
Evidence for an Ordering Transition near 120 K in an Intrinsically Disordered Protein, Casein



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
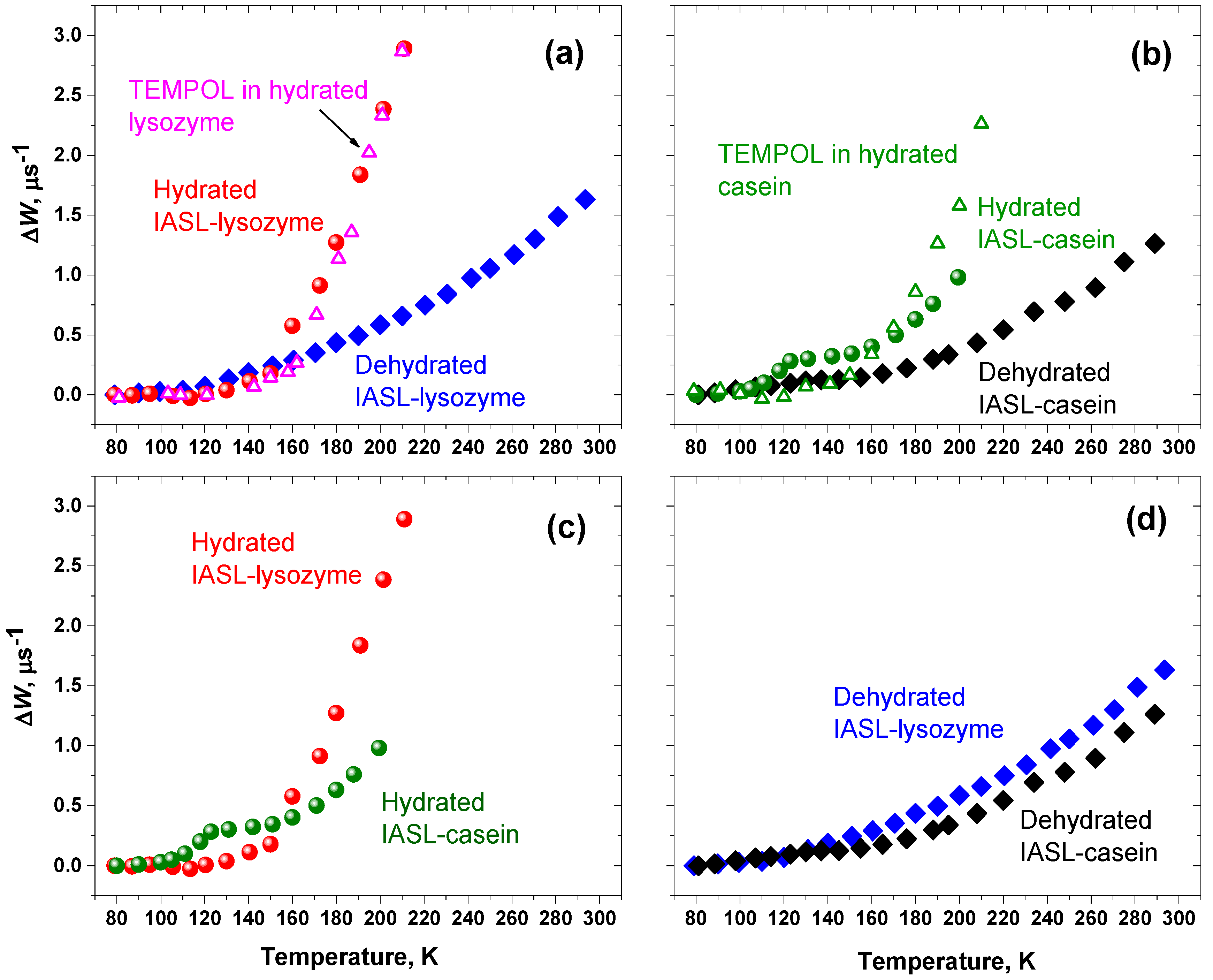
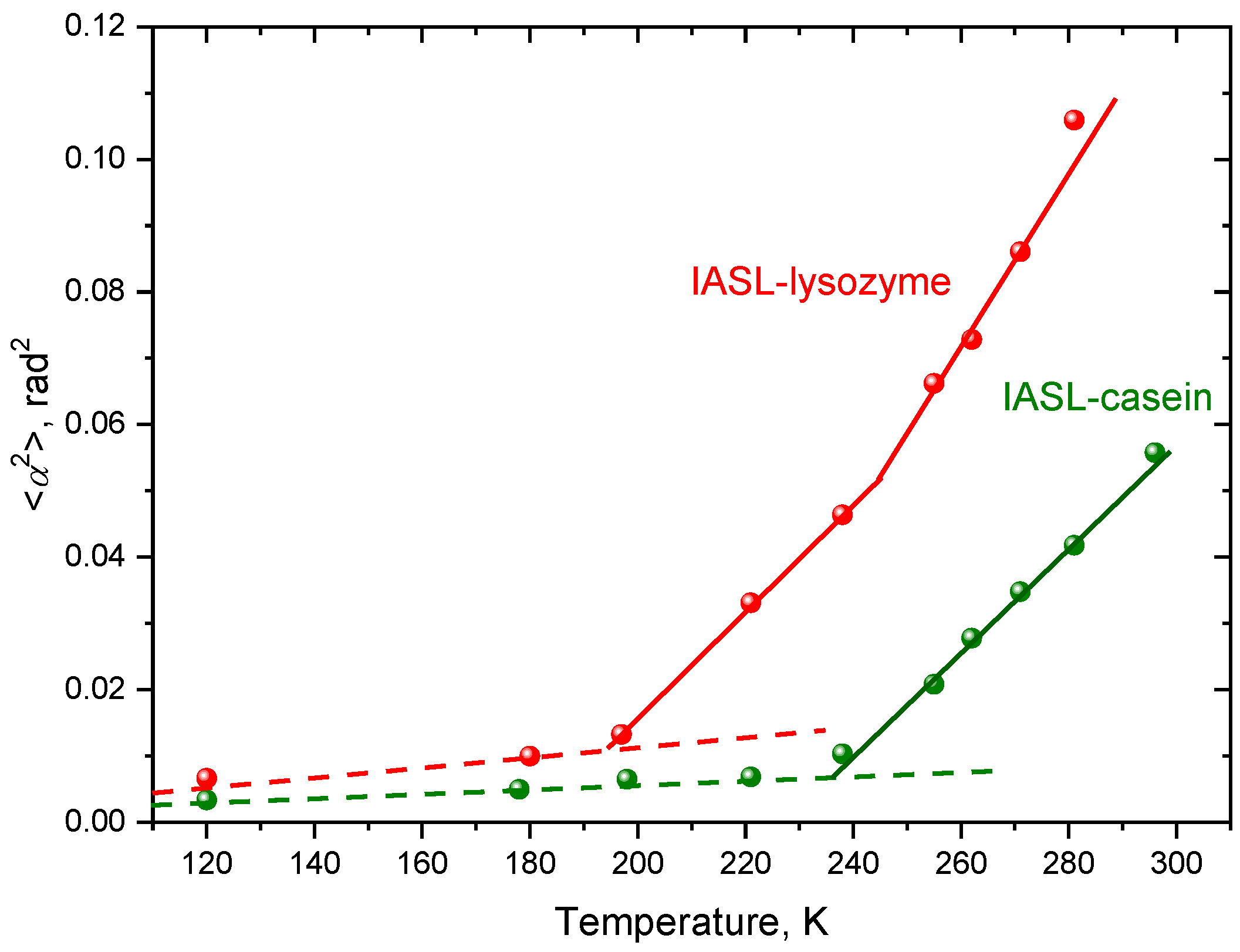
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Sample Availability
References
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef]
- Uversky, V.N. Unusual biophysics of intrinsically disordered proteins. Biochim. Biophys. Acta Proteins Proteomics 2013, 1834, 932–951. [Google Scholar] [CrossRef]
- Oldfield, C.J.; Dunker, A.K. Intrinsically Disordered Proteins and Intrinsically Disordered Protein Regions. Annu. Rev. Biochem. 2014, 83, 553–584. [Google Scholar] [CrossRef]
- Deiana, A.; Forcelloni, S.; Porrello, A.; Giansanti, A. Intrinsically disordered proteins and structured proteins with intrinsically disordered regions have different functional roles in the cell. PLoS ONE 2019, 14, e0217889. [Google Scholar] [CrossRef] [Green Version]
- Drake, J.A.; Pettitt, B.M. Physical chemistry of the protein backbone: Enabling the mechanisms of intrinsic protein disorder. J. Phys. Chem. B 2020, 124, 4379–4390. [Google Scholar] [CrossRef]
- Hosoya, Y.; Ohkanda, J. Intrinsically Disordered Proteins as Regulators of Transient Biological Processes and as Untapped Drug Targets. Molecules 2021, 26, 2118. [Google Scholar] [CrossRef]
- Uversky, V.N. Supramolecular Fuzziness of Intracellular Liquid Droplets: Liquid-Liquid Phase Transitions, Membrane-Less Organelles, and Intrinsic Disorder. Molecules 2019, 24, 3265. [Google Scholar] [CrossRef] [Green Version]
- Dyson, H.J.; Wright, P.E. NMR illuminates intrinsic disorder. Curr. Opinion Struct. Biol. 2021, 70, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Weickert, S.; Cattani, J.; Drescher, M. Intrinsically disordered proteins (IDPs) studied by EPR and in-cell EPR. In Electron Paramagnetic Resonance; Gilbert, B.C., Chechik, V., Murphy, D.M., Eds.; Royal Society of Chemistry: Cambridge, UK, 2018; Volume 26, pp. 1–37. [Google Scholar] [CrossRef]
- Eschmann, N.A.; Georgieva, E.R.; Ganguly, P.P.; Borbat, P.; Rappaport, M.D.; Akdogan, Y.; Freed, J.H.; Shea, J.-E.; Han, S. Signature of an aggregation-prone conformation of tau. Sci. Rep. 2017, 7, 44739. [Google Scholar] [CrossRef] [PubMed]
