Abstract
Phenolic compounds are an important class of plant secondary metabolites which play crucial physiological roles throughout the plant life cycle. Phenolics are produced under optimal and suboptimal conditions in plants and play key roles in developmental processes like cell division, hormonal regulation, photosynthetic activity, nutrient mineralization, and reproduction. Plants exhibit increased synthesis of polyphenols such as phenolic acids and flavonoids under abiotic stress conditions, which help the plant to cope with environmental constraints. Phenylpropanoid biosynthetic pathway is activated under abiotic stress conditions (drought, heavy metal, salinity, high/low temperature, and ultraviolet radiations) resulting in accumulation of various phenolic compounds which, among other roles, have the potential to scavenge harmful reactive oxygen species. Deepening the research focuses on the phenolic responses to abiotic stress is of great interest for the scientific community. In the present article, we discuss the biochemical and molecular mechanisms related to the activation of phenylpropanoid metabolism and we describe phenolic-mediated stress tolerance in plants. An attempt has been made to provide updated and brand-new information about the response of phenolics under a challenging environment.
1. Introduction
Plants face a plethora of biotic and abiotic stresses during their entire life which have negative impact on their growth, development, and productivity [1,2,3]. Biotic factors include insect pests, fungi, and weeds whereas abiotic stresses include salinity, drought, heavy metals, pesticides, ultraviolet (UV) radiation, as well as heat and cold stress [3,4,5,6,7,8,9,10,11,12,13,14,15,16,17,18]. The amplitude of these abiotic stresses has increased severely in recent years principally due to anthropogenic activities [7,19]. Plants, being sessile, are persistently exposed to these factors and require a set of effective mechanisms which can be activated under unfavorable circumstances to sustain their life cycle [20]. According to some reports, the projection of these stresses contributes significantly and affects the growth and productivity by reducing crop yield and overall crop production by 70% and 50%, respectively [21,22]. Thus, it is imperative to reduce the crop productivity losses by improving crop performance through various approaches, including application of plant bio-stimulant products as well as stimulation of plant secondary metabolism [11,23,24]. Plants need to endure different abiotic stresses and polyphenols accumulate in response to these stresses helping plants to acclimatize to unfavorable environments [25,26]. Hence, the concentration of phenols in plant tissue is a good indicator to predict the extent of abiotic stress tolerance in plants which varies greatly in different plant species under an array of external factors.
Phenolic compounds influence the plant growth and development, including seed germination, biomass accumulation, and improved plant metabolism [27,28,29,30]. In this regard, we summarized different studies showing a broad spectrum of different effects of abiotic stresses and discussed how endogenous phenol levels can help in mitigating abiotic stress in plants. Moreover, physiological and molecular mechanisms connected to the phenylpropanoid pathway underlying abiotic stress tolerance have extensively discussed. At the end, we explained phenol-mediated stress tolerance and suggestions have been made to further escalate the extent of deep mechanistic studies.
2. Biosynthetic Pathway of Polyphenols in Plants
Phenolics are known to be the largest groups of secondary metabolites in plants varying from simpler aromatic rings to more complex ones, such as lignins. These compounds originated from phenylalanine therefore are also called as phenylpropanoids. Polyphenols are characterized by the presence of large multiples of phenol structural units. The number and characteristics of these phenol structures underlie the unique physical, chemical, and biological properties of particular members of each class. Phenols are indeed divided into several groups such as phenolic acids, flavonoids, stilbenes, and lignans with peculiar properties. Plant phenolics are synthesized biogenetically through a shikimate/phenylpropanoid pathway, whereas a mevalonate pathway generates terpenoids. Both these secondary pathways produce a wide array of monomeric and polymeric structures encompassing a comprehensive array of physiological and biochemical roles in plants. The term “secondary metabolites” refers to the metabolites or phytochemicals synthesized through secondary metabolism. During the biosynthesis of phenolic compounds, erythrose 4-phosphate is combined with phosphoenolpyruvate (PEP) to form phenylalanine. Then phenylalanine ammonia lyase (PAL) catalyzes the conversion of phenylalanine to trans-cinnamic acid. Several other phenolic compound such as flavonoids, coumarins, hydrolysable tannins, monolignols, lignans, and lignins are formed through this pathway, formally known as the phenylpropanoid pathway (see complete details in [26,31,32,33]).
3. Physiological Roles of Phenolics in Plants
Phenolics are widely distributed and are involved in key metabolic and physiological process in plants [34,35]. Phenolics influence different physiological processes related to growth and development in plants including seed germination, cell division, and synthesis of photosynthetic pigments [36]. Phenolic compounds have been exploited for several application including bioremediation, allelochemical, promotion of plant growth, and antioxidants as food additives [37]. In plants, phenolic accumulation is usually a consistent feature of plants under stress, which represents a defensive mechanism to cope with multiple abiotic stresses [31]. Plant phenolics play an important role in several physiological processes to improve the tolerance and adaptability of plants under suboptimal conditions [38,39,40]. In particular, a large number of secondary metabolites having antioxidant properties belong to this group [41] which can ameliorate plant performance under stress conditions.
Plants interact with their living environment through secondary metabolites. Polyphenols are, for example, involved in signal transduction from the root to the shoot and also help in nutrient mobilization. The roots exudates contain phenolic compounds which alter the physiochemical properties of the rhizosphere. Soil microbes transform phenolics into compounds which help in N mineralization and humus formation [42]. Furthermore, phenolics improve nutrient uptake through chelation of metallic ions, enhanced active absorption sites, and soil porosity with accelerated mobilization of elements like calcium (Ca), magnesium (Mg), potassium (K), zinc (Zn), iron (Fe), and manganese (Mn) [43]. Recently, Rehman, et al. [44] found that Zn application and plant growth promoting rhizobacteria (PGPRs) treatment enhanced the contents of phenolics and organic acids (pyruvic acid, tartaric acid, citric acid, malonic acid, malic acid, succinic acid, oxaloacetic acid, oxalic acid, and methyl malonic acid) in root exudates of wheat, which helped in nutrient mobilization of Zn, N, and Ca and their uptake [44,45]. Phenolic compounds also help in N fixation in legumes. Legumes release several secondary metabolites from roots, principally flavonoid compounds (flavanols and iso-flavonoids) which play crucial role in Nod factors synthesis and in the production of infection tube during nodulation, given that they inhibits auxin transport and facilitate cell division [46].
Plant phenolics, as physiological regulator or chemical messenger, inhibit the IAA catabolism (dihydroxy B-ring flavonoids) or limit the IAA synthesis (monohydroxy B-ring flavonoids) [47]. Flavonoids play a key role in the development of functional pollen. For instance, addition of a very small dose of flavonol aglycones kaempferol or quercetin restored the fertility in mature pollen during pollination [48,49]. Some phenolic compounds (trans-cinnamic acid, coumarin, p-hydroxybenzoic acid, and benzoic acid) might be potentially phytotoxic if accumulated in high quantity and can inhibit germination and seedling growth [50] due to the disruption of cellular enzyme functioning and impairment of cell division. For instance, some phenolic compounds inhibit the prolyl aminopeptidase and phosphatase enzyme involved in seed germination in legumes [51]. Conversely, high phenolic acid contents have been reported to exert positive effects in seed germinating. In a recent study, Chen et al. [52] found a substantial increase in free (1042%), bound (120%), and total phenolic acid content (741%) in canary grass during germination. The spruce bark extract containing polyphenols stimulated the germination rate in Lycopersicon esculentum while inhibited root elongation [53]. Phenolics reduced the thickness and increased the seed tegument porosity which help in water imbibition and boost the germination rate [54]. Polyphenolic extracts of spruce bark intensified the photosynthetic activity and biosynthesis of assimilatory pigment (chlorophyll a and b) in maize and sunflower [55,56]. Phenolics reduced the energy required for ion transfer by modifying the structure of thylakoids and mitochondrial membranes [57]. As antioxidants, phenolic compounds participate in the scavenging of reactive oxygen species (ROS), catalyzing oxygenation reactions through formation of metallic complexes, and inhibiting the activities of oxidizing enzymes [58].
In conclusion, polyphenols are produced under optimal and (with higher levels) in suboptimal conditions in plants and play crucial role in the development encompassing signal transduction, cell division, hormonal regulation, photosynthetic activity regulation, germination, and reproduction rate. Plants exhibiting increased synthesis of polyphenols under abiotic stresses usually show a better adaptability to limiting environments.
