A Rice Immunophilin Homolog, OsFKBP12, Is a Negative Regulator of Both Biotic and Abiotic Stress Responses



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
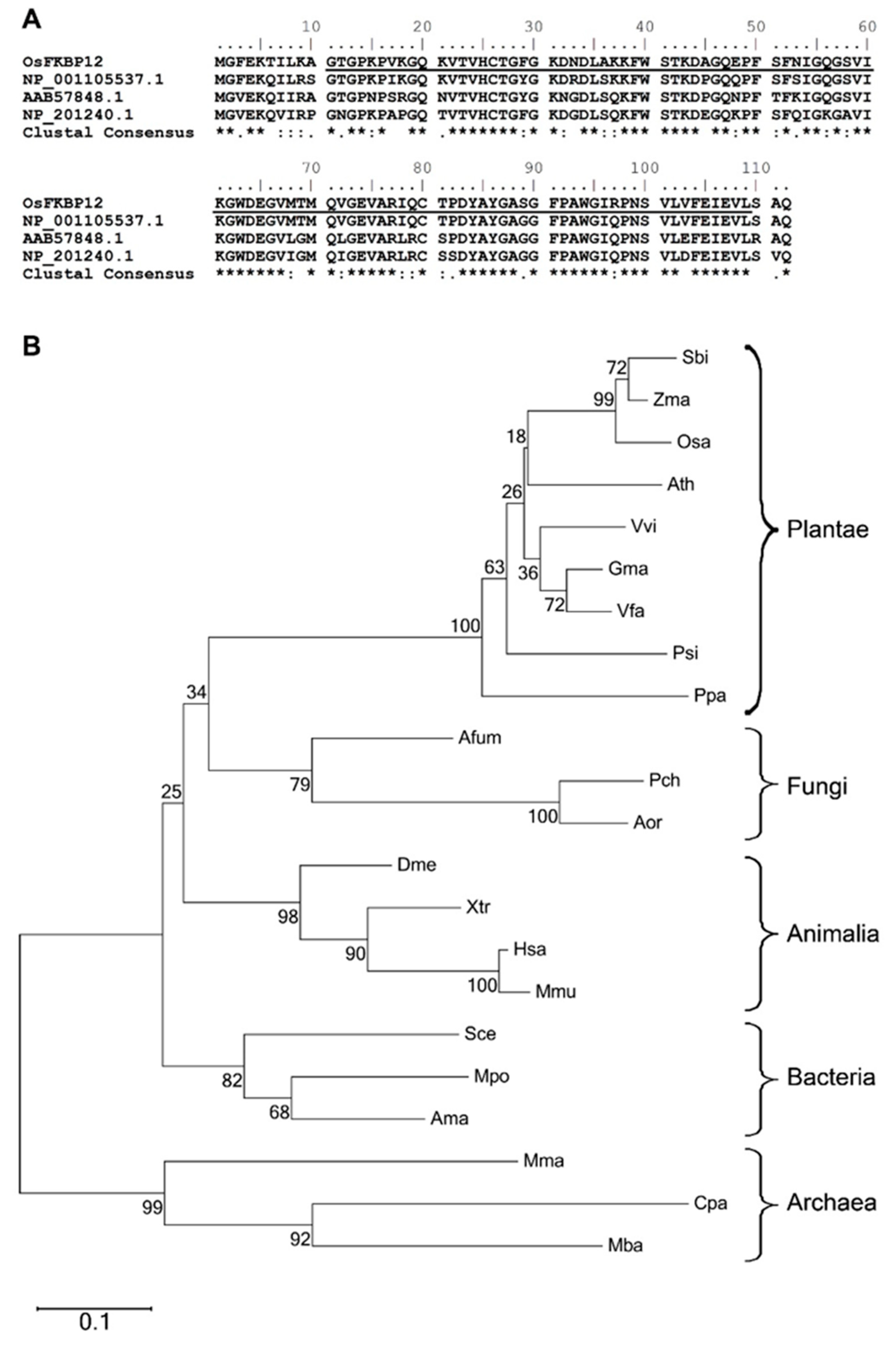
2.1. Identification of OsFKBP12, a Gene Encoding an FK506-Binding Protein (FKBP) in Oryza sativa
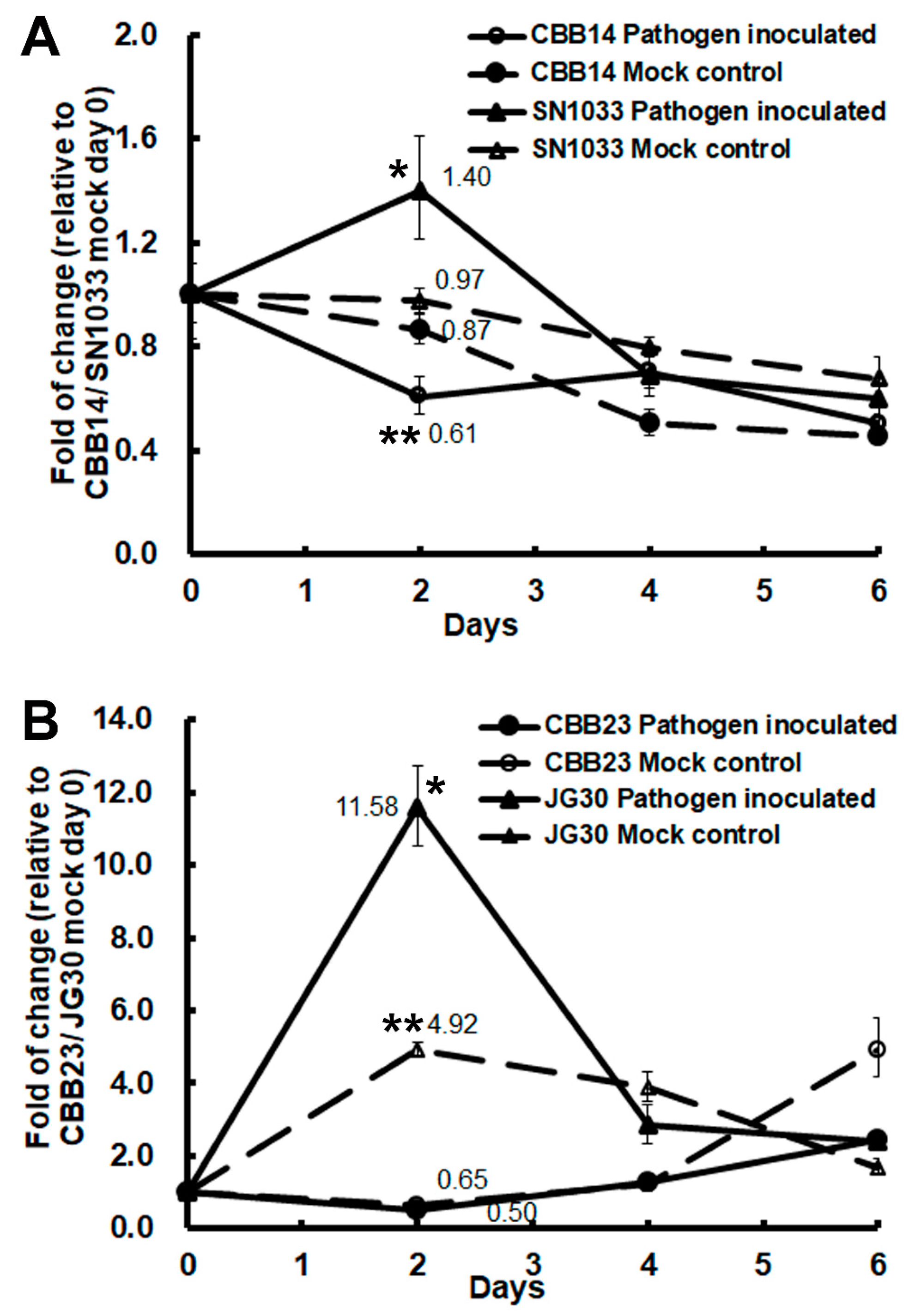
2.2. OsFKBP12 Expression was induced by the Pathogen Xanthomonas oryza pv. oryza (Xoo) in the Susceptible Recurrent Parents of Resistant NILs
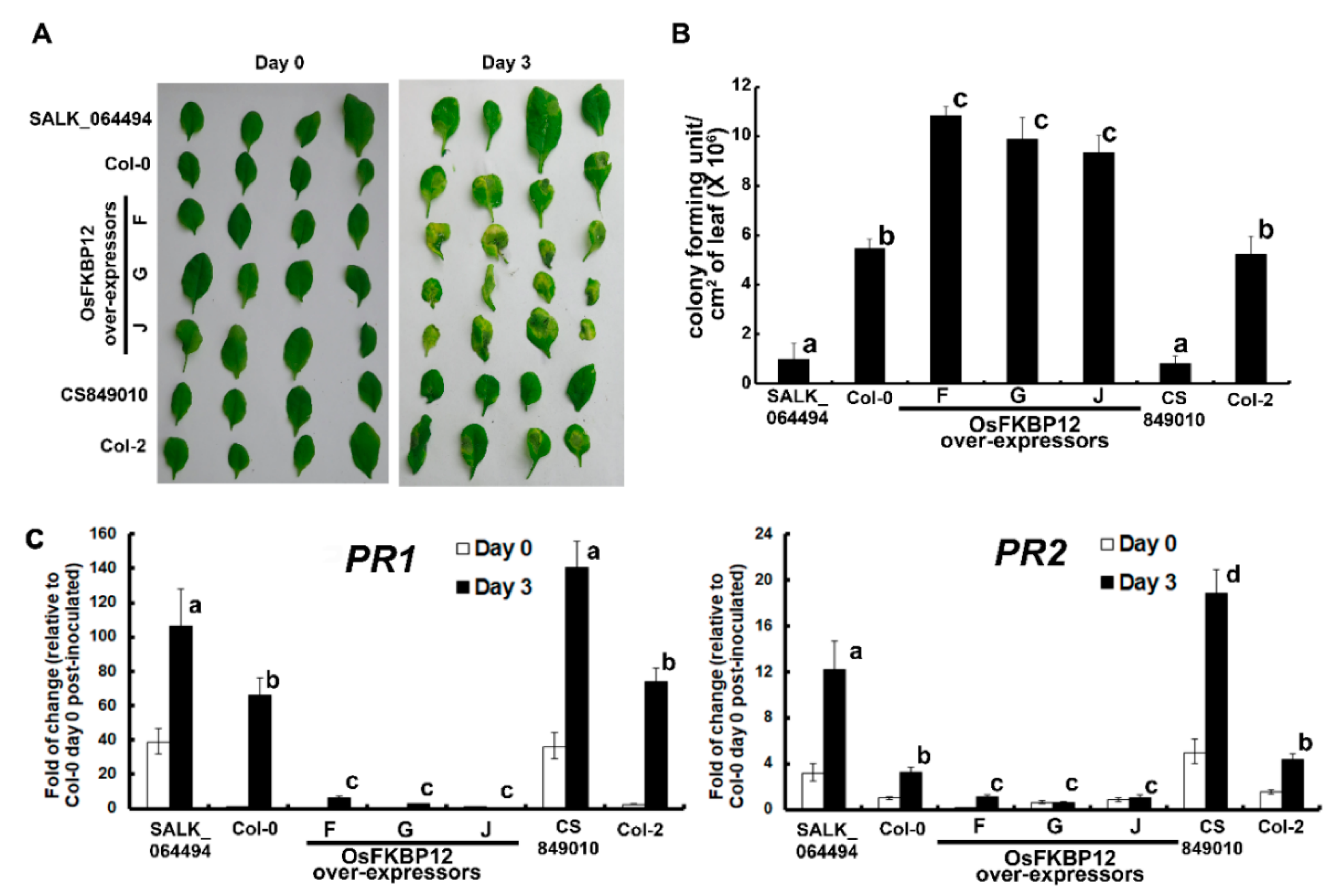
2.3. Ectopic Expression of OsFKBP12 Repressed Salicylic Acid (SA)-Related Defense Marker Genes and Enhanced the Susceptibility to Pseudomonas syringae pv. Tomato DC3000 (Pst DC3000) in Transgenic A. thaliana
2.4. Ectopic Expression of OsFKBP12 in Arabidopsis Increased Salt Sensitivity and Reduced Expressions of Genes Related to Salt Tolerance
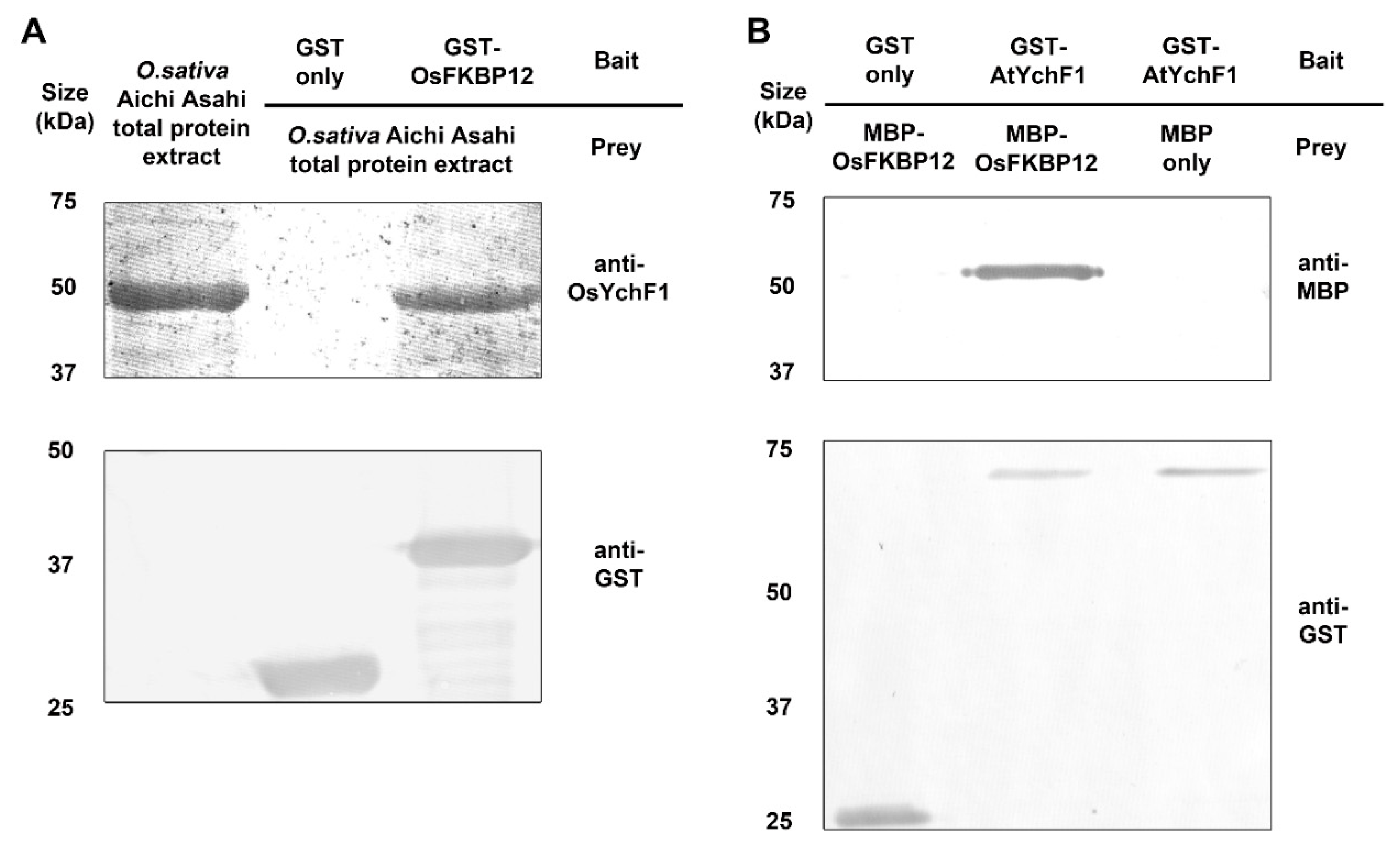
2.5. OsFKBP12 Protein Interacted with an Ancient Unconventional G-Protein, OsYchF1
3. Discussion
3.1. OsFKBP12 Encodes an FKBP (FK506-Binding Protein) with a Conserved FKBP-Specific Domain in Oryza sativa
3.2. OsFKBP12 Encodes a Negative Regulator of Defense Responses
3.3. OsFKBP12 Increased Salt Sensitivity in Transgenic Arabidopsis
3.4. OsFKBP12 Interacts with a Defense-Related GTPase, OsYchF1
4. Materials and Methods
4.1. Plant Material and Chemicals
4.2. Pathogen Inoculations of Rice with Xanthomonas oryza pv. oryza (Xoo) and Arabidopsis with Pseudomonas syringae pv. Tomato (Pst DC3000)
4.3. Salt Treatment of Arabidopsis
4.4. Cloning of OsFKBP12, and cDNA Library Construction Using Suppression Subtractive Hybridization Technique
4.5. Phylogenetic Analyses and Sequence Alignments
4.6. Yeast Two-Hybrid, Fusion Protein Constructions and In-Vitro Pulldown Assays
