Abstract
Chromosome 3-specific NotI microarray (NMA) containing 180 clones with 188 genes was used in the study to analyze 18 high grade serous ovarian cancer (HGSOC) samples and 7 benign ovarian tumors. We aimed to find novel methylation-dependent biomarkers for early detection and prognosis of HGSOC. Thirty five NotI markers showed frequency of methylation/deletion more or equal to 17%. To check the results of NMA hybridizations several samples for four genes (LRRC3B, THRB, ITGA9 and RBSP3 (CTDSPL)) were bisulfite sequenced and confirmed the results of NMA hybridization. A set of eight biomarkers: NKIRAS1/RPL15, THRB, RBPS3 (CTDSPL), IQSEC1, NBEAL2, ZIC4, LOC285205 and FOXP1, was identified as the most prominent set capable to detect both early and late stages of ovarian cancer. Sensitivity of this set is equal to (72 ± 11)% and specificity (94 ± 5)%. Early stages represented the most complicated cases for detection. To distinguish between Stages I + II and Stages III + IV of ovarian cancer the most perspective set of biomarkers would include LOC285205, CGGBP1, EPHB1 and NKIRAS1/RPL15. The sensitivity of the set is equal to (80 ± 13)% and the specificity is (88 ± 12)%. Using this technique we plan to validate this panel with new epithelial ovarian cancer samples and add markers from other chromosomes.
1. Introduction
Epithelial ovarian cancer (EOC) remains a highly lethal malignancy. It is the fifth leading cause of cancer deaths among women in the United States (22,000 new cases and 16,000 deaths annually) and causes more than 140,000 deaths annually in women worldwide. Despite intensive research efforts over the past decade directed toward improved detection and treatment of ovarian cancer, the majority of women diagnosed with ovarian cancer die from the disease [1].
The epithelium is the tissue where most ovarian cancers arise [2]. Ovarian cancer is classified into several stages according to the American Joint Committee on Cancer/Tumor Node Metastasis (AJCC/TNM) and International Federation of Gynecology and Obstetrics (FIGO) staging systems which are based on how far the cancer has spread. In Stages I and II, the tumor is confined to the ovaries, while there is local metastasis (usually lymph) in Stage III and there is distal organ metastases in Stage IV [3].
Currently, the two principle obstacles in treating this life threatening disease are lack of effective biomarkers for early detection and drug resistance after initial chemotherapy.
Due to the atypical syndrome of the early stage of ovarian cancer, it is difficult to diagnose in its early stages. EOC (90% of ovarian cancer) is diagnosed at an advanced Stages III and IV in 75% of all cases, where the disease has spread throughout the abdomen. Patients with advanced stage disease have a 5-year survival of only 30% in contrast to early-stage disease (confined to the ovaries), where 5-year survival exceeds 80% [4].
Ovarian cancer is a heterogeneous disease both histologically and in patterns of disease progression. EOC is comprised of four major histologic subtypes: endometrioid, mucinous, clear cell and serous ovarian cancer. The high grade variant of serous ovarian cancer (HGSOC) is typically diagnosed in late stage, and is mostly responsible for the high lethality rate of ovarian cancer. It is also the subtype with the highest prevalence, estimated at ~70% of all cases [5,6].
Similar to other cancers, the initiation and development of ovarian cancer is characterized by activation of oncogenes and disruption of tumor suppressor genes (TSGs) by both genetic and epigenetic mechanisms. While it is well known that it is challenging to treat ovarian cancer through a genetic strategy due in part to its heterogeneity, the reversibility of epigenetic mechanisms involved in ovarian cancer opens exciting new avenues for treatment. The epigenomics of ovarian cancer has therefore become a rapidly expanding field leading to intense investigation.
Hypermethylation of CpG islands in gene promoter regions has been observed as a frequent mechanism associated with inactivation of TSGs which contributes to malignant transformation. As aberrant methylation is thought to be one of the earliest observable molecular changes in carcinogenesis, the detection of alterations in DNA methylation patterns has potential applicability to the detection of early-stage or pre-malignant disease [7,8]. Specific methylated DNA markers can be detected in the serum, plasma and peritoneal fluid of ovarian cancer patients [9].
Thus, cancer epigenetic studies hold great promise in revealing potent biomarkers for improved cancer detection [10,11]. Candidate gene and whole-genome studies have identified methylation signatures that may serve as biomarkers for EOC characterization including classification [7], progression [12] and response to therapy [13].
We constructed a new generation (~20 years ago) of lambda based cloning vectors [14]. These vectors opened new possibilities for gene cloning and analysis. Using these vectors we invented new approaches for construction of NotI linking and jumping libraries that have several advantages compared to previous techniques; they enabled efficient construction of such libraries representative and in plasmid form [15,16]. We experimentally confirmed that there is a direct association between CpG islands, NotI sites and expressed sequences in the human genome [17,18]. We constructed numerous linking libraries with different restriction enzymes in an attempt to generate representative NotI linking libraries, covering the whole human genome [19]. We generated more than 100,000 NotI flanking sequences and identified among them approximately 22,000 unique NotI sequences comprising 17 Mb information [20,21]. It was believed at this time that human genome contains only 3100 NotI sites and we showed for the first time that there are more than 10,000 NotI sites.
With this sequencing information we suggested to construct and to use NotI microarrays (NMA, see Figure 1) for comparison of normal and malignant cells [22]. Here we present our results of analysis using NMA of 18 HGSOC and 7 benign ovarian adenomas (BOA) and developed a set of novel epigenetic biomarkers for early detection and diagnosis of EOC.
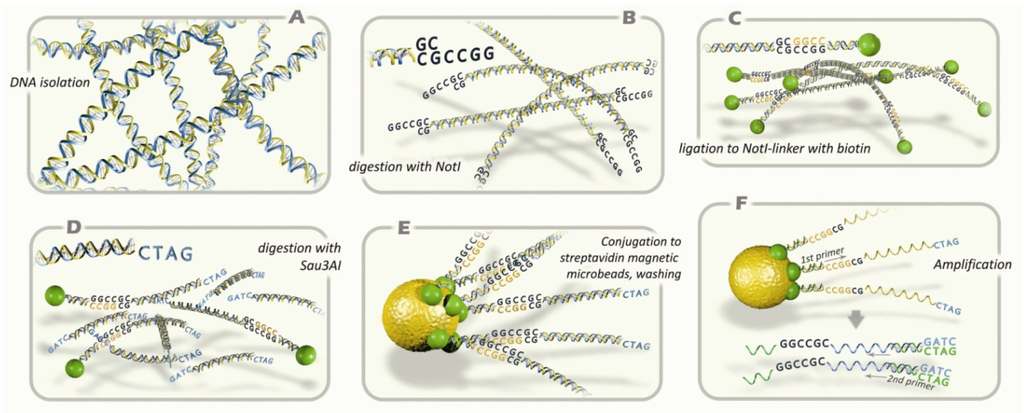
Figure 1.
Principal scheme of NotI microarray analysis protocol. (A) Isolation of genomic DNA; (B) digestion with methyl-specific rare-cutter enzyme NotI; (C) ligation of fragments with NotI-linker containing biotin; (D) digestion with 4-base pair recognizing restriction enzyme Sau3AI; (E) conjugation to microbeads containing streptavidin; washing; (F) amplification of DNA sequences that has been attached to microbeads. The standard procedures are performed: microarray hybridization, cloning, and sequencing analysis.
2. Results
2.1. Analysis of Methylation Frequency Using NotI Microarrays
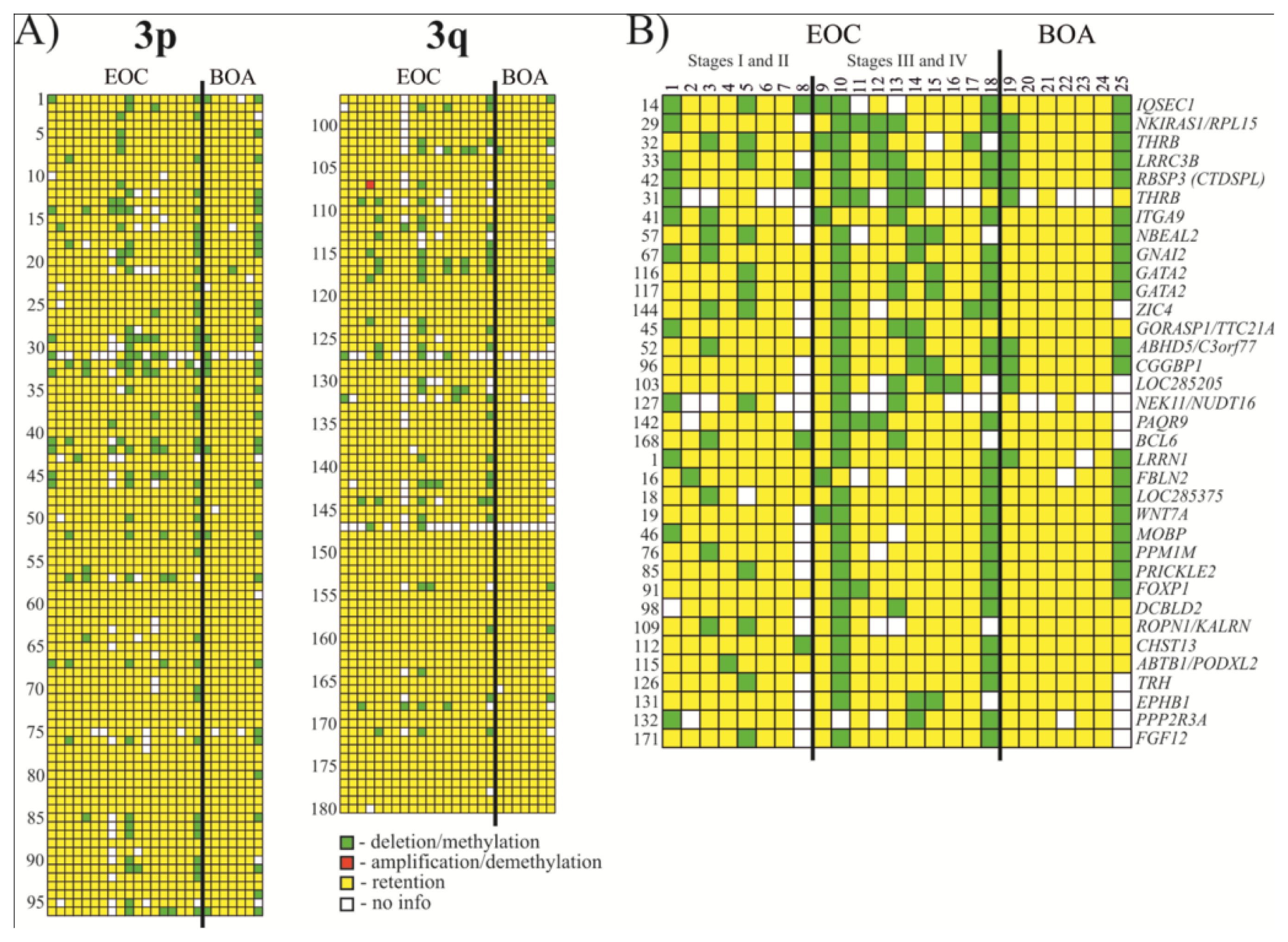
Thirty five NotI markers showed frequency of methylation/deletion more or equal to 17% (see Figure 2 and Tables 1 and S1). Among the most affected genes were IQSEC1, NKIRAS1/RPL15, THRB, LRRC3B and RBSP3 (CTDSPL) that showed 33% methylation/deletion (up to 38% when not counting samples with “no information”).