4. Abiotic Stresses and Their Toxic Effects on Plants
In recent times, producing more food and preventing crop losses to meet the demands of ever-increasing human populations has gained unprecedented importance. Nevertheless, a large proportion of arable land face abiotic stresses (drought, salinity, cold, heat, heavy metal toxicity, UV radiation, etc.) which are expected to increase due to climate change and the incidence of these environmental stresses are further fueled by anthropogenic activities. These abiotic stresses cause alteration in physiological and biochemical processes of plants which results in diminished plant growth and poor yield [59]. These stresses bring rapid changes in cellular redox homeostasis with excessive reactive oxygen species (ROS) generation which eventually damage cell organelles and interfere in ROS-promoted signaling pathways [60]. Contrary to over production of ROS, a physiological redox state hampers normal cell functions and affects the plant immune system, suggesting that a threshold level of ROS is necessary for normal plant functioning (Figure 1; Farooq, et al. [61]). Increased ROS generation under abiotic stresses enhanced itself exponentially the production of ROS [62], which result in peroxidation and destabilization of cellular membranes. Recently Rehman, et al. [63] observed that heat stress and Zn deficiency cause reductions in growth (shoot and root biomass, and root length), and consequently impeded nutrient uptake, enhanced lipid peroxidation and impaired photosynthetic performance. In plants, ROS is produced from 1–2% of total O2 consumed in high active cell organelles like chloroplasts, mitochondria, and peroxisomes (Figure 1; [64]). The most common ROS are singlet oxygen (1O2), superoxide radicle (O2•–), hydrogen peroxide (H2O2), and hydroxyl radicle (•OH) [65].
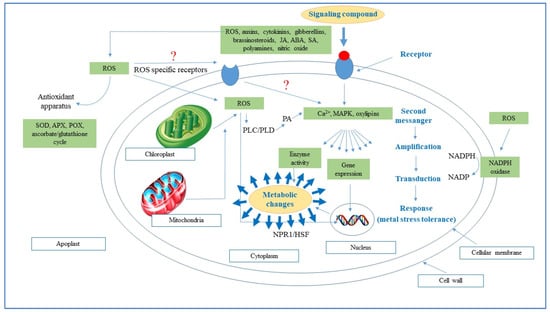
Figure 1.
Schematization of signal transmission and transduction in plant cells. Abbreviation: ABA, abscisic acid; APX, ascorbate peroxidase; HSF, redox-sensitive transcription factor; JA, jasmonic acid; MAPK, mitogen-activated protein kinase; NADP, oxidized nicotinamide adenine dinucleotide; NADPH, reduced nicotinamide adenine dinucleotide; NPR1, redox-sensitive transcription factor; OXI1, serine/threonine kinase; PA, phosphatidic acid; PLC/PLD, phospholipases class C and D; POX, peroxidase; ROS, reactive oxygen species; SA, salicylic acid; SOD, superoxide dismutase.
Abiotic stresses disturb the balance between ROS generation and scavenge and accelerate ROS propagation which damages vital macromolecules (nucleic acids, proteins, carbohydrates, and lipids) and eventually leads to cell death. ROS-induced protein damage is caused by oxidation of amino acid residues (e.g., cysteine) for disulphide bond formation, oxidation of arginine, lysine, and threonine residues resulting in irreversible carbonylation in side chains and oxidation of methionine residue to form methionine sulphoxide [66]. ROS production also limits CO2 fixation in chloroplasts which are the main site for ROS generation in green plants [67]. ROS reacts with chlorophyll during photosynthesis and forms the chlorophyll triplet state which can rapidly generate (1O2), thus causing damage to photosynthetic complexes (principally PSII) and perturbing the molecular reaction of the photosynthetic pathway [68]. Apart from the chloroplast, the mitochondria also increase ROS production under abiotic stress which influences plant cellular processes [24]. In mitochondria, ~1–5% of O2 consumed leads to H2O2 formation which is subsequently transformed in •OH during the Fenton reaction [69]. Furthermore, intensive respiratory/photorespiratory metabolism demands high electron input leading to escalated ROS production which results in protein oxidation [61]. Peroxisomes are also major sites for ROS production, particularly H2O2, and have two- and 50-fold higher concentration of H2O2 than chloroplasts and mitochondria, respectively [70]. This H2O2 is involved in stress induced oxidative damage given that it can freely pass lipid membranes. Under the physiological level, different antioxidant defense mechanism detoxify ROS. However, over production of ROS can overwhelm the defense system, resulting in oxidative stress, cell damage, and cell death (Figure 1) [71].
5. Response and Role of Endogenous Phenolics in Plants against Abiotic Stress
In response to abiotic stresses, biosynthesis of secondary metabolites, including polyphenols, is usually increased in plants. Phenolics confer indeed higher tolerance to plants against various stress conditions like heavy metals, salinity, drought, temperature, pesticides, and UV radiations [33,72,73,74,75,76,77]. Plants growing under stressful environments have the ability to biosynthesize more phenolic compounds in comparison to plants growing under normal conditions [78]. These compounds have antioxidative properties and are capable of scavenging free radicals, resulting in reduction of cell membrane peroxidation [79], hence protecting plant cells from ill effects of oxidative stress. Biosynthesis of phenolics under stressful environments is regulated by the altered activities of various key enzymes of phenolic biosynthetic pathways like PAL and CHS (chalcone synthase). Enhanced performance of enzymes is also accompanied by the up-regulation of the transcript levels of genes encoding key biosynthetic enzymes like PAL, C4H (cinnamate 4-hydroxylase), 4CL (4-coumarate: CoA ligase), CHS, CHI (chalcone isomerase), F3H (flavanone3-hydroxylase), F3′H (flavonoid 3′-hydroxylase), F3′5′H (flavonoid 3′5′-hydroxylase), DFR (dihydroflavonol 4-reductase), FLS (flavonol synthase), IFS (isoflavone synthase), IFR (isoflavone reductase), and UFGT (UDP flavonoid glycosyltransferase) [74,80,81,82,83,84,85,86]. The responses of phenolic compounds under different abiotic stresses have been discussed in individual sections mentioned below.
5.1. Heavy Metal
Metal stress causes oxidative stress to plants by triggering the generation of harmful ROSs and ultimately cause toxicity and retarded growth [11,87,88]. However, enhanced biosynthesis of phenolics in plants under metal stress helps in protecting plants from oxidative stress [72,89,90]. Flavonoids can enhance the metal chelation process which helps in reducing the levels of harmful hydroxyl radical in plant cells [91,92] and this fits well with the observation that the levels of flavonoids in plants have found to be enhanced by metal excess [90,93]. Under metal toxicity, accumulation of specific flavonoids which are involved in aiding to the plant’s defense mechanism is also enhanced including anthocyanins and flavonols [72,94,95,96]. Accumulation of phenolic compounds is due to the up-regulation of the biosynthesis of phenylpropanoid enzymes including phenylalanine ammonia-lyase, chalcone synthase, shikimate dehydrogenase, cinnamyl alcohol dehydrogenase, and polyphenol oxidase [95,97], which in turn, is dependent on the modulation of transcript levels of genes encoding biosynthetic enzymes under metal stress [72,85]. Flavonoids are also known for their scavenging capability of H2O2 and are considered to play a crucial role in the phenolic/ascorbate-peroxidase cycle [98,99].
Shikimate dehydrogenase (SKDH) and glucose-6-phosphate dehydrogenase (G6PDH) are two important enzymes which catalyze the biological reaction required for the production of important precursors of phenylpropanoid pathways [100]. Another enzyme cinnamyl alcohol dehydrogenase (CADH) catalyzes biochemical reactions which produce precursors required for synthesis of lignin [101]. Heavy metals stimulate phenylpropanoid the biosynthetic pathway in plants by up-regulating the activities of key biosynthetic enzymes like PAL, SKDH, G6PDH, and CADH [101]. Additionally, polyphenol oxidase (PPO) helps during the process of ROS scavenging, and enhancing a plant’s resistance to abiotic stress conditions like heavy metals [100,101,102]. Table 1 summarizes the impact of metal stress on phenolic composition of plants.

Table 1.
Summary table describing the impact of heavy metal stress on the endogenous levels of various phenolic compounds in plants.
5.2. Drought
Phenolic accumulation is very crucial to counteract the negative impacts of drought stress in plants [33]. Transcriptomic and metabolomic studies carried out on Arabidopsis plants confirmed that enhanced flavonoid accumulation under drought stress is very helpful to provide resistance [107]. Biosynthesis and accumulation of flavonols were also stimulated in plants under water deficit conditions accompanied by enhanced resistance against drought stress [108,109]. Drought stress also regulated the biosynthetic pathways of phenolic acids and flavonoids, leading to enhanced accumulation of these compounds [82,110,111] which acted as antioxidants and prevented plants from adverse effects of water deficit conditions [112]. For example, contents of flavonoids like kaempferol and quercetin were enhanced in tomato plants accompanied by enhanced drought tolerance [113]. Flavonoid accumulation in cytoplasm can efficiently detoxify harmful H2O2 molecules generated as a result of drought stress and, at the end oxidation of flavonoids is followed by ascorbic acid mediated re-conversion of flavonoids into primary metabolites [114]. The main reason for this drought-induced accumulation of phenolic compounds is the modulation of phenylpropanoid biosynthetic pathway. Drought regulates many key genes encoding main enzymes of phenylpropanoid pathway, which results in stimulated biosynthesis of phenolic compounds. The impact of drought stress on accumulation of phenolics and related processes has been summarized in Table 2.
