
Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
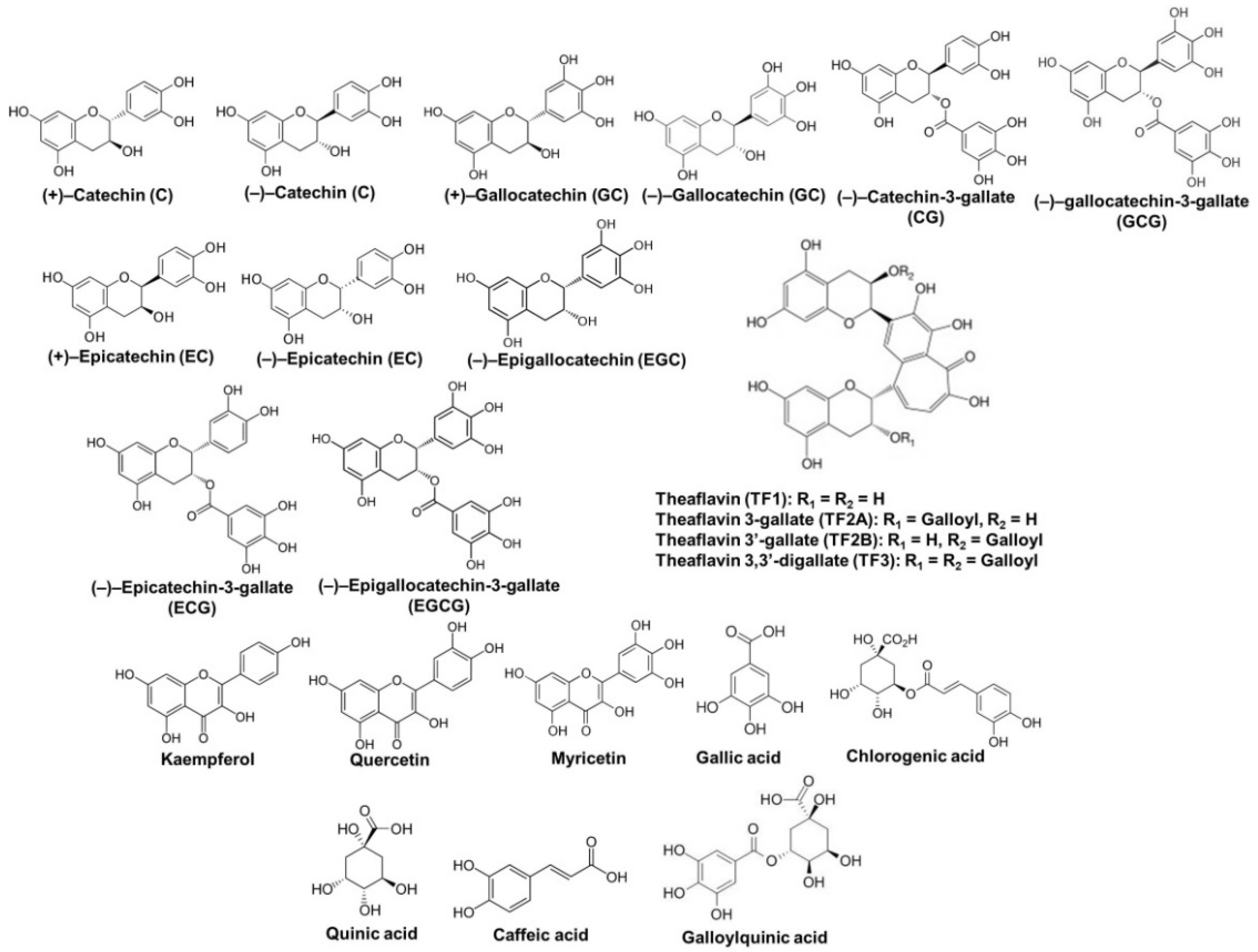
2. Chemical Properties of Tea Polyphenols
3. Antioxidant Activity of Tea Polyphenols
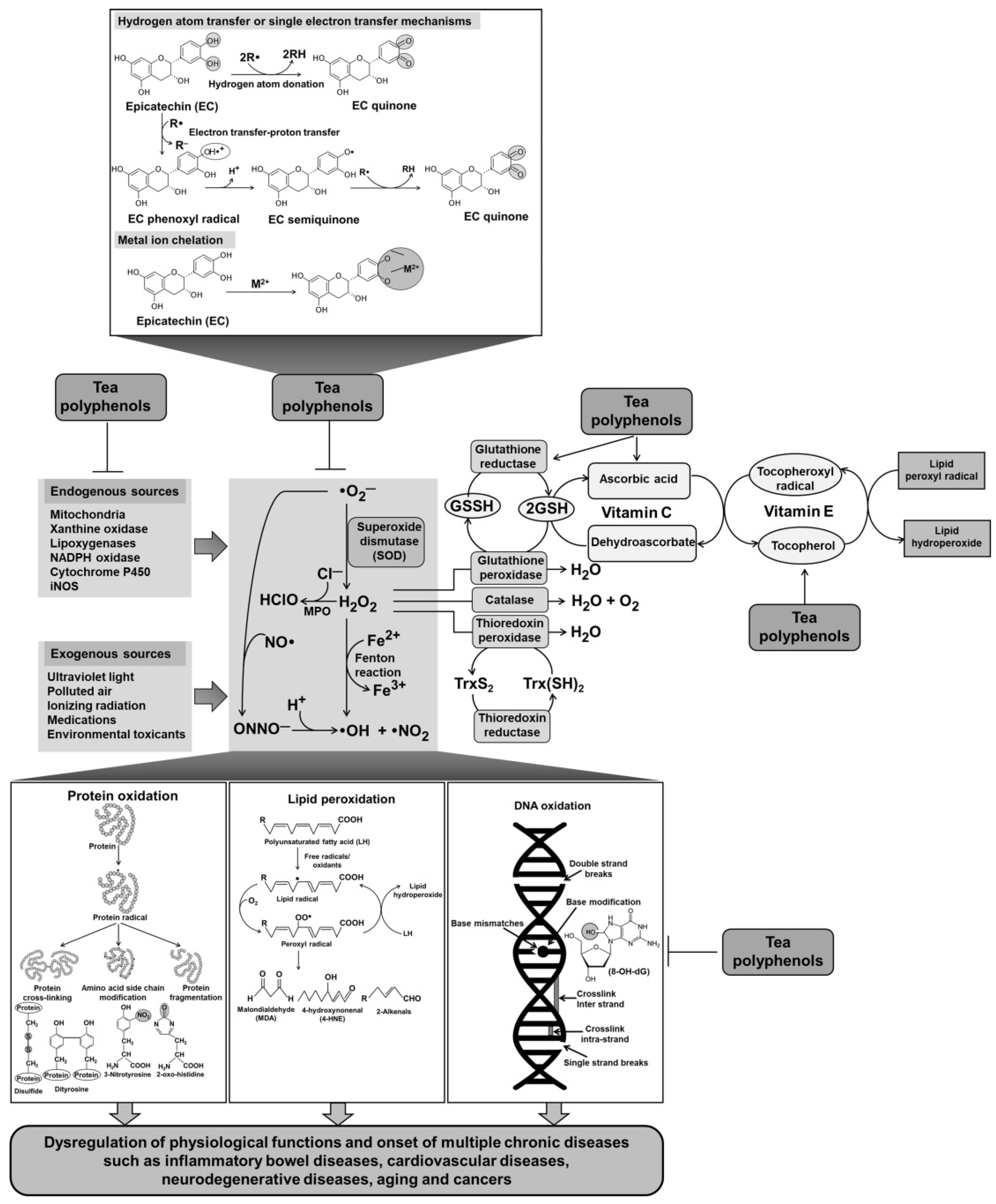
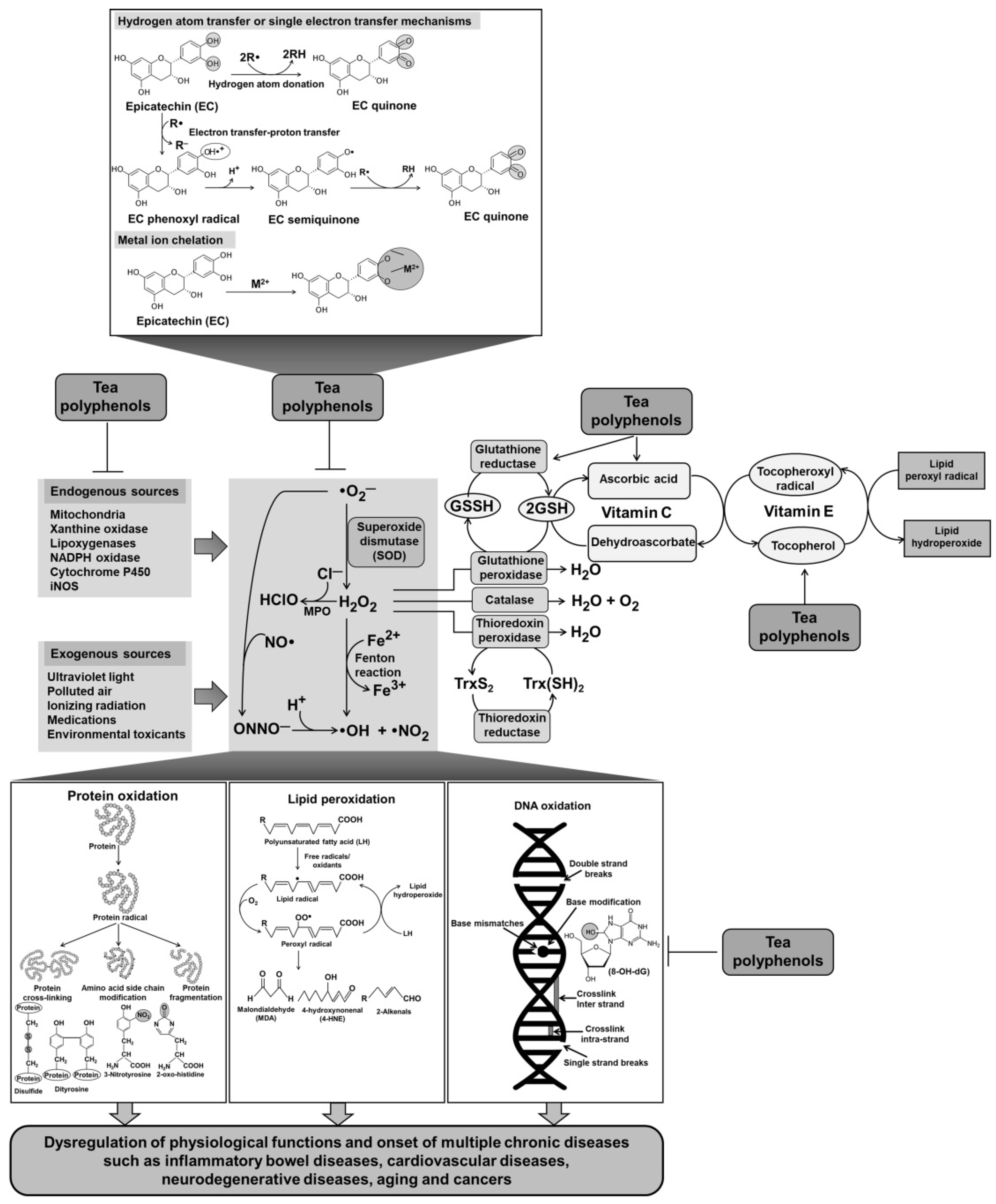
3.1. Direct Antioxidant Activity of Tea Polyphenols
3.1.1. Radical and Oxidant Scavenging Activities of Tea Polyphenols
3.1.2. Inhibition of Lipid Peroxidation by Tea Polyphenols
3.1.3. Inhibition of DNA Oxidation by Tea Polyphenols
3.1.4. Inhibition of Protein Oxidation by Tea Polyphenols
3.1.5. Metal Chelation by Tea Polyphenols
3.2. Indirect Antioxidant Activity of Tea Polyphenols
3.2.1. Inhibition of “Pro-Oxidant” Enzymes by Tea Polyphenols
3.2.2. Induction of Intracellular Antioxidant Defense System by Tea Polyphenols
3.2.3. Antioxidant Synergisms of Tea Polyphenols and Vitamins
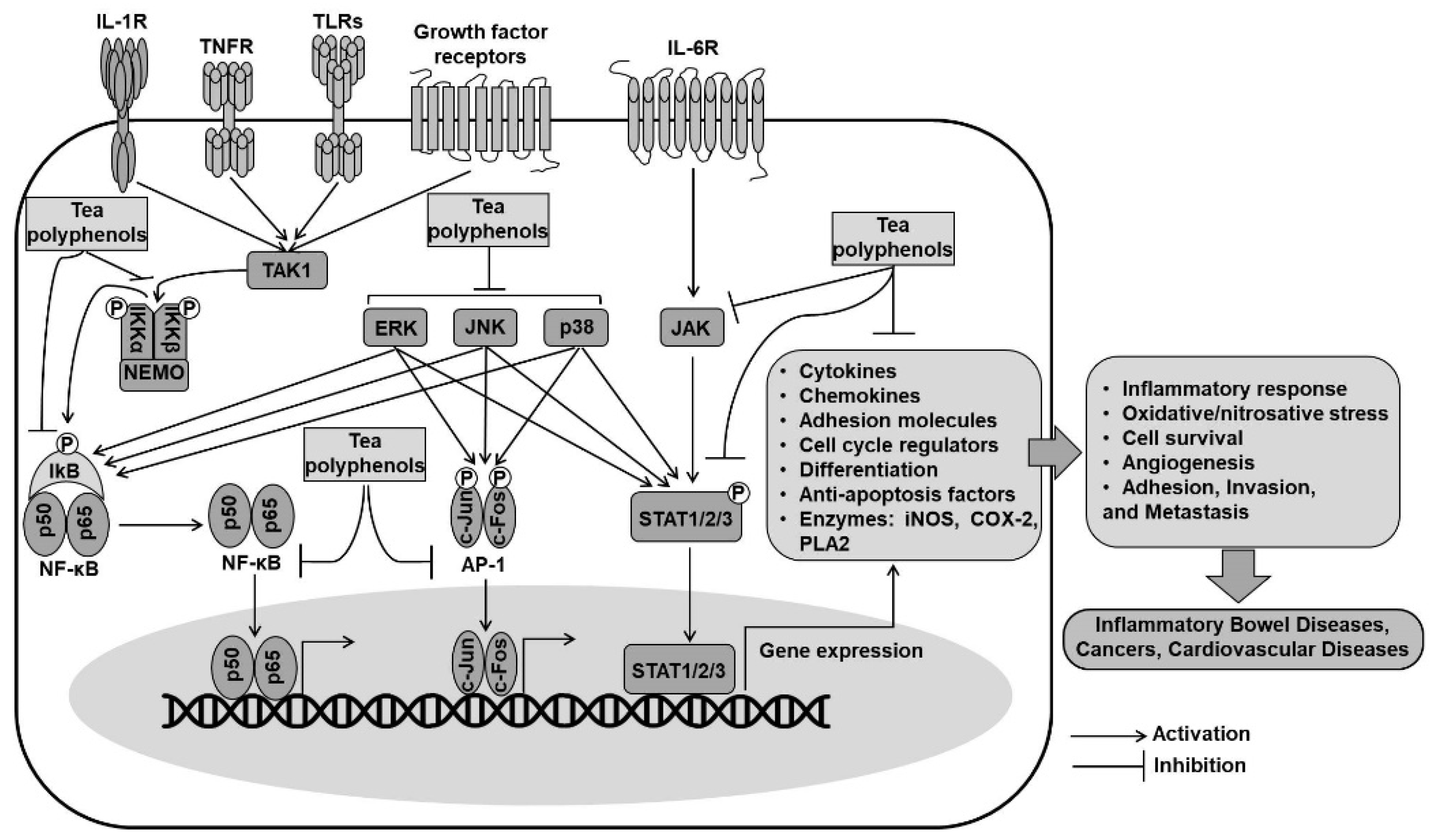
4. Cellular Defensive Mechanisms of Tea Polyphenols through the Regulation of Signaling Pathways
4.1. Regulation of Phase I and Phase II Enzymes by Tea Polyphenols
4.2. Inhibition of NF-κB and AP1 Pathways by Tea Polyphenols
4.3. Inhibition of the Signal Transducer and Activator of Transcription (STAT) Pathway by Tea Polyphenols
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Narotzki, B.; Reznick, A.Z.; Aizenbud, D.; Levy, Y. Green tea: A promising natural product in oral health. Arch. Oral Biol. 2012, 57, 429–435. [Google Scholar] [CrossRef]
- Xing, L.; Zhang, H.; Qi, R.; Tsao, R.; Mine, Y. Recent Advances in the Understanding of the Health Benefits and Molecular Mechanisms Associated with Green Tea Polyphenols. J. Agric. Food Chem. 2019, 67, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Sajilata, M.G.; Bajaj, P.R.; Singhal, R.S. Tea Polyphenols as Nutraceuticals. Compr. Rev. Food Sci. Food Saf. 2008, 7, 229–254. [Google Scholar] [CrossRef]
- Zhang, L.; Ho, C.-T.; Zhou, J.; Santos, J.S.; Armstrong, L.; Granato, D. Chemistry and Biological Activities of Processed Camellia sinensis Teas: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1474–1495. [Google Scholar] [CrossRef] [Green Version]
- Namal Senanayake, S.P.J. Green tea extract: Chemistry, antioxidant properties and food applications—A review. J. Funct. Foods 2013, 5, 1529–1541. [Google Scholar] [CrossRef]