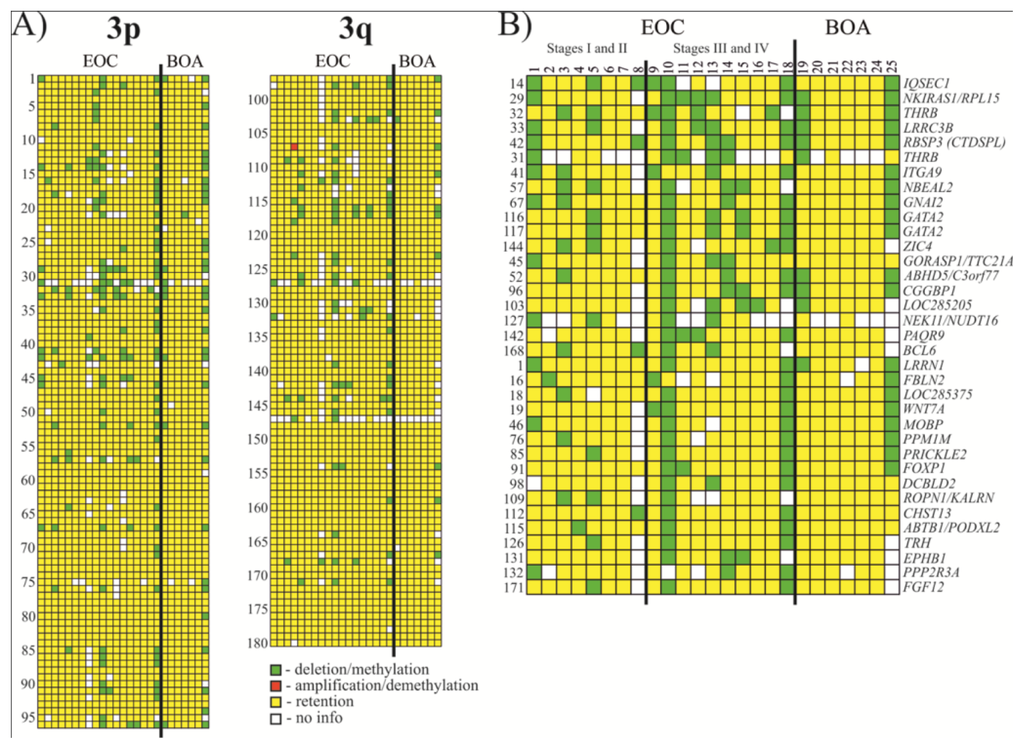
Figure 2.
Hybridization pattern of DNA from Epithelial ovarian cancer (EOC) and benign ovarian adenomas (BOA) samples on NotI-microarrays. (A) Vertically, 180 NotI sites arranged according to their localization on chromosome 3 (from 3p26.2 to 3p11.1 and from 3q11.2 to 3q29). Horizontally, 25 ovarian samples (18 EOC and 7 BOA); (B) Vertically, 35 NotI sites arranged by methylation/deletion frequency (from 33% to 17%).

Table 1.
Methylation/deletion frequencies for 35 genes with the highest percent of changes in ovarian cancer.
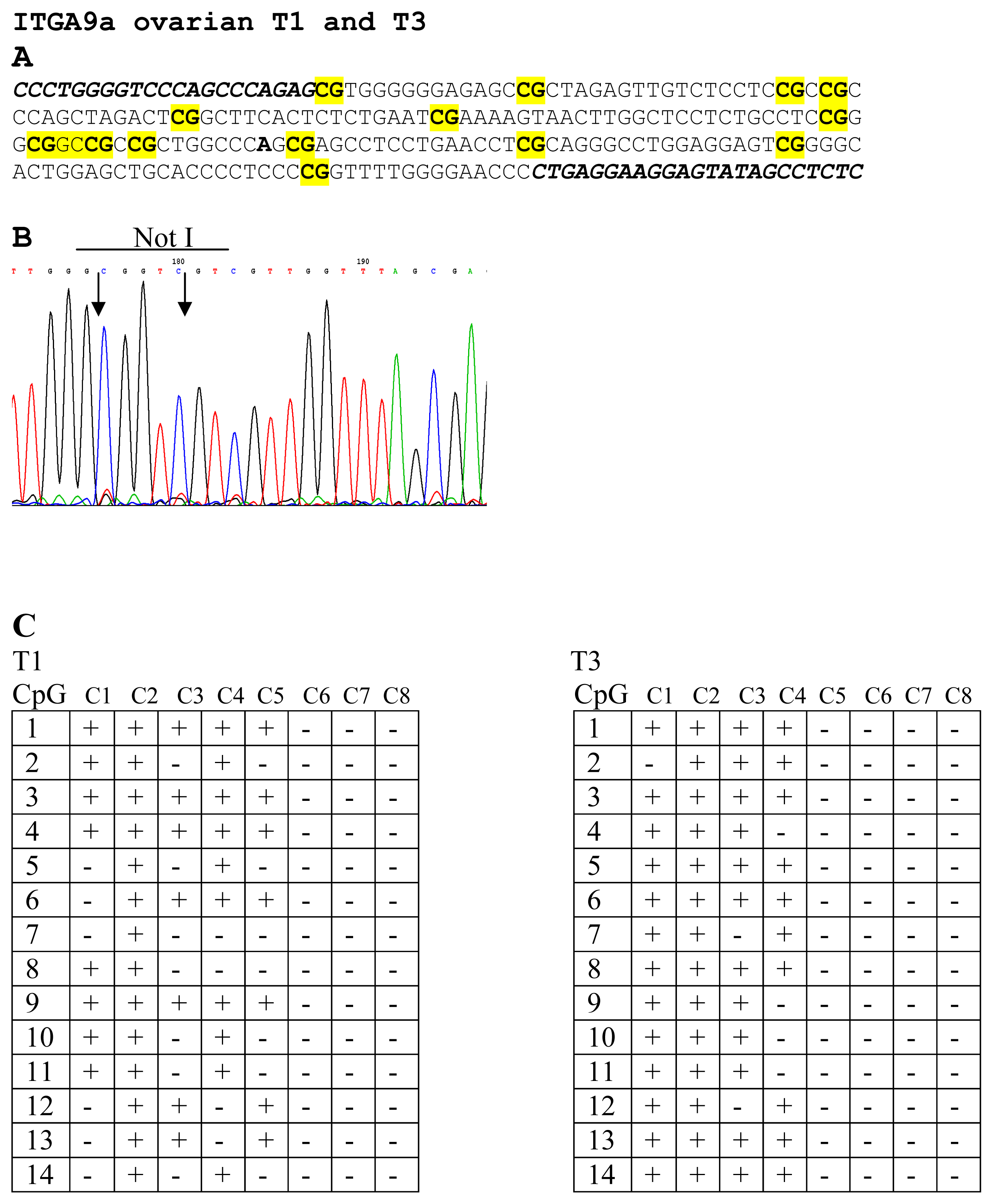
To prove the results of NMA hybridizations, several samples were sequenced. For genes LRRC3B (No. 12 and 13) and THRB (No. 9 and 10) were selected two samples and both of them were found methylated. For ITGA9 three samples were selected (No. 1, 2 and 3) and only sample No. 2 was found unmethylated. For RBSP3 (CTDSPL) three samples were also selected (No. 14, 15 and 16) and as it follows from the Figure 1B only one sample was found to be methylated (No. 14). Thus results of bisulfite sequencing confirmed the results of NMA (see Figure 3).