Table 2.
Summary table describing the impact of drought stress on the endogenous levels of various phenolic compounds in plants.
5.3. Salinity
Salt stress results in generation of ROS like superoxide anions, hydrogen peroxide, and hydroxyl ions [126,127] and require activation of well-orchestrated and finely-tuned plants antioxidant system to contrast ROS propagation [128,129]. Phenolic compounds have powerful antioxidant properties and help in scavenging of harmful ROS in plants under salt stress [130,131,132]. Moreover, in response to salt stress, phenylpropanoid biosynthetic pathway gets stimulated and results in production of various phenolic compounds which have strong antioxidative potential [131,133,134].
Some genes like VvbHLH1 are involved in the enhanced production of flavonoids by regulating the genes of the biosynthetic pathways and confer salt tolerance to plants [135,136]. In tobacco plants, NtCHS1 plays a crucial role in the biosynthesis of flavonoids under salt stress, where accumulation directly favors the scavenging of ROS [130]. Flavone biosynthesis also was enhanced under saline conditions and in Glycine max, it was observed that salinity up-regulates the expression of flavone synthase genes, GmFNSII-1 and GmFNSII-2 [137]. Some phenolic acids also accumulate in plants under saline conditions including caffeic acid, caftaric acid, cinnamylmalic acid, gallic acid, ferulic acid, and vanillic acid [131,138,139,140]. Biosynthesis of anthocyanins also was promoted in plants growing under saline conditions [141,142]. A detailed explanation about the effect of salt stress on phenolic composition has been provided in Table 3.
Table 3.
Summary table describing the impact of salt stress on the endogenous levels of various phenolic compounds in plants.
5.4. UV Light
Exposure of UV-B radiations to plants causes damage to their protein structure, causes harmful mutations to DNA and generates harmful ROS. To counteract the negative effects of UV-B exposure, endogenous phenolics accumulated in plant cells and protect cell components by making a shield under epidermal layer. They further reduce DNA damage by preventing dimerization of thymine along with reducing photo-damage of important enzymes like NAD/NADP [33,148]. Moreover, flavonoids also act as light screens due to their capability of absorbing both visible (anthocyanins) and UV radiations (anthocyanins and colorless flavonoids), hence protecting plants from these harmful radiations [26,149]. This fact was supported by various researchers who observed enhanced biosynthesis of flavonoids in plants under UV radiations, accompanied by enhanced UV absorption and plant tolerance to these radiations [98,150] and powerful antioxidant capacity [151]. Moreover, it is also well known that plants growing at high altitude accumulate more phenolics like flavonoids than plants of a temperate region. This enhanced flavonoid accumulation under high light/UV exposure is because of stimulated flavonoid biosynthetic pathways and their corresponding gene transcript levels [33,83,152,153]. The key genes which are up-regulated in plants upon UV exposure include: CHS (chalcone synthase); CHI (chalcone isomerase); FLS (flavonol synthase); DFR (dihydroflavonol 4-reductase); FHT (flavanone 3β-hydroxylase), FGT (flavonoid glycosyltransferases); and PAL (phenylalanine ammonia lyase) [154,155]. It is also believed that UV light also utilizes jasmonate dependent/independent pathways to stimulate the biosynthesis of phenols in plants [156]. Additionally, abscisic acid (ABA) is also known to modulate the phenolic biosynthetic pathway in presence of UV light [157]. Table 4 provides a brief summary about impact of UV exposure on the endogenous phenolic composition of plants.
Table 4.
Summary table describing the impact of UV light exposure on the endogenous levels of various phenolic compounds in plants.
5.5. Other Abiotic Factors
Other abiotic factors like temperature, nanoparticles, and pesticides also stimulate the endogenous phenolic biosynthesis in plants and help in providing resistance against phytotoxic effects of these abiotic stresses [74,80,153,168,169,170,171]. Phenolic biosynthetic pathways also get activated in plants growing under pesticide stress conditions. This leads to more accumulation of phenolic compounds in plants, which confer resistance to survive against pesticide toxicity [73,170]. This stimulated phenolic biosynthesis is due to the activation of key biosynthetic enzymes and up-regulation of key genes of phenylpropanoid branch, including PAL and CHS [74,80]. Increased accumulation of anthocyanins in plant leaves promote by application of insecticides also helps in recovery of plant photosynthetic efficiency [172]. Similarly, under temperature stress (both heat and chilling), plants synthesize more phenolic compounds such as anthocyanins, flavonoids, flavonols, and phenolic acids, which ultimately protect plant cells [75,129,168,169,173]. In Festuca trachyphylla plants growing under heat stress, enhancement in the phenolic compounds was noticed including 4-hydroxybenzoic acid, benzoic acid, caffeic acid, coumaric acid, cinnamic acid, gallic acid, homovanillic acid, ferulic acid, salicylic acid, and vanillic acid [76]. The increased accumulation of these phenolic compounds is accompanied by enhanced tolerance of F. trachyphylla plants against high temperature [76]. In carrot, phenolics like coumaric acid, caffeic acid, and anthocyanins are suggested to prevent heat induced oxidative damage by enhancing their accumulation [174]. Some phenolics like salicylic acid also act as stimulant for phenol biosynthesis in plants under high temperature stress. This leads to enhanced accumulation of phenolic compounds which further help in detoxification of ROS and providing heat resistance to plants [175]. Under chilling stress, phenolic compounds like suberin or lignin start accumulating in plant cell walls which helps in enhancing resistance against chilling stress [176]. This enhanced thickness of cell wall due to phenolic accumulation is beneficial for prevention of chilling injury and cell collapse under cold stress [33]. Stimulated phenolic biosynthesis under low temperature stress is due to the enhanced expression of PAL, CAD (cinnamylalcohol dehydrogenase), and HCT (hydroxycinnamoyl transferase), and increased phenolic levels play crucial role in protection plants against chilling stress [86]. This fact is further supported by the research carried out on peach under chilling stress by Gao et al. [177]. These researchers suggested that 24-epibrassinolide stimulated biosynthesis of phenolics is involved in reduction of heat generated oxidative stress by helping to scavenge of ROS. Table 5 provides a detailed overview about how different abiotic factors affect phenolic metabolism in plants.
Table 5.
Summary table describing the impact of various abiotic factors on the endogenous levels of various phenolic compounds in plants.
6. Conclusions
Phenylpropanoid pathway is likely the most studied pathway of secondary metabolism in planta. In plants growing under challenging environments, accumulation of phenolic compounds usually parallels enhanced plant tolerance as summarized in Figure 2. Abiotic stresses also activate the cell signaling process, resulting in transcriptional up-regulation of phenylpropanoid pathway. The increase in plant’s resistance is correlated with the multiple function of polyphenols in plants, principally consisting in their ROS scavenging ability and/or the capacity of some polyphenol classes to protect the plant from excessive light such as UV (flavonoids) and visible light (anthocyanins). In addition, polyphenols might play other key ecological roles under abiotic stress, acting for example as infochemicals for other plants. Aside from the huge body of papers on the matter, further research is needed to deepen, for example, the role of specialized polyphenols as a response to certain abiotic stresses and to describe the intimal mechanisms which shift from primary metabolism to the up-regulation of phenylpropanoid pathway, which is as a cross response to several environmental stressors.
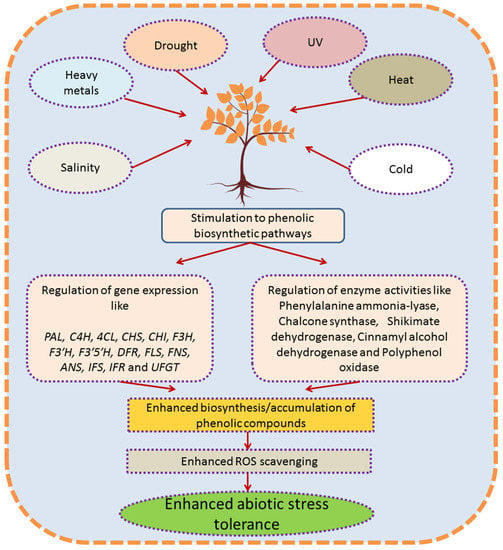
Figure 2.
Diagrammatic explanation for response and role of phenolic compounds in plants growing under abiotic stress conditions. ROS (reactive oxygen species); PAL (phenylalanine ammonia lyase); CHS (chalcone synthase); CHI (chalcone isomerase); C4H (cinnamate 4-hydroxylase); 4CL (4-coumarate: CoA ligase); F3H (flavanone3-hydroxylase); F3′H (flavonoid 3′-hydroxylase); F3′5′H (flavonoid 3′5′-hydroxylase); FLS (flavonol synthase); FNS (flavone synthase) UFGT (UDP flavonoid glycosyltransferase); IFS (isoflavone synthase); IFR (isoflavone reductase); DFR (dihydroflavonol 4-reductase); ANS (anthocyanidin synthase).
