
Evening Primrose Oil Enhances Tamoxifen’s Anticancer Activity against Breast Cancer Cells by Inducing Apoptosis, Inhibiting Angiogenesis, and Arresting the Cell Cycle

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Cell Line
2.3. Ethical Approval
2.4. Cell Viability Assay
- TAM concentrations (1, 0.5, 0.25, 0.125, and 0.0652 μg/mL) or EPO concentrations (250, 125, 62.5, 31.25, and 15.63 μg/mL) were added to all MCF-7 wells, except the control wells.
- TAM concentrations (8, 4, 2, 1, and 0.5 μg/mL) or EPO concentrations (250, 125, 62.5, 31.25, and 15.63 μg/mL) were added to all MDA-MB-231 wells except the control wells.
2.5. Treatment of MCF-7 and MDA-MB-231 Cells for Biomolecular Investigations
- Control: cells were treated with 1% DMSO, in both cell lines.
- EPO: cells were treated with (IC50) concentration of EPO, in both cell lines.
- TAM: cells were treated with (IC50) concentration of TAM, in both cell lines.
- Combination-1: cells were treated with 50% of (IC50) concentration of EPO +50% of (IC50) concentration of TAM, in both cell lines.
- Combination-2: cells were treated with (IC50) concentration of EPO + (IC50) concentration of TAM, in both cell lines.
2.6. Preparation of Cell Lysates
2.7. Protein Quantitation Using BCA Assay
2.8. Biomarker Analysis Using ELISA Technique
2.9. Caspase-3 Activity Assay
2.10. qRT-PCR Determination of mRNA Genes Expression of VEGF, Bax, and Bcl-2
2.11. Annexin V—FITC/PI Double Staining for Flow-Cytometric Apoptosis Detection
2.12. Cell-Cycle Analysis by Flow Cytometry