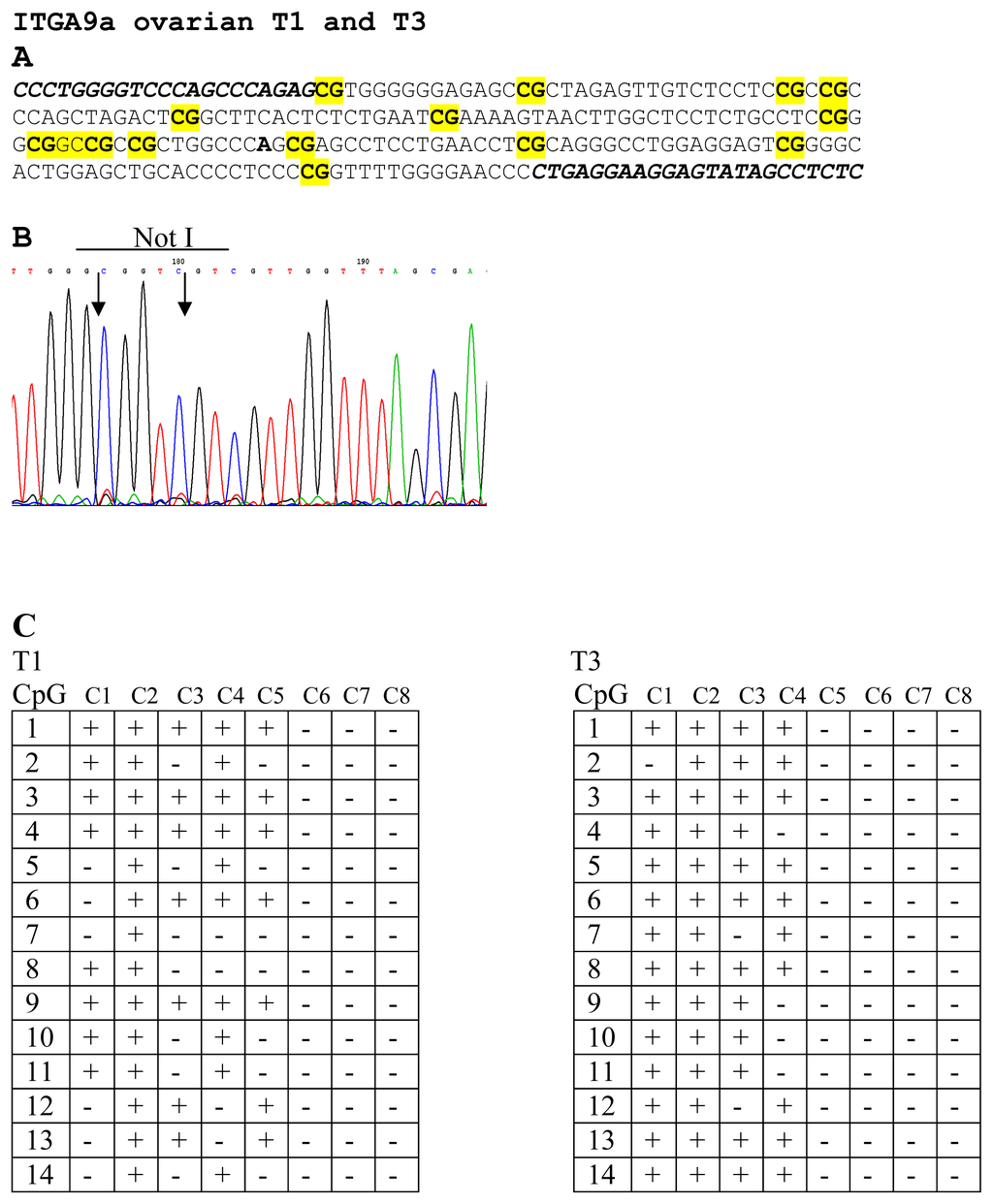
Figure 3.
Bisulfite sequencing of ITGA9 in EOC samples. CG-pairs containing methylated cytosine are shown in bold and yellow (A). Primers for bisulfite sequencing (A) are shown in italics below example of sequencing diagrams (B) demonstrating methylated sequence of ITGA9 is shown. In the two tables (C) methylated (+) and unmethylated (−) CG pairs are shown in eight sequenced clones for T1 and T3 samples.
Eight genes showed the tendency to increase methylation/deletion frequency during ovarian cancer progression (stages III + IV relative to stages I + II, see Table 2).
Table 2.
Methylation/Deletion frequency for eight genes in two groups of samples.
2.2. Selection of Genes/Biomarkers for Detection and Discrimination EOC with Different Histological Characteristics
For detection cancer in ovarian biopsy on all stages including early one of the most perspective set from analyzed genes included 8 biomarkers: NKIRAS1/RPL15, THRB, RBPS3 (CTDSPL), IQSEC1, NBEAL2, ZIC4, LOC285205 and FOXP1. If methylation/deletion was found in two or more of these biomarkers then sample would be recognized as cancer. Sensitivity of this set is equal (72 ± 11)% and specificity (94 ± 5)%. Early stages represented the most complicated cases for detection.
BOA samples had no changes in five cases from seven analyzed, so in order to distinguish them from cancer samples it is possible to use the same set as for cancer detection. If methylation/deletion was found in two or more of the above-mentioned biomarkers then the sample would be recognized as cancer. Sensitivity of this set is equal (72 ± 11)% and specificity (71 ± 17)%.
To distinguish between Stages I + II and Stages III + IV of ovarian cancer the most perspective set would include LOC285205, CGGBP1, EPHB1, and NKIRAS1/RPL15 biomarkers. If we found methylation/deletion in 1 or more of these biomarkers then sample would be recognized as a sample from III + IV stages. In this case the sensitivity of the set is equal to (80 ± 13)% and the specificity is (88 ± 12)%. Stages III + IV methylation/deletion assumed as positive result and Stages I + II as negative.
In summary, the suggested set of 10 markers (NKIRAS1/RPL15, THRB, RBPS3 (CTDSPL), IQSEC1, NBEAL2, ZIC4, LOC285205, FOXP1, CGGBP1, EPHB1) would allow us to discriminate/diagnose the majority of EOC cases with sensitivity and specificity higher than 71% (up to 94%) (Tables 3 and 4).
Table 3.
Early detection and discrimination of ovarian cancer groups with different histological characteristics using the set of 10 selected markers.
Table 4.
Annotations for gene markers involved in ovarian cancer and their protein products.
3. Discussion
In this work we used chromosome 3 specific NotI-microarrays with 180 clones containing 188 genes to analyze 18 HGSOC and 7 BOA samples. The main idea of the approach is that NotI enzyme cuts only unmethylated CpG pairs inside the recognition site of the enzyme (5′-GCGGCCGC-3′) and only small fraction (0.1%–0.05%) of the human genome containing NotI digested fragments is labeled. The NR probes were prepared as described earlier (see Materials and Methods and [84,103–105]). Thus, in contrast to all other methods where undigested by methylation sensitive enzymes DNA fragments are labeled, we label only digested DNA fragments. As a consequence, our probe contains 10-fold less repeats, it is hotter, not very sensitive to incomplete digestion and gives less background. To confirm results of NMA hybridizations, several samples for four genes, namely, LRRC3B, THRB, ITGA9 and RBSP3 (CTDSPL) were sequenced (see Figure 3). All the genes were found to be methylated. It is important to note that for bisulfite sequencing we always clone PCR product and then sequence 8 clones. Of course, we could not exclude the possibility that unmethylated clones, i.e., green in Figure 2, were in fact deleted. However, this issue can be considered as an advantage when search for TSGs is being performed: TSGs are inactivated either by methylation or deletions, or both of them, and we can detect these events simultaneously. If it is necessary, discrimination between these changes can be done using e.g., qPCR, bisulfite sequencing, and using another NR probe prepared with tumor DNA after amplification with Phi29 DNA polymerase.
This study clearly demonstrated that NotI-microarrays are powerful tools to find methylated genes and resulted in identification of many novel genes/biomarkers that can be important for the development of more specific biomarker sets for early diagnosis and new approaches to therapy of EOC.
As it was mentioned in the Introduction the major problems in HGSOC are its early diagnosis, discrimination between Stages I + II and Stages III + IV and absence of specific molecular markers.
For example, ovarian cancer screening with transvaginal ultrasound (TVU) and CA125 was evaluated in the Prostate, Lung, Colorectal, and Ovarian (PLCO) trials. However, it was revealed that the predictive value of both tests was relatively low [106]. Of 39,115 women randomized to receive screening, 28,816 received at least 1 test. Abnormal TVU was found in 1338 (4.7%), and abnormal CA-125 in 402 (1.4%). Twenty-nine neoplasms were identified (26 ovarian, 2 fallopian, and 1 primary peritoneal neoplasm). Nine were tumors of low malignant potential and 20 were invasive. The positive predictive value for invasive cancer was 3.7% for an abnormal CA-125, 1.0% for an abnormal TVU, and 23.5% if both tests were abnormal. The authors concluded that nothing in the findings reported in the paper suggests a need to revise the present (1996) ovarian cancer screening guidelines of the US Preventive Services Task Force, which state “routine screening for ovarian cancer by ultrasound, the measurement of serum tumor markers, or pelvic examination is not recommended.”
Increasing evidences indicate that epigenetic mechanisms may play a major role in the development of ovarian cancer [2]. Aberrant DNA methylation occurs commonly in tumors and is considered to be one of the earliest molecular changes in carcinogenesis [8,107–110]. Furthermore, studies have identified tumor-specific gene methylation in blood DNA of patients with EOC [9,111–113], indicating that methylation patterns in plasma DNA have potential to serve as non-invasive biomarkers of EOC.
Thus, Caceres et al. [9] using the panel of 6 TSGs: BRCA1, RASSF1A, APC, p14ARF, p16INK4A and DAP-Kinase, demonstrated that promoter hypermethylation is common in ovarian cancer, including stage I disease, and can be readily detected in a specific manner in serum and peritoneal fluid DNA. In this initial feasibility study, they observed a sensitivity of 82% and 100% specificity in serum. However, hypermethylation was observed in all histologic cell types, grades, and stages of ovarian tumor examined and moreover TSGs that were used in the study were not ovarian cancer specific.
In the study by Melnikov et al. [114] previously developed microarray-based technique was used; authors evaluated differences in DNA methylation profiles in a panel of 56 genes using sections of serous papillary adenocarcinomas and uninvolved ovaries (n = 30) from women in a high-risk group. Methylation profiles were also generated for circulating DNA from blood of patients (n = 33) and healthy controls (n = 33). Using the most differentially methylated genes for naïve Bayesian analysis, they identified 10 of these profiles as potentially informative in tissues. Various combinations of these genes produced 69% sensitivity and 70% specificity for cancer detection as estimated under a stratified, fivefold cross-validation protocol. In plasma, five genes were identified as informative; their combination had 85% sensitivity and 61% specificity for cancer detection. These results suggest that differential methylation profiling in heterogeneous samples has the potential to identify components of a composite biomarker that may detect ovarian cancer in blood with significant accuracy.