Author Contributions
A.S. and B.Z. drafted the outline of review. A.S., B.S., and A.R. participated in literature collection and writing the initial draft. R.B. and M.L. participated in revision of initial draft.
Funding
We acknowledge the different funding agencies: This study was supported by National Key Research and Development Program of China (2018YFD1000600); Independent research topics of the State Key Laboratory of Subtropical Silviculture (ZY20180208, ZY20180308); Key Research and Development Program of Zhejiang Province (2018C02004); Key Project of Zhejiang Provincial Natural Science Foundation (LZ18C160001); Fruit Innovation Team Project of Zhejiang Province (2016C02052-12); National Undergraduate Innovation and Entrepreneurship Training Project (201610341010); Undergraduate Science and Technology Innovation Plan of Zhejiang Province (2017R412006); Undergraduate Research Training Program in Zhejiang A & F University (102-2013200005, 102-2013200041, 102-2013200042,KX20180047, KX20180043, KX20180065).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Plant Sci. 2018, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Kreps, J.A.; Wu, Y.; Chang, H.-S.; Zhu, T.; Wang, X.; Harper, J.F. Transcriptome changes for Arabidopsis in response to salt, osmotic, and cold stress. Plant Physiol. 2002, 130, 2129–2141. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.-B.; Guo, Q.-J.; Chu, L.-Y.; Zhao, X.-N.; Su, Z.-L.; Hu, Y.-C.; Cheng, J.-F. Understanding molecular mechanism of higher plant plasticity under abiotic stress. Colloids Surf. B Biointerfaces 2007, 54, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Gill, S.S.; Gill, R. Plant Adaptation to Environmental Change: Significance of Amino Acids and Their Derivatives; CABI: Wallingford, UK, 2014. [Google Scholar]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Bao, M.; Wang, L.; Khan, I.; Ullah, E.; Tung, S.A.; Samad, R.A.; Shahzad, B. Cadmium toxicity in Maize (Zea mays L.): Consequences on antioxidative systems, reactive oxygen species and cadmium accumulation. Environ. Sci. Pollut. Res. 2015, 22, 17022–17030. [Google Scholar] [CrossRef]
- Shahzad, B.; Tanveer, M.; Che, Z.; Rehman, A.; Cheema, S.A.; Sharma, A.; Song, H.; Ur Rehman, S.; Zhaorong, D. Role of 24-epibrassinolide (EBL) in mediating heavy metal and pesticide induced oxidative stress in plants: A review. Ecotoxicol. Environ. Saf. 2018, 147, 935–944. [Google Scholar] [CrossRef]
- Shahzad, B.; Tanveer, M.; Rehman, A.; Cheema, S.A.; Fahad, S.; Rehman, S.; Sharma, A. Nickel; whether toxic or essential for plants and environment-A review. Plant Physiol. Biochem. 2018, 132, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kumar, V.; Kumar, R.; Shahzad, B.; Thukral, A.K.; Bhardwaj, R. Brassinosteroid-mediated pesticide detoxification in plants: A mini-review. Cogent Food Agric. 2018, 4, 1436212. [Google Scholar] [CrossRef]
- Fahad, S.; Rehman, A.; Shahzad, B.; Tanveer, M.; Saud, S.; Kamran, M.; Ihtisham, M.; Khan, S.U.; Turan, V.; Ur Rahman, M.H. Rice Responses and Tolerance to Metal/Metalloid Toxicity. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 299–312. [Google Scholar]
- Soares, C.; Carvalho, M.E.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges—A little help from the antioxidant networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Guo, H.; Feng, X.; Hong, C.; Chen, H.; Zeng, F.; Zheng, B.; Jiang, D. Malate secretion from the root system is an important reason for higher resistance of Miscanthus sacchariflorus to cadmium. Physiol. Planta 2017, 159, 340–353. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Chen, H.; Hong, C.; Jiang, D.; Zheng, B. Exogenous malic acid alleviates cadmium toxicity in Miscanthus sacchariflorus through enhancing photosynthetic capacity and restraining ROS accumulation. Ecotoxicol. Environ. Saf. 2017, 141, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Qiu, L.; Guo, H.; Wang, Y.; Yuan, H.; Yan, D.; Zheng, B. Spermidine induces physiological and biochemical changes in southern highbush blueberry under drought stress. Braz. J. Bot. 2017, 40, 841–851. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Khan, N.A. Salicylic acid and jasmonates: Approaches in abiotic stress tolerance. J. Plant Biochem. Physiol. 2013, 1, 4. [Google Scholar] [CrossRef]
- Rao, K.M.; Raghavendra, A.; Reddy, K.J. Physiology and Molecular Biology of Stress Tolerance in Plants; Springer Science & Business Media: New York, NY, USA, 2006. [Google Scholar]
- Kaur, G.; Kumar, S.; Nayyar, H.; Upadhyaya, H. Cold stress injury during the pod-filling phase in chickpea (Cicer arietinum L.): Effects on quantitative and qualitative components of seeds. J. Agron. Crop Sci. 2008, 194, 457–464. [Google Scholar]
- Mantri, N.; Patade, V.; Penna, S.; Ford, R.; Pang, E. Abiotic stress responses in plants: Present and future. In Abiotic Stress Responses in Plants; Springer: New York, NY, USA, 2012; pp. 1–19. [Google Scholar]
- Shahzad, B.; Cheema, S.; Farooq, M.; Cheema, Z.; Rehman, A.; Abbas, T. Growth Stimulating Influence of Foliage Applied Brassica Water Extracts on Morphological and Yield Attributes of Bread Wheat under Different Fertilizer Regimes. Planta Daninha 2018, 36, e018178331. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 26. [Google Scholar] [CrossRef]
- Pereira, A. Plant abiotic stress challenges from the changing environment. Front. Plant Sci. 2016, 7, 1123. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V. Phenolic Compounds: Introduction. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1543–1580. [Google Scholar] [CrossRef]
- Raskin, I. Role of salicylic acid in plants. Ann. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Yalpani, N.; Enyedi, A.J.; León, J.; Raskin, I. Ultraviolet light and ozone stimulate accumulation of salicylic acid, pathogenesis-related proteins and virus resistance in tobacco. Planta 1994, 193, 372–376. [Google Scholar] [CrossRef]
- Senaratna, T.; Touchell, D.; Bunn, E.; Dixon, K. Acetyl salicylic acid (Aspirin) and salicylic acid induce multiple stress tolerance in bean and tomato plants. Plant Growth Regul. 2000, 30, 157–161. [Google Scholar] [CrossRef]
- Nazar, R.; Iqbal, N.; Syeed, S.; Khan, N.A. Salicylic acid alleviates decreases in photosynthesis under salt stress by enhancing nitrogen and sulfur assimilation and antioxidant metabolism differentially in two mungbean cultivars. J. Plant Physiol. 2011, 168, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Saltveit, M.E. Synthesis and metabolism of phenolic compounds. In Fruit and Vegetable Phytochemicals Chemistry, Nutritional Value, and Stability; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and Regulation of Plants Phenolics in Abiotic Stress Tolerance: An Overview. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 157–168. [Google Scholar]
- Boudet, A.M. Evolution and current status of research in phenolic compounds. Phytochemistry 2007, 68, 2722–2735. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A.; Kohli, S.K.; Bali, S.; Sharma, M.; Kumar, R.; Bhardwaj, R.; Thukral, A.K. Differential distribution of polyphenols in plants using multivariate techniques. Biotech. Res. Innov. 2019, 3, 1–21. [Google Scholar] [CrossRef]
- Tanase, C.; Bujor, O.-C.; Popa, V.I. Phenolic Natural Compounds and Their Influence on Physiological Processes in Plants. In Polyphenols in Plants, 2nd ed.; Watson, R.R., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 45–58. [Google Scholar] [CrossRef]
- Bujor, O.-C.; Talmaciu, I.A.; Volf, I.; Popa, V.I. Biorefining to recover aromatic compounds with biological properties. TAPPI J. 2015, 14, 187–193. [Google Scholar]
- Andersen, C.P. Source–sink balance and carbon allocation below ground in plants exposed to ozone. New Phytol. 2003, 157, 213–228. [Google Scholar] [CrossRef]