- Sicoli, G.; Kress, T.; Vezin, H.; Ledolter, K.; Kurzbach, D. A switch between two intrinsically disordered conformational ensembles modulates the active site of a basic-helix−loop−helix transcription factor. J. Phys. Chem. Lett. 2020, 11, 8944–8951. [Google Scholar] [CrossRef] [PubMed]
- Chinak, O.A.; Shernyukov, A.V.; Ovcherenko, S.S.; Sviridov, E.A.; Golyshev, V.M.; Fomin, A.S.; Pyshnaya, I.A.; Kuligina, E.V.; Richter, V.A.; Bagryanskaya, E.G. Structural and aggregation features of a human k-casein fragment with antitumor and cell-penetrating properties. Molecules 2019, 24, 2919. [Google Scholar] [CrossRef] [Green Version]
- Kirilina, E.P.; Dzuba, S.A.; Maryasov, A.G.; Tsvetkov, Y.D. Librational dynamics of nitroxide molecules in a molecular glass studied by echo-detected EPR. Appl. Magn. Reson. 2021, 21, 203–221. [Google Scholar] [CrossRef]
- Erilov, D.A.; Bartucci, R.; Guzzi, R.; Marsh, D.; Dzuba, S.A.; Sportelli, L. Echo-detected electron paramagnetic resonance spectra of spin-labeled lipids in membrane model systems. J. Phys. Chem. B 2004, 108, 4501–4507. [Google Scholar] [CrossRef] [Green Version]
- Isaev, N.P.; Dzuba, S.A. Fast stochastic librations and slow rotations of spin labeled stearic acids in a model phospholipid bilayer at cryogenic temperatures. J. Phys. Chem. B 2008, 112, 13285–13291. [Google Scholar] [CrossRef] [PubMed]
- Golysheva, E.A.; De Zotti, M.; Toniolo, C.; Formaggio, F.; Dzuba, S.A. Low-temperature dynamical transition in lipid bilayers detected by spin-label ESE spectroscopy. Appl. Magn. Reson. 2018, 49, 1369–1383. [Google Scholar] [CrossRef]
- Golysheva, E.A.; Dzuba, S.A. Lipid chain mobility and packing in DOPC bilayers at cryogenic temperatures. Chem. Phys. Lipids 2021, 226, 104817. [Google Scholar] [CrossRef]
- Bartucci, R.; Guzzi, R.; De Zotti, M.; Toniolo, C.; Sportelli, L.; Marsh, D. Backbone Dynamics of Alamethicin Bound to Lipid Membranes: Spin-Echo Electron Paramagnetic Resonance of TOAC-Spin Labels. Biophys. J. 2008, 94, 2698–2705. [Google Scholar] [CrossRef] [Green Version]
- Aloi, E.; Oranges, M.; Guzzi, R.; Bartucci, R. Low-temperature dynamics of chain-labeled lipids in ester- and ether-linked phosphatidylcholine membranes. J. Phys. Chem. B 2017, 121, 9239–9246. [Google Scholar] [CrossRef]