4.7. Bimolecular Fluorescence Complementation (BiFC) Experiment
4.8. Online Bioinformatics Tools
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galat, A.; Rivière, S. Peptidyl-Prolyl Cis/Trans Isomerases; Oxford University Press: Oxford, UK, 1998; ISBN 0198502885. [Google Scholar]
- Choi, J.; Chen, J.; Schreiber, S.L.; Clardy, J. Structure of the FKBP12-Rapamycin Complex Interacting with Binding Domain of Human FRAP. Science 1996, 273, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.G.; Gray, J.; Horton, P.; Luan, S. Plant immunophilins: Functional versatility beyond protein maturation. New Phytol. 2005, 166, 753–769. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; He, Z.; Luan, S. Introducing Immunophilins. From Organ Transplantation to Plant Biology. Plant Physiol. 2004, 134, 1241–1243. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gollan, P.J.; Bhave, M. Genome-wide analysis of genes encoding FK506-binding proteins in rice. Plant Mol. Biol. 2009, 72, 1–16. [Google Scholar] [CrossRef]
- He, Z.; Li, L.; Luan, S. Immunophilins and Parvulins. Superfamily of Peptidyl Prolyl Isomerases in Arabidopsis. Plant Physiol. 2004, 134, 1248–1267. [Google Scholar] [CrossRef]
- Aghdasi, B.; Ye, K.; Resnick, A.; Huang, A.; Ha, H.C.; Guo, X.; Dawson, T.M.; Dawson, V.L.; Snyder, S.H. FKBP12, the 12-kDa FK506-binding protein, is a physiologic regulator of the cell cycle. Proc. Natl. Acad. Sci. USA 2001, 98, 2425–2430. [Google Scholar] [CrossRef]
- Wang, W.-W.; Ma, Q.; Xiang, Y.; Zhu, S.-W.; Cheng, B.-J. Genome-wide analysis of immunophilin FKBP genes and expression patterns in Zea mays. Genet. Mol. Res. 2012, 11, 1690–1700. [Google Scholar] [CrossRef]
