily falsifiable theory of consciousness. Cogn. Neurosci.-UK 2021, 12, 74–76. [Google Scholar] [CrossRef]
- Rosenthal, D. Competing models of consciousness. Cogn. Neuropsychol. 2020, 37, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Dubrovsky, D.I. “The Hard Problem of Consciousness”. Theoretical solution of its main questions. Aims Neurosci. 2019, 6, 85–103. [Google Scholar] [CrossRef]
- Koch, C. What is consciousness? Nature 2018, 557, S8–S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.F.H.; Kallio-Tamminen, T. Brain-mind operational architectonics: At the boundary between quantum physics and Eastern metaphysics. Phys. Life Rev. 2019, 31, 122–133. [Google Scholar] [CrossRef]
- Fingelkurts, A.A.; Fingelkurts, A.A.; Neves, C.E.H. Natural world physical, brain operational, and mind phenomenal space-time. Phys. Life Rev. 2010, 7, 195–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, H.C.; Changeux, J.P.; Rosenstand, A. Towards a cognitive neuroscience of self-awareness. Neurosci. Biobehav. Rev. 2017, 83, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Changeux, J.P. Climbing Brain Levels of Organisation from Genes to Consciousness. Trends Cogn. Sci. 2017, 21, 168–181. [Google Scholar] [CrossRef] [Green Version]
- Changeux, J.P. Wider than the sky: The phenomenal gift of consciousness. Nature 2004, 428, 603–604. [Google Scholar] [CrossRef] [Green Version]
- Dehaene, S. Consciousness and the Brain: Deciphering How the Brain Codes Our Thoughts; Viking: New York, NY, USA, 2014. [Google Scholar]
- Changeux, J.P.; Goulas, A.; Hilgetag, C.C. A Connectomic Hypothesis for the Hominization of the Brain. Cereb. Cortex 2021, 31, 2425–2449. [Google Scholar] [CrossRef]
- Machado, T.A.; Kauvar, I.V.; Deisseroth, K. Multiregion neuronal activity: The forest and the trees. Nat. Rev. Neurosci. 2022, 23, 683–704. [Google Scholar] [CrossRef] [PubMed]
- Gibson, J.J. The Ecological Approach to Visual Perception; Houghton Mifflin: Boston, MA, USA, 1979. [Google Scholar]
- Koban, L.; Gianaros, P.J.; Kober, H.; Wager, T.D. The self in context: Brain systems linking mental and physical health. Nat. Rev. Neurosci. 2021, 22, 309–322. [Google Scholar] [CrossRef]
- Baars, B.J.; Franklin, S. How conscious experience and working memory interact. Trends Cogn. Sci. 2003, 7, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Hahn, L.A.; Rose, J. Working Memory as an Indicator for Comparative Cognition—Detecting Qualitative and Quantitative Differences. Front. Psychol. 2020, 11, 01954. [Google Scholar] [CrossRef]
- Coolidge, F.L.; Wynn, T. The evolution of working memory. Annee Psychol. 2020, 120, 103–134. [Google Scholar] [CrossRef]
- Ma, W.J.; Husain, M.; Bays, P.M. Changing concepts of working memory. Nat. Neurosci. 2014, 17, 347–356. [Google Scholar] [CrossRef]
- Christophel, T.B.; Klink, P.C.; Spitzer, B.; Roelfsema, P.R.; Haynes, J.D. The distributed nature of working memory. Trends Cogn. Sci. 2017, 21, 111–124. [Google Scholar] [CrossRef]
- Barbosa, J.; Stein, H.; Martinez, R.L.; Galan-Gadea, A.; Li, S.H.; Dalmau, J.; Adam, K.C.S.; Valls-Sole, J.; Constantinidis, C.; Compte, A. Interplay between persistent activity and activity-silent dynamics in the prefrontal cortex underlies serial biases in working memory. Nat. Neurosci. 2020, 23, 1016–1024. [Google Scholar] [CrossRef] [PubMed]
- Finn, E.S.; Huber, L.; Jangraw, D.C.; Molfese, P.J.; Bandettini, P.A. Layer-dependent activity in human prefrontal cortex during working memory. Nat. Neurosci. 2019, 22, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Quentin, R.; King, J.R.; Sallard, E.; Fishman, N.; Thompson, R.; Buch, E.R.; Cohen, L.G. Differential Brain Mechanisms of Selection and Maintenance of Information during Working Memory. J. Neurosci. 2019, 39, 3728–3740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Dai, C.; Ba, Y.; Zhang, L.; Shao, Y.; Tian, J. Effect of Sleep Deprivation on the Working Memory-Related N2-P3 Components of the Event-Related Potential Waveform. Front. Neurosci. 2020, 14, 360. [Google Scholar] [CrossRef]