- Ivanov, M.Y.; Prikhod’ko, S.A.; Adonin, N.Y.; Kirilyuk, I.A.; Adichtchev, S.V.; Surovtsev, N.V.; Dzuba, S.A.; Fedin, M.V. Structural anomalies in ionic liquids near the glass transition revealed by pulse EPR. J. Phys. Chem. Lett. 2018, 9, 4607–4612. [Google Scholar] [CrossRef]
- Ivanov, M.Y.; Poryvaev, A.S.; Polyukhov, D.M.; Prikhod’ko, S.A.; Adonin, N.Y.; Fedin, M.V. Nanoconfinement effects on structural anomalies in imidazolium ionic liquids. Nanoscale 2020, 12, 23480–23487. [Google Scholar] [CrossRef]
- Bakulina, O.D.; Ivanov, M.Y.; Prikhod’ko, S.A.; Pylaeva, S.I.; Zaytseva, V.; Surovtsev, N.V.; Adonin, N.Y.; Fedin, M.V. Nanocage formation and structural anomalies in imidazolium ionic liquid glasses governed by alkyl chains of cations. Nanoscale 2020, 12, 19982–19991. [Google Scholar] [CrossRef]
- Ivanov, M.Y.; Prikhod’ko, S.A.; Adonin, N.Y.; Fedin, M.V. Structural anomalies in binary mixtures of ionic liquid [Bmim]BF4 with water studied by EPR. J. Phys. Chem. B 2019, 123, 9956–9962. [Google Scholar] [CrossRef]
- De Simone, F.; Guzzi, R.; Sportelli, L.; Marsh, D.; Bartucci, R. Electron spin-echo studies of spin-labelled lipid membranes and free fatty acids interacting with human serum albumin. Biochim. Biophys. Acta 2007, 1768, 1541–1549. [Google Scholar] [CrossRef] [Green Version]
- Konov, K.B.; Isaev, N.P.; Dzuba, S.A. Low-temperature molecular motions in lipid bilayers in the presence of sugars: Insights into cryoprotective mechanisms. J. Phys. Chem. B 2014, 118, 12478–12485. [Google Scholar] [CrossRef] [PubMed]
- Aloi, E.; Bartucci, R. Interdigitated lamellar phases in the frozen state: Spin-label CW- and FT-EPR. Biophys. Chem. 2019, 253, 106229. [Google Scholar] [CrossRef] [PubMed]
- Golysheva, E.A.; Shevelev, G.Y.; Dzuba, S.A. Dynamical transition in molecular glasses and proteins observed by spin relaxation of nitroxide spin probes and labels. J. Chem. Phys. 2017, 147, 064501. [Google Scholar] [CrossRef] [PubMed]
- Aloi, E.; Guzzi, R.; Bartucci, R. Unsaturated lipid bilayers at cryogenic temperature: Librational dynamics of chain-labeled lipids from pulsed and CW-EPR. Phys. Chem. Chem. Phys. 2019, 21, 18699–18705. [Google Scholar] [CrossRef]
- Kämpf, K.; Demuth, D.; Zamponi, M.; Wuttke, J.; Vogel, M. Quasielastic neutron scattering studies on couplings of protein and water dynamics in hydrated elastin. J. Chem. Phys. 2020, 152, 245101. [Google Scholar] [CrossRef]