- Lattanzio, V.; Cardinali, A.; Ruta, C.; Fortunato, I.M.; Lattanzio, V.M.T.; Linsalata, V.; Cicco, N. Relationship of secondary metabolism to growth in oregano (Origanum vulgare L.) shoot cultures under nutritional stress. Environ. Exp. Bot. 2009, 65, 54–62. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Oszmanski, J. Polyphenols as antioxidants in food. Przem. Spo. 1995, 3, 94–96. [Google Scholar]
- Halvorson, J.J.; Gonzalez, J.M.; Hagerman, A.E.; Smith, J.L. Sorption of tannin and related phenolic compounds and effects on soluble-N in soil. Soil Biol. Biochem. 2009, 41, 2002–2010. [Google Scholar] [CrossRef]
- Seneviratne, G.; Jayasinghearachchi, H.S. Mycelial colonization by bradyrhizobia and azorhizobia. J. Biosci. 2003, 28, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Farooq, M.; Naveed, M.; Nawaz, A.; Shahzad, B. Seed priming of Zn with endophytic bacteria improves the productivity and grain biofortification of bread wheat. Eur. J. Agron. 2018, 94, 98–107. [Google Scholar] [CrossRef]
- Rehman, A.; Farooq, M.; Naveed, M.; Ozturk, L.; Nawaz, A. Pseudomonas aided zinc application improves the productivity and biofortification of bread wheat. Crop Pasture Sci. 2018, 69, 659–672. [Google Scholar] [CrossRef]
- Zhang, J.; Subramanian, S.; Stacey, G.; Yu, O. Flavones and flavonols play distinct critical roles during nodulation of Medicago truncatula by Sinorhizobium meliloti. Plant J. 2009, 57, 171–183. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoids induced in cells undergoing nodule organogenesis in white clover are regulators of auxin breakdown by peroxidase. J. Exp. Bot. 2001, 52, 419–426. [Google Scholar] [CrossRef]
- Van Der Meer, I.M.; Stam, M.E.; Van Tunen, A.J.; Mol, J.N.; Stuitje, A.R. Antisense inhibition of flavonoid biosynthesis in petunia anthers results in male sterility. Plant Cell 1992, 4, 253–262. [Google Scholar] [CrossRef]
- Taylor, L.P.; Grotewold, E. Flavonoids as developmental regulators. Curr. Opin. Plant Biol. 2005, 8, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Baleroni, C.R.S.; Ferrarese, M.L.L.; Souza, N.E.; Ferrarese-Filho, O. Lipid Accumulation during Canola Seed Germination in Response to Cinnamic Acid Derivatives. Biol. Planta 2000, 43, 313–316. [Google Scholar] [CrossRef]
- Shankar, S.; Girish, R.; Karthik, N.; Rajendran, R.; Mahendran, V. Allelopathic effects of phenolics and terpenoids extracted from Gimelina arborea on germination of Black gram (Vigna mungo) and Green gram (Vigna radiata). Allelopath. J. 2009, 23, 323–332. [Google Scholar]
- Chen, Z.; Yu, L.; Wang, X.; Gu, Z.; Beta, T. Changes of phenolic profiles and antioxidant activity in canaryseed (Phalaris canariensis L.) during germination. Food Chem. 2016, 194, 608–618. [Google Scholar] [CrossRef] [PubMed]
- Balas, A.; Popa, V. Bioactive compounds extracted from Picea abies bark. In Proceedings of the 10th European Workshop on Lignocellulosics and Pulp, Stockholm, Sweden, 25–28 August 2008; pp. 345–356. [Google Scholar]
- Tobe, K.; Zhang, L.; Qiu, G.Y.; Shimizu, H.; Omasa, K. Characteristics of seed germination in five non-halophytic Chinese desert shrub species. J. Arid. Environ. 2001, 47, 191–201. [Google Scholar] [CrossRef]
- Tanase, C.; Boz, I.; Stingu, A.; Volf, I.; Popa, V.I. Physiological and biochemical responses induced by spruce bark aqueous extract and deuterium depleted water with synergistic action in sunflower (Helianthus annuus L.) plants. Ind. Crops Prod. 2014, 60, 160–167. [Google Scholar] [CrossRef]
- Tanase, C.; Bara, C.I.; Popa, V.I. Cytogenetical effect of some polyphenol compounds separated from industrial by-products on. Cell. Chem. Technol. 2015, 49, 799–805. [Google Scholar]
- Moreland, D.E.; Novitzky, W.P. Effects of Phenolic Acids, Coumarins, and Flavonoids on Isolated Chloroplasts and Mitochondria. In Allelochemicals: Role in Agriculture and Forestry; American Chemical Society: Washington, DC, USA, 1987; pp. 247–261. [Google Scholar] [CrossRef]
- Amarowicz, R.; Weidner, S. Biological activity of grapevine phenolic compounds. In Grapevine Molecular Physiology & Biotechnology; Springer: New York, NY, USA, 2009; pp. 389–405. [Google Scholar]
- Wani, S.H.; Sah, S. Biotechnology and abiotic stress tolerance in rice. J. Rice Res. 2014, 2, e105. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.-P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef]
- Farooq, M.A.; Niazi, A.K.; Akhtar, J.; Farooq, M.; Souri, Z.; Karimi, N.; Rengel, Z. Acquiring control: The evolution of ROS-Induced oxidative stress and redox signaling pathways in plant stress responses. Plant Physiol. Biochem. 2019, 141, 353–369. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Farooq, M.; Asif, M.; Ozturk, L. Supra-optimal growth temperature exacerbates adverse effects of low Zn supply in wheat. J. Plant Nutr. Soil Sci. 2019. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; Palma, J.M.; Rodriguez-Ruiz, M. Plant peroxisomes: A nitro-oxidative cocktail. Redox Biol. 2017, 11, 535–542. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, B.K.; Askerlund, P.; Bykova, N.V.; Egsgaard, H.; Moller, I.M. Identification of oxidised proteins in the matrix of rice leaf mitochondria by immunoprecipitation and two-dimensional liquid chromatography-tandem mass spectrometry. Phytochemistry 2004, 65, 1839–1851. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Buchert, F.; Forreiter, C. Singlet oxygen inhibits ATPase and proton translocation activity of the thylakoid ATP synthase CF1CFo. FEBS Lett. 2010, 584, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Popov, V.N.; Simonian, R.A.; Skulachev, V.P.; Starkov, A.A. Inhibition of the alternative oxidase stimulates H2O2 production in plant mitochondria. FEBS Lett. 1997, 415, 87–90. [Google Scholar] [CrossRef]
- Foyer, C.H.; Bloom, A.J.; Queval, G.; Noctor, G. Photorespiratory metabolism: Genes, mutants, energetics, and redox signaling. Annu. Rev. Plant Biol. 2009, 60, 455–484. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol. 2006, 141, 312–322. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Thukral, A.K.; Bhardwaj, R. Epibrassinolide-imidacloprid interaction enhances non-enzymatic antioxidants in Brassica juncea L. Ind. J. Plant Physiol. 2016, 21, 70–75. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, S.; Kumar, V.; Kanwar, M.K.; Kesavan, A.K.; Thukral, A.K.; Bhardwaj, R.; Alam, P.; Ahmad, P. Pre-sowing Seed Treatment with 24-Epibrassinolide Ameliorates Pesticide Stress in Brassica juncea L. through the Modulation of Stress Markers. Front. Plant Sci. 2016, 7, 1569. [Google Scholar] [CrossRef] [PubMed]
- Ancillotti, C.; Bogani, P.; Biricolti, S.; Calistri, E.; Checchini, L.; Ciofi, L.; Gonnelli, C.; Del Bubba, M. Changes in polyphenol and sugar concentrations in wild type and genetically modified Nicotiana langsdorffii Weinmann in response to water and heat stress. Plant Physiol. Biochem. 2015, 97, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yuan, B.; Huang, B. Differential Heat-Induced Changes in Phenolic Acids Associated with Genotypic Variations in Heat Tolerance for Hard Fescue. Crop Sci. 2019, 59, 667–674. [Google Scholar] [CrossRef]
- Smirnov, O.E.; Kosyan, A.M.; Kosyk, O.I.; Taran, N.Y. Response of phenolic metabolism induced by aluminium toxicity in Fagopyrum esculentum moench. plants. Ukr. Biochem. J. 2015, 87, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D. Potential of salt and drought stress to increase pharmaceutical significant secondary compounds in plants. Landbauforschung Volkenrode 2008, 58, 139. [Google Scholar]
- Schroeter, H.; Boyd, C.; Spencer, J.P.; Williams, R.J.; Cadenas, E.; Rice-Evans, C. MAPK signaling in neurodegeneration: Influences of flavonoids and of nitric oxide. Neurobiol. Aging 2002, 23, 861–880. [Google Scholar] [CrossRef]