2.13. Characterization of EPO Using Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
2.14. Statistical Analysis
3. Results
3.1. GC-MS Analysis of EPO
3.2. Effect of EPO and TAM on MCF-7 and MDA-MB-231 Cell Viability
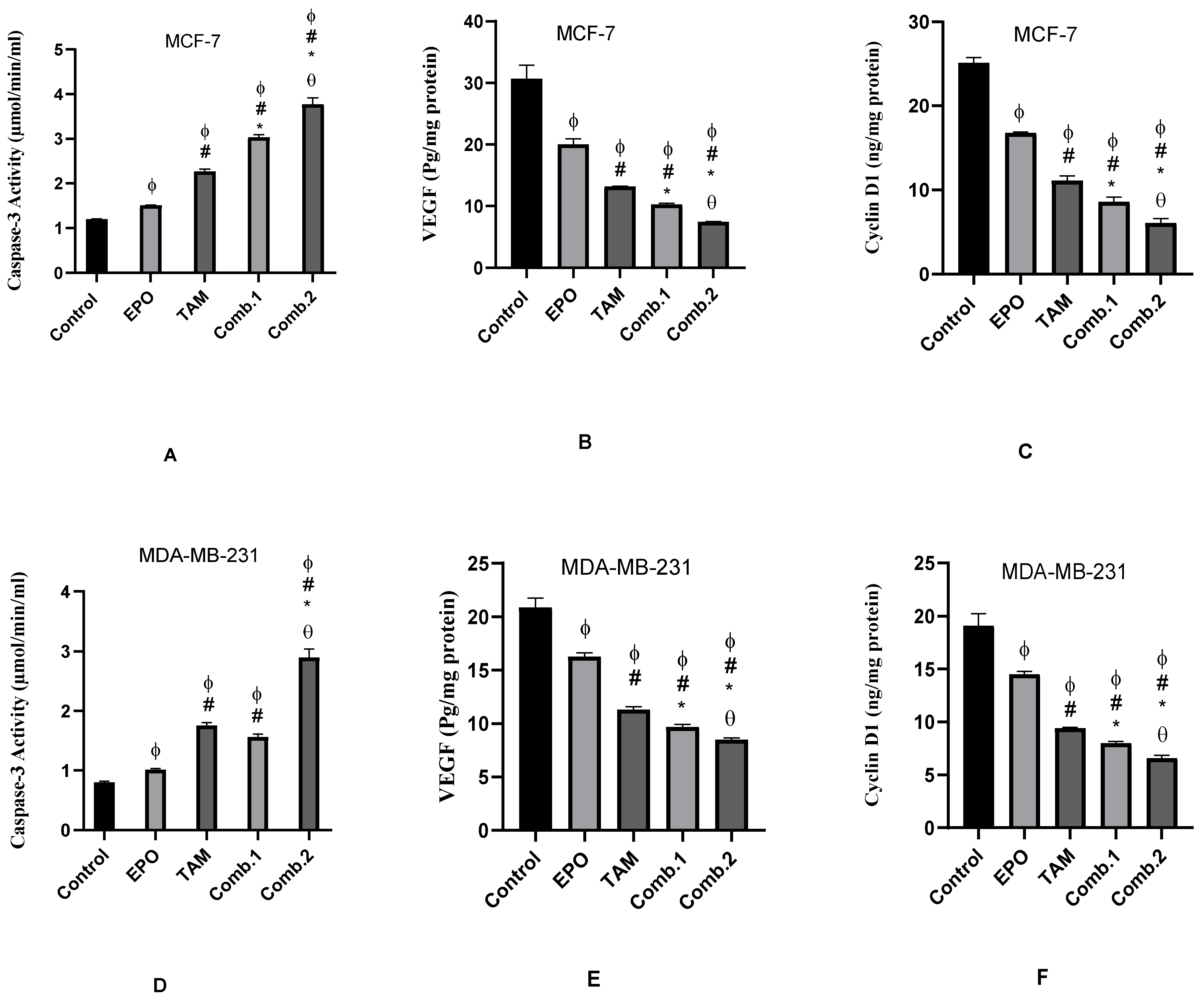
3.3. Effect of TAM and EPO, Alone or in Combination, on Caspase-3 Activity, VEGF and Cyclin D-1 Levels
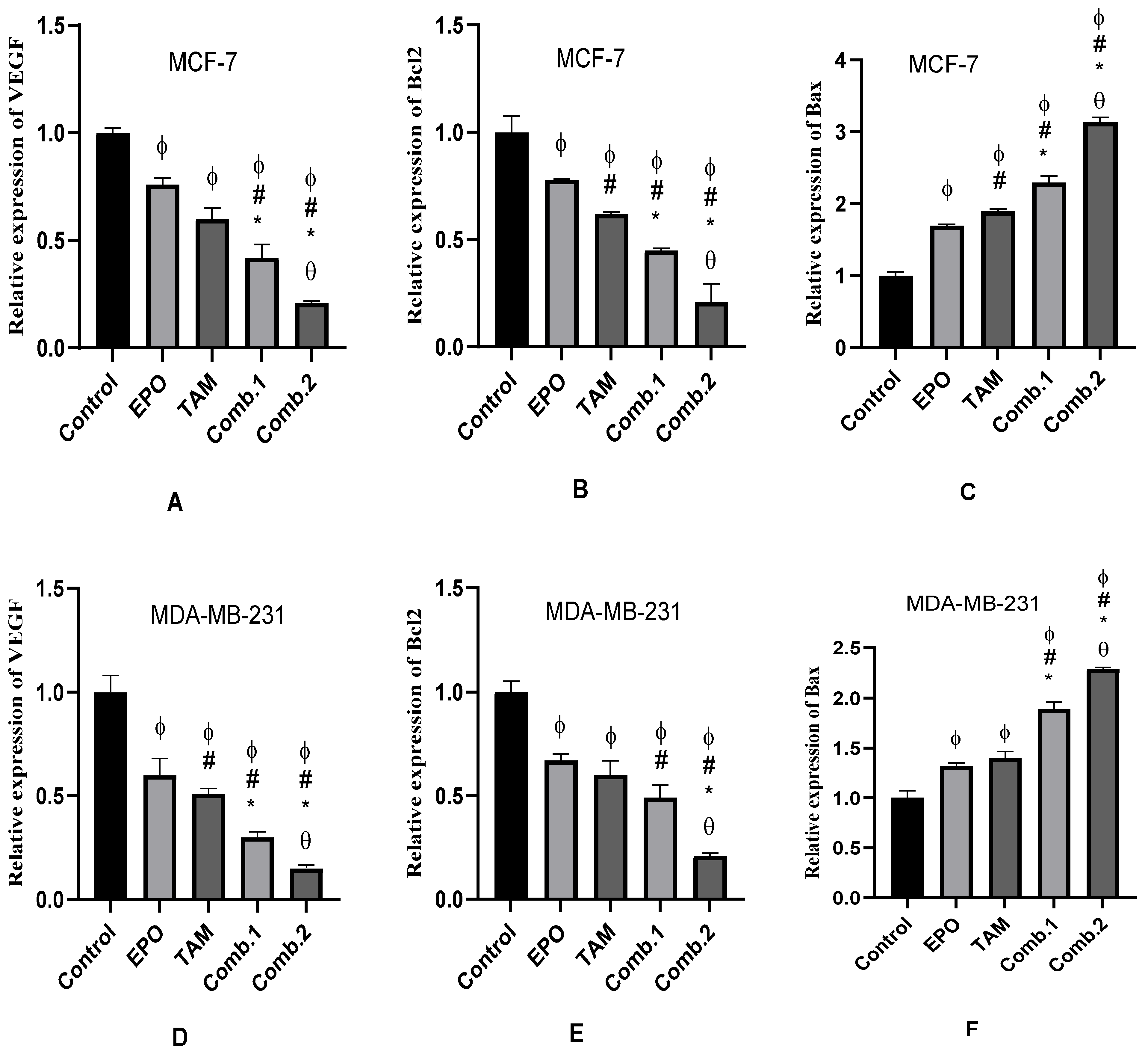
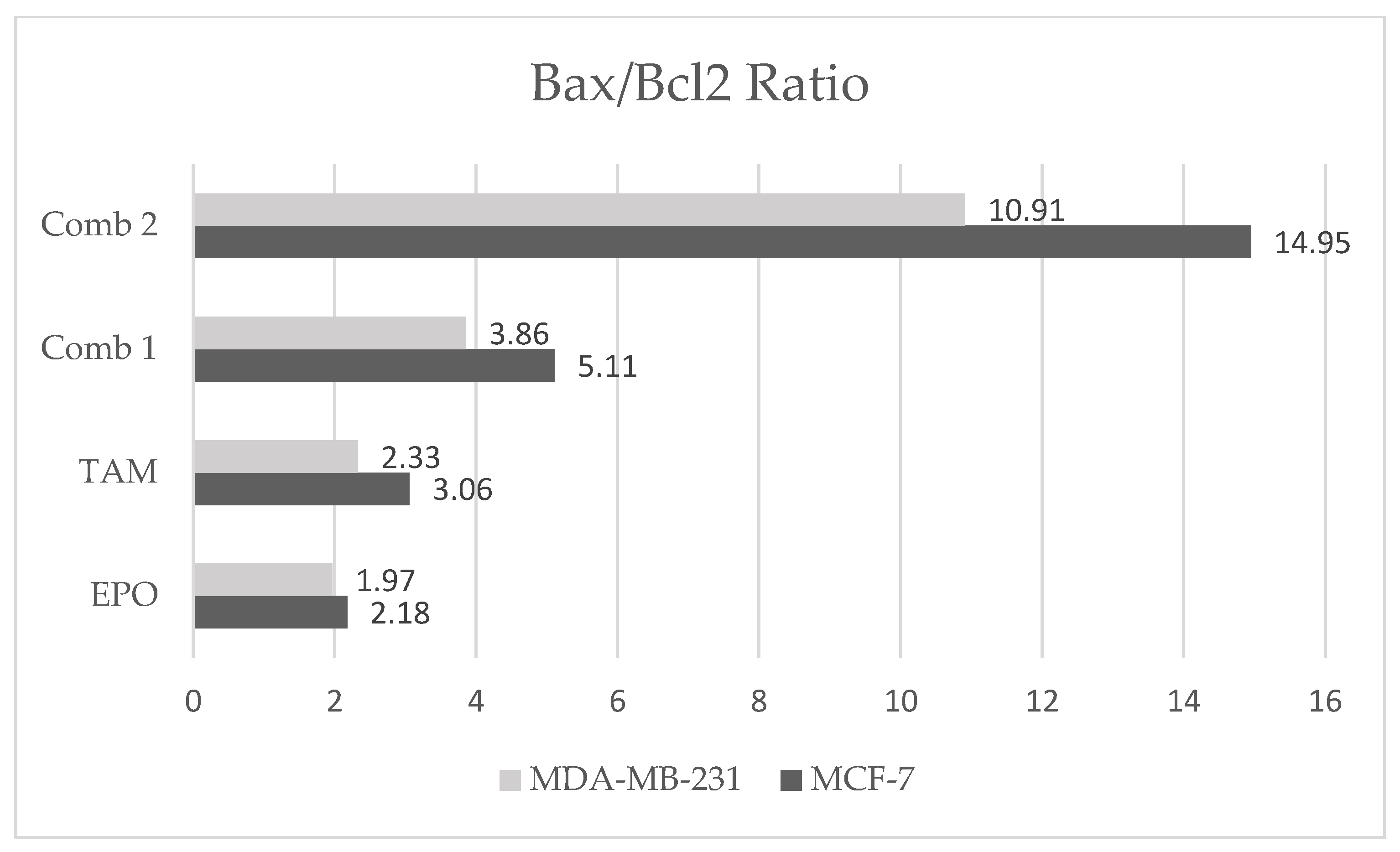
3.4. Effect of TAM and EPO, Alone and in Combination, on VEGF, Bcl-2, and Bax Gene Expression