- Schirò, G.; Weik, M. Role of hydration water in the onset of protein structural dynamics. J. Phys. Condens. Matter 2019, 31, 463002. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xiang, S.; Zhu, H.; Li, L. The Structural and Dynamical Properties of the Hydration of SNase Based on a Molecular Dynamics Simulation. Molecules 2021, 26, 5403. [Google Scholar] [CrossRef]
- Moitzi, C.; Portnaya, I.; Glatter, O.; Ramon, O.; Danino, D. Effect of temperature on self-assembly of bovine β-casein above and below isoelectric pH. Structural analysis by cryogenic-transmission electron microscopy and small-angle X-ray scattering. Langmuir 2008, 24, 3020–3029. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Singh, L.R.; Dar, T.A. Taurine induces an ordered but functionally inactive conformation in intrinsically disordered casein proteins. Sci. Rep. 2020, 10, 3503. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, D.; Carnerup, A.M.; Schillén, K.; Miguel, M.D.; Lindman, B. Phase behavior and coassembly of DNA and lysozyme in dilute aqueous mixtures: A model investigation of DNA−protein interactions. Langmuir 2010, 26, 2986–2988. [Google Scholar] [CrossRef]
- Rupley, J.A.; Careri, G. Protein hydration and function. Adv. Protein Chem. 1991, 41, 37–172. [Google Scholar] [CrossRef]
- Dhindsa, G.K.; Tyagi, M.; Chu, X.-Q. Temperature-dependent dynamics of dry and hydrated β-casein studied by quasielastic neutron scattering. J. Phys. Chem. B 2014, 118, 10821–10829. [Google Scholar] [CrossRef] [PubMed]
- Perticaroli, S.; Nickels, J.D.; Ehlers, G.; Mamontov, E.; Sokolov, A.P. Dynamics and rigidity in an intrinsically disordered protein, β-casein. J. Phys. Chem. B 2014, 118, 7317–7326. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D. Spin-Label Electron Paramagnetic Resonance Spectroscopy; SRC Press: Boca Raton, FL, USA, 2020. [Google Scholar] [CrossRef]
- Erilov, D.A.; Bartucci, R.; Guzzi, R.; Shubin, A.A.; Maryasov, A.G.; Marsh, D.; Dzuba, S.A.; Sportelli, L. Water concentration profiles in membranes measured by ESEEM of spin-labeled lipids. J. Phys. Chem. B 2005, 109, 12003–12013. [Google Scholar] [CrossRef] [PubMed]