- Xu, Q.; Liang, S.; Kudla, J.; Luan, S. Molecular characterization of a plant FKBP12 that does not mediate action of FK506 and rapamycin. Plant J. 1998, 15, 511–519. [Google Scholar] [CrossRef]
- Sormani, R.; Yao, L.; Menand, B.; Ennar, N.; Lecampion, C.; Meyer, C.; Robaglia, C. Saccharomyces cerevisiae FKBP12 binds Arabidopsis thaliana TOR and its expression in plants leads to rapamycin susceptibility. BMC Plant Biol. 2007, 7, 26. [Google Scholar] [CrossRef]
- Faure, J.-D.; Gingerich, D.; Howell, S.H. An Arabidopsis immunophilin, AtFKBP12, binds to AtFIP37 (FKBP interacting protein) in an interaction that is disrupted by FK506. Plant J. 1998, 15, 783–789. [Google Scholar] [CrossRef]
- Gupta, R.; Mould, R.M.; He, Z.; Luan, S. Nonlinear partial differential equations and applications: A chloroplast FKBP interacts with and affects the accumulation of Rieske subunit of cytochrome bf complex. Proc. Natl. Acad. Sci. USA 2002, 99, 15806–15811. [Google Scholar] [CrossRef] [PubMed]
- Kamphausen, T.; Fanghänel, J.; Neumann, D.; Schulz, B.; Rahfeld, J.-U. Characterization of Arabidopsis thaliana AtFKBP42 that is membrane-bound and interacts with Hsp90. Plant J. 2002, 32, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Vespa, L.; Vachon, G.; Berger, F.; Perazza, D.; Faure, J.-D.; Herzog, M. The Immunophilin-Interacting Protein AtFIP37 from Arabidopsis Is Essential for Plant Development and Is Involved in Trichome Endoreduplication. Plant Physiol. 2004, 134, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.; Lima, S.; Wong, J.H.; Phillips, R.S.; Buchanan, B.B.; Luan, S. A redox-active FKBP-type immunophilin functions in accumulation of the photosystem II supercomplex in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2006, 103, 12631–12636. [Google Scholar] [CrossRef]