- Yilmaz, Y. Novel uses of catechins in foods. Trends Food Sci. Technol. 2006, 17, 64–71. [Google Scholar] [CrossRef]
- Hashimoto, F.; Nonaka, G.-I.; Nishioka, I. Tannins and Related Compounds. XC.: 8-C-Ascorbyl (−)-Epigallocatechin 3-O-Gallate and Novel Dimeric Flavan-3-ols, Oolonghomobisflavans A and B, from Oolong Tea. (3). Chem. Pharm. Bull. 1989, 37, 3255–3263. [Google Scholar] [CrossRef]
- Corey, M.E.; Kerr, W.L.; Mulligan, J.H.; Lavelli, V. Phytochemical stability in dried apple and green tea functional products as related to moisture properties. LWT 2011, 44, 67–74. [Google Scholar] [CrossRef]
- Wang, H.; Helliwell, K. Determination of flavonols in green and black tea leaves and green tea infusions by high-performance liquid chromatography. Food Res. Int. 2001, 34, 223–227. [Google Scholar] [CrossRef]
- Monobe, M.; Nomura, S.; Ema, K.; Matsunaga, A.; Nesumi, A.; Yoshida, K.; Maeda-Yamamoto, M.; Horie, H. Quercetin Glycosides-rich Tea Cultivars (Camellia sinensis L.) in Japan. Food Sci. Technol. Res. 2015, 21, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Rusak, G.; Komes, D.; Likić, S.; Horžić, D.; Kovač, M. Phenolic content and antioxidative capacity of green and white tea extracts depending on extraction conditions and the solvent used. Food Chem. 2008, 110, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.S.; Deolindo, C.T.P.; Hoffmann, J.F.; Chaves, F.C.; do Prado-Silva, L.; Sant’Ana, A.S.; Azevedo, L.; do Carmo, M.A.V.; Granato, D. Optimized Camellia sinensis var. sinensis, Ilex paraguariensis, and Aspalathus linearis blend presents high antioxidant and antiproliferative activities in a beverage model. Food Chem. 2018, 254, 348–358. [Google Scholar] [CrossRef]
- Limón-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2009, 674, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Piechota-Polanczyk, A.; Fichna, J. Review article: The role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 605–620. [Google Scholar] [CrossRef] [Green Version]
- Forester, S.C.; Lambert, J.D. The role of antioxidant versus pro-oxidant effects of green tea polyphenols in cancer prevention. Mol. Nutr. Food Res. 2011, 55, 844–854. [Google Scholar] [CrossRef] [Green Version]