- Sharma, A.; Yuan, H.; Kumar, V.; Ramakrishnan, M.; Kohli, S.K.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Zheng, B. Castasterone attenuates insecticide induced phytotoxicity in mustard. Ecotoxicol. Environ. Saf. 2019, 179, 50–61. [Google Scholar] [CrossRef]
- Perin, E.C.; Da Silva Messias, R.; Borowski, J.M.; Crizel, R.L.; Schott, I.B.; Carvalho, I.R.; Rombaldi, C.V.; Galli, V. ABA-dependent salt and drought stress improve strawberry fruit quality. Food Chem. 2019, 271, 516–526. [Google Scholar] [CrossRef]
- Gharibi, S.; Sayed Tabatabaei, B.E.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech.f. Phytochemistry 2019, 162, 90–98. [Google Scholar] [CrossRef]
- Chen, Z.; Ma, Y.; Yang, R.; Gu, Z.; Wang, P. Effects of exogenous Ca2+ on phenolic accumulation and physiological changes in germinated wheat (Triticum aestivum L.) under UV-B radiation. Food Chem. 2019, 288, 368–376. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Li, Y.; Guo, T. Expression of flavonoid biosynthesis genes and accumulation of flavonoid in wheat leaves in response to drought stress. Plant Physiol. Biochem. 2014, 80, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Leng, X.; Jia, H.; Sun, X.; Shangguan, L.; Mu, Q.; Wang, B.; Fang, J. Comparative transcriptome analysis of grapevine in response to copper stress. Sci. Rep. 2015, 5, 17749. [Google Scholar] [CrossRef]
- Zhou, P.; Li, Q.; Liu, G.; Xu, N.; Yang, Y.; Zeng, W.; Chen, A.; Wang, S. Integrated analysis of transcriptomic and metabolomic data reveals critical metabolic pathways involved in polyphenol biosynthesis in Nicotiana tabacum under chilling stress. Funct. Plant Biol. 2018, 46, 30–43. [Google Scholar] [CrossRef]
- Pandey, N.; Sharma, C.P. Effect of heavy metals Co2+, Ni2+ and Cd2+ on growth and metabolism of cabbage. Plant Sci. 2002, 163, 753–758. [Google Scholar] [CrossRef]
- Villiers, F.; Ducruix, C.; Hugouvieux, V.; Jarno, N.; Ezan, E.; Garin, J.; Junot, C.; Bourguignon, J. Investigating the plant response to cadmium exposure by proteomic and metabolomic approaches. Proteomics 2011, 11, 1650–1663. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Interaction of 24-epibrassinolide and salicylic acid regulates pigment contents, antioxidative defense responses, and gene expression in Brassica juncea L. seedlings under Pb stress. Environ. Sci. Pollut. Res. 2018, 25, 15159–15173. [Google Scholar] [CrossRef]
- Kaur, R.; Yadav, P.; Sharma, A.; Kumar Thukral, A.; Kumar, V.; Kaur Kohli, S.; Bhardwaj, R. Castasterone and citric acid treatment restores photosynthetic attributes in Brassica juncea L. under Cd(II) toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 466–475. [Google Scholar] [CrossRef]
- Mira, L.; Fernandez, M.T.; Santos, M.; Rocha, R.; Florencio, M.H.; Jennings, K.R. Interactions of flavonoids with iron and copper ions: A mechanism for their antioxidant activity. Free Radic. Res. 2002, 36, 1199–1208. [Google Scholar] [CrossRef]
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free. Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Selenium ameliorates chromium toxicity through modifications in pigment system, antioxidative capacity, osmotic system, and metal chelators in Brassica juncea seedlings. S. Afr. J. Bot. 2018, 119, 1–10. [Google Scholar] [CrossRef]
- Trejo-Tapia, G.; Jimenez-Aparicio, A.; Rodriguez-Monroy, M.; De Jesus-Sanchez, A.; Gutierrez-Lopez, G. Influence of cobalt and other microelements on the production of betalains and the growth of suspension cultures of Beta vulgaris. Plant Cell Tiss. Org. Cult. 2001, 67, 19–23. [Google Scholar] [CrossRef]
- Zafari, S.; Sharifi, M.; Ahmadian Chashmi, N.; Mur, L.A. Modulation of Pb-induced stress in Prosopis shoots through an interconnected network of signaling molecules, phenolic compounds and amino acids. Plant Physiol. Biochem. 2016, 99, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kısa, D.; Elmastaş, M.; Öztürk, L.; Kayır, Ö. Responses of the phenolic compounds of Zea mays under heavy metal stress. Appl. Biol. Chem. 2016, 59, 813–820. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia Obovata under Cd and Zn stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Keilig, K.; Ludwig-Mueller, J. Effect of flavonoids on heavy metal tolerance in Arabidopsis thaliana seedlings. Bot. Stud. 2009, 50, 311–318. [Google Scholar]
- Kováčik, J.; Klejdus, B.; Hedbavny, J.; Štork, F.; Bačkor, M. Comparison of cadmium and copper effect on phenolic metabolism, mineral nutrients and stress-related parameters in Matricaria chamomilla plants. Plant Soil 2009, 320, 231. [Google Scholar] [CrossRef]
- Mishra, B.; Sangwan, N.S. Amelioration of cadmium stress in Withania somnifera by ROS management: Active participation of primary and secondary metabolism. Plant Growth Regul. 2019, 87, 403–412. [Google Scholar] [CrossRef]
- Mishra, B.; Sangwan, R.S.; Mishra, S.; Jadaun, J.S.; Sabir, F.; Sangwan, N.S. Effect of cadmium stress on inductive enzymatic and nonenzymatic responses of ROS and sugar metabolism in multiple shoot cultures of Ashwagandha (Withania somnifera Dunal). Protoplasma 2014, 251, 1031–1045. [Google Scholar] [CrossRef] [PubMed]
- Poonam, R.K.; Bhardwaj, R.; Sirhindi, G. Castasterone regulated polyphenolic metabolism and photosynthetic system in Brassica juncea plants under copper stress. J. Pharmacogn. Phytochem. 2015, 4, 282–289. [Google Scholar]
- Kaur, P.; Bali, S.; Sharma, A.; Vig, A.P.; Bhardwaj, R. Effect of earthworms on growth, photosynthetic efficiency and metal uptake in Brassica juncea L. plants grown in cadmium-polluted soils. Environ. Sci. Pollut. Res. 2017, 24, 13452–13465. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Bali, S.; Sharma, A.; Vig, A.P.; Bhardwaj, R. Role of earthworms in phytoremediation of cadmium (Cd) by modulating the antioxidative potential of Brassica juncea L. Appl. Soil Ecol. 2018, 124, 306–316. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Kumar, V.; Kaur, P.; Bhardwaj, R. Synergistic effect of 24-epibrassinolide and salicylic acid on photosynthetic efficiency and gene expression in Brassica juncea L. under Pb stress. Turk. J. Biol. 2017, 41, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Kirakosyan, A.; Seymour, E.; Kaufman, P.B.; Warber, S.; Bolling, S.; Chang, S.C. Antioxidant capacity of polyphenolic extracts from leaves of Crataegus laevigata and Crataegus monogyna (Hawthorn) subjected to drought and cold stress. J. Agric. Food Chem. 2003, 51, 3973–3976. [Google Scholar] [CrossRef]
- Ballizany, W.L.; Hofmann, R.W.; Jahufer, M.Z.Z.; Barrett, B.A. Multivariate associations of flavonoid and biomass accumulation in white clover (Trifolium repens) under drought. Funct. Plant Biol. 2012, 39, 167–177. [Google Scholar] [CrossRef]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Differential responses of phenolic compounds of Brassica napus under drought stress. Iran. J. Plant Physiol. 2018, 8, 2417–2425. [Google Scholar]
- Li, M.; Li, Y.; Zhang, W.; Li, S.; Gao, Y.; Ai, X.; Zhang, D.; Liu, B.; Li, Q. Metabolomics analysis reveals that elevated atmospheric CO2 alleviates drought stress in cucumber seedling leaves. Anal. Biochem. 2018, 559, 71–85. [Google Scholar] [CrossRef]
- Nichols, S.N.; Hofmann, R.W.; Williams, W.M. Physiological drought resistance and accumulation of leaf phenolics in white clover interspecific hybrids. Environ. Exp. Bot. 2015, 119, 40–47. [Google Scholar] [CrossRef]
- Sanchez-Rodriguez, E.; Moreno, D.A.; Ferreres, F.; Rubio-Wilhelmi Mdel, M.; Ruiz, J.M. Differential responses of five cherry tomato varieties to water stress: Changes on phenolic metabolites and related enzymes. Phytochemistry 2011, 72, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, I.; Alegre, L.; Van Breusegem, F.; Munne-Bosch, S. How relevant are flavonoids as antioxidants in plants? Trends Plant Sci. 2009, 14, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Gharibi, S.; Tabatabaei, B.E.; Saeidi, G.; Goli, S.A. Effect of Drought Stress on Total Phenolic, Lipid Peroxidation, and Antioxidant Activity of Achillea Species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crops Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Galieni, A.; Di Mattia, C.; De Gregorio, M.; Speca, S.; Mastrocola, D.; Pisante, M.; Stagnari, F. Effects of nutrient deficiency and abiotic environmental stresses on yield, phenolic compounds and antiradical activity in lettuce (Lactuca sativa L.). Sci. Hortic. 2015, 187, 93–101. [Google Scholar] [CrossRef]