- Aviezer-Hagai, K.; Skovorodnikova, J.; Galigniana, M.; Farchi-Pisanty, O.; Maayan, E.; Bocovza, S.; Efrat, Y.; Von Koskull-Döring, P.; Ohad, N.; Breiman, A. Arabidopsis immunophilins ROF1 (AtFKBP62) and ROF2 (AtFKBP65) exhibit tissue specificity, are heat-stress induced, and bind HSP90. Plant Mol. Biol. 2007, 63, 237–255. [Google Scholar] [CrossRef]
- Gupta, D.; Tuteja, N. Chaperones and foldases in endoplasmic reticulum stress signaling in plants. Plant Signal. Behav. 2011, 6, 232–236. [Google Scholar] [CrossRef]
- Kromina, K.A.; Ignatov, A.N.; Abdeeva, I.A. Role of peptidyl-prolyl-cis/trans-isomerases in pathologic processes. Biochem. Suppl. Ser. A Membr. Cell Biol. 2008, 2, 195–202. [Google Scholar] [CrossRef]
- Coaker, G.; Falick, A.; Staskawicz, B. Activation of a Phytopathogenic Bacterial Effector Protein by a Eukaryotic Cyclophilin. Science 2005, 308, 548–550. [Google Scholar] [CrossRef]
- Coaker, G.; Zhu, G.; Ding, Z.; Van Doren, S.R.; Staskawicz, B. Eukaryotic cyclophilin as a molecular switch for effector activation. Mol. Microbiol. 2006, 61, 1485–1496. [Google Scholar] [CrossRef]
- Abdeeva, I.; Pogorelko, G.V.; Bruskin, S.A.; Piruzian, E.S.; Abdeeva, I.A. The role of peptidyl-prolyl cis/trans isomerase genes of Arabidopsis thaliana in plant defense during the course of Xanthomonas campestris infection. Russ. J. Genet. 2014, 50, 140–148. [Google Scholar] [CrossRef]
- Alavilli, H.; Lee, H.; Park, M.; Yun, D.-J.; Lee, B.-H. Enhanced multiple stress tolerance in Arabidopsis by overexpression of the polar moss peptidyl prolyl isomerase FKBP12 gene. Plant Cell Rep. 2017, 37, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.-Y.; Zeng, N.-Y.; Tong, S.-W.; Li, W.-Y.F.; Xue, Y.; Zhao, K.-J.; Wang, C.; Zhang, Q.; Fu, Y.; Sun, Z.; et al. Constitutive expression of a rice GTPase-activating protein induces defense responses. New Phytol. 