Ten of the most well-known methylated genes in ovarian cancer were selected in Montavon et al., 2012 study: BRCA1, CDH1, DLEC1, EN1, GATA4, GATA5, HOXa9, HSULF1; RASSF1A and SFN. Although some of them showed very high frequency of methylation in HGSOC (e.g., SFN 100%) and HOXa9 and EN1 showed sensitivity of 98.8% and specificity of 91.7%, none of the markers after correction for multiple testing, gene methylation was not significantly associated with any clinico pathological characteristics including discrimination of Stages I + II from Stages III + IV [6].
It has become clear that even with the great promise of DNA methylation biomarkers in epithelial ovarian cancer, the identification of highly specific, sensitive and robust panels of markers and the standardization of analysis techniques are still required in order to improve detection, treatment and thus patients’ outcome [115].
Among 11 genes included in our set there are six genes for which no information about their involvement in ovarian carcinogenesis have been shown: NKIRAS1/RPL15, THRB, IQSEC1, NBEAL2 and ZIC4 (see Table 4). However, for all 11 genes in the set it was shown involvement in some of cancers. In some cases it was shown only decreased expression (LOC285205, EPHB1 and RBSP3 (CTDSPL)) and in other cases loss of heterozygosity and copy number changes (CGGBP1 and RBSP3 (CTDSPL)) in ovarian cancer. Some other genes showed decreased expression in other cancer types, like EPHB1 in gastric carcinoma, NKIRAS1 in kidney cancer, THRB in many cancers, etc. FOXP1 is found to be significantly down-regulated in stage III serous ovarian carcinoma (see Table 4). Among two genes from the Table 2 not included in the set is the well-known tumor suppressor WNT7A. WNT (Wingless-Type Mouse mammary tumor virus Integration Site Family) growth factors have diverse roles in governing cell fate, proliferation, migration, polarity, and death in multicellular organisms. WNT7A has been demonstrated to be a TSG in lung cancer. Normally WNT7A maintains epithelial differentiation and inhibits growth of the transformed cell in a subset of human Non-Small Cell Lung Cancer (NSCLC). It was shown that WNT7A regulates tumor growth and progression in ovarian cancer. GATA2 is also involved in the development of bladder and breast cancer and acute myelogenous leukemia. Other genes were also shown to be involved in the process of carcinogenesis (see Table 4). Among the genes represented in the set there are genes encoding transcriptional regulators (CGGBP1, GATA2), receptors (EPHB1, THRB), phosphatase (RBSP3 (CTDSPL)), proteins interacting with other proteins (NKIRAS1, IQSEC1, NBEAL2, ZIC4).
These proteins are involved in different cancer-related pathways:
- EPHB1 for example participates in the Ephrin-EphR Signaling Pathway. In this pathway members of RAS family, MAPK, oncogene NCK1 and other genes important for cell developmental processes are included. Ephrin receptors make up the largest subgroup of the receptor tyrosine kinase (RTK) family. The protein encoded by this gene is a receptor for ephrin-B family members.
- NKIRAS1 is a potent regulator of NF-κ-B activity by preventing the degradation of NF-κ-B inhibitor beta (NFKBIB) by most signals, explaining why NFKBIB is more resistant to degradation.
- IQSEC1 links epidermal growth factor receptor signaling to ARF6 activation to induce breast cancer invasion (see Table 4).
- RBSP3 (CTDSPL) is a potential activator of the RB1 gene pathway [81].
At the same time little is known about function and involvement of LOC285205 and CGGBP1 genes in carcinogenesis. Thus this novel set of markers open perspectives for further improvement in early detection and diagnosis of EOC. Of course, this set needs further validation and testing with new samples of ovarian cancer including blood samples. Also this panel should be enriched with additional markers from other chromosomes; we are planning to construct NMA containing 10,000–15,000 genes.
4. Materials and Methods
4.1. Tissue Specimens
Eighteen paired specimens of epithelial ovarian carcinoma (all the samples were HGSOC) and seven cases of benign ovarian adenoma (BOA) were obtained after surgical resection of primary EOC or adenoma prior radiation or chemotherapy and stored in liquid nitrogen. “Normal” controls were obtained minimum at 2 cm distance from the tumor and confirmed histologically as normal ovarian epithelial cells. The diagnosis was verified by histopathology and only samples containing 70%–80% or more tumor cells were used in the study. The samples were collected in accordance to the guidelines issued by the Ethics Committee of Blokhin Cancer Research Center, Russian Academy of Medical Sciences (Moscow). All patients gave written informed consent that is available upon request. The Ethics Committee of Blokhin Cancer Research Center, Russian Academy of Medical Sciences specifically approved this study. The study was done in accordance with the principles outlined in the Declaration of Helsinki. All tumor specimens were characterized according to the International System of Clinico-Morphological Classification of Tumors, based on the tumor-node-metastasis (TNM) and staging classification of 2002 and AJCC/TNM criteria classification of 1999 [3].
4.2. NotI-Microarrays
One hundred and eighty NotI linking clones from human chromosome 3 containing 188 genes [22,116] with inserts up to 15 kb were immobilized on the glass slides in six replications. Plasmid DNA for immobilization on the glasses was isolated with a HiPure Plasmid Midiprep kit (Invitrogen) and printed on the silanized glasses at a concentration of 0.25 μg/μL with a QarrayMini microarrayer (Genetix, UK). DNA from E. coli was used as negative hybridization control.
4.3. NotI Probes and Hybridization
Preparation of NotI representations (NotI probes, NR, see Figure 1) with special labeling procedure, where only sequences surrounding NotI sites were labeled (0.1%–0.05% of the total human genomic DNA) was done using paired normal (adjacent control) and tumor DNA essentially as described previously [84,103–105]. In brief, this involved DNA digestion with NotI restriction enzyme, ligation to NotI-linkers with biotin, digestion with Sau3A restriction enzyme, immobilization on Dynabeads M-280 Streptavidin “Dynal” and finally washing and ligation to DNA bound with magnetic beads with Sau3A-linkers. The enriched DNA was amplified by PCR using universal and linker-primers. PCR conditions were the following: 2 min at 95 °C, then 35 cycles of denaturation (45 s at 95 °C), annealing (40 s at 64 °C) and synthesis (2 min 20 s at 72 °C). Thereafter, 200–400 ng of NR was labelled by PCR as described above but in the presence of 1.25 nM of Cy5-dCTP (or Cy3-dCTP). This probe detected both deleted/amplified and methylated/unmethylated sequences. If it is necessary, discrimination between these changes can be done using qPCR, bisulfite sequencing, using another NR probe prepared with tumor DNA after amplification with Phi29 DNA polymerase, etc. It is clear from Figure 1 that deleted NotI sites will give no signal, the same as methylated, i.e., will be green whereas amplified DNA would give stronger red signal.
Hybridization of coupled NotI samples was carried out at 42 °C for 15 h in a Lucidea Base device (Amersham Pharmacia Biotech) according to manufacturer’s recommendations. Microarrays were scanned in a GenePix 4000A. The results were processed with GenePix Pro 6.0 software (Amersham Pharmacia Biotech). Then data were analyzed using our program NIMAN (NotI-Microarray ANalysis, see [117]).
4.4. Statistical Analysis
Fisher’s exact test and χ2 criteria were used for analysis of methylation changes in ovarian cancer groups with different histological characteristics. p-Values < 0.05 were considered as statistically significant. All statistical procedures were performed using NIMAN [117] and BioStat software [118]. Sensitivity was calculated as the proportion of true positives that were correctly identified by the set. Specificity was calculated as the proportion of true negatives that were correctly identified by the set [119].
4.5. PCR, Cloning, Bisulfite Sequencing
PCR, cloning, bisulfite sequencing were done as described earlier [120].
5. Conclusions
We selected novel epigenetic markers including NKIRAS1/RPL15, THRB RBPS3 (CTDSPL), IQSEC1, NBEAL2, ZIC4, LOC285205, CGGBP1, EPHB1 and FOXP1 that allowed detection of cancer in ovarian biopsies on all stages with sensitivity equal to (72 ± 11)% and specificity (94 ± 5)%. This set allowed us to discriminate between Stages I+II and III+IV with sensitivity equal to (80 ± 13)% and specificity (88 ± 12)%. We will confirm this with a validation set of new EOC samples and add markers from other chromosomes. Among eleven genes included in our set, there are six genes for which no information about their involvement in ovarian carcinogenesis have been shown: NKIRAS1/RPL15, THRB, IQSEC1, NBEAL2 and ZIC4. For the 5 remaining genes except FOXP1 only slightly relevant information was published.
Acknowledgements
This work was supported by research grants from the Swedish Cancer Society, the Swedish Institute, the Swedish Research Council and Karolinska Institute, State Contracts 02.740.11.5227 and 16.552.11.7034 with the Russian Ministry of Education and Science, grants 10-04-01213-a and 11-04-00269 from the Russian Foundation for Basic Research.
- Conflict of interestThe authors declare no conflict of interest.