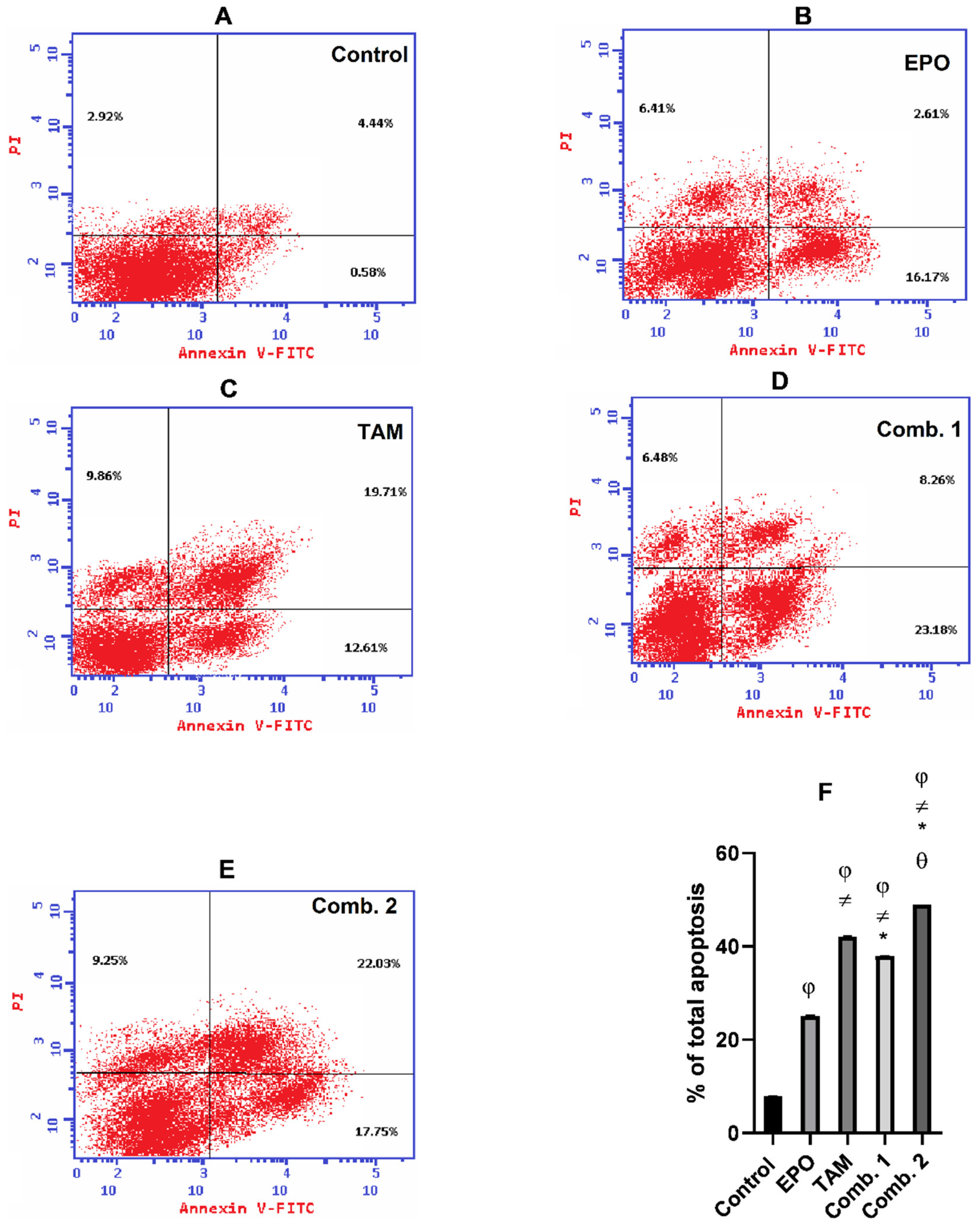
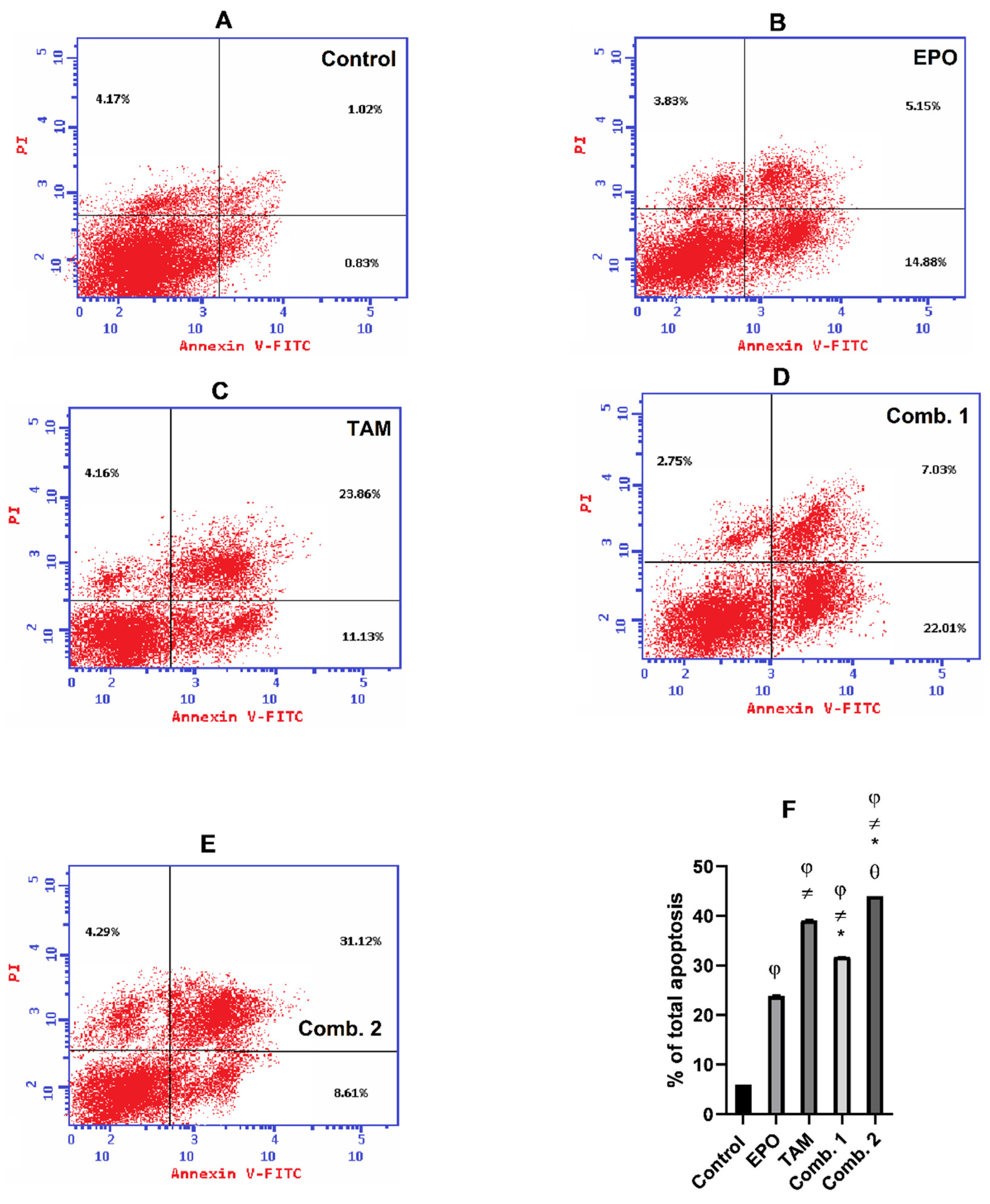
3.5. Effect of TAM and EPO, Alone or in Combination, on Apoptosis
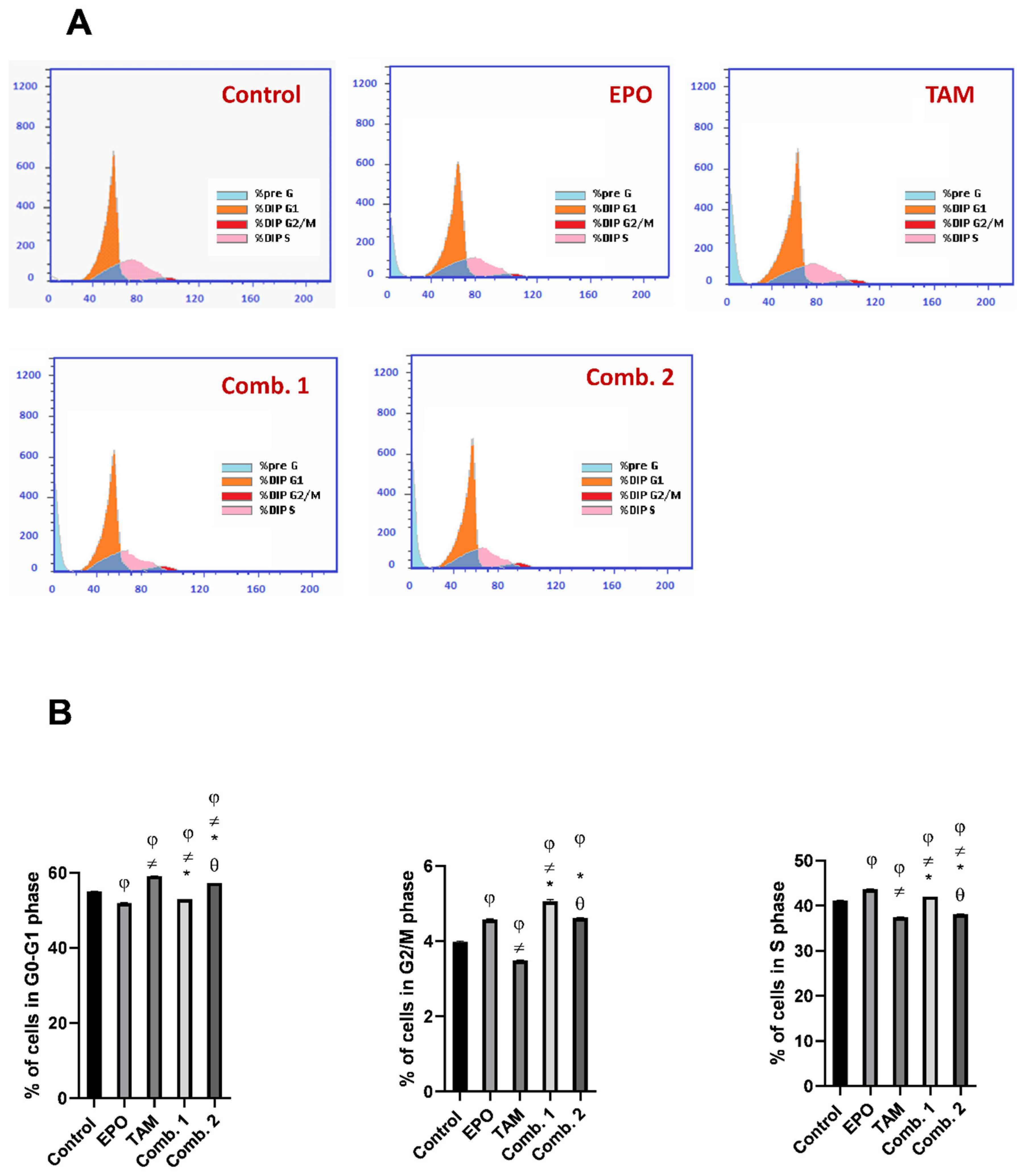
3.6. Effect of TAM and EPO, Alone or in Combination, on Cell Cycle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Hasan, S.A.W. Breast Cancer. World Health Organization WHO Regional Websites. 26 March 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer (accessed on 4 March 2022).
