
Lepidium sativum Secondary Metabolites (Essential Oils): In Vitro and In Silico Studies on Human Hepatocellular Carcinoma Cell Lines


 ,
,  and
and 
Abstract
:
1. Introduction
2. Results
2.1. Extraction
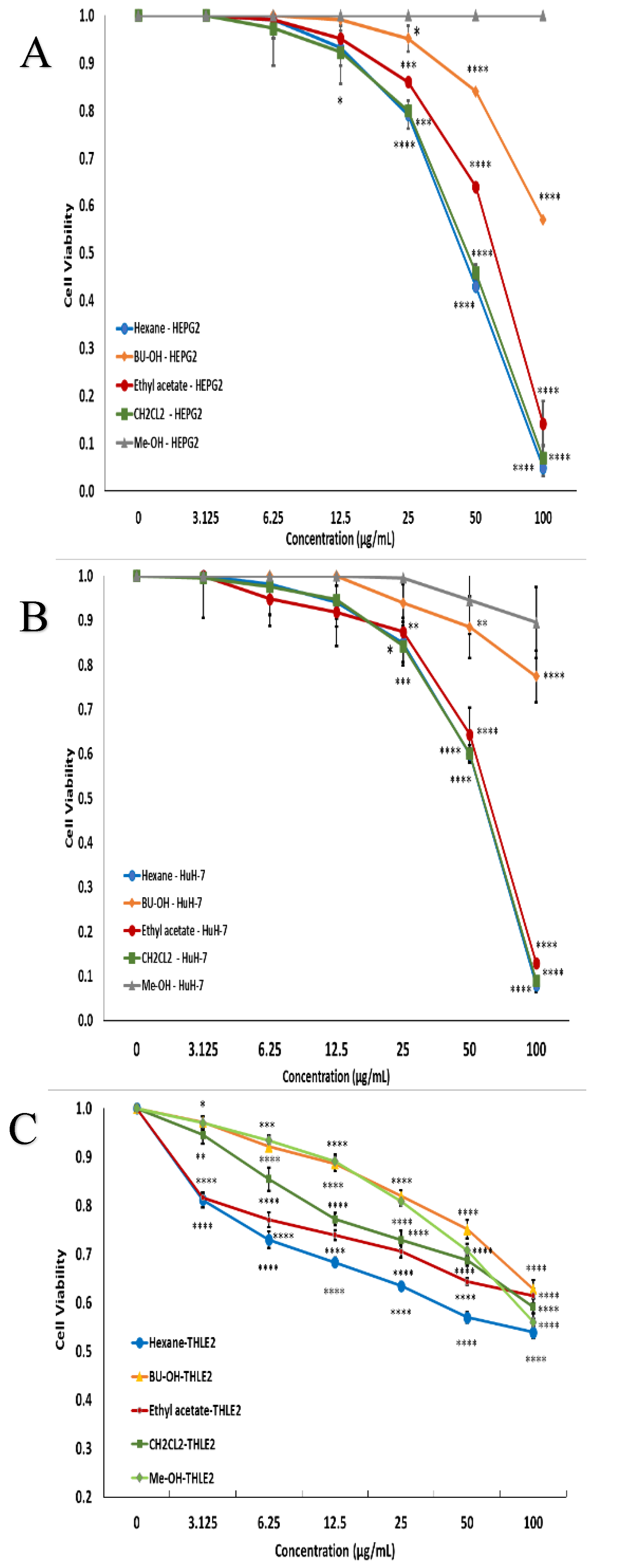
2.2. Cell Cytotoxicity
2.3. Gas Chromatography (GC) Analysis
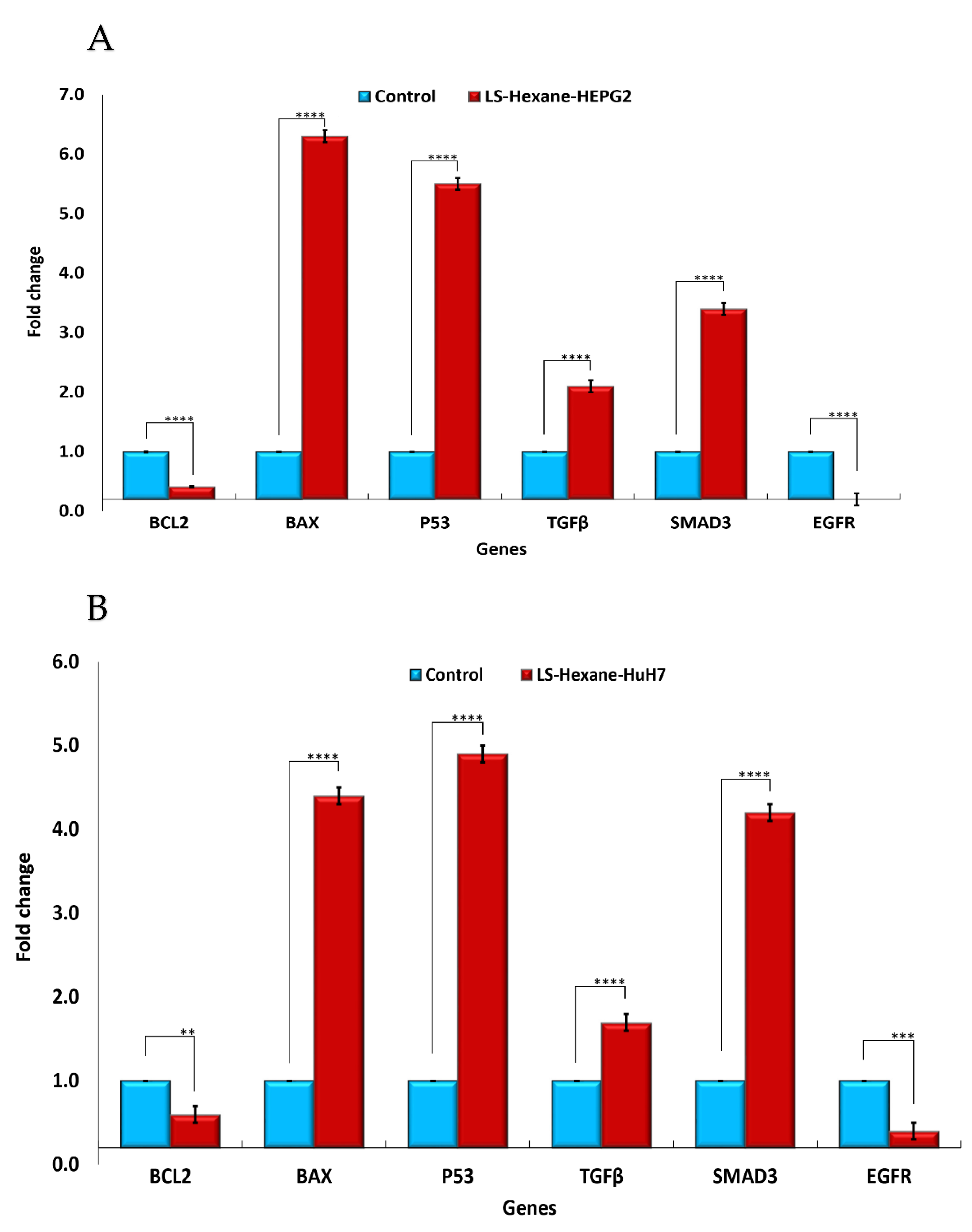
2.4. Effect of L. sativum Extracts on Expression of EGFR and BCL2, TGF-β, SMAD3, BAX, and P53 Genes
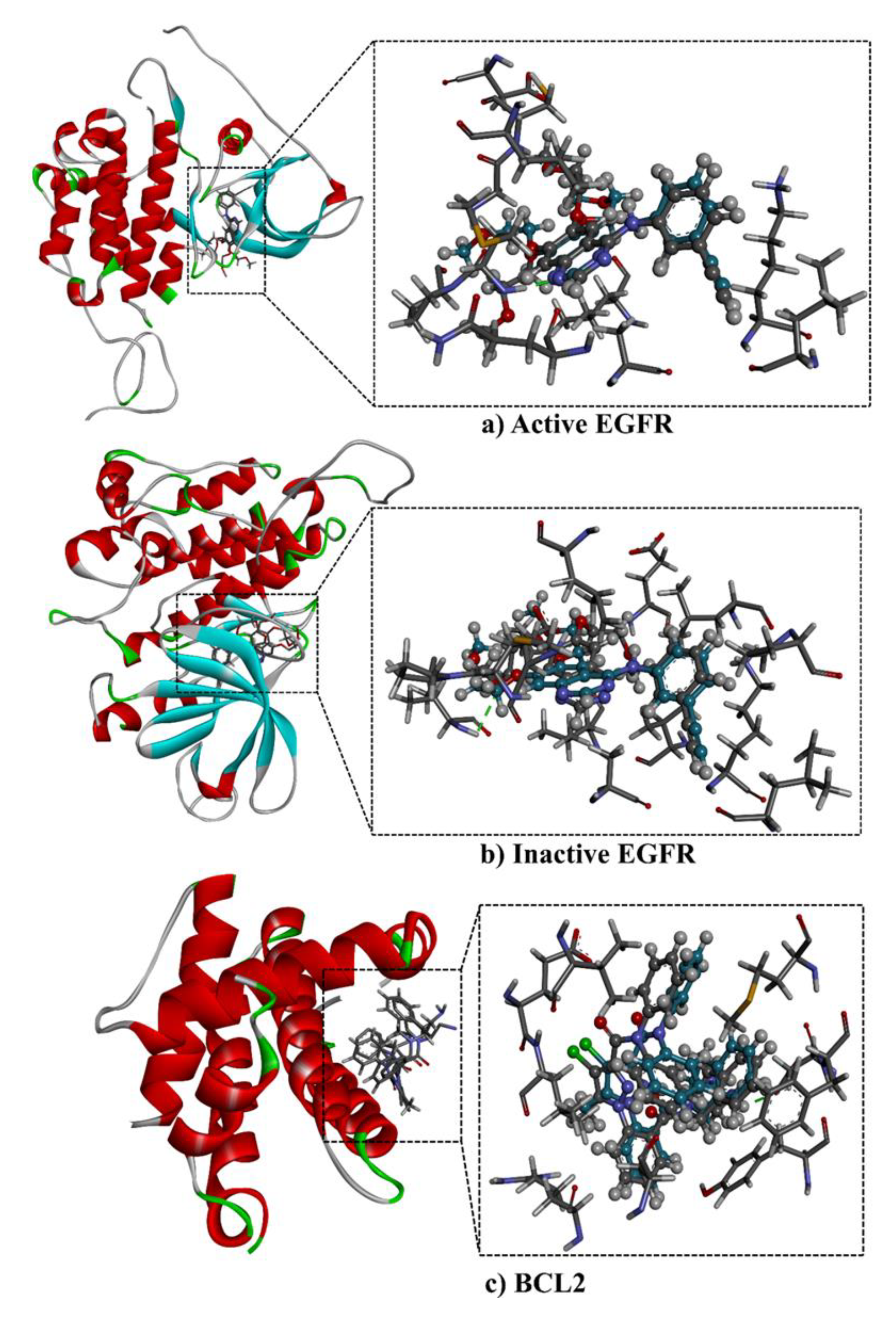
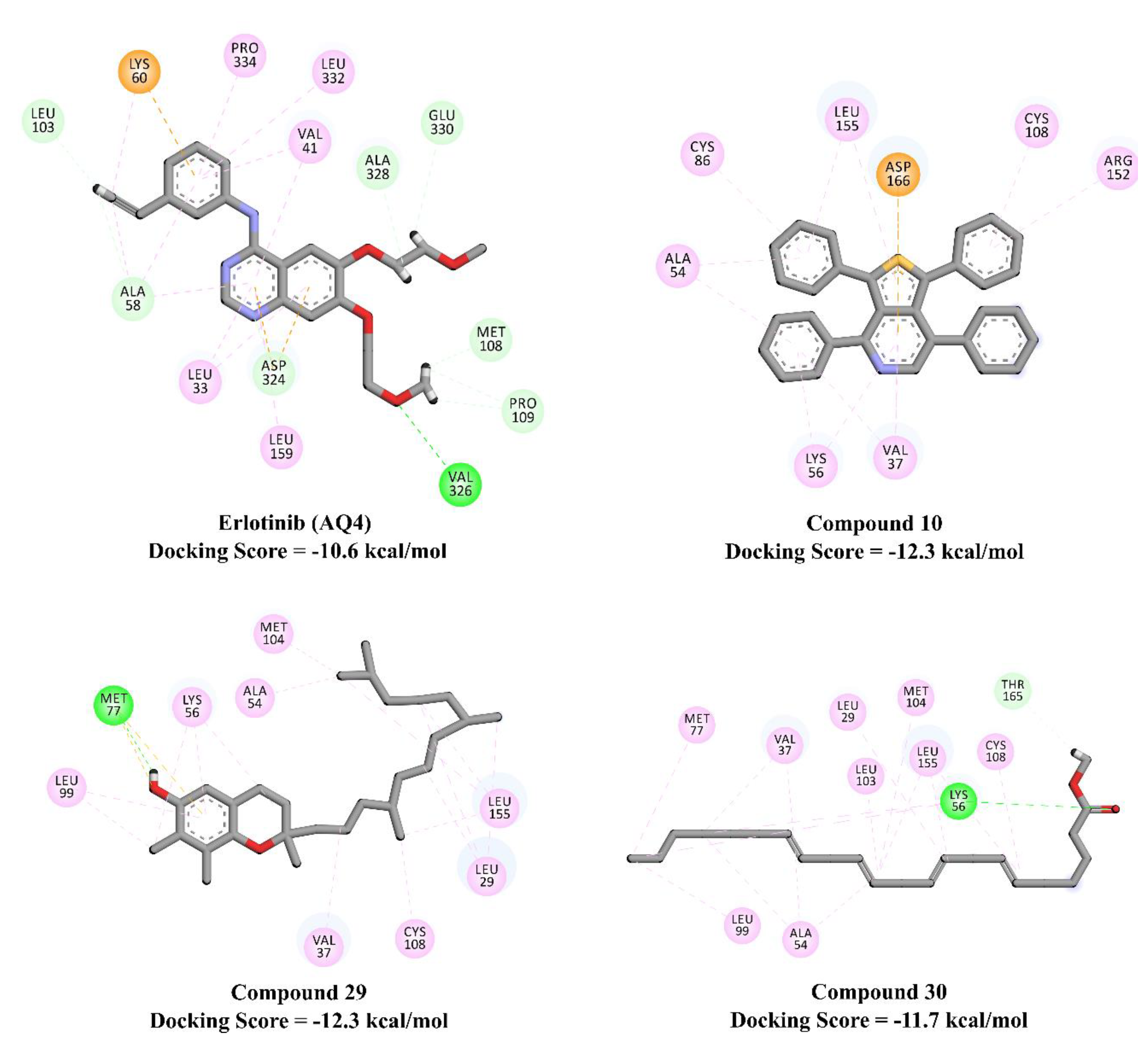
2.5. Molecular Docking Calculations and Pathway Enrichment Analysis (PEA)
3. Discussion
4. Materials and Methods
4.1. Method of Extraction
4.2. Cell Line Culture
4.3. MTT Assay
4.4. Gas Chromatography
4.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.6. Molecular Docking and Pathway Enrichment Analysis (PEA)
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castelli, G.; Pelosi, E.; Testa, U. Liver Cancer: Molecular Characterization, Clonal Evolution and Cancer Stem Cells. Cancers 2017, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Rashed, W.M.; Kandeil, M.A.M.; Mahmoud, M.; Ezzat, S. Hepatocellular Carcinoma (HCC) in Egypt: A comprehensive overview. J. Egypt. Natl. Cancer Inst. 2020, 32, 1–11. [Google Scholar] [CrossRef] [Green Version]