- Varela, M.C.; Arslan, I.; Reginato, M.A.; Cenzano, A.M.; Luna, M.V. Phenolic compounds as indicators of drought resistance in shrubs from Patagonian shrublands (Argentina). Plant Physiol. Biochem. 2016, 104, 81–91. [Google Scholar] [CrossRef]
- Garcia-Calderon, M.; Pons-Ferrer, T.; Mrazova, A.; Pal’ove-Balang, P.; Vilkova, M.; Perez-Delgado, C.M.; Vega, J.M.; Eliasova, A.; Repcak, M.; Marquez, A.J.; et al. Modulation of phenolic metabolism under stress conditions in a Lotus japonicus mutant lacking plastidic glutamine synthetase. Front. Plant Sci. 2015, 6, 760. [Google Scholar] [CrossRef]
- Silva, F.L.B.; Vieira, L.G.E.; Ribas, A.F.; Moro, A.L.; Neris, D.M.; Pacheco, A.C. Proline accumulation induces the production of total phenolics in transgenic tobacco plants under water deficit without increasing the G6PDH activity. Theor. Exp. Plant Physiol. 2018, 30, 251–260. [Google Scholar] [CrossRef]
- Ghasemi Pirbalouti, A.; Malekpoor, F.; Salimi, A.; Golparvar, A. Exogenous application of chitosan on biochemical and physiological characteristics, phenolic content and antioxidant activity of two species of basil (Ocimum ciliatum and Ocimum basilicum) under reduced irrigation. Sci. Hortic. 2017, 217, 114–122. [Google Scholar] [CrossRef]
- Khalil, N.; Fekry, M.; Bishr, M.; El-Zalabani, S.; Salama, O. Foliar spraying of salicylic acid induced accumulation of phenolics, increased radical scavenging activity and modified the composition of the essential oil of water stressed Thymus vulgaris L. Plant Physiol. Biochem. 2018, 123, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kaur, L.; Zhawar, V.K. Phenolic parameters under exogenous ABA, water stress, salt stress in two wheat cultivars varying in drought tolerance. Ind. J. Plant Physiol. 2015, 20, 151–156. [Google Scholar] [CrossRef]
- Griesser, M.; Weingart, G.; Schoedl-Hummel, K.; Neumann, N.; Becker, M.; Varmuza, K.; Liebner, F.; Schuhmacher, R.; Forneck, A. Severe drought stress is affecting selected primary metabolites, polyphenols, and volatile metabolites in grapevine leaves (Vitis vinifera cv. Pinot noir). Plant Physiol. Biochem. 2015, 88, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; Di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [PubMed]
- Taïbi, K.; Taïbi, F.; Ait Abderrahim, L.; Ennajah, A.; Belkhodja, M.; Mulet, J.M. Effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidant defence systems in Phaseolus vulgaris L. S. Afr. J. Bot. 2016, 105, 306–312. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant Responses to Salt Stress: Adaptive Mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; Abreu, C.E.B.D.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Martinez, V.; Mestre, T.C.; Rubio, F.; Girones-Vilaplana, A.; Moreno, D.A.; Mittler, R.; Rivero, R.M. Accumulation of Flavonols over Hydroxycinnamic Acids Favors Oxidative Damage Protection under Abiotic Stress. Front. Plant Sci. 2016, 7, 838. [Google Scholar] [CrossRef]
- Chen, S.; Wu, F.; Li, Y.; Qian, Y.; Pan, X.; Li, F.; Wang, Y.; Wu, Z.; Fu, C.; Lin, H.; et al. NtMYB4 and NtCHS1 Are Critical Factors in the Regulation of Flavonoid Biosynthesis and Are Involved in Salinity Responsiveness. Front. Plant Sci. 2019, 10, 178. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V. Effects of salt stress on volatile compounds, total phenolic content and antioxidant activities of Salvia mirzayanii. S. Afr. J. Bot. 2014, 93, 92–97. [Google Scholar] [CrossRef]
- Rossi, L.; Borghi, M.; Francini, A.; Lin, X.; Xie, D.Y.; Sebastiani, L. Salt stress induces differential regulation of the phenylpropanoid pathway in Olea europaea cultivars Frantoio (salt-tolerant) and Leccino (salt-sensitive). J. Plant Physiol. 2016, 204, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Biochem. 2018, 129, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Golkar, P.; Taghizadeh, M. In vitro evaluation of phenolic and osmolite compounds, ionic content, and antioxidant activity in safflower (Carthamus tinctorius L.) under salinity stress. Plant Cell Tiss. Org. Cult. 2018, 134, 357–368. [Google Scholar] [CrossRef]
- Wang, F.; Zhu, H.; Chen, D.; Li, Z.; Peng, R.; Yao, Q. A grape bHLH transcription factor gene, VvbHLH1, increases the accumulation of flavonoids and enhances salt and drought tolerance in transgenic Arabidopsis thaliana. Plant Cell Tiss. Org. Cult. 2016, 125, 387–398. [Google Scholar] [CrossRef]
- Yan, J.; Wang, B.; Jiang, Y.; Cheng, L.; Wu, T. GmFNSII-controlled soybean flavone metabolism responds to abiotic stresses and regulates plant salt tolerance. Plant Cell Physiol. 2014, 55, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Ben-Abdallah, S.; Zorrig, W.; Amyot, L.; Renaud, J.; Hannoufa, A.; Lachâal, M.; Karray-Bouraoui, N. Potential production of polyphenols, carotenoids and glycoalkaloids in Solanum villosum Mill. under salt stress. Biologia 2019, 74, 309–324. [Google Scholar] [CrossRef]
- Scagel, C.F.; Lee, J.; Mitchell, J.N. Salinity from NaCl changes the nutrient and polyphenolic composition of basil leaves. Ind. Crops Prod. 2019, 127, 119–128. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef]
- Dkhil, B.B.; Denden, M. Effect of salt stress on growth, anthocyanins, membrane permeability and chlorophyll fluorescence of Okra (Abelmoschus esculentus L.) seedlings. Am. J. Plant Physiol. 2012, 7, 174–183. [Google Scholar] [CrossRef]
- Eryılmaz, F. The Relationships between Salt Stress and Anthocyanin Content in Higher Plants. Biotechnol. Biotechnol. Equip. 2006, 20, 47–52. [Google Scholar] [CrossRef]
- Aloisi, I.; Parrotta, L.; Ruiz, K.B.; Landi, C.; Bini, L.; Cai, G.; Biondi, S.; Del Duca, S. New Insight into Quinoa Seed Quality under Salinity: Changes in Proteomic and Amino Acid Profiles, Phenolic Content, and Antioxidant Activity of Protein Extracts. Front. Plant Sci. 2016, 7, 656. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Borgognone, D.; Rouphael, Y.; Cardarelli, M.; Bernardi, J.; Colla, G. Mild Potassium Chloride Stress Alters the Mineral Composition, Hormone Network, and Phenolic Profile in Artichoke Leaves. Front. Plant Sci. 2016, 7, 948. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, P.; Gu, Z.; Tao, Y.; Shen, C.; Zhou, Y.; Han, Y.; Yang, R. Ca2+ involved in GABA signal transduction for phenolics accumulation in germinated hulless barley under NaCl stress. Food Chem. X 2019, 2, 100023. [Google Scholar] [CrossRef]
- Çoban, Ö.; Göktürk Baydar, N. Brassinosteroid effects on some physical and biochemical properties and secondary metabolite accumulation in peppermint (Mentha piperita L.) under salt stress. Ind. Crops Prod. 2016, 86, 251–258. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Niazi, A.; Moghadam, A. Phenylalanine ammonia lyase isolation and functional analysis of phenylpropanoid pathway under salinity stress in ‘Salvia’ species. Aust. J. Crop Sci. 2015, 9, 656–665. [Google Scholar]
- Daayf, F.; Lattanzio, V. Recent Advances in Polyphenol Research; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Kolb, C.A.; Kaser, M.A.; Kopecky, J.; Zotz, G.; Riederer, M.; Pfundel, E.E. Effects of natural intensities of visible and ultraviolet radiation on epidermal ultraviolet screening and photosynthesis in grape leaves. Plant. Physiol. 2001, 127, 863–875. [Google Scholar] [CrossRef]