2008, 179, 530–545. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wasaki, J.; Yonetani, R.; Shinano, T.; Kai, M.; Osaki, M. Expression of the OsPI1 gene, cloned from rice roots using cDNA microarray, rapidly responds to phosphorus status. New Phytol. 2003, 158, 239–248. [Google Scholar] [CrossRef]
- Katagiri, F.; Thilmony, R.; He, S.Y. The Arabidopsis Thaliana-Pseudomonas Syringae Interaction. Arab. Book 2002, 1, e0039. [Google Scholar] [CrossRef]
- Czechowski, T.; Bari, R.P.; Stitt, M.; Scheible, W.-R.; Udvardi, M.K. Real-time RT-PCR profiling of over 1400Arabidopsistranscription factors: Unprecedented sensitivity reveals novel root- and shoot-specific genes. Plant J. 2004, 38, 366–379. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Cheung, M.-Y.; Zeng, N.-Y.; Tong, S.-W.; Li, F.W.-Y.; Zhao, K.-J.; Zhang, Q.; Sun, S.S.-M.; Lam, H.-M. Expression of a RING-HC protein from rice improves resistance to Pseudomonas syringae pv. tomato DC3000 in transgenic Arabidopsis thaliana. J. Exp. Bot. 2007, 58, 4147–4159. [Google Scholar] [CrossRef]
- Zhou, L.; Cheung, M.; Zhang, Q.; Zhang, S.-H.; Sun, S.S.; Lam, H.-M.; Lei, C.-L. A novel simple extracellular leucine-rich repeat (eLRR) domain protein from rice (OsLRR1) enters the endosomal pathway and interacts with the hypersensitive-induced reaction protein 1 (OsHIR1). Plant Cell Environ. 2009, 32, 1804–1820. [Google Scholar] [CrossRef]
- Cheung, M.-Y.; Xue, Y.; Zhou, L.; Li, M.-W.; Sun, S.S.-M.; Lam, H.-M. An Ancient P-Loop GTPase in Rice Is Regulated by a Higher Plant-specific Regulatory Protein. J. Biol. Chem. 2010, 285, 37359–37369. [Google Scholar] [CrossRef] [PubMed]