- El-Zayadi, A.-R.; Badran, H.M.; Barakat, E.M.F.; Attia, M.E.-D.; Shawky, S.; Mohamed, M.K.; Selim, O.; Saeid, A. Hepatocellular carcinoma in Egypt: A single center study over a decade. World J. Gastroenterol. 2005, 11, 5193–5198. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Khaled, H.M.; Mikhail, N.N.; Baraka, H.; Kamel, H. Cancer Incidence in Egypt: Results of the National Population-Based Cancer Registry Program. J. Cancer Epidemiol. 2014, 2014, 1–18. [Google Scholar] [CrossRef]
- Xiao, Q.; Zhu, W.; Feng, W.; Lee, S.S.; Leung, A.W.; Shen, J.; Gao, L.; Xu, C. A Review of Resveratrol as a Potent Chemoprotective and Synergistic Agent in Cancer Chemotherapy. Front. Pharmacol. 2019, 9, 1534. [Google Scholar] [CrossRef]
- Shukla, S.; Ohnuma, S.; Ambudkar, S.V. Improving cancer chemotherapy with modulators of ABC drug transporters. Curr. Drug Targets 2011, 12, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.A.; Hegazy, M.-E.F.; Hassan, N.M.; Wojcinska, M.; Karchesy, J.; Pare, P.W.; Mabry, T.J. Constituents of Chrysothamnus viscidiflorus. Phytochemistry 2006, 67, 1547–1553. [Google Scholar] [CrossRef]
- Xiao, Q.; Wu, J.; Pang, X.; Jiang, Y.; Wang, P.; Leung, A.W.; Gao, L.; Jiang, S.; Xu, C.; Xiao, J.W.Q. Discovery and Development of Natural Products and their Derivatives as Photosensitizers for Photodynamic Therapy. Curr. Med. Chem. 2018, 25, 839–860. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, M.-E.F.; Abdelfatah, S.; Hamed, A.R.; Mohamed, T.A.; Elshamy, A.A.; Saleh, I.A.; Reda, E.H.; Abdel-Azim, N.S.; Shams, K.A.; Sakr, M.; et al. Cytotoxicity of 40 Egyptian plant extracts targeting mechanisms of drug-resistant cancer cells. Phytomedicine 2018, 59, 152771. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. Int. J. Kuwait Univ. Health Sci. Cent. 2015, 25 (Suppl. 2), 41–59. [Google Scholar] [CrossRef]
- Al-Yahya, M.; Mossa, J.; Ageel, A.; Rafatullah, S. Pharmacological and safety evaluation studies on Lepidium sativum L., Seeds. Phytomed. Int. J. Phytother. Phytopharm. 1994, 1, 155–159. [Google Scholar] [CrossRef]
- Falana, H.; Nofal, W.; Nakhleh, H. A review article Lepidium sativum (Garden cress). In Pharm-D Program, College of Nursing, Pharmacy and Health Professions; Birzeit University: Birzeit, Palestine, 2014; pp. 1–8. [Google Scholar]
- Alqahtani, F.Y.; Aleanizy, F.S.; Mahmoud, A.Z.; Farshori, N.N.; Alfaraj, R.; Al-Sheddi, E.S.; Alsarra, I. Chemical composition and antimicrobial, antioxidant, and anti-inflammatory activities of Lepidium sativum seed oil. Saudi J. Biol. Sci. 2018, 26, 1089–1092. [Google Scholar] [CrossRef] [PubMed]