- Louzada, S.; Adega, F.; Chaves, R. Defining the sister rat mammary tumor cell lines HH-16 cl. 2/1 and HH-16. cl. 4 as an in vitro cell model for Erbb2. PLoS ONE 2012, 7, e29923. [Google Scholar] [CrossRef] [PubMed]
- Salahuddin, M.M.; Omran, G.A.; Helmy, M.W.; Houssen, M.E. Effect of Regorafenib on P2X7 Receptor Expression and Different Oncogenic Signaling Pathways in a Human Breast Cancer Cell Line: A Potential of New Insight of the Antitumor Effects of Regorafenib. Curr. Issues Mol. Biol. 2021, 43, 2199–2209. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yu, P.; Tang, J. Characterization of triple-negative breast cancer MDA-MB-231 cell spheroid model. OncoTargets Ther. 2020, 13, 5395. [Google Scholar] [CrossRef] [PubMed]
- Eeckhoute, J.; Carroll, J.S.; Geistlinger, T.R.; Torres-Arzayus, M.I.; Brown, M. A cell-type-specific transcriptional network required for estrogen regulation of cyclin D1 and cell cycle progression in breast cancer. Genes Dev. 2006, 20, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I. Tamoxifen a pioneering drug: An update on the therapeutic potential of tamoxifen derivatives. Eur. J. Med. Chem. 2018, 143, 515–531. [Google Scholar]
- Mangla, B.; Neupane, Y.R.; Singh, A.; Kohli, K. Tamoxifen and Sulphoraphane for the breast cancer management: A synergistic nanomedicine approach. Med. Hypotheses 2019, 132, 109379. [Google Scholar] [CrossRef]
- Clifford, R.; Bowden, D.; Blower, E.; Kirwan, C.C.; Vimalachandran, D. Does tamoxifen have a therapeutic role outside of breast cancer? A systematic review of the evidence. Surg. Oncol. 2020, 33, 100–107. [Google Scholar] [CrossRef]
- Khamis, A.A.; Ali, E.M.; Abd El-Moneim, M.A.; Abd-Alhaseeb, M.M.; El-Magd, M.A.; Salim, E.I. Hesperidin, piperine and bee venom synergistically potentiate the anticancer effect of tamoxifen against breast cancer cells. Biomed. Pharmacother. 2018, 105, 1335–1343. [Google Scholar] [CrossRef]
- Roberts, C.G.; Gurisik, E.; Biden, T.J.; Sutherland, R.L.; Butt, A.J. Synergistic cytotoxicity between tamoxifen and the plant toxin persin in human breast cancer cells is dependent on Bim expression and mediated by modulation of ceramide metabolism. Mol. Cancer Ther. 2007, 6, 2777–2785. [Google Scholar] [CrossRef] [Green Version]
- Bayles, B.; Usatine, R. Evening primrose oil. Am. Fam. Physician 2009, 80, 1405–1408. [Google Scholar]
- Atteia, H.H.; Alzahrani, S.; El-Sherbeeny, N.A.; Youssef, A.M.; Farag, N.E.; Mehanna, E.T.; Elhawary, R.; Ibrahim, G.A.; Elmistekawy, A.; Zaitone, S.A. Evening primrose oil ameliorates hyperleptinemia and reproductive hormone disturbances in obese female rats: Impact on estrus cyclicity. Front. Endocrinol. 2020, 10, 942. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.; Li, Y.; Luo, X.; Wang, X.; Wang, C.; Wen, B.; Guan, X.; Xu, Y.; Liu, B. Effect of the chemical refining process on composition and oxidative stability of evening primrose oil. J. Food Processing 2020, 44, e14800. [Google Scholar] [CrossRef]
- Timoszuk, M.; Bielawska, K.; Skrzydlewska, E. Evening Primrose (Oenothera biennis) Biological Activity Dependent on Chemical Composition. Antioxidants 2018, 7, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Shi, J.; Gong, L. Gamma linolenic acid suppresses hypoxia-induced gastric cancer cell growth and epithelial-mesenchymal transition by inhibiting the Wnt/b-catenin signaling pathway. Folia Histochem. Cytobiol. 2020, 58, 117–126. [Google Scholar] [CrossRef]
- El-Hanboshy, S.M.; Helmy, M.W.; Abd-Alhaseeb, M.M. Catalpol synergistically potentiates the anti-tumour effects of regorafenib against hepatocellular carcinoma via dual inhibition of PI3K/Akt/mTOR/NF-κB and VEGF/VEGFR2 signaling pathways. Mol. Biol. Rep. 2021, 48, 7233–7242. [Google Scholar] [CrossRef] [PubMed]
- Wirasnita, R.; Hadibarata, T.; Novelina, Y.M.; Yusoff, A.R.M.; Yusop, Z. A modified methylation method to determine fatty acid content by gas chromatography. Bull. Korean Chem. Soc. 2013, 34, 3239–3242. [Google Scholar] [CrossRef] [Green Version]