- Groß, D.; Kohlmann, C.W. Predicting self-control capacity—Taking into account working memory capacity, motivation, and heart rate variability. Acta Psychol. 2020, 209, 103131. [Google Scholar] [CrossRef] [PubMed]
- Sauseng, P.; Liesefeld, H.R. Cognitive Control: Brain Oscillations Coordinate Human Working Memory. Curr. Biol. 2020, 30, R405–R407. [Google Scholar] [CrossRef]
- Carruthers, P. Evolution of working memory. Proc. Natl. Acad. Sci. USA 2013, 110, 10371–10378. [Google Scholar] [CrossRef] [Green Version]
- Balter, M. Evolution of behavior. Did working memory spark creative culture? Science 2010, 328, 160–163. [Google Scholar] [CrossRef]
- Bouchacourt, F.; Buschman, T.J. A flexible model of working memory. Neuron 2019, 103, 147–160. [Google Scholar] [CrossRef]
- Tononi, G.; Boly, M.; Massimini, M.; Koch, C. Integrated information theory: From consciousness to its physical substrate. Nat. Rev. Neurosci. 2016, 17, 450–461. [Google Scholar] [CrossRef]
- Tononi, G.; Koch, C. Consciousness: Here, there and everywhere? Philos. Trans. R. Soc. B 2015, 370, 20140167. [Google Scholar] [CrossRef] [Green Version]
- Tononi, G. Consciousness as integrated information: A provisional manifesto. Biol. Bull. 2008, 215, 216–242. [Google Scholar] [CrossRef]
- Mallatt, J. A Traditional Scientific Perspective on the Integrated Information Theory of Consciousness. Entropy 2021, 23, 360. [Google Scholar] [CrossRef] [PubMed]
- Toker, D.; Sommer, F.T. Information integration in large brain networks. PLoS Comput. Biol. 2019, 15, e1006807. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Li, R.; Shu, L.; He, Z.Y.; Wang, J.N.; Zhang, C.M.; Ma, H.F.; Aihara, K.; Chen, L.N. Predicting future dynamics from short-term time series using an Anticipated Learning Machine. Natl. Sci. Rev. 2020, 7, 1079–1091. [Google Scholar] [CrossRef] [Green Version]
- Palmer, T. Human creativity and consciousness: Unintended consequences of the brain‘s extraordinary energy efficiency? Entropy 2020, 22, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaefer, K.; Stella, F.; McNaughton, B.L.; Battaglia, F.P. Replay, the default mode network and the cascaded memory systems model. Nat. Rev. Neurosci. 2022, 23, 628–640. [Google Scholar] [CrossRef]
- Schreiner, T.; Staudigl, T. Electrophysiological signatures of memory reactivation in humans. Philos. Trans. R. Soc. B 2020, 375, 20190293. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Z.; Nour, M.M.; Schuck, N.W.; Behrens, T.E.J.; Dolan, R.J. Decoding cognition from spontaneous neural activity. Nat. Rev. Neurosci. 2022, 23, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Whittington, J.C.R.; McCaffary, D.; Bakermans, J.J.W.; Behrens, T.E.J. How to build a cognitive map. Nat. Neurosci. 2022, 25, 1257–1272. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.P.; Prescott, T.J. Scaffolding layered control architectures through constraint closure: Insights into brain evolution and development. Philos. Trans. R. Soc. B 2022, 377, 20200519. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.; Di Giovanni, D.A.; Gomez-Pilar, J.; Nakao, T.; Huang, Z.; Longtin, A.; Northoff, G. The temporal signature of self: Temporal measures of resting-state EEG predict self-consciousness. Hum. Brain Mapp. 2019, 40, 789–803. [Google Scholar] [CrossRef] [Green Version]