- Diwakar, B.T.; Lokesh, B.R.; Naidu, K.A. Modulatory effect of α-linolenic acid-rich garden cress (Lepidium sativum L.) seed oil on inflammatory mediators in adult albino rats. Br. J. Nutr. 2011, 106, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigoniya, P.; Shukla, A. Phytopharmacological screening of Lepidium sativum seeds total alkaloid: Hepatoprotective, antidiabetic and in vitro antioxidant activity along with identification by LC/MS/MS. PharmaNutrition 2014, 2, 90. [Google Scholar] [CrossRef]
- Raghavendra, R.H. Eugenol and n-3 Rich Garden Cress Seed Oil as Modulators of Platelet Aggregation and Eicosanoids in Wistar Albino Rats. Open Nutraceuticals J. 2011, 4, 144–150. [Google Scholar] [CrossRef]
- Adam, S.I.; Salih, S.A.; Abdelgadir, W.S. “In vitro” Antimicrobial Assessment of “Lepidium sativum” L. Seeds Extracts. Asian J. Med. Sci. 2011, 3, 261–266. [Google Scholar]
- Al-Fatimi, M.; Friedrich, U.; Jenett-Siems, K. Cytotoxicity of plants used in traditional medicine in Yemen. Fitoterapia 2005, 76, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Aslani, E.; Naghsh, N.; Ranjbar, M. Cytotoxic effects of hydroalcoholic extracts of cress (Lepidium sativum)—Made from different stages of the plant—On k562 Leukemia cell line. Hormozgan Med. J. 2015, 18, 411–419. Available online: https://www.sid.ir/en/journal/ViewPaper.aspx?id=531695 (accessed on 15 February 2021).