- Konov, K.B.; Isaev, N.P.; Dzuba, S.A. Glycerol penetration profile in phospholipid bilayers measured by ESEEM of spin-labelled lipids. Mol. Phys. 2013, 111, 2882–2886. [Google Scholar] [CrossRef]
- Milov, A.D.; Samoilova, R.I.; Shubin, A.A.; Grishin, Y.A.; Dzuba, S.A. ESEEM measurements of local water concentration in D2O-containing spin-labeled systems. Appl. Magn. Reson. 2008, 35, 73–94. [Google Scholar] [CrossRef]
- Du, J.L.; Eaton, G.R.; Eaton, S.S. Temperature, Orientation, and Solvent Dependence of Electron Spin-Lattice Relaxation Rates for Nitroxyl Radicals in Glassy Solvents and Doped Solids. J. Magn. Reason. Ser. A 1995, 115, 213–221. [Google Scholar] [CrossRef]
- Sato, H.; Kathirvelu, V.; Spagnol, G.; Rajca, S.; Rajca, A.; Eaton, S.S.; Eaton, G.R. Impact of Electron-Electron Spin Interaction on Electron Spin Relaxation of Nitroxide Diradicals and Tetraradical in Glassy Solvents Between 10 and 300 K. J. Phys. Chem. B 2008, 112, 2818–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Bottle, S.E.; Blinco, J.P.; Micallef, A.S.; Eaton, G.R.; Eaton, S.S. Electron spin-lattice relaxation of nitroxyl radicals in temperature ranges that span glassy solutions to low-viscosity liquids. J. Magn. Reson. 2008, 191, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sannikova, N.; Timofeev, I.; Bagryanskaya, E.; Bowman, M.; Fedin, M.; Krumkacheva, O. Electron Spin Relaxation of Photoexcited Porphyrin in Water—Glycerol Glass. Molecules 2020, 25, 2677. [Google Scholar] [CrossRef] [PubMed]
- Paschenko, S.V.; Toropov, Y.V.; Dzuba, S.A.; Tsvetkov, Y.D.; Vorobiev, A.K. Temperature dependence of amplitudes of libration motion of guest spin probe molecules in organic glasses. J. Chem. Phys. 1999, 110, 8150–8154. [Google Scholar] [CrossRef]
- Poluektov, O.G.; Utschig, L.M.; Dalosto, S.; Thurnauer, M.C. Probing Local Dynamics of the Photosynthetic Bacterial Reaction Center with a Cysteine Specific Spin Label. J. Phys. Chem. B 2003, 107, 6239–6244. [Google Scholar] [CrossRef]
- Scarpelli, F.; Bartucci, R.; Sportelli, L.; Guzzi, R. Solvent effect on librational dynamics of spin-labelled haemoglobin by ED- and CW-EPR. Eur. Biophys J. 2011, 40, 273–279. [Google Scholar] [CrossRef]
- Marsh, D.; Bartucci, R.; Guzzi, R.; Sportelli, L.; Esmann, M. Librational fluctuations in protein glasses. Biochim. Biophys. Acta 2013, 1834, 1591–1595. [Google Scholar] [CrossRef]