References
- Hunn, J.; Rodriguez, G.C. Ovarian cancer: Etiology, risk factors, and epidemiology. Clin. Obstet. Gynecol 2012, 55, 3–23. [Google Scholar]
- Chen, H.; Hardy, T.M.; Tollefsbol, T.O. Epigenomics of ovarian cancer and its chemoprevention. Front. Genet 2011, 2, 67. [Google Scholar]
- Yarbro, J.W.; Page, D.L.; Fielding, L.P.; Partridge, E.E.; Murphy, G.P. American Joint Committee on Cancer prognostic factors consensus conference. Cancer 1999, 86, 2436–2446. [Google Scholar]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin 2010, 60, 277–300. [Google Scholar]
- Landen, C.N., Jr; Birrer, M.J.; Sood, A.K. Early events in the pathogenesis of epithelial ovarian cancer. J. Clin. Oncol 2008, 26, 995–1005. [Google Scholar]
- Montavon, C.; Gloss, B.S.; Warton, K.; Barton, C.A.; Statham, A.L.; Scurry, J.P.; Tabor, B.; Nguyen, T.V.; Qu, W.; Samimi, G.; et al. Prognostic and diagnostic significance of DNA methylation patterns in high grade serous ovarian cancer. Gynecol. Oncol 2012, 124, 582–588. [Google Scholar]
- Barton, C.A.; Hacker, N.F.; Clark, S.J.; O’Brien, P.M. DNA methylation changes in ovarian cancer: Implications for early diagnosis, prognosis and treatment. Gynecol. Oncol 2008, 109, 129–139. [Google Scholar]
- Jones, P.A.; Baylin, S.B. The fundamental role of epigenetic events in cancer. Nat. Rev. Genet 2002, 3, 415–428. [Google Scholar]
- Ibanez de Caceres, I.; Battagli, C.; Esteller, M.; Herman, J.G.; Dulaimi, E.; Edelson, M.I.; Bergman, C.; Ehya, H.; Eisenberg, B.L.; Cairns, P. Tumor cell-specific BRCA1 and RASSF1A hypermethylation in serum, plasma, and peritoneal fluid from ovarian cancer patients. Cancer Res 2004, 64, 6476–6481. [Google Scholar]
- Kagan, J.; Srivastava, S.; Barker, P.E.; Belinsky, S.A.; Cairns, P. Towards Clinical Application of Methylated DNA Sequences as Cancer Biomarkers: A Joint NCI’s EDRN and NIST Workshop on Standards, Methods, Assays, Reagents and Tools. Cancer Res 2007, 67, 4545–4549. [Google Scholar]
- Laird, P.W. The power and the promise of DNA methylation markers. Nat. Rev. Cancer 2003, 3, 253–266. [Google Scholar]
- Wei, S.H.; Chen, C.M.; Strathdee, G.; Harnsomburana, J.; Shyu, C.R.; Rahmatpanah, F.; Shi, H.; Ng, S.W.; Yan, P.S.; Nephew, K.P.; et al. Methylation microarray analysis of late-stage ovarian carcinomas distinguishes progression-free survival in patients and identifies candidate epigenetic markers. Clin. Cancer Res 2002, 8, 2246–2252. [Google Scholar]
- Wei, S.H.; Balch, C.; Paik, H.H.; Kim, Y.S.; Baldwin, R.L.; Liyanarachchi, S.; Li, L.; Wang, Z.; Wan, J.C.; Davuluri, R.V.; et al. Prognostic DNA methylation biomarkers in ovarian cancer. Clin. Cancer Res 2006, 12, 2788–2794. [Google Scholar]
- Zabarovsky, E.R.; Klein, G.; Winberg, G. Lambda SK diphasmids: phage lambda vectors for genomic, jumping, linking and cDNA libraries. Gene 1993, 127, 1–14. [Google Scholar]
- Zabarovsky, E.R.; Boldog, F.; Erlandsson, R.; Kashuba, V.I.; Allikmets, R.L.; Marcsek, Z.; Kisselev, L.L.; Stanbridge, E.; Klein, G.; Sumegi, J.; et al. New strategy for mapping the human genome based on a novel procedure for construction of jumping libraries. Genomics 1991, 11, 1030–1039. [Google Scholar]
- Zabarovsky, E.R.; Boldog, F.; Thompson, T.; Scanlon, D.; Winberg, G.; Marcsek, Z.; Erlandsson, R.; Stanbridge, E.J.; Klein, G.; Sumegi, J. Construction of a human chromosome 3 specific NotI linking library using a novel cloning procedure. Nucleic Acids Res 1990, 18, 6319–6324. [Google Scholar]
- Allikmets, R.L.; Kashuba, V.I.; Pettersson, B.; Gizatullin, R.; Lebedeva, T.; Kholodnyuk, I.D.; Bannikov, V.M.; Petrov, N.; Zakharyev, V.M.; Winberg, G.; et al. NotI linking clones as a tool for joining physical and genetic maps of the human genome. Genomics 1994, 19, 303–309. [Google Scholar]
- Wang, J.Y.; Zabarovsky, E.R.; Talmadge, C.; Berglund, P.; Chan, K.W.; Pokrovskaya, E.S.; Kashuba, V.I.; Zhen, D.K.; Boldog, F.; Zabarovskaya, V.I.; et al. Somatic cell hybrid panel and NotI linking clones for physical mapping of human chromosome 3. Genomics 1994, 20, 105–113. [Google Scholar]
- Zabarovsky, E.R.; Allikmets, R.; Kholodnyuk, I.; Zabarovska, V.I.; Paulsson, N.; Bannikov, V.M.; Kashuba, V.I.; Dean, M.; Kisselev, L.L.; Klein, G. Construction of representative NotI linking libraries specific for the total human genome and for human chromosome 3. Genomics 1994, 20, 312–316. [Google Scholar]
- Kutsenko, A.S.; Gizatullin, R.Z.; Al-Amin, A.N.; Wang, F.; Kvasha, S.M.; Podowski, R.M.; Matushkin, Y.G.; Gyanchandani, A.; Muravenko, O.V.; Levitsky, V.G.; et al. NotI flanking sequences: A tool for gene discovery and verification of the human genome. Nucleic Acids Res 2002, 30, 3163–3170. [Google Scholar]
- Zabarovsky, E.R.; Gizatullin, R.; Podowski, R.M.; Zabarovska, V.V.; Xie, L.; Muravenko, O.V.; Kozyrev, S.; Petrenko, L.; Skobeleva, N.; Li, J.; et al. NotI clones in the analysis of the human genome. Nucleic Acids Res 2000, 28, 1635–1639. [Google Scholar]
- Li, J.; Protopopov, A.; Wang, F.; Senchenko, V.; Petushkov, V.; Vorontsova, O.; Petrenko, L.; Zabarovska, V.; Muravenko, O.; Braga, E.; et al. NotI subtraction and NotI-specific microarrays to detect copy number and methylation changes in whole genomes. Proc. Natl. Acad. Sci. USA 2002, 99, 10724–10729. [Google Scholar]
- Lind, G.E.; Skotheim, R.I.; Fraga, M.F.; Abeler, V.M.; Esteller, M.; Lothe, R.A. Novel epigenetically deregulated genes in testicular cancer include homeobox genes and SCGB3A1 (HIN-1). J. Pathol 2006, 210, 441–449. [Google Scholar]
- Singh, U.; Roswall, P.; Uhrbom, L.; Westermark, B. CGGBP1 regulates cell cycle in cancer cells. BMC Mol. Biol 2011, 12, 28. [Google Scholar]
- Cody, N.A.; Shen, Z.; Ripeau, J.S.; Provencher, D.M.; Mes-Masson, A.M.; Chevrette, M.; Tonin, P.N. Characterization of the 3p12.3-pcen region associated with tumor suppression in a novel ovarian cancer cell line model genetically modified by chromosome 3 fragment transfer. Mol. Carcinog 2009, 48, 1077–1092. [Google Scholar]
- Dong, Y.; Mao-Ying, Q.L.; Chen, J.W.; Yang, C.J.; Wang, Y.Q.; Tan, Z.M. Involvement of EphB1 receptor/ephrinB1 ligand in bone cancer pain. Neurosci. Lett 2011, 496, 163–167. [Google Scholar]
- Liu, S.; Liu, W.T.; Liu, Y.P.; Dong, H.L.; Henkemeyer, M.; Xiong, L.Z.; Song, X.J. Blocking EphB1 receptor forward signaling in spinal cord relieves bone cancer pain and rescues analgesic effect of morphine treatment in rodents. Cancer Res 2011, 71, 4392–4402. [Google Scholar]
- Kuang, S.Q.; Bai, H.; Fang, Z.H.; Lopez, G.; Yang, H.; Tong, W.; Wang, Z.Z.; Garcia-Manero, G. Aberrant DNA methylation and epigenetic inactivation of Eph receptor tyrosine kinases and ephrin ligands in acute lymphoblastic leukemia. Blood 2010, 115, 2412–2419. [Google Scholar]
- Sheng, Z.; Wang, J.; Dong, Y.; Ma, H.; Zhou, H.; Sugimura, H.; Lu, G.; Zhou, X. EphB1 is underexpressed in poorly differentiated colorectal cancers. Pathobiology 2008, 75, 274–280. [Google Scholar]
- Wang, J.D.; Dong, Y.C.; Sheng, Z.; Ma, H.H.; Li, G.L.; Wang, X.L.; Lu, G.M.; Sugimura, H.; Jin, J.; Zhou, X.J. Loss of expression of EphB1 protein in gastric carcinoma associated with invasion and metastasis. Oncology 2007, 73, 238–245. [Google Scholar]
- Berardi, A.C.; Marsilio, S.; Rofani, C.; Salvucci, O.; Altavista, P.; Perla, F.M.; Diomedi-Camassei, F.; Uccini, S.; Kokai, G.; Landuzzi, L.; et al. Up-regulation of EphB and ephrin-B expression in rhabdomyosarcoma. Anticancer Res 2008, 28, 763–769. [Google Scholar]
- Huynh-Do, U.; Vindis, C.; Liu, H.; Cerretti, D.P.; McGrew, J.T.; Enriquez, M.; Chen, J.; Daniel, T.O. Ephrin-B1 transduces signals to activate integrin-mediated migration, attachment and angiogenesis. J. Cell Sci 2002, 115, 3073–3081. [Google Scholar]
- Pejovic, T.; Pande, N.T.; Mori, M.; Mhawech-Fauceglia, P.; Harrington, C.; Mongoue-Tchokote, S.; Dim, D.; Andrews, C.; Beck, A.; Tarumi, Y.; et al. Expression profiling of the ovarian surface kinome reveals candidate genes for early neoplastic changes. Translational Oncology 2009, 2, 341–349. [Google Scholar]
- Chen, Z.; Xiao, Y.; Zhang, J.; Li, J.; Liu, Y.; Zhao, Y.; Ma, C.; Luo, J.; Qiu, Y.; Huang, G.; Korteweg, C.; Gu, J. Transcription factors E2A, FOXO1 and FOXP1 regulate recombination activating gene expression in cancer cells. PLoS One 2011, 6, e20475. [Google Scholar]