- Ricci, A.; Parpinello, G.P.; Versari, A. The Nutraceutical Impact of Polyphenolic Composition in Commonly Consumed Green Tea, Green Coffee and Red Wine Beverages: A Review. Recent. Adv. Food Sci. Nutr. Res. 2018, 2018, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-W.; Zhu, Z.-Q.; Hu, T.-X.; Zhu, D.-Y. Structure-activity relationship of natural flavonoids in hydroxyl radical-scavenging effects. Acta. Pharmacol. Sin. 2002, 23, 667–672. [Google Scholar]
- Frei, B.; Higdon, J.V. Antioxidant Activity of Tea Polyphenols In Vivo: Evidence from Animal Studies. J. Nutr. 2003, 133, 3275–3284. [Google Scholar] [CrossRef] [Green Version]
- Janeiro, P.; Oliveira Brett, A.M. Catechin electrochemical oxidation mechanisms. Anal. Chim. Acta 2004, 518, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant Polyphenols as Antioxidant and Antibacterial Agents for Shelf-Life Extension of Meat and Meat Products: Classification, Structures, Sources, and Action Mechanisms. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Zhao, B.; Shen, S.; Hou, J.; Hu, J.; Xin, W. ESR study on the structure–antioxidant activity relationship of tea catechins and their epimers. Biochim. Biophys. Acta 1999, 1427, 13–23. [Google Scholar] [CrossRef]
- Nakagawa, T.; Yokozawa, T. Direct scavenging of nitric oxide and superoxide by green tea. Food Chem. Toxicol. 2002, 40, 1745–1750. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Szeto, Y.T. Total Antioxidant Capacity of Teas by the Ferric Reducing/Antioxidant Power Assay. J. Agric. Food Chem. 1999, 47, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Fajardo-Lira, C.; Lee, H.W.; Youssefian, A.A.; Go, V.L.W.; Heber, D. Catechin Content of 18 Teas and a Green Tea Extract Supplement Correlates With the Antioxidant Capacity. Nutr. Cancer 2003, 45, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, Y.; Ohie, T.; Yonekawa, Y.; Yonemoto, K.; Aizawa, H.; Mori, Y.; Watanabe, M.; Takeuchi, M.; Hasegawa, M.; Taguchi, C.; et al. Coffee and Green Tea As a Large Source of Antioxidant Polyphenols in the Japanese Population. J. Agric. Food Chem. 2009, 57, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-N.; Tang, G.