- Guzzi, R.; Bartucci, R.; Esmann, M.; Marsh, D. Lipid librations at the interface with the Na, K-ATPase. Biophys. J. 2015, 108, 2825–2832. [Google Scholar] [CrossRef] [Green Version]
- Fajer, P.G. Determination of spin-label orientation within the myosin head. Proc. Nati. Acad. Sci. USA 1994, 91, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Busi, B.; Yarava, J.R.; Bertarello, A.; Freymond, F.; Adamski, W.; Maurin, D.; Hiller, M.; Oschkinat, H.; Blackledge, M.; Emsley, L. Similarities and differences among protein dynamics studied by variable temperature nuclear magnetic resonance relaxation. J. Phys. Chem. B 2021, 125, 2212–2221. [Google Scholar] [CrossRef]
- Gallat, F.-X.; Laganowsky, A.; Wood, K.; Gabel, F.; van Eijck, L.; Wuttke, J.; Moulin, M.; Härtlein, M.; Eisenberg, D.; Colletier, J.-P.; et al. Dynamical coupling of intrinsically disordered proteins and their hydration water: Comparison with folded soluble and membrane proteins. Biophys. J. 2012, 103, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golysheva, E.A.; Samoilova, R.I.; De Zotti, M.; Toniolo, C.; Formaggio, F.; Dzuba, S.A. Electron spin echo detection of stochastic molecular librations: Non-cooperative motions on solid surface. J. Magn. Reson. 2019, 309, 106621. [Google Scholar] [CrossRef] [PubMed]
- Golysheva, E.A.; Samoilova, R.I.; De Zotti, M.; Formaggio, F.; Gobbo, M.; Dzuba, S.A. ESE-detected molecular motions of spin-labeled molecules on a solid inorganic surface: Motional models and onset temperatures. Appl. Magn. Reson. 2020, 51, 1019–1029. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maslennikova, N.A.; Golysheva, E.A.; Dzuba, S.A. Evidence for an Ordering Transition near 120 K in an Intrinsically Disordered Protein, Casein. Molecules 2021, 26, 5971. https://doi.org/10.3390/molecules26195971
Maslennikova NA, Golysheva EA, Dzuba SA. Evidence for an Ordering Transition near 120 K in an Intrinsically Disordered Protein, Casein. Molecules. 2021; 26(19):5971. https://doi.org/10.3390/molecules26195971
Chicago/Turabian StyleMaslennikova, Natalya A., Elena A. Golysheva, and Sergei A. Dzuba. 2021. "Evidence for an Ordering Transition near 120 K in an Intrinsically Disordered Protein, Casein" Molecules 26, no. 19: 5971. https://doi.org/10.3390/molecules26195971