- Jaiyesimi, I.A.; Buzdar, A.U.; Decker, D.A.; Hortobagyi, G.N. Use of tamoxifen for breast cancer: Twenty-eight years later. J. Clin. Oncol. 1995, 13, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Vong, C.T.; Chen, H.; Gao, Y.; Lyu, P.; Qiu, L.; Zhao, M.; Liu, Q.; Cheng, Z.; Zou, J.; et al. Naturally occurring anti-cancer compounds: Shining from Chinese herbal medicine. Chin. Med. 2019, 14, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard-Gallon, D.J.; Satih, S.; Chalabi, N.; Rabiau, N.; Bosviel, R.; Fontana, L.; Bignon, Y.-J. Phytoestrogens regulate the expression of genes involved in different biological processes in BRCA2 knocked down MCF-7, MDA-MB-231 and MCF-10a cell lines. Oncol. Rep. 2010, 23, 647–653. [Google Scholar]
- Henriksson, E.; Kjellén, E.; Wahlberg, P.; Wennerberg, J.; Kjellström, J.H. Differences in estimates of cisplatin-induced cell kill in vitro between colorimetric and cell count/colony assays. Vitr. Cell. Dev. Biol. Anim. 2006, 42, 320–323. [Google Scholar] [CrossRef]
- Lewandowska, U.; Owczarek, K.; Szewczyk, K.; Sosnowska, D.; Koziołkiewicz, M.; Hrabec, E. Differentiated impact of procyanidins from evening primrose on human breast cancer cells. Cent. Eur. J. Biol. 2014, 9, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Miyake, J.A.; Gomes, R.N.; Colquhoun, A. Gamma-linolenic acid alters migration, proliferation and apoptosis in human and rat glioblastoma cells. Prostaglandins Other Lipid Mediat. 2020, 150, 106452. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.Y.; Xu, Y. Anti-cancer activities of ω-6 polyunsaturated fatty acids. Biomed. J. 2014, 37, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Sansone, A.; Ferreri, R.; Amézaga, J.; Tueros, I. Fatty Acids and Membrane Lipidomics in Oncology: A Cross-Road of Nutritional, Signaling and Metabolic Pathways. Metabolites 2020, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-S.; Huang, S.-M.; Wang, Y.-C. Palmitate Enhances the Efficacy of Cisplatin and Doxorubicin against Human Endometrial Carcinoma Cells. Int. J. Mol. Sci. 2021, 23, 80. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Ding, Y.; Wang, Y.; Wang, Z.; Yin, X.; Yan, G.; Zhang, L.; Yang, P.; Shen, H. Functional lipidomics: Palmitic acid impairs hepatocellular carcinoma development by modulating membrane fluidity and glucose metabolism. Hepatology 2017, 66, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Hardy, S.; Langelier, Y.; Prentki, M. Oleate activates phosphatidylinositol 3-kinase and promotes proliferation and reduces apoptosis of MDA-MB-231 breast cancer cells, whereas palmitate has opposite effects. Cancer Res. 2000, 60, 6353–6358. [Google Scholar]
- Hsiao, Y.-H.; Lin, C.-I.; Liao, H.; Chen, Y.-H.; Lin, S.-H. Palmitic acid-induced neuron cell cycle G2/M arrest and endoplasmic reticular stress through protein palmitoylation in SH-SY5Y human neuroblastoma cells. Int. J. Mol. Sci. 2014, 15, 20876–20899. [Google Scholar] [CrossRef] [Green Version]
- Baumann, J.; Wong, J.; Sun, Y.; Conklin, D.S. Palmitate-induced ER stress increases trastuzumab sensitivity in HER2/neu-positive breast cancer cells. BMC Cancer 2016, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Harada, H.; Yamashita, U.; Kurihara, H.; Fukushi, E.; Kawabata, J.; Kamei, Y. Antitumor activity of palmitic acid found as a selective cytotoxic substance in a marine red alga. Anticancer Res. 2003, 22, 2587–2590. [Google Scholar]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Orfanos, C.E.; Geilen, C.C.; Wieder, T.; Sturm, I.; Daniel, P.T. The Bax/Bcl-2 ratio determines the susceptibility of human melanoma cells to CD95/Fas-mediated apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowska, U.; Szewczyk, K.; Owczarek, K.; Hrabec, Z.; Podsedek, A.; Koziołkiewicz, M.; Hrabec, E. Flavanols from evening primrose (Oenothera paradoxa) defatted seeds inhibit prostate cells invasiveness and cause changes in Bcl-2/Bax mRNA ratio. J. Agric. Food Chem. 2013, 61, 2987–2998. [Google Scholar] [CrossRef] [PubMed]