-Y.; Cao, S.-Y.; Xu, X.-Y.; Gan, R.-Y.; Liu, Q.; Mao, Q.-Q.; Shang, A.; Li, H.-B. Phenolic Profiles and Antioxidant Activities of 30 Tea Infusions from Green, Black, Oolong, White, Yellow and Dark Teas. Antioxidants 2019, 8, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nibir, Y.M.; Sumit, A.F.; Akhand, A.A.; Ahsan, N.; Hossain, M.S. Comparative assessment of total polyphenols, antioxidant and antimicrobial activity of different tea varieties of Bangladesh. Asian Pac. J. Trop. Biomed. 2017, 7, 352–357. [Google Scholar] [CrossRef]
- Ananingsih, V.K.; Sharma, A.; Zhou, W. Green tea catechins during food processing and storage: A review on stability and detection. Food Res. Int. 2013, 50, 469–479. [Google Scholar] [CrossRef]
- Chen, C.-W.; Ho, C.-T. Antioxidant properties of polyphenols extracted from green and black teas. J. Food Lipids 1995, 2, 35–46. [Google Scholar] [CrossRef]
- Yang, Z.; Tu, Y.; Xia, H.; Jie, G.; Chen, X.; He, P. Suppression of free-radicals and protection against H2O2-induced oxidative damage in HPF-1 cell by oxidized phenolic compounds present in black tea. Food Chem. 2007, 105, 1349–1356. [Google Scholar] [CrossRef]
- Yang, Z.; Jie, G.; Dong, F.; Xu, Y.; Watanabe, N.; Tu, Y. Radical-scavenging abilities and antioxidant properties of theaflavins and their gallate esters in H2O2-mediated oxidative damage system in the HPF-1 cells. Toxicol. Vitr. 2008, 22, 1250–1256. [Google Scholar] [CrossRef]
- Shiraki, M.; Hara, Y.; Osawa, T.; Kumon, H.; Nakayama, T.; Kawakishi, S. Antioxidative and antimutagenic effects of theaflavins from black tea. Mutat. Res. Lett. 1994, 323, 29–34. [Google Scholar] [CrossRef]
- Miller, N.J.; Castelluccio, C.; Tijburg, L.; Rice-Evans, C. The antioxidant properties of theaflavins and their gallate esters—Radical scavengers or metal chelators? FEBS Lett. 1996, 392, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Bezard, E.; Zhao, B. Protective effect of green tea polyphenols on the SH-SY5Y cells against 6-OHDA induced apoptosis through ROS–NO pathway. Free Radic. Biol. Med. 2005, 39, 682–695. [Google Scholar] [CrossRef]