- Goatly, A.; Bacon, C.M.; Nakamura, S.; Ye, H.; Kim, I.; Brown, P.J.; Ruskone-Fourmestraux, A.; Cervera, P.; Streubel, B.; Banham, A.H.; Du, M.Q. FOXP1 abnormalities in lymphoma: translocation breakpoint mapping reveals insights into deregulated transcriptional control. Mod. Pathol 2008, 21, 902–911. [Google Scholar]
- Streubel, B.; Vinatzer, U.; Lamprecht, A.; Raderer, M.; Chott, A. T(3;14)(p14.1;q32) involving IGH and FOXP1 is a novel recurrent chromosomal aberration in MALT lymphoma. Leukemia 2005, 19, 652–658. [Google Scholar]
- Fenton, J.A.; Schuuring, E.; Barrans, S.L.; Banham, A.H.; Rollinson, S.J.; Morgan, G.J.; Jack, A.S.; van Krieken, J.H.; Kluin, P.M. t(3;14)(p14;q32) results in aberrant expression of FOXP1 in a case of diffuse large B-cell lymphoma. Genes Chromosomes Cancer 2006, 45, 164–168. [Google Scholar]
- Banham, A.H.; Beasley, N.; Campo, E.; Fernandez, P.L.; Fidler, C.; Gatter, K.; Jones, M.; Mason, D.Y.; Prime, J.E.; Trougouboff, P.; et al. The FOXP1 winged helix transcription factor is a novel candidate tumor suppressor gene on chromosome 3p. Cancer Res 2001, 61, 8820–8829. [Google Scholar]
- Toma, M.I.; Grosser, M.; Herr, A.; Aust, D.E.; Meye, A.; Hoefling, C.; Fuessel, S.; Wuttig, D.; Wirth, M.P.; Baretton, G.B. Loss of heterozygosity and copy number abnormality in clear cell renal cell carcinoma discovered by high-density affymetrix 10K single nucleotide polymorphism mapping array. Neoplasia 2008, 10, 634–642. [Google Scholar]
- Wlodarska, I.; Veyt, E.; de Paepe, P.; Vandenberghe, P.; Nooijen, P.; Theate, I.; Michaux, L.; Sagaert, X.; Marynen, P.; Hagemeijer, A.; de Wolf-Peeters, C. FOXP1, a gene highly expressed in a subset of diffuse large B-cell lymphoma, is recurrently targeted by genomic aberrations. Leukemia 2005, 19, 1299–1305. [Google Scholar]
- Giatromanolaki, A.; Koukourakis, M.I.; Sivridis, E.; Gatter, K.C.; Harris, A.L.; Banham, A.H. Loss of expression and nuclear/cytoplasmic localization of the FOXP1 forkhead transcription factor are common events in early endometrial cancer: Relationship with estrogen receptors and HIF-1alpha expression. Mod. Pathol 2006, 19, 9–16. [Google Scholar]
- Brown, P.J.; Ashe, S.L.; Leich, E.; Burek, C.; Barrans, S.; Fenton, J.A.; Jack, A.S.; Pulford, K.; Rosenwald, A.; Banham, A.H. Potentially oncogenic B-cell activation-induced smaller isoforms of FOXP1 are highly expressed in the activated B cell-like subtype of DLBCL. Blood 2008, 111, 2816–2824. [Google Scholar]
- Green, M.R.; Gandhi, M.K.; Courtney, M.J.; Marlton, P.; Griffiths, L. Relative abundance of full-length and truncated FOXP1 isoforms is associated with differential NFκB activity in Follicular Lymphoma. Leuk. Res 2009, 33, 1699–1702. [Google Scholar]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative genomic profiling of human prostate cancer. Cancer Cell 2010, 18, 11–22. [Google Scholar]
- Kim, Y.S.; Hwan, J.D.; Bae, S.; Bae, D.H.; Shick, W.A. Identification of differentially expressed genes using an annealing control primer system in stage III serous ovarian carcinoma. BMC Cancer 2010, 10, 576. [Google Scholar]
- Winn, R.A.; van Scoyk, M.; Hammond, M.; Rodriguez, K.; Crossno, J.T., Jr; Heasley, L.E.; Nemenoff, R.A. Antitumorigenic effect of Wnt 7a and Fzd 9 in non-small cell lung cancer cells is mediated through ERK-5-dependent activation of peroxisome proliferator-activated receptor gamma. J. Biol. Chem 2006, 281, 26943–26950. [Google Scholar]
- Bren-Mattison, Y.; van Putten, V.; Chan, D.; Winn, R.; Geraci, M.W.; Nemenoff, R.A. Peroxisome proliferator-activated receptor-gamma (PPAR(gamma)) inhibits tumorigenesis by reversing the undifferentiated phenotype of metastatic non-small-cell lung cancer cells (NSCLC). Oncogene 2005, 24, 1412–1422. [Google Scholar]
- Leung, W.K.; Bai, A.H.; Chan, V.Y.; Yu, J.; Chan, M.W.; To, K.F.; Wu, J.R.; Chan, K.K.; Fu, Y.G.; Chan, F.K.; Sung, J.J. Effect of peroxisome proliferator activated receptor gamma ligands on growth and gene expression profiles of gastric cancer cells. Gut 2004, 53, 331–338. [Google Scholar]
- Tennis, M.A.; Vanscoyk, M.M.; Wilson, L.A.; Kelley, N.; Winn, R.A. Methylation of Wnt7a is modulated by DNMT1 and cigarette smoke condensate in non-small cell lung cancer. PLoS One 2012, 7, e32921. [Google Scholar]
- Yoshioka, S.; King, M.L.; Ran, S.; Okuda, H.; Maclean, J.A., 2nd; McAsey, M.E.; Sugino, N.; Brard, L.; Watabe, K.; Hayashi, K. WNT7A Regulates Tumor Growth and Progression in Ovarian Cancer through the WNT/beta-Catenin Pathway. Mol. Cancer Res 2012, 10, 496–482. [Google Scholar]
- Calvo, R.; West, J.; Franklin, W.; Erickson, P.; Bemis, L.; Li, E.; Helfrich, B.; Bunn, P.; Roche, J.; Brambilla, E.; et al. Altered HOX and WNT7A expression in human lung cancer. Proc. Natl. Acad. Sci. USA 2000, 97, 12776–12781. [Google Scholar]
- Winn, R.A.; Marek, L.; Han, S.Y.; Rodriguez, K.; Rodriguez, N.; Hammond, M.; van Scoyk, M.; Acosta, H.; Mirus, J.; Barry, N.; et al. Restoration of Wnt-7a expression reverses non-small cell lung cancer cellular transformation through frizzled-9-mediated growth inhibition and promotion of cell differentiation. J. Biol. Chem 2005, 280, 19625–19634. [Google Scholar]
- Li, S.; Chiang, T.C.; Davis, G.R.; Williams, R.M.; Wilson, V.P.; McLachlan, J.A. Decreased expression of Wnt7a mRNA is inversely associated with the expression of estrogen receptor-alpha in human uterine leiomyoma. J. Clin. Endocrinol. Metabol 2001, 86, 454–457. [Google Scholar]
- Fluge, O.; Bruland, O.; Akslen, L.A.; Lillehaug, J.R.; Varhaug, J.E. Gene expression in poorly differentiated papillary thyroid carcinomas. Thyroid 2006, 16, 161–175. [Google Scholar]
- Merritt, M.A.; Parsons, P.G.; Newton, T.R.; Martyn, A.C.; Webb, P.M.; Green, A.C.; Papadimos, D.J.; Boyle, G.M. Expression profiling identifies genes involved in neoplastic transformation of serous ovarian cancer. BMC Cancer 2009, 9, 378. [Google Scholar]
- Zhang, X.L.; Peng, C.J.; Peng, J.; Jiang, L.Y.; Ning, X.M.; Zheng, J.H. Prognostic role of Wnt7a expression in ovarian carcinoma patients. Neoplasma 2010, 57, 545–551. [Google Scholar]
- Prado, S.M.D.; Villaamil, V.M.; Gallego, G.A.; Calvo, M.B.; Cedrun, J.L.L.; Soliva, S.S.; Ayerbes, M.V.; Campelo, R.G.; Aparicio, L.M.A. Expression of Wnt gene family and frizzled receptors in head and neck squamous cell carcinomas. Virchows. Arch 2009, 455, 67–75. [Google Scholar]
- Hsu, Y.A.; Lin, H.J.; Sheu, J.J.; Shieh, F.K.; Chen, S.Y.; Lai, C.H.; Tsai, F.J.; Wan, L.; Chen, B.H. A novel interaction between interferon-inducible protein p56 and ribosomal protein L15 in gastric cancer cells. DNA Cell Biol 2011, 30, 671–679. [Google Scholar]
- Gerashchenko, G.V.; Bogatyrova, O.O.; Rudenko, E.E.; Kondratov, A.G.; Gordiyuk, V.V.; Zgonnyk, Y.M.; Vozianov, O.F.; Pavlova, T.V.; Zabarovsky, E.R.; Rynditch, A.V.; et al. Genetic and epigenetic changes of NKIRAS1 gene in human renal cell carcinomas. Exp. Oncol 2010, 32, 71–75. [Google Scholar]
- Lin, H.; Wang, Y.; Zhang, X.; Liu, B.; Zhang, W.; Cheng, J. Prognostic significance of kappaB-Ras1 expression in gliomas. Med. Oncol 2011, 29, 1272–1279. [Google Scholar]
- Wang, H.; Zhao, L.N.; Li, K.Z.; Ling, R.; Li, X.J.; Wang, L. Overexpression of ribosomal protein L15 is associated with cell proliferation in gastric cancer. BMC Cancer 2006, 6, 91. [Google Scholar]
- Dang, C.; Gottschling, M.; Manning, K.; O’Currain, E.; Schneider, S.; Sterry, W.; Stockfleth, E.; Nindl, I. Identification of dysregulated genes in cutaneous squamous cell carcinoma. Oncol. Rep 2006, 16, 513–519. [Google Scholar]
- Willett, R.T.; Greene, L.A. Gata2 is required for migration and differentiation of retinorecipient neurons in the superior colliculus. J. Neurosci 2011, 31, 4444–4455. [Google Scholar]
- Kashiwabara, Y.; Sasaki, S.; Matsushita, A.; Nagayama, K.; Ohba, K.; Iwaki, H.; Matsunaga, H.; Suzuki, S.; Misawa, H.; Ishizuka, K.; et al. Functions of PIT1 in GATA2-dependent transactivation of the thyrotropin beta promoter. J. Mol. Endocrinol 2009, 42, 225–237. [Google Scholar]
- Kandimalla, R.; van Tilborg, A.A.; Kompier, L.C.; Stumpel, D.J.; Stam, R.W.; Bangma, C.H.; Zwarthoff, E.C. Genome-wide Analysis of CpG Island Methylation in Bladder Cancer Identified TBX2, TBX3, GATA2, and ZIC4 as pTa-Specific Prognostic Markers. Eur. Urol 2012, 61, 1245–1256. [Google Scholar]
- Bodor, C.; Renneville, A.; Smith, M.; Charazac, A.; Iqbal, S.; Etancelin, P.; Cavenagh, J.; Barnett, M.J.; Kramarzova, K.; Krishnan, B.; et al. Germ-line GATA2 p.THR354MET mutation in familial myelodysplastic syndrome with acquired monosomy 7 and ASXL1 mutation demonstrating rapid onset and poor survival. Haematologica 2012, 97, 890–894. [Google Scholar]