- Riccardi, C.; Napolitano, E.; Platella, C.; Musumeci, D.; Melone, M.A.; Montesarchio, D. Anti-VEGF DNA-based aptamers in cancer therapeutics and diagnostics. Med. Res. Rev. 2021, 41, 464–506. [Google Scholar] [CrossRef]
- Carmeliet, P. VEGF as a key mediator of angiogenesis in cancer. Oncology 2005, 69, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Frezzetti, D.; Gallo, M.; Maiello, M.R.; D’Alessio, A.; Esposito, C.; Chicchinelli, N.; Normanno, N.; De Luca, A. VEGF as a potential target in lung cancer. Expert Opin. Ther. Targets 2017, 21, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Mohammadian, M.; Feizollahzadeh, S.; Mahmoudi, R.; Toofani Milani, A.; Rezapour-Firouzi, S.; Karimi Douna, B. Hsp90 Inhibitor; NVP-AUY922 in Combination with Doxorubicin Induces Apoptosis and Downregulates VEGF in MCF-7 Breast Cancer Cell Line. Asian Pac. J. Cancer Prev. 2020, 21, 1773–1778. [Google Scholar] [CrossRef]
- Ahlin, C.; Lundgren, C.; Embretsén-Varro, E.; Jirström, K.; Blomqvist, C.; Fjällskog, M.L. High expression of cyclin D1 is associated to high proliferation rate and increased risk of mortality in women with ER-positive but not in ER-negative breast cancers. Breast Cancer Res. Treat. 2017, 164, 667–678. [Google Scholar] [CrossRef]
- Shan, Y.S.; Hsu, H.P.; Lai, M.D.; Hung, Y.H.; Wang, C.Y.; Yen, M.C.; Chen, Y.L. Cyclin D1 overexpression correlates with poor tumor differentiation and prognosis in gastric cancer. Oncol. Lett. 2017, 14, 4517–4526. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| VEGF | AGGAGGAGGGCAGAATCATC | GGCACACAGGATGGCTTGAA |
| Bax | CCTGTGCACCAAGGTGCCGGAACT | CCACCCTGGTCTTGGATCCAGCCC |
| Bcl-2 | AGGAAGTGAACATTTCGGTGAC | GCTCAGTTCCAGGACCAGGC |
| β actin | CACCAACTGGGACGACAT | ACAGCCTGGATAGCAACG |
| Peak | Retention Time (min) | Name | Area % | Molecular Weight | Molecular Formula | Retention Index |
|---|---|---|---|---|---|---|
| 1 | 7.99 | Methyl tetradecanoate (Myristic acid, Methyl ester) | 0.23 | 242 | C15H30O2 | 1725 |
| 2 | 11.55 | Methyl hexadecanoate (Palmitic acid, Methyl ester) | 40.36 | 270 | C17H34O2 | 1926 |
| 3 | 14.81 | Methyl γ-linolenate (γ-Linolenic acid, methyl ester) | 7.32 | 292 | C19H32O2 | 2092 |
| 4 | 15.77 | Methyl linoleate (Linoleic acid, methyl ester) | 43.05 | 294 | C19H34O2 | 2092 |
| 5 | 16.25 | Methyl stearate (Stearic acid, Methyl ester) | 7.91 | 298 | C19H38O2 | 2128 |
| 6 | 17.75 | 6,9-Octadecadienoic acid (E, E), methyl ester | 0.72 | 294 | C19H34O2 | 2350 |
| 7 | 18.07 | 9,12-Octadecadienoic acid (Z, Z) (Linoleic acid) | 0.41 | 280 | C18H32O2 | 2133 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Alhaseeb, M.M.; Massoud, S.M.; Elsayed, F.; Omran, G.A.; Salahuddin, A. Evening Primrose Oil Enhances Tamoxifen’s Anticancer Activity against Breast Cancer Cells by Inducing Apoptosis, Inhibiting Angiogenesis, and Arresting the Cell Cycle. Molecules 2022, 27, 2391. https://doi.org/10.3390/molecules27082391
Abd-Alhaseeb MM, Massoud SM, Elsayed F, Omran GA, Salahuddin A. Evening Primrose Oil Enhances Tamoxifen’s Anticancer Activity against Breast Cancer Cells by Inducing Apoptosis, Inhibiting Angiogenesis, and Arresting the Cell Cycle. Molecules. 2022; 27(8):2391. https://doi.org/10.3390/molecules27082391
Chicago/Turabian StyleAbd-Alhaseeb, Mohammad M., Sarah M. Massoud, Fatma Elsayed, Gamal A. Omran, and Ahmad Salahuddin. 2022. "Evening Primrose Oil Enhances Tamoxifen’s Anticancer Activity against Breast Cancer Cells by Inducing Apoptosis, Inhibiting Angiogenesis, and Arresting the Cell Cycle" Molecules 27, no. 8: 2391. https://doi.org/10.3390/molecules27082391
APA StyleAbd-Alhaseeb, M. M., Massoud, S. M., Elsayed, F., Omran, G. A., & Salahuddin, A. (2022). Evening Primrose Oil Enhances Tamoxifen’s Anticancer Activity against Breast Cancer Cells by Inducing Apoptosis, Inhibiting Angiogenesis, and Arresting the Cell Cycle. Molecules, 27(8), 2391. https://doi.org/10.3390/molecules27082391