- Shanmugam, T.; Selvaraj, M.; Poomalai, S. Epigallocatechin gallate potentially abrogates fluoride induced lung oxidative stress, inflammation via Nrf2/Keap1 signaling pathway in rats: An in-vivo and in-silico study. Int. Immunopharmacol. 2016, 39, 128–139. [Google Scholar] [CrossRef]
- Young, J.F.; Dragsted, L.O.; Haraldsdóttir, J.; Daneshvar, B.; Kall, M.A.; Loft, S.; Nilsson, L.; Nielsen, S.E.; Mayer, B.; Skibsted, L.H.; et al. Green tea extract only affects markers of oxidative status postprandially: Lasting antioxidant effect of flavonoid-free diet. Br. J. Nutr. 2002, 87, 343–355. [Google Scholar] [CrossRef]
- Henning, S.M.; Niu, Y.; Lee, N.H.; Thames, G.D.; Minutti, R.R.; Wang, H.; Go, V.L.W.; Heber, D. Bioavailability and antioxidant activity of tea flavanols after consumption of green tea, black tea, or a green tea extract supplement. Am. J. Clin. Nutr. 2004, 80, 1558–1564. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Nah, J.; Chun, S.; Park, H.; Yang, S.E.; Min, W.K. In vivo antioxidant effect of green tea. Eur. J. Clin. Nutr. 2000, 54, 527–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benzie, I.F.F.; Szeto, Y.T.; Strain, J.J.; Tomlinson, B. Consumption of Green Tea Causes Rapid Increase in Plasma Antioxidant Power in Humans. Nutr. Cancer 1999, 34, 83–87. [Google Scholar] [CrossRef]
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mol. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free Radical Lipid Peroxidation: Mechanisms and Analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef]
- Truong, V.-L.; Jun, M.; Jeong, W.-S. Role of resveratrol in regulation of cellular defense systems against oxidative stress. BioFactors 2018, 44, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Alexander, D.M. Green tea catechins as inhibitors of oxidation of meat lipids. J. Food Lipids 1998, 5, 125–133. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, B.; Li, M.; Shen, S.; Xin, W. Studies on protective mechanisms of four components of green tea polyphenols against lipid peroxidation in synaptosomes. Biochim. Biophys. Acta-Lipid Lipid Metab. 1996, 1304, 210–222. [Google Scholar] [CrossRef]
- Hodgson, J.M.; Proudfoot, J.M.; Croft, K.D.; Puddey, I.B.; Mori, T.A.; Beilin, L.J. Comparison of the effects of black and green tea on in vitro lipoprotein oxidation in human serum. J. Sci. Food Agric. 1999, 79, 561–566. [Google Scholar] [CrossRef]