- Brookshire, G. Putative rhythms in attentional switching can be explained by aperiodic temporal structure. Nat. Hum. Behav. 2022, 6, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Yeshurun, Y.; Nguyen, M.; Hasson, U. The default mode network: Where the idiosyncratic self meets the shared social world. Nat. Rev. Neurosci. 2021, 22, 181–192. [Google Scholar] [CrossRef]
- Jang, S.H.; Kwon, Y.H. Effect of repetitive transcranial magnetic stimulation on the ascending reticular activating system in a patient with disorder of consciousness: A case report. BMC Neurol. 2020, 20, 37. [Google Scholar] [CrossRef]
- Xia, X.Y.; Wang, Y.; Li, C.; Li, X.L.; He, J.H.; Bai, Y. Transcranial magnetic stimulation-evoked connectivity reveals modulation effects of repetitive transcranial magnetic stimulation on patients with disorders of consciousness. Neuroreport 2019, 30, 1307–1315. [Google Scholar] [CrossRef]
- Iimori, T.; Nakajima, S.; Miyazaki, T.; Tarumi, R.; Ogyu, K.; Wada, M.; Tsugawa, S.; Masuda, F.; Daskalakis, Z.J.; Blumberger, D.M.; et al. Effectiveness of the prefrontal repetitive transcranial magnetic stimulation on cognitive profiles in depression, schizophrenia, and Alzheimer‘s disease: A systematic review. Prog. Neuro–Psychoph. 2019, 88, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Q.; Gao, T.Y.; Zhang, T.; Li, W.X.; Wu, T.Y.; Han, X.C.; Han, S.H. Neural dynamics of racial categorization predicts racial bias in face recognition and altruism. Nat. Hum. Behav. 2020, 4, 69–87. [Google Scholar] [CrossRef]
- He, Z.; Muhlert, N.; Elliott, R. Emotion regulation of social exclusion: A cross-cultural study. Hum. Soc. Sci. Comm. 2021, 8, 173. [Google Scholar] [CrossRef]
- Han, S. The Sociocultural Brain: A Cultural Neuroscience Approach to Human Nature; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Luo, S.Y.; Zhu, Y.Y.; Han, S.H. Functional connectome fingerprint of holistic-analytic cultural style. Soc. Cogn. Affect. Neurosci. 2022, 17, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.Y.; Han, X.C.; Bang, D.; Han, S.H. Cultural differences in neurocognitive mechanisms underlying believing. Neuroimage 2022, 250, 118954. [Google Scholar] [CrossRef] [PubMed]
- von Suchodoletz, A.; Hepach, R. Cultural values shape the expression of self-evaluative social emotions. Sci. Rep. 2021, 11, 1369. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Jorgensen, N.A.; Telzer, E.H. A Call for Greater Attention to Culture in the Study of Brain and Development. Perspect. Psychol. Sci. 2021, 16, 275–293. [Google Scholar] [CrossRef]
- Richerson, P.J.; Boyd, R. Not by Genes Alone: How Culture Transformed Human Evolution; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Hari, R.; Henriksson, L.; Malinen, S.; Parkkonen, L. Centrality of social interaction in human brain function. Neuron 2015, 88, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raichle, M.E. The Brain’s Default Mode Network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smallwood, J.; Bernhardt, B.C.; Leech, R.; Bzdok, D.; Jefferies, E.; Margulies, D.S. The default mode network in cognition: A topographical perspective. Nat. Rev. Neurosci. 2021, 22, 503–513. [Google Scholar] [CrossRef]
- O‘Doherty, J.P.; Lee, S.W.; Tadayonnejad, R.; Cockburn, J.; Iigaya, K.; Charpentier, C.J. Why and how the brain weights contributions from a mixture of experts. Neurosci. Biobehav. Rev. 2021, 123, 14–23. [Google Scholar] [CrossRef]
- Sharot, T.; Sunstein, C.R. How people decide what they want to know. Nat. Hum. Behav. 2020, 4, 14–19. [Google Scholar] [CrossRef]
- LeDoux, J.E. As soon as there was life, there was danger: The deep history of survival behaviours and the shallower history of consciousness. Philos. Trans. R. Soc. B 2022, 377, 20210292. [Google Scholar] [CrossRef]
- Cisek, P. Evolution of behavioural control from chordates to primates. Philos. Trans. R. Soc. B 2022, 377, 20200522. [Google Scholar] [CrossRef]