- Mahassni, S.H.; Al-Reemi, R.M. Apoptosis and necrosis of human breast cancer cells by an aqueous extract of garden cress (Lepidium sativum) seeds. Saudi J. Biol. Sci. 2013, 20, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Raish, M.; Ahmad, A.; Alkharfy, K.M.; Ahamad, S.R.; Mohsin, K.; Al-Jenoobi, F.I.; Al-Mohizea, A.M.; Ansari, M.A. Hepatoprotective activity of Lepidium sativum seeds against D-galactosamine/lipopolysaccharide induced hepatotoxicity in animal model. BMC Complement. Altern. Med. 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Saleem, T.S.M.; Chetty, C.M.; Ramkanth, S.; Rajan, V.S.T.; Kumar, K.M.; Gauthaman, K. Hepatoprotective Herbs—A Review. Int. J. Res. Pharm. Sci. 2010, 1, 1–5. Available online: https://www.pharmascope.org/index.php/ijrps/article/view/78 (accessed on 18 January 2021).
- Aboul-Soud, M.A.M.; Ashour, A.E.; Challis, J.K.; Ahmed, A.F.; Kumar, A.; Nassrallah, A.; AlAhmari, T.A.; Saquib, Q.; Siddiqui, M.A.; Al-Sheikh, Y.; et al. Biochemical and Molecular Investigation of In Vitro Antioxidant and Anticancer Activity Spectrum of Crude Extracts of Willow Leaves Salix safsaf. Plants 2020, 9, 1295. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhang, Q.; Zhao, J.; Liao, M.; Wen, S.; Yang, M. Phosphorylation of Bcl-2 plays an important role in glycochenodeoxycholate-induced survival and chemoresistance in HCC. Oncol. Rep. 2017, 38, 1742–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Murata, M.; Yamaguchi, T.; Zaki, K.M. TGF-β/Smad signaling during hepatic fibro-carcinogenesis (Review). Int. J. Oncol. 2014, 45, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, B.C.; Hoshida, Y.; Fujii, T.; Wei, L.; Yamada, S.; Lauwers, G.Y.; McGinn, C.M.; Deperalta, D.K.; Chen, X.; Kuroda, T.; et al. Epidermal growth factor receptor inhibition attenuates liver fibrosis and development of hepatocellular carcinoma. Hepatology 2013, 59, 1577–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazy, M.-E.F.; ElShamy, A.I.; Mohamed, T.A.; Hamed, A.R.; Ibrahim, M.A.A.; Ohta, S.; Paré, P.W. Cembrene Diterpenoids with Ether Linkages from Sarcophyton ehrenbergi: An Anti-Proliferation and Molecular-Docking Assessment. Mar. Drugs 2017, 15, 192. [Google Scholar] [CrossRef]
- Elsisi, G.H.; Aburawash, A.; Waked, E. Cost-Effectiveness Analysis of New HCV Treatments in Egyptian Cirrhotic and Non-Cirrhotic Patients: A Societal Perspective. Value Health Reg. Issues 2017, 13, 7–15. [Google Scholar] [CrossRef]
- Desai, A.; Qazi, G.; Ganju, R.; El-Tamer, M.; Singh, J.; Saxena, A.; Bedi, Y.; Taneja, S.; Bhat, H. Medicinal Plants and Cancer Chemoprevention. Curr. Drug Metab. 2008, 9, 581–591. [Google Scholar] [CrossRef] [Green Version]
- AlObaidi, L.A. Study the anticancer effect of Lepidium sativum leaves extract on squamous cell carcinoma (CAL-27) cell lines. J. Nat. Sci. Res. 2014, 4, 17. [Google Scholar]
- El-Kaream, S.A.A. Biochemical and biophysical study of chemopreventive and chemotherapeutic anti-tumor potential of some Egyptian plant extracts. Biochem. Biophys. Rep. 2019, 18, 100637. [Google Scholar] [CrossRef]
- Abuelgasim, A.I.; Nuha, H.S.; Mohammed, A.H. Hepatoprotective effect of Lepidium sativum against carbon tetrachloride induced damage in rats. Res. J. Anim. Vet. Sci. 2008, 3, 20–23. [Google Scholar]