- Geisler, M.; Girin, M.; Brandt, S.; Vincenzetti, V.; Plaza, S.; Paris, N.; Kobae, Y.; Maeshima, M.; Billion, K.; Kolukisaoglu, Ü.H.; et al. ArabidopsisImmunophilin-like TWD1 Functionally Interacts with Vacuolar ABC Transporters. Mol. Biol. Cell 2004, 15, 3393–3405. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nat. Cell Biol. 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Eggermont, K.; Penninckx, I.A.M.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Genes controlling expression of defense responses in Arabidopsis—2001 status. Curr. Opin. Plant Biol. 2001, 4, 301–308. [Google Scholar] [CrossRef]
- Spoel, S.H.; Koornneef, A.; Claessens, S.M.C.; Korzelius, J.P.; Van Pelt, J.A.; Mueller, M.J.; Buchala, A.J.; Métraux, J.-P.; Brown, R.; Kazan, K.; et al. NPR1 Modulates Cross-Talk between Salicylate- and Jasmonate-Dependent Defense Pathways through a Novel Function in the Cytosol. Plant Cell 2003, 15, 760–770. [Google Scholar] [CrossRef]
- Devoto, A.; Turner, J.G. Jasmonate-regulated Arabidopsis stress signalling network. Physiol. Plant. 2005, 123, 161–172. [Google Scholar] [CrossRef]
- Van Loon, L.; Van Strien, E. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Uknes, S.; Mauch-Mani, B.; Moyer, M.; Potter, S.; Williams, S.; Dincher, S.; Chandler, D.; Slusarenko, A.; Ward, E.; Ryals, J. Acquired resistance in Arabidopsis. Plant Cell Online 1992, 4, 645–656. [Google Scholar]
- Greenberg, J.T.; Ausubel, F.M. Arabidopsis mutants compromised for the control of cellular damage during pathogenesis and aging. Plant J. 1993, 4, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Crooks, C.; Creissen, G.; Hill, L.; Fairhurst, S.; Doerner, P.; Lamb, C. Pseudomonas sax Genes Overcome Aliphatic Isothiocyanate-Mediated Non-Host Resistance in Arabidopsis. Science 2011, 331, 1185–1188. [Google Scholar] [CrossRef] [PubMed]