- Leung, L.K.; Su, Y.; Chen, R.; Zhang, Z.; Huang, Y.; Chen, Z.-Y. Theaflavins in Black Tea and Catechins in Green Tea Are Equally Effective Antioxidants. J. Nutr. 2001, 131, 2248–2251. [Google Scholar] [CrossRef]
- Yoshida, H.; Ishikawa, T.; Hosoai, H.; Suzukawa, M.; Ayaori, M.; Hisada, T.; Sawada, S.; Yonemura, A.; Higashi, K.; Ito, T.; et al. Inhibitory effect of tea flavonoids on the ability of cells to oxidize low density lipoprotein. Biochem. Pharmacol. 1999, 58, 1695–1703. [Google Scholar] [CrossRef]
- Murakami, C.; Hirakawa, Y.; Inui, H.; Nakano, Y.; Yoshida, H. Effect of Tea Catechins on Cellular Lipid Peroxidation and Cytotoxicity in HepG2 Cells. Biosci. Biotechnol. Biochem. 2002, 66, 1559–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Cai, S.; Li, J.; Xiong, L.; Tian, L.; Liu, J.; Huang, J.; Liu, Z. Neuroprotective Effects of Theaflavins Against Oxidative Stress-Induced Apoptosis in PC12 Cells. Neurochem. Res. 2016, 41, 3364–3372. [Google Scholar] [CrossRef]
- Fatima, M.; Kesharwani, R.K.; Misra, K.; Rizvi, S.I. Protective Effect of Theaflavin on Erythrocytes Subjected to In Vitro Oxidative Stress. Biochem. Res. Int. 2013, 2013, 649759. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.-J.; Ma, L.-P.; Hou, L.-F.; Zhou, B.; Yang, L.; Liu, Z.-L. Antioxidant effects of green tea polyphenols on free radical initiated peroxidation of rat liver microsomes. Chem. Phys. Lipids 2002, 120, 109–117. [Google Scholar] [CrossRef]
- Skrzydlewska, E.; Ostrowska, J.; Farbiszewski, R.; Michalak, K. Protective effect of green tea against lipid peroxidation in the rat liver, blood serum and the brain. Phytomedicine 2002, 9, 232–238. [Google Scholar] [CrossRef]
- Guo, S.; Yan, J.; Yang, T.; Yang, X.; Bezard, E.; Zhao, B. Protective Effects of Green Tea Polyphenols in the 6-OHDA Rat Model of Parkinson’s Disease Through Inhibition of ROS-NO Pathway. Biol. Psychiatry 2007, 62, 1353–1362. [Google Scholar] [CrossRef]