- Balanis, N.; Carlin, C.R. Stress-induced EGF receptor signaling through STAT3 and tumor progression in triple-negative breast cancer. Mol. Cell. Endocrinol. 2017, 451, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Lanaya, H.; Natarajan, A.; Komposch, K.; Li, L.; Amberg, N.; Chen, L.; Wculek, S.; Hammer, M.; Zenz, R.; Peck-Radosavljevic, M.; et al. EGFR has a tumour-promoting role in liver macrophages during hepatocellular carcinoma formation. Nature 2014, 16, 972–981. [Google Scholar] [CrossRef]
- Komposch, K.; Sibilia, M. EGFR Signaling in Liver Diseases. Int. J. Mol. Sci. 2015, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 2017, 12, 3–20. [Google Scholar] [CrossRef]
- Principe, D.R.; Doll, J.A.; Bauer, J.; Jung, B.; Munshi, H.G.; Bartholin, L.; Pasche, B.; Lee, C.; Grippo, P.J. TGF-β: Duality of Function between Tumor Prevention and Carcinogenesis. J. Natl. Cancer Inst. 2014, 106, djt369. [Google Scholar] [CrossRef]
- Chen, J.; Gingold, J.A.; Su, X. Immunomodulatory TGF-β Signaling in Hepatocellular Carcinoma. Trends Mol. Med. 2019, 25, 1010–1023. [Google Scholar] [CrossRef] [Green Version]
- Dooley, S.; Dijke, P.T. TGF-β in progression of liver disease. Cell Tissue Res. 2011, 347, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, I.; Moreno-Caceres, J.; Sánchez, A.; Dooley, S.; Dewidar, B.; Giannelli, G.; Dijke, P.T.; IT-LIVER Consortium. TGF-β signalling and liver disease. FEBS J. 2016, 283, 2219–2232. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, I.; Caballero-Díaz, D. Transforming Growth Factor-β-Induced Cell Plasticity in Liver Fibrosis and Hepatocarcinogenesis. Front. Oncol. 2018, 8, 357. [Google Scholar] [CrossRef] [Green Version]
- Tu, S.; Huang, W.; Huang, C.; Luo, Z.; Yan, X. Contextual Regulation of TGF-β Signaling in Liver Cancer. Cells 2019, 8, 1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massagué, J. TGF-β signal transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-A.; Zhang, G.-M.; Feigenbaum, L.; Zhang, Y.E. Smad3 reduces susceptibility to hepatocarcinoma by sensitizing hepatocytes to apoptosis through downregulation of Bcl-2. Cancer Cell 2006, 9, 445–457. [Google Scholar] [CrossRef] [Green Version]
- Ventura, A.; Kirsch, D.G.; McLaughlin, M.E.; Tuveson, D.A.; Grimm, J.; Lintault, L.; Newman, J.; Reczek, E.E.; Weissleder, R.; Jacks, T. Restoration of p53 function leads to tumour regression in vivo. Nature 2007, 445, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Swapana, N.; Tominaga, T.; Elshamy, A.I.; Ibrahim, M.A.; Hegazy, M.-E.F.; Singh, C.B.; Suenaga, M.; Imagawa, H.; Noji, M.; Umeyama, A. Kaemgalangol A: Unusual seco-isopimarane diterpenoid from aromatic ginger Kaempferia galanga. Fitoterapia 2018, 129, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- El-Kareem, M.S.M.A.; Rabbih, M.A.E.F.; Selim, E.T.M.; Elsherbiny, E.A.E.-M.; El-Khateeb, A. Application of GC/EIMS in Combination with Semi-Empirical Calculations for Identification and Investigation of Some Volatile Components in Basil Essential Oil. Int. J. Anal. Mass Spectrom. Chromatogr. 2016, 04, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Morris, G.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Stamos, J.; Sliwkowski, M.X.; Eigenbrot, C. Structure of the Epidermal Growth Factor Receptor Kinase Domain Alone and in Complex with a 4-Anilinoquinazoline Inhibitor. J. Biol. Chem. 2002, 277, 46265–46272. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Liu, Y.; Lemmon, M.A.; Radhakrishnan, R. Erlotinib binds both inactive and active conformations of the EGFR tyrosine kinase domain. Biochem. J. 2012, 448, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, J.; Payne, A.; de Candole, B.; Ford, D.; Hutchinson, B.; Trevitt, G.; Turner, J.; Edwards, C.; Watkins, C.; Whitcombe, I.; et al. Tetrahydroisoquinoline amide substituted phenyl pyrazoles as selective Bcl-2 inhibitors. Bioorganic. Med. Chem. Lett. 2009, 19, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.C.; Myers, J.B.; Folta, T.; Shoja, V.; Heath, L.S.; Onufriev, A. H++: A server for estimating pKas and adding missing hydrogens to macromolecules. Nucleic Acids Res. 2005, 33, W368–W371. [Google Scholar] [CrossRef] [PubMed]
- OpenEye Scientific Software. SZYBKI, 1.9.0.3; OpenEye Scientific Software: Santa Fe, NM, USA, 2016. [Google Scholar]
- Halgren, T.A. MMFF94s option for energy minimization studies. J. Comput. Chem. 1999, 20, 720–729. [Google Scholar] [CrossRef]
- Gasteiger, J.; Marsili, M. Iterative partial equalization of orbital electronegativity—A rapid access to atomic charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]
- Mohamed, T.A.; Elshamy, A.I.; Ibrahim, M.A.A.; Atia, M.A.M.; Ahmed, R.F.; Ali, S.K.; Mahdy, K.A.; Alshammari, S.O.; Al-Abd, A.M.; Moustafa, M.F.; et al. Gastroprotection against Rat Ulcers by Nephthea Sterol Derivative. Biomolecules 2021, 11, 1247. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2017, 46, D649–D655. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT (min) | Compound Name | Formula | M.W | Area% |
|---|---|---|---|---|---|
| Mono-Terpenoids | 13.38% | ||||
| 1 | 11.29 | Estragole | C10H12O | 148 | 1.44% |
| 2 | 13.03 | Alpha-terpinyl acetate | C12H20O2 | 196 | 2.36% |
| 3 | 13.27 | cis-p-Propenylanisole | C10H12O | 148 | 4.13% |
| 4 | 18.51 | gamma-Asarone | C12H16O3 | 208 | 0.68% |
| 5 | 15.77 | Methyleugenol | C11H14O2 | 178 | 4.77% |
| Sesqui-Terpenoids | 5.42% | ||||
| 6 | 12.3 | Copaene | C15H24 | 204 | 1.09% |
| 7 | 13.54 | Caryophyllene | C15H24 | 204 | 3.82% |
| 8 | 15.59 | Sesquisabinene isomer | C15H24 | 204 | 0.74% |
| 9 | 17.56 | Caryophyllene oxide | C15H24O | 220 | 0.86% |
| Hydrocarbons | 69.16% | ||||
| 10 | 4.06 | Thieno[3,4-c]pyridine-1,3-dione | C31H21NS | 439 | 1.65% |
| 11 | 19.6 | 2,4,6-Trimethylmandelic acid | C11H14O3 | 194 | 1.41% |
| 12 | 21.51 | Palmitic acid, methyl ester | C17H34O2 | 270 | 3.30% |
| 13 | 23.98 | 2-Nonadecanone | C19H38O | 282 | 2.40% |
| 14 | 24.14 | Oleic acid, methyl ester | C19H36O2 | 296 | 11.76% |
| 15 | 24.72 | Linolenic acid, methyl ester | C19H32O2 | 292 | 26.44% |
| 16 | 26.59 | 11-Eicosenoic acid, methyl ester | C21H40O2 | 324 | 4.80% |
| 17 | 27.61 | Cis-8,11,14-Eicosatrienoic Acid | C20H34O2 | 306 | 1.48% |
| 18 | 27.72 | 6,9,12,15-Docosatetraenoic acid, methyl ester | C23H38O2 | 498 | 4.15% |
| 19 | 28.88 | Methyl (Z)-13-docosenoate | C23H44O2 | 352 | 1.18% |
| 20 | 29.23 | 2-Hydroxy-1-(hydroxymethyl)ethyl (9z)-9-octadecenoate | C21H40O4 | 356 | 1.30% |
| 21 | 29.84 | 2-[(9Z,12Z)-9,12-Octadecadien-1-yloxy]ethanol | C20H38O2 | 310 | 0.52% |
| 22 | 31.18 | Meadowlactone | C20H38O2 | 310 | 2.65% |
| 23 | 31.46 | Dotriacontane | C32H66 | 450 | 0.94% |
| 24 | 33.35 | (Z)-18-Octadec-9-enolide | C18H32O2 | 280 | 5.18% |
| Others | 9.71% | ||||
| 25 | 14.98 | [2-methoxy-4-[(Z)-prop-1-enyl]phenyl] acetate | C10H12O2 | 164 | 1.15% |
| 26 | 18.91 | N-[(3-fluorophenyl)methyl]-7H-purin-6-amine | C12H10FN5 | 243 | 1.79% |
| 27 | 30.72 | 10-Nethoxy-Nb-alpha-methylcorynantheol | C21H29N2O2 | 341 | 1.03% |