- Bogdanović, M.; Milica, B.; Živković, S.; Sabovljevic, A.; Grubišić, D.; Sabovljević, M. Comparative study on the effects of NaCl on selected moss and fern representatives. Aust. J. Bot. 2011, 59, 734–740. [Google Scholar] [CrossRef]
- Alavilli, H.; Lee, H.; Park, M.; Lee, B.-H. Antarctic Moss Multiprotein Bridging Factor 1c Overexpression in Arabidopsis Resulted in Enhanced Tolerance to Salt Stress. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef]
- Amir, A.I.; Van Rosmalen, M.; Mayer, G.; Lebendiker, M.; Danieli, T.; Friedler, A. Highly homologous proteins exert opposite biological activities by using different interaction interfaces. Sci. Rep. 2015, 5, 11629. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the International Conference on Intelligent Systems for Molecular Biology, San Diego, CA, USA, 14–17 August 1994; Volume 2, pp. 28–36. [Google Scholar]
- Cunningham, J.J.; Estrella, V.; Lloyd, M.; Gillies, R.; Frieden, B.R.; Gatenby, R. Intracellular Electric Field and pH Optimize Protein Localization and Movement. PLoS ONE 2012, 7, e36894. [Google Scholar] [CrossRef]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Walker, J.M. (Ed.) The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–608. [Google Scholar] [CrossRef]
- Caldon, C.E.; Yoong, P.; March, P.E. Evolution of a molecular switch: Universal bacterial GTPases regulate ribosome function. Mol. Microbiol. 2001, 41, 289–297. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, A.N.; Yang, W.C.; Wang, C.L. Breeding of three near-isogenic japonica rice lines with different major genes for resistance to bacterial blight. Acta Agric. Sin. 1996, 22, 135–141. [Google Scholar]