- Snell-Rood, E.; Snell-Rood, C. The developmental support hypothesis: Adaptive plasticity in neural development in response to cues of social support. Philos. Trans. R. Soc. B 2020, 375, 20190491. [Google Scholar] [CrossRef]
- Kanaev, I.A. Evolutionary origin and the development of consciousness. Neurosci. Biobehav. Rev. 2022, 133, 104511. [Google Scholar] [CrossRef]
- Tavoni, G.; Doi, T.; Pizzica, C.; Balasubramanian, V.; Gold, J.I. Human inference reflects a normative balance of complexity and accuracy. Nat. Hum. Behav. 2022, 6, 1153–1168. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; McAvoy, A.; Mori, Y.; Plotkin, J.B. Evolution of prosocial behaviours in multilayer populations. Nat. Hum. Behav. 2022, 6, 338–348. [Google Scholar] [CrossRef]
- Alu, F.; Miraglia, F.; Orticoni, A.; Judica, E.; Cotelli, M.; Rossini, P.M.; Vecchio, F. Approximate Entropy of Brain Network in the Study of Hemispheric Differences. Entropy 2020, 22, 1220. [Google Scholar] [CrossRef] [PubMed]
- Luppi, A.I.; Craig, M.M.; Pappas, I.; Finoia, P.; Williams, G.B.; Allanson, J.; Pickard, J.D.; Owen, A.M.; Naci, L.; Menon, D.K.; et al. Consciousness-specific dynamic interactions of brain integration and functional diversity. Nat. Comm. 2019, 10, 4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burioka, N.; Miyata, M.; Cornelissen, G.; Halberg, F.; Takeshima, T.; Kaplan, D.T.; Suyama, H.; Endo, M.; Maegaki, Y.; Nomura, T.; et al. Approximate entropy in the electroencephalogram during wake and sleep. Clin. EEG Neurosci. 2005, 36, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Volz, L.J.; Gazzaniga, M.S. Interaction in isolation: 50 years of insights from split-brain research. Brain 2017, 140, 2051–2060. [Google Scholar] [CrossRef] [PubMed]
- Fried, I. Neurons as will and representation. Nat. Rev. Neurosci. 2022, 23, 104–114. [Google Scholar] [CrossRef]
- Padilla-Coreano, N.; Tye, K.M.; Zelikowsky, M. Dynamic influences on the neural encoding of social valence. Nat. Rev. Neurosci. 2022, 23, 535–550. [Google Scholar] [CrossRef]
- Tonelli, G. Genesis: The Story of How Everything Began; Farrar, Straus and Giroux: New York, NY, USA, 2021. [Google Scholar]
- Kauffman, S.A. A World Beyond Physics: The Emergence and Evolution of Life; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Richerson, P.J.; Gavrilets, S.; de Waal, F.B.M. Modern theories of human evolution foreshadowed by Darwin’s Descent of Man. Science 2021, 372, eaba3776. [Google Scholar] [CrossRef]
- Schrödinger, E. What Is Life? & Mind and Matter; Cambridge University Press: Cambridge, UK, 1974. [Google Scholar]
- Anokhin, P.K. Selected Works. Philosophical Aspects of the Theory of Functional Systems; Science: Moscow, Russia, 1978. [Google Scholar]
- Jekely, G. The chemical brain hypothesis for the origin of nervous systems. Philos. Trans. R. Soc. B 2021, 376, 20190761. [Google Scholar] [CrossRef]
- Puscian, A.; Bryksa, A.; Kondrakiewicz, L.; Kostecki, M.; Winiarski, M.; Knapska, E. Ability to share emotions of others as a foundation of social learning. Neurosci. Biobehav. Rev. 2022, 132, 23–36. [Google Scholar] [CrossRef]
- Popiel, N.J.M.; Khajehabdollahi, S.; Abeyasinghe, P.M.; Riganello, F.; Nichols, E.S.; Owen, A.M.; Soddu, A. The emergence of integrated information, complexity, and ‘consciousness’ at criticality. Entropy 2020, 22, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigogine, I.; Stengers, I. Order out of Chaos: Man‘s New Dialogue with Nature; Flamingo: Temecula, CA, USA, 1984. [Google Scholar]
- Wang, R. Yinyang: The Way of Heaven and Earth in Chinese Thought and Culture. Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Jékely, G.; Godfrey-Smith, P.; Keijzer, F. Reafference and the origin of the self in early nervous system evolution. Philos. Trans. R. Soc. B 2021, 376, 20190764. [Google Scholar] [CrossRef] [PubMed]