- Hahn, C.N.; Chong, C.E.; Carmichael, C.L.; Wilkins, E.J.; Brautigan, P.J.; Li, X.C.; Babic, M.; Lin, M.; Carmagnac, A.; Lee, Y.K.; et al. Heritable GATA2 mutations associated with familial myelodysplastic syndrome and acute myeloid leukemia. Nat. Genet 2011, 43, 1012–1017. [Google Scholar]
- Acosta, D.; Suzuki, M.; Connolly, D.; Thompson, R.F.; Fazzari, M.J.; Greally, J.M.; Montagna, C. DNA methylation changes in murine breast adenocarcinomas allow the identification of candidate genes for human breast carcinogenesis. Mamm. Genome 2011, 22, 249–259. [Google Scholar]
- Wang, Y.; He, X.; Ngeow, J.; Eng, C. GATA2 negatively regulates PTEN by preventing nuclear translocation of androgen receptor and by androgen-independent suppression of PTEN transcription in breast cancer. Hum. Mol. Genet 2012, 21, 569–576. [Google Scholar]
- Jazdzewski, K.; Boguslawska, J.; Jendrzejewski, J.; Liyanarachchi, S.; Pachucki, J.; Wardyn, K.A.; Nauman, A.; de la Chapelle, A. Thyroid hormone receptor beta (THRB) is a major target gene for microRNAs deregulated in papillary thyroid carcinoma (PTC). J. Clin. Endocrinol. Metab 2011, 96, E546–E553. [Google Scholar]
- Master, A.; Wojcicka, A.; Piekielko-Witkowska, A.; Boguslawska, J.; Poplawski, P.; Tanski, Z.; Darras, V.M.; Williams, G.R.; Nauman, A. Untranslated regions of thyroid hormone receptor beta 1 mRNA are impaired in human clear cell renal cell carcinoma. Biochim. Biophys. Acta 2010, 1802, 995–1005. [Google Scholar]
- Ling, Y.; Xu, X.; Hao, J.; Ling, X.; Du, X.; Liu, X.; Zhao, X. Aberrant methylation of the THRB gene in tissue and plasma of breast cancer patients. Cancer Genet. Cytogenet 2010, 196, 140–145. [Google Scholar]
- Vasiljevic, N.; Wu, K.; Brentnall, A.R.; Kim, D.C.; Thorat, M.A.; Kudahetti, S.C.; Mao, X.; Xue, L.; Yu, Y.; Shaw, G.L.; et al. Absolute quantitation of DNA methylation of 28 candidate genes in prostate cancer using pyrosequencing. Dis. Markers 2011, 30, 151–161. [Google Scholar]
- Muller, I.; Urban, K.; Pantel, K.; Schwarzenbach, H. Comparison of genetic alterations detected in circulating microsatellite DNA in blood plasma samples of patients with prostate cancer and benign prostatic hyperplasia. Ann. N. Y. Acad. Sci 2006, 1075, 222–229. [Google Scholar]
- Schwarzenbach, H.; Chun, F.K.; Muller, I.; Seidel, C.; Urban, K.; Erbersdobler, A.; Huland, H.; Pantel, K.; Friedrich, M.G. Microsatellite analysis of allelic imbalance in tumour and blood from patients with prostate cancer. BJU Int 2008, 102, 253–258. [Google Scholar]
- Qiu, H.; Lotan, R.; Lippman, S.M.; Xu, X.C. Lack of correlation between expression of retinoic acid receptor-beta and loss of heterozygosity on chromosome band 3p24 in esophageal cancer. Genes Chromosomes Cancer 2000, 28, 196–202. [Google Scholar]
- Mannion, C.; Park, W.S.; Man, Y.G.; Zhuang, Z.; Albores-Saavedra, J.; Tavassoli, F.A. Endocrine tumors of the cervix: Morphologic assessment, expression of human papillomavirus, and evaluation for loss of heterozygosity on 1p,3p, 11q, and 17p. Cancer 1998, 83, 1391–1400. [Google Scholar]
- Rowley, H.; Jones, A.; Spandidos, D.; Field, J. Definition of a tumor suppressor gene locus on the short arm of chromosome 3 in squamous cell carcinoma of the head and neck by means of microsatellite markers. Arch. Otolaryngol. Head Neck Surg 1996, 122, 497–501. [Google Scholar]
- Buchhagen, D.L. Frequent involvement of chromosome 3p alterations in lung carcinogenesis: allelotypes of 215 established cell lines at six chromosome 3p loci. J. Cell. Biochem. Suppl 1996, 24, 198–209. [Google Scholar]
- Guigon, C.J.; Kim, D.W.; Willingham, M.C.; Cheng, S.Y. Mutation of thyroid hormone receptor-beta in mice predisposes to the development of mammary tumors. Oncogene 2011, 30, 3381–3390. [Google Scholar]
- Kashuba, V.I.; Li, J.; Wang, F.; Senchenko, V.N.; Protopopov, A.; Malyukova, A.; Kutsenko, A.S.; Kadyrova, E.; Zabarovska, V.I.; Muravenko, O.V.; et al. RBSP3 (HYA22) is a tumor suppressor gene implicated in major epithelial malignancies. Proc. Natl. Acad. Sci. USA 2004, 101, 4906–4911. [Google Scholar]
- Anedchenko, E.A.; Dmitriev, A.A.; Krasnov, G.S.; Kondrat’eva, T.T.; Kopantsev, E.P.; Vinogradova, T.V.; Zinov’eva, M.V.; Zborovskaia, I.B.; Polotskii, B.E.; Sakharova, O.V.; et al. Down-regulation of RBSP3/CTDSPL, NPRL2/G21, RASSF1A, ITGA9, HYAL1 and HYAL2 genes in non-small cell lung cancer. Mol. Biol (Mosk) 2008, 42, 965–976. [Google Scholar]
- Anedchenko, E.A.; Kiseleva, N.P.; Dmitriev, A.A.; Kiselev, F.L.; Zabarovskii, E.R.; Senchenko, V.N. Tumor suppressor gene RBSP3 in cervical carcinoma: Copy number and transcriptional level. Mol. Biol (Mosk) 2007, 41, 86–95. [Google Scholar]
- Pavlova, T.V.; Kashuba, V.I.; Muravenko, O.V.; Yenamandra, S.P.; Ivanova, T.A.; Zabarovskaia, V.I.; Rakhmanaliev, E.R.; Petrenko, L.A.; Pronina, I.V.; Loginov, V.I.; et al. Technology of analysis of epigenetic and structural changes of epithelial tumors genome with NotI-microarrays by the example of human chromosome. Mol. Biol (Mosk) 2009, 43, 339–347. [Google Scholar]
- Senchenko, V.N.; Anedchenko, E.A.; Kondratieva, T.T.; Krasnov, G.S.; Dmitriev, A.A.; Zabarovska, V.I.; Pavlova, T.V.; Kashuba, V.I.; Lerman, M.I.; Zabarovsky, E.R. Simultaneous down-regulation of tumor suppressor genes RBSP3/CTDSPL, NPRL2/G21 and RASSF1A in primary non-small cell lung cancer. BMC Cancer 2010, 10, 75. [Google Scholar]
- Sinha, S.; Singh, R.K.; Alam, N.; Roy, A.; Roychoudhury, S.; Panda, C.K. Frequent alterations of hMLH1 and RBSP3/HYA22 at chromosomal 3p22.3 region in early and late-onset breast carcinoma: Clinical and prognostic significance. Cancer Sci 2008, 99, 1984–1991. [Google Scholar]
- Ghosh, A.; Ghosh, S.; Maiti, G.P.; Sabbir, M.G.; Zabarovsky, E.R.; Roy, A.; Roychoudhury, S.; Panda, C.K. Frequent alterations of the candidate genes hMLH1, ITGA9 and RBSP3 in early dysplastic lesions of head and neck: Clinical and prognostic significance. Cancer Sci 2010, 101, 1511–1520. [Google Scholar]
- Mitra, S.; Mazumder Indra, D.; Bhattacharya, N.; Singh, R.K.; Basu, P.S.; Mondal, R.K.; Roy, A.; Zabarovsky, E.R.; Roychoudhury, S.; Panda, C.K. RBSP3 is frequently altered in premalignant cervical lesions: Clinical and prognostic significance. Genes Chromosomes Cancer 2010, 49, 155–170. [Google Scholar]
- Kashuba, V.I.; Pavlova, T.V.; Grigorieva, E.V.; Kutsenko, A.; Yenamandra, S.P.; Li, J.; Wang, F.; Protopopov, A.I.; Zabarovska, V.I.; Senchenko, V.; et al. High mutability of the tumor suppressor genes RASSF1 and RBSP3 (CTDSPL) in cancer. PLoS One 2009, 4, e5231. [Google Scholar]
- Zheng, Y.S.; Zhang, H.; Zhang, X.J.; Feng, D.D.; Luo, X.Q.; Zeng, C.W.; Lin, K.Y.; Zhou, H.; Qu, L.H.; Zhang, P.; et al. MiR-100 regulates cell differentiation and survival by targeting RBSP3, a phosphatase-like tumor suppressor in acute myeloid leukemia. Oncogene 2012, 31, 80–92. [Google Scholar]
- Shoubridge, C.; Walikonis, R.S.; Gecz, J.; Harvey, R.J. Subtle functional defects in the Arf-specific guanine nucleotide exchange factor IQSEC2 cause non-syndromic X-linked intellectual disability. Small GTPases 2010, 1, 98–103. [Google Scholar]
- Someya, A.; Moss, J.; Nagaoka, I. The guanine nucleotide exchange protein for ADP-ribosylation factor 6, ARF-GEP100/BRAG2, regulates phagocytosis of monocytic phagocytes in an ARF6-dependent process. J. Biol. Chem 2010, 285, 30698–30707. [Google Scholar]
- Morishige, M.; Hashimoto, S.; Ogawa, E.; Toda, Y.; Kotani, H.; Hirose, M.; Wei, S.; Hashimoto, A.; Yamada, A.; Yano, H.; et al. GEP100 links epidermal growth factor receptor signalling to Arf6 activation to induce breast cancer invasion. Nat. Cell. Biol 2008, 10, 85–92. [Google Scholar]
- Sabe, H.; Hashimoto, S.; Morishige, M.; Ogawa, E.; Hashimoto, A.; Nam, J.M.; Miura, K.; Yano, H.; Onodera, Y. The EGFR-GEP100-Arf6-AMAP1 signaling pathway specific to breast cancer invasion and metastasis. Traffic 2009, 10, 982–993. [Google Scholar]
- Gunay-Aygun, M.; Falik-Zaccai, T.C.; Vilboux, T.; Zivony-Elboum, Y.; Gumruk, F.; Cetin, M.; Khayat, M.; Boerkoel, C.F.; Kfir, N.; Huang, Y.; et al. NBEAL2 is mutated in gray platelet syndrome and is required for biogenesis of platelet alpha-granules. Nat. Genet 2011, 43, 732–734. [Google Scholar]
- SABiosciences Gene Network Central Home Page. Available online: http://www.sabiosciences.com/genenetwork/genenetworkcentral.php accessed on 20 June 2012.