- Horwitz, A.H.; Morandi, C.; Wilcox, G. Deoxyribonucleic acid sequence of araBAD promoter mutants of Escherichia coli. J. Bacteriol. 1980, 142, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Moran, R. Formulae for Determination of Chlorophyllous Pigments Extracted with N,N-Dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.-Y.; Li, M.-W.; Yung, Y.-L.; Wen, C.-Q.; Lam, H.-M. The unconventional P-loop NTPase OsYchF1 and its regulator OsGAP1 play opposite roles in salinity stress tolerance. Plant Cell Environ. 2013, 36, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Chou, H.-L.; Zhang, L.; Hwang, S.-K.; Starkenburg, S.R.; Doroshenk, K.A.; Kumamaru, T.; Okita, T.W. RNA-Binding Protein RBP-P Is Required for Glutelin and Prolamine mRNA Localization in Rice Endosperm Cells. Plant Cell 2018, 30, 2529–2552. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheung, M.-Y.; Auyeung, W.-K.; Li, K.-P.; Lam, H.-M. A Rice Immunophilin Homolog, OsFKBP12, Is a Negative Regulator of Both Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2020, 21, 8791. https://doi.org/10.3390/ijms21228791
Cheung M-Y, Auyeung W-K, Li K-P, Lam H-M. A Rice Immunophilin Homolog, OsFKBP12, Is a Negative Regulator of Both Biotic and Abiotic Stress Responses. International Journal of Molecular Sciences. 2020; 21(22):8791. https://doi.org/10.3390/ijms21228791
Chicago/Turabian StyleCheung, Ming-Yan, Wan-Kin Auyeung, Kwan-Pok Li, and Hon-Ming Lam. 2020. "A Rice Immunophilin Homolog, OsFKBP12, Is a Negative Regulator of Both Biotic and Abiotic Stress Responses" International Journal of Molecular Sciences 21, no. 22: 8791. https://doi.org/10.3390/ijms21228791
APA StyleCheung, M.-Y., Auyeung, W.-K., Li, K.-P., & Lam, H.-M. (2020). A Rice Immunophilin Homolog, OsFKBP12, Is a Negative Regulator of Both Biotic and Abiotic Stress Responses. International Journal of Molecular Sciences, 21(22), 8791. https://doi.org/10.3390/ijms21228791