- Suryanarayana, S.M.; Robertson, B.; Grillner, S. The neural bases of vertebrate motor behaviour through the lens of evolution. Philos. Trans. R. Soc. B 2022, 377, 20200521. [Google Scholar] [CrossRef] [PubMed]
- Liden, W.H.; Phillips, M.L.; Herberholz, J. Neural control of behavioural choice in juvenile crayfish. Proc. R. Soc. B-Biol. Sci. 2010, 277, 3493–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, K.Y.; Jekely, G. Origins of eukaryotic excitability. Philos. Trans. R. Soc. B 2021, 376, 20190758. [Google Scholar] [CrossRef]
- Lyon, P.; Keijzer, F.; Arendt, D.; Levin, M. Reframing cognition: Getting down to biological basics. Philos. Trans. R. Soc. B 2021, 376, 20190750. [Google Scholar] [CrossRef]
- Bryer, M.A.H.; Koopman, S.E.; Cantlon, J.F.; Piantadosi, S.T.; MacLean, E.L.; Baker, J.M.; Beran, M.J.; Jones, S.M.; Jordan, K.E.; Mahamane, S.; et al. The evolution of quantitative sensitivity. Philos. Trans. R. Soc. B 2022, 377, 20200529. [Google Scholar] [CrossRef]
- Bechtel, W.; Bich, L. Grounding cognition: Heterarchical control mechanisms in biology. Philos. Trans. R. Soc. B 2021, 376, 20190751. [Google Scholar] [CrossRef]
- Schaap, P. From environmental sensing to developmental control: Cognitive evolution in dictyostelid social amoebas. Philos. Trans. R. Soc. B 2021, 376, 20190756. [Google Scholar] [CrossRef] [PubMed]
- Wielgoss, S.; Wolfensberger, R.; Sun, L.; Fiegna, F.; Velicer, G.J. Social genes are selection hotspots in kin groups of a soil microbe. Science 2019, 363, 1342–1345. [Google Scholar] [CrossRef] [PubMed]
- Pen, I.; Flatt, T. Asymmetry, division of labour and the evolution of ageing in multicellular organisms. Philos. Trans. R. Soc. B 2021, 376, 20190729. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, M.; Cini, A.; Alberti, S.; Salaris, E.; Theodorou, P.; Floris, I.; Satta, A. Honey bees increase social distancing when facing the ectoparasite Varroa destructor. Sci. Adv. 2021, 7, eabj1398. [Google Scholar] [CrossRef]
- Turner, L. Emotional bonds: Bridging the gap between evolutionary and humanistic accounts of religious belief. Arch. Psychol. Relig. 2020, 42, 6–28. [Google Scholar] [CrossRef]
- Hrdy, S.B.; Burkart, J.M. The emergence of emotionally modern humans: Implications for language and learning. Philos. Trans. R. Soc. B 2020, 375, 20190499. [Google Scholar] [CrossRef]
- Tomasello, M. The adaptive origins of uniquely human sociality. Philos. Trans. R. Soc. B 2020, 375, 20190493. [Google Scholar] [CrossRef]
- Richerson, P.J.; Boyd, R. The human life history is adapted to exploit the adaptive advantages of culture. Philos. Trans. R. Soc. B 2020, 375, 20190498. [Google Scholar] [CrossRef] [PubMed]
- Uomini, N.; Fairlie, J.; Gray, R.D.; Griesser, M. Extended parenting and the evolution of cognition. Philos. Trans. R. Soc. B 2020, 375, 20190495. [Google Scholar] [CrossRef]
- Gurven, M.; Davison, R.J.; Kraft, T. The optimal timing of teaching and learning across the life course. Philos. Trans. R. Soc. B 2020, 375, 20190500. [Google Scholar] [CrossRef] [PubMed]
- Strassberg, S.S.; Creanza, N. Cultural evolution and prehistoric demography. Philos. Trans. R. Soc. B 2021, 376, 20190713. [Google Scholar] [CrossRef]
- Deffner, D.; McElreath, R. The importance of life history and population regulation for the evolution of social learning. Philos. Trans. R. Soc. B 2020, 375, 20190492. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Parkinson, C. Navigating social knowledge. Nat. Neurosci. 2021, 24, 1195–1197. [Google Scholar] [CrossRef] [PubMed]
- O’Madagain, C.; Tomasello, M. Joint attention to mental content and the social origin of reasoning. Synthese 2021, 198, 4057–4078. [Google Scholar] [CrossRef] [Green Version]