- Chen, J.-H.; Tipoe, G.L.; Liong, E.C.; So, H.S.H.; Leung, K.-M.; Tom, W.-M.; Fung, P.C.W.; Nanji, A.A. Green tea polyphenols prevent toxin-induced hepatotoxicity in mice by down-regulating inducible nitric oxide–derived prooxidants. Am. J. Clin. Nutr. 2004, 80, 742–751. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, S.; Castro, E.; Rocha-Pereira, P.; Rebelo, I.; Rocha, S.; Santos-Silva, A. The effect of green tea in oxidative stress. Clin. Nutr. 2006, 25, 790–796. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhou, Y.; Lyons, K.E.; Pahwa, R.; Reddy, M.B. Green Tea Consumption Reduces Oxidative Stress in Parkinson’s Disease Patients. J. Behav. Brain Sci. 2015, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.F.; Fisher, L.J.; Hara, Y.; Harris, T.; Mak, W.B.; Melton, L.D.; Packer, J.E. Green tea catechins partially protect DNA from ·OH radical-induced strand breaks and base damage through fast chemical repair of DNA radicals. Carcinogenesis 2001, 22, 1189–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Q.; Torii, Y.; Uchida, K.; Nakamura, Y.; Hara, Y.; Osawa, T. Black Tea Polyphenols, Theaflavins, Prevent Cellular DNA Damage by Inhibiting Oxidative Stress and Suppressing Cytochrome P450 1A1 in Cell Cultures. J. Agric. Food Chem. 2002, 50, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, J.; Xue, X.; Zhao, Y.; Lu, L.; Cui, M.; Miao, W.; Fan, S. Theaflavin ameliorates ionizing radiation-induced hematopoietic injury via the NRF2 pathway. Free Radic. Biol. Med. 2017, 113, 59–70. [Google Scholar] [CrossRef]
- Chen, C.H.; Liu, T.Z.; Chen, C.H.; Wong, C.H.; Chen, C.H.; Lu, F.J.; Chen, S.C. The efficacy of protective effects of tannic acid, gallic acid, ellagic acid, and propyl gallate against hydrogen peroxide-induced oxidative stress and DNA damages in IMR-90 cells. Mol. Nutr. Food Res. 2007, 51, 962–968. [Google Scholar] [CrossRef]
- Vidjaya Letchoumy, P.; Chandra Mohan, K.V.P.; Stegeman, J.J.; Gelboin, H.V.; Hara, Y.; Nagini, S. Pretreatment with black tea polyphenols modulates xenobiotic-metabolizing enzymes in an experimental oral carcinogenesis model. Oncol. Res. 2008, 17, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Kager, N.; Ferk, F.; Kundi, M.; Wagner, K.-H.; Mišík, M.; Knasmüller, S. Prevention of oxidative DNA damage in inner organs and lymphocytes of rats by green tea extract. Eur. J. Nutr. 2010, 49, 227–234. [Google Scholar] [CrossRef]