- Ghasemi, S.; Kumleh, H.H.; Kordrostami, M. Changes in the expression of some genes involved in the biosynthesis of secondary metabolites in Cuminum cyminum L. under UV stress. Protoplasma 2019, 256, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Siddiqui, S.; Upadhyay, N.; Soni, J. Effects of ultraviolet irradiation, pulsed electric field, hot water and ethanol vapours treatment on functional properties of mung bean sprouts. J. Food Sci. Technol. 2014, 51, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Charles, M.T.; Luo, Z.; Mimee, B.; Veronneau, P.Y.; Rolland, D.; Roussel, D. Preharvest Ultraviolet C Irradiation Increased the Level of Polyphenol Accumulation and Flavonoid Pathway Gene Expression in Strawberry Fruit. J. Agric. Food Chem. 2017, 65, 9970–9979. [Google Scholar] [CrossRef] [PubMed]
- Demkura, P.V.; Abdala, G.; Baldwin, I.T.; Ballare, C.L. Jasmonate-dependent and -independent pathways mediate specific effects of solar ultraviolet B radiation on leaf phenolics and antiherbivore defense. Plant Physiol. 2010, 152, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Berli, F.J.; Fanzone, M.; Piccoli, P.; Bottini, R. Solar UV-B and ABA are involved in phenol metabolism of Vitis vinifera L. increasing biosynthesis of berry skin polyphenols. J. Agric. Food Chem. 2011, 59, 4874–4884. [Google Scholar] [CrossRef] [PubMed]
- Nenadis, N.; Llorens, L.; Koufogianni, A.; Diaz, L.; Font, J.; Gonzalez, J.A.; Verdaguer, D. Interactive effects of UV radiation and reduced precipitation on the seasonal leaf phenolic content/composition and the antioxidant activity of naturally growing Arbutus unedo plants. J. Photochem. Photobiol. B 2015, 153, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Rodriguez, M.; Nair, V.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velazquez, D.A. UVA, UVB Light, and Methyl Jasmonate, Alone or Combined, Redirect the Biosynthesis of Glucosinolates, Phenolics, Carotenoids, and Chlorophylls in Broccoli Sprouts. Int. J. Mol. Sci. 2017, 18, 2330. [Google Scholar] [CrossRef]
- Liu, M.; Cao, B.; Zhou, S.; Liu, Y. Responses of the flavonoid pathway to UV-B radiation stress and the correlation with the lipid antioxidant characteristics in the desert plant Caryopteris mongolica. Acta Ecol. Sin. 2012, 32, 150–155. [Google Scholar] [CrossRef]
- Nascimento, L.; Leal-Costa, M.V.; Menezes, E.A.; Lopes, V.R.; Muzitano, M.F.; Costa, S.S.; Tavares, E.S. Ultraviolet-B radiation effects on phenolic profile and flavonoid content of Kalanchoe pinnata. J. Photochem. Photobiol. B 2015, 148, 73–81. [Google Scholar] [CrossRef]
- Sytar, O.; Zivcak, M.; Bruckova, K.; Brestic, M.; Hemmerich, I.; Rauh, C.; Simko, I. Shift in accumulation of flavonoids and phenolic acids in lettuce attributable to changes in ultraviolet radiation and temperature. Sci. Hortic. 2018, 239, 193–204. [Google Scholar] [CrossRef]
- Lee, M.J.; Son, J.E.; Oh, M.M. Growth and phenolic compounds of Lactuca sativa L. grown in a closed-type plant production system with UV-A, -B, or -C lamp. J. Sci. Food Agric. 2014, 94, 197–204. [Google Scholar] [CrossRef]
- Huyskens-Keil, S.; Eichholz, I.; Kroh, L.; Rohn, S. UV-B induced changes of phenol composition and antioxidant activity in black currant fruit (Ribes nigrum L.). J. App. Bot. Food Qual. 2012, 81, 140–144. [Google Scholar]
- Mariz-Ponte, N.; Mendes, R.J.; Sario, S.; De Oliveira, J.F.; Melo, P.; Santos, C. Tomato plants use non-enzymatic antioxidant pathways to cope with moderate UV-A/B irradiation: A contribution to the use of UV-A/B in horticulture. J. Plant Physiol. 2018, 221, 32–42. [Google Scholar] [CrossRef]
- Chen, Z.; Ma, Y.; Weng, Y.; Yang, R.; Gu, Z.; Wang, P. Effects of UV-B radiation on phenolic accumulation, antioxidant activity and physiological changes in wheat (Triticum aestivum L.) seedlings. Food Biosci. 2019, 30, 100409. [Google Scholar] [CrossRef]
- Alonso, R.; Berli, F.J.; Fontana, A.; Piccoli, P.; Bottini, R. Malbec grape (Vitis vinifera L.) responses to the environment: Berry phenolics as influenced by solar UV-B, water deficit and sprayed abscisic acid. Plant Physiol. Biochem. 2016, 109, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Swieca, M. Elicitation with abiotic stresses improves pro-health constituents, antioxidant potential and nutritional quality of lentil sprouts. Saudi J. Biol. Sci. 2015, 22, 409–416. [Google Scholar] [CrossRef]
- Wang, L.; Shan, T.; Xie, B.; Ling, C.; Shao, S.; Jin, P.; Zheng, Y. Glycine betaine reduces chilling injury in peach fruit by enhancing phenolic and sugar metabolisms. Food Chem. 2019, 272, 530–538. [Google Scholar] [CrossRef]
- Mahdavi, V.; Farimani, M.M.; Fathi, F.; Ghassempour, A. A targeted metabolomics approach toward understanding metabolic variations in rice under pesticide stress. Anal. Biochem. 2015, 478, 65–72. [Google Scholar] [CrossRef]
- Korosi, L.; Bouderias, S.; Csepregi, K.; Bognar, B.; Teszlak, P.; Scarpellini, A.; Castelli, A.; Hideg, E.; Jakab, G. Nanostructured TiO2-induced photocatalytic stress enhances the antioxidant capacity and phenolic content in the leaves of Vitis vinifera on a genotype-dependent manner. J. Photochem. Photobiol. B 2019, 190, 137–145. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Singh, R.; Thukral, A.K.; Bhardwaj, R. Effect of seed pre-soaking with 24-epibrassinolide on growth and photosynthetic parameters of Brassica juncea L. in imidacloprid soil. Ecotoxicol. Environ. Saf. 2016, 133, 195–201. [Google Scholar] [CrossRef]
- Zlotek, U.; Szymanowska, U.; Baraniak, B.; Karas, M. Antioxidant activity of polyphenols of adzuki bean (Vigna angularis) germinated in abiotic stress conditions. Acta Sci. Pol. Technol. Aliment. 2015, 14, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Commisso, M.; Toffali, K.; Strazzer, P.; Stocchero, M.; Ceoldo, S.; Baldan, B.; Levi, M.; Guzzo, F. Impact of phenylpropanoid compounds on heat stress tolerance in carrot cell cultures. Front. Plant Sci. 2016, 7, 1439. [Google Scholar] [CrossRef] [PubMed]
- Cingoz, G.S.; Gurel, E. Effects of salicylic acid on thermotolerance and cardenolide accumulation under high temperature stress in Digitalis trojana Ivanina. Plant Physiol. Biochem. 2016, 105, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.; Yaish, M.W. Antifreeze proteins in overwintering plants: A tale of two activities. Trends Plant. Sci. 2004, 9, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, Z.; Lv, X.; Cheng, N.; Peng, B.; Cao, W. Effect of 24-epibrassinolide on chilling injury of peach fruit in relation to phenolic and proline metabolisms. Post. Biol. Technol. 2016, 111, 390–397. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic Acid Seed Treatment Stimulates Insecticide Detoxification in Brassica juncea L. Front. Plant Sci. 2018, 9, 1609. [Google Scholar] [CrossRef]
- Nourozi, E.; Hosseini, B.; Maleki, R.; Mandoulakani, B.A. Pharmaceutical important phenolic compounds overproduction and gene expression analysis in Dracocephalum kotschyi hairy roots elicited by SiO2 nanoparticles. Ind. Crops Prod. 2019, 133, 435–446. [Google Scholar] [CrossRef]
- Girilal, M.; Fayaz, A.M.; Elumalai, L.K.; Sathiyaseelan, A.; Gandhiappan, J.; Kalaichelvan, P.T. Comparative Stress Physiology Analysis of Biologically and Chemically Synthesized Silver Nanoparticles on Solanum lycopersicum L. Colloid Interface Sci. Commun. 2018, 24, 1–6. [Google Scholar] [CrossRef]
- Raigond, P.; Raigond, B.; Kaundal, B.; Singh, B.; Joshi, A.; Dutt, S. Effect of zinc nanoparticles on antioxidative system of potato plants. J. Environ. Biol. 2017, 38, 435–439. [Google Scholar] [CrossRef]
- Singh, O.S.; Pant, N.C.; Laishram, M.L.; Tewari, R.D.; Joshi, K.; Pandey, C. Effect of CuO nanoparticles on polyphenols content and antioxidant activity in Ashwagandha (Withania somnifera L. Dunal). J. Pharmacogn. Phytochem. 2018, 7, 3433–3439. [Google Scholar]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).