- Mammen, M.; Koeymen, B.; Tomasello, M. Young children’s moral judgments depend on the social relationship between agents. Cogn. Dev. 2021, 57, 100973. [Google Scholar] [CrossRef]
- Li, L.; Britvan, B.; Tomasello, M. Young children conform more to norms than to preferences. PLoS ONE 2021, 16, e0251228. [Google Scholar] [CrossRef]
- Kanaev, I.A.; Dryaeva, E.D. Evolutionary Approach to Consciousness in Modern Philosophy and Interdisciplinary Studies. Vop. Filos. 2022, 2, 106–116. [Google Scholar] [CrossRef]
- Prinz, W. Seeing some one. Front. Psychol. 2018, 9, 1747. [Google Scholar] [CrossRef]
- Antón, S.C.; Potts, R.; Aiello, L.C. Human evolution. Evolution of early Homo: An integrated biological perspective. Science 2014, 345, 1236828. [Google Scholar] [CrossRef]
- Callaway, E. Oldest human DNA reveals recent Neanderthal mixing. Nature 2021, 592, 339. [Google Scholar] [CrossRef]
- Vaesen, K.; Scherjon, F.; Hemerik, L.; Verpoorte, A. Inbreeding, Allee effects and stochasticity might be sufficient to account for Neanderthal extinction. PLoS ONE 2019, 14, e0225117. [Google Scholar] [CrossRef]
- Read, D.W. Working Memory: A Cognitive Limit to Non-Human Primate Recursive Thinking Prior to Hominid Evolution. Evol. Psychol. 2008, 6, 676–714. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Shultz, S. Social complexity and the fractal structure of group size in primate social evolution. Biol. Rev. 2021, 96, 1889–1906. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, R.I.M. Religion, the social brain and the mystical stance. Arch. Psychol. Relig. 2020, 42, 46–62. [Google Scholar] [CrossRef]
- Pettitt, P. Hominin evolutionary thanatology from the mortuary to funerary realm: The palaeoanthropological bridge between chemistry and culture. Philos. Trans. R. Soc. B 2018, 373, 20180212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettitt, P.; Anderson, J.R. Primate thanatology and hominoid mortuary archeology. Primates 2020, 61, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Northoff, G.; Vogeley, K.; Wexler, B.E.; Kitayama, S.; Varnum, M.E. A cultural neuroscience approach to the biosocial nature of the human brain. Annu. Rev. Psychol. 2013, 64, 335–359. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Northoff, G. Culture-sensitive neural substrates of human cognition: A transcultural neuroimaging approach. Nat. Rev. Neurosci. 2008, 9, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Han, S. Neurocognitive Basis of Racial Ingroup Bias in Empathy. Trends Cogn. Sci. 2018, 22, 400–421. [Google Scholar] [CrossRef] [PubMed]
- Zmigrod, L.; Tsakiris, M. Computational and neurocognitive approaches to the political brain: Key insights and future avenues for political neuroscience. Philosop Philos. Trans. R. Soc. B 2021, 376, 20200130. [Google Scholar] [CrossRef]
- Bélanger, J.J. The sociocognitive processes of ideological obsession: Review and policy implications. Philos. Trans. R. Soc. B 2021, 376, 20200144. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanaev, I.A. Entropy and Cross-Level Orderliness in Light of the Interconnection between the Neural System and Consciousness. Entropy 2023, 25, 418. https://doi.org/10.3390/e25030418
Kanaev IA. Entropy and Cross-Level Orderliness in Light of the Interconnection between the Neural System and Consciousness. Entropy. 2023; 25(3):418. https://doi.org/10.3390/e25030418
Chicago/Turabian StyleKanaev, Ilya A. 2023. "Entropy and Cross-Level Orderliness in Light of the Interconnection between the Neural System and Consciousness" Entropy 25, no. 3: 418. https://doi.org/10.3390/e25030418
APA StyleKanaev, I. A. (2023). Entropy and Cross-Level Orderliness in Light of the Interconnection between the Neural System and Consciousness. Entropy, 25(3), 418. https://doi.org/10.3390/e25030418