| 28 | 28.94 | Propanoic acid,2-(3-acetoxy-4,4,14-trimethylandrost-8-en-17-yl) | C27H42O4 | 430 | 1.19% |
| 29 | 35.91 | gamma -Tocopherol | C28H48O2 | 416 | 3.55% |
| 30 | 36.79 | 9,12,15-Octadecatrienoic acid,2-phenyl-1,3-dioxan-5-yl ester | C28H40O4 | 440 | 1.00% |
| Total | 97.67% | ||||
| No. | Compound Name | Docking Score (kcal/mol) | ||
|---|---|---|---|---|
| EGFR | BCL2 | |||
| Active | Inactive | |||
| 1 | Estragole | −4.6 | −5.6 | −4.2 |
| 2 | Alpha-Terpinyl acetate | −5.8 | −7.7 | −5.3 |
| 3 | Anisole, p-propenyl-, cis- | −4.8 | −5.8 | −4.4 |
| 4 | Gamma-Asarone | −4.4 | −6.3 | −4.2 |
| 5 | Methyleugenol | −4.5 | −6.1 | −4.3 |
| 6 | Copaene | −6.8 | −9.0 | −6.5 |
| 7 | Caryophyllene | −6.3 | −7.1 | −6.0 |
| 8 | Sesquisabinene isomer | −6.4 | −8.4 | −6.0 |
| 9 | Caryophyllene oxide | −6.0 | −7.4 | −6.1 |
| 10 | Thieno[3,4-c]pyridine-1,3-dione | −8.5 | −12.3 | −7.4 |
| 11 | 2,4,6-Trimethylmandelic acid | −5.6 | −6.7 | −4.5 |
| 12 | Palmitic acid, methyl ester | −4.7 | −8.2 | −4.0 |
| 13 | 2-Nonadecanone | −4.8 | −9.1 | −3.9 |
| 14 | Oleic acid, methyl ester | −5.3 | −8.5 | −4.7 |
| 15 | Linolenic acid, methyl ester | −5.9 | −8.7 | −4.5 |
| 16 | 11-Eicosenoic acid, methyl ester | −5.4 | −7.8 | −4.7 |
| 17 | cis-8,11,14-Eicosatrienoic Acid | −5.3 | −9.8 | −5.4 |
| 18 | 6,9,12,15-Docosatetraenoic acid, methyl ester | −6.0 | −10.3 | −4.9 |
| 19 | Methyl (Z)-13-docosenoate | −3.1 | −9.9 | −4.5 |
| 20 | 2-Hydroxy-1-(hydroxymethyl)ethyl (9z)-9-octadecenoate | −2.4 | −8.3 | −4.4 |
| 21 | 2-[(9Z,12Z)-9,12-Octadecadien-1-yloxy]ethanol | −4.6 | −8.8 | −3.8 |
| 22 | Meadowlactone | −6.0 | −10.1 | −5.5 |
| 23 | Dotriacontane | −4.8 | −9.7 | −3.4 |
| 24 | (Z)-18-Octadec-9-enolide | −7.2 | −7.9 | −7.2 |
| 25 | 2-Methoxy-4-[(1e)-1-propenyl]phenol | −4.7 | −6.4 | −4.1 |
| 26 | N-[(3-fluorophenyl)methyl]-7H-purin-6-amine | −6.3 | −8.0 | −4.9 |
| 27 | 10-Nethoxy-Nb-alpha-methylcorynantheol | −7.9 | −7.3 | −5.9 |
| 28 | Propanoic acid,2-(3-acetoxy-4,4,14-trimethylandrost-8-en-17-yl) | −8.0 | −5.2 | −7.7 |
| 29 | gamma-Tocopherol | −8.5 | −12.3 | −7.0 |
| 30 | 9,12,15-Octadecatrienoic acid, 2-phenyl-1,3-dioxan-5-yl ester | −6.3 | −11.7 | −6.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazir, S.; El-Sherif, A.A.; Abdel-Ghani, N.T.; Ibrahim, M.A.A.; Hegazy, M.-E.F.; Atia, M.A.M. Lepidium sativum Secondary Metabolites (Essential Oils): In Vitro and In Silico Studies on Human Hepatocellular Carcinoma Cell Lines. Plants 2021, 10, 1863. https://doi.org/10.3390/plants10091863
Nazir S, El-Sherif AA, Abdel-Ghani NT, Ibrahim MAA, Hegazy M-EF, Atia MAM. Lepidium sativum Secondary Metabolites (Essential Oils): In Vitro and In Silico Studies on Human Hepatocellular Carcinoma Cell Lines. Plants. 2021; 10(9):1863. https://doi.org/10.3390/plants10091863
Chicago/Turabian StyleNazir, Shaimaa, Ahmed A. El-Sherif, Nour T. Abdel-Ghani, Mahmoud A. A. Ibrahim, Mohamed-Elamir F. Hegazy, and Mohamed A. M. Atia. 2021. "Lepidium sativum Secondary Metabolites (Essential Oils): In Vitro and In Silico Studies on Human Hepatocellular Carcinoma Cell Lines" Plants 10, no. 9: 1863. https://doi.org/10.3390/plants10091863
APA StyleNazir, S., El-Sherif, A. A., Abdel-Ghani, N. T., Ibrahim, M. A. A., Hegazy, M.-E. F., & Atia, M. A. M. (2021). Lepidium sativum Secondary Metabolites (Essential Oils): In Vitro and In Silico Studies on Human Hepatocellular Carcinoma Cell Lines. Plants, 10(9), 1863. https://doi.org/10.3390/plants10091863