- Tohyama, J.; Kato, M.; Kawasaki, S.; Harada, N.; Kawara, H.; Matsui, T.; Akasaka, N.; Ohashi, T.; Kobayashi, Y.; Matsumoto, N. Dandy-Walker malformation associated with heterozygous ZIC1 and ZIC4 deletion: Report of a new patient. Am. J. Med. Genet. A 2011, 155A, 130–133. [Google Scholar]
- Aruga, J.; Nozaki, Y.; Hatayama, M.; Odaka, Y.S.; Yokota, N. Expression of ZIC family genes in meningiomas and other brain tumors. BMC Cancer 2010, 10, 79. [Google Scholar]
- Pourebrahim, R.; Van Dam, K.; Bauters, M.; De Wever, I.; Sciot, R.; Cassiman, J.J.; Tejpar, S. ZIC1 gene expression is controlled by DNA and histone methylation in mesenchymal proliferations. FEBS Lett 2007, 581, 5122–5126. [Google Scholar]
- Bataller, L.; Wade, D.F.; Graus, F.; Stacey, H.D.; Rosenfeld, M.R.; Dalmau, J. Antibodies to Zic4 in paraneoplastic neurologic disorders and small-cell lung cancer. Neurology 2004, 62, 778–782. [Google Scholar]
- Sabater, L.; Bataller, L.; Suarez-Calvet, M.; Saiz, A.; Dalmau, J.; Graus, F. ZIC antibodies in paraneoplastic cerebellar degeneration and small cell lung cancer. J. Neuroimmunol 2008, 201–202, 163–165. [Google Scholar]
- GeneCards Home Page. Available online: http://genecards.org/ accessed on 20 June 2012.
- Dunwell, T.L.; Hesson, L.B.; Pavlova, T.; Zabarovska, V.; Kashuba, V.; Catchpoole, D.; Chiaramonte, R.; Brini, A.T.; Griffiths, M.; Maher, E.R.; et al. Epigenetic analysis of childhood acute lymphoblastic leukemia. Epigenetics 2009, 4, 185–193. [Google Scholar]
- Haraldson, K.; Kashuba, V.I.; Dmitriev, A.A.; Senchenko, V.N.; Kudryavtseva, A.V.; Pavlova, T.V.; Braga, E.A.; Pronina, I.V.; Kondratov, A.G.; Rynditch, A.V.; et al. LRRC3B gene is frequently epigenetically inactivated in several epithelial malignancies and inhibits cell growth and replication. Biochimie 2012, 94, 1151–1157. [Google Scholar]
- Law, E.W.; Cheung, A.K.; Kashuba, V.I.; Pavlova, T.V.; Zabarovsky, E.R.; Lung, H.L.; Cheng, Y.; Chua, D.; Lai-Wan Kwong, D.; Tsao, S.W.; et al. Anti-angiogenic and tumor-suppressive roles of candidate tumor-suppressor gene, Fibulin-2, in nasopharyngeal carcinoma. Oncogene 2012, 31, 728–738. [Google Scholar]
- Buys, S.S.; Partridge, E.; Greene, M.H.; Prorok, P.C.; Reding, D.; Riley, T.L.; Hartge, P.; Fagerstrom, R.M.; Ragard, L.R.; Chia, D.; et al. Ovarian cancer screening in the Prostate, Lung, Colorectal and Ovarian (PLCO) cancer screening trial: Findings from the initial screen of a randomized trial. Am. J. Obstet. Gynecol 2005, 193, 1630–1639. [Google Scholar]
- Balch, C.; Fang, F.; Matei, D.E.; Huang, T.H.; Nephew, K.P. Minireview: epigenetic changes in ovarian cancer. Endocrinology 2009, 150, 4003–4011. [Google Scholar]
- Baylin, S.B.; Jones, P.A. A decade of exploring the cancer epigenome—Biological and translational implications. Nat. Rev. Cancer 2011, 11, 726–734. [Google Scholar]
- Baylin, S.B.; Ohm, J.E. Epigenetic gene silencing in cancer—A mechanism for early oncogenic pathway addiction? Nat. Rev. Cancer 2006, 6, 107–116. [Google Scholar]
- Widschwendter, M.; Jiang, G.; Woods, C.; Muller, H.M.; Fiegl, H.; Goebel, G.; Marth, C.; Muller-Holzner, E.; Zeimet, A.G.; Laird, P.W.; et al. DNA hypomethylation and ovarian cancer biology. Cancer Res 2004, 64, 4472–4480. [Google Scholar]
- Collins, Y.; Dicioccio, R.; Keitz, B.; Lele, S.; Odunsi, K. Methylation of death-associated protein kinase in ovarian carcinomas. Int. J. Gynecol. Cancer 2006, 16 Suppl 1, 195–199. [Google Scholar]
- Gifford, G.; Paul, J.; Vasey, P.A.; Kaye, S.B.; Brown, R. The acquisition of hMLH1 methylation in plasma DNA after chemotherapy predicts poor survival for ovarian cancer patients. Clin. Cancer Res 2004, 10, 4420–4426. [Google Scholar]
- Liggett, T.E.; Melnikov, A.; Yi, Q.; Replogle, C.; Hu, W.; Rotmensch, J.; Kamat, A.; Sood, A.K.; Levenson, V. Distinctive DNA methylation patterns of cell-free plasma DNA in women with malignant ovarian tumors. Gynecol. Oncol 2011, 120, 113–120. [Google Scholar]
- Melnikov, A.; Scholtens, D.; Godwin, A.; Levenson, V. Differential methylation profile of ovarian cancer in tissues and plasma. J. Molecular Diagn 2009, 11, 60–65. [Google Scholar]
- Gloss, B.S.; Samimi, G. Epigenetic biomarkers in epithelial ovarian cancer. Cancer Lett 2012. [Google Scholar] [CrossRef]
- Kashuba, V.I.; Gizatullin, R.Z.; Protopopov, A.I.; Li, J.; Vorobieva, N.V.; Fedorova, L.; Zabarovska, V.I.; Muravenko, O.V.; Kost-Alimova, M.; Domninsky, D.A.; et al. Analysis of NotI linking clones isolated from human chromosome 3 specific libraries. Gene 1999, 239, 259–271. [Google Scholar]
- Dmitriev, A.A.; Kashuba, V.I.; Haraldson, K.; Senchenko, V.N.; Pavlova, T.V.; Kudryavtseva, A.V.; Anedchenko, E.A.; Krasnov, G.S.; Pronina, I.V.; Loginov, V.I.; et al. Genetic and epigenetic analysis of non-small cell lung cancer with NotI-microarrays. Epigenetics 2012, 7, 502–513. [Google Scholar]
- AnalystSoft BioStat home page. Available online: http://www.analystsoft.com/en/products/biostat/ accessed on 27 November 2009.
- Altman, D.G.; Bland, J.M. Diagnostic tests. 1: Sensitivity and specificity. BMJ 1994, 308, 1552. [Google Scholar]
- Dreijerink, K.; Braga, E.; Kuzmin, I.; Geil, L.; Duh, F.M.; Angeloni, D.; Zbar, B.; Lerman, M.I.; Stanbridge, E.J.; Minna, J.D.; et al. The candidate tumor suppressor gene, RASSF1A, from human chromosome 3p21.3 is involved in kidney tumorigenesis. Proc. Natl. Acad. Sci. USA 2001, 98, 7504–7509. [Google Scholar]
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).