- Hakim, I.A.; Harris, R.B.; Brown, S.; Chow, H.-H.S.; Wiseman, S.; Agarwal, S.; Talbot, W. Effect of Increased Tea Consumption on Oxidative DNA Damage among Smokers: A Randomized Controlled Study. J. Nutr. 2003, 133, 3303–3309. [Google Scholar] [CrossRef] [Green Version]
- Hakim, I.A.; Harris, R.B.; Chow, H.H.S.; Dean, M.; Brown, S.; Ali, I.U. Effect of a 4-Month Tea Intervention on Oxidative DNA Damage among Heavy Smokers. Cancer Epidemiol. Biomark. Prev. 2004, 13, 242–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlett, B.S.; Stadtman, E.R. Protein Oxidation in Aging, Disease, and Oxidative Stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vayalil, P.K.; Mittal, A.; Hara, Y.; Elmets, C.A.; Katiyar, S.K. Green Tea Polyphenols Prevent Ultraviolet Light-Induced Oxidative Damage and Matrix Metalloproteinases Expression in Mouse Skin. J. Invest. Dermatol. 2004, 122, 1480–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özyurt, H.; Luna, C.; Estévez, M. Redox chemistry of the molecular interactions between tea catechins and human serum proteins under simulated hyperglycemic conditions. Food Funct. 2016, 7, 1390–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, J.; Assunção, M.; Lukoyanov, N.; Cardoso, A.; Carvalho, F.; Andrade, J.P. Protective effects of a catechin-rich extract on the hippocampal formation and spatial memory in aging rats. Behav. Brain Res. 2013, 246, 94–102. [Google Scholar] [CrossRef]
- Rice-Evans, C. Implications of the Mechanisms of Action of Tea Polyphenols as Antioxidants in vitro for Chemoprevention in Humans. Proc. Soc. Exp. Biol. Med. 1999, 220, 262–266. [Google Scholar] [CrossRef]
- Pannala, A.; Rice-Evans, C.A.; Halliwell, B.; Singh, S. Inhibition of Peroxynitrite-Mediated Tyrosine Nitration by Catechin Polyphenols. Biochem. Biophys. Res. Commun. 1997, 232, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Mazzon, E.; Muià, C.; Paola, R.D.; Genovese, T.; Menegazzi, M.; De Sarro, A.; Suzuki, H.; Cuzzocrea, S. Green tea polyphenol extract attenuates colon injury induced by experimental colitis. Free Radic. Res. 2005, 39, 1017–1025. [Google Scholar] [CrossRef]
- Pham, A.N.; Xing, G.; Miller, C.J.; Waite, T.D. Fenton-like copper redox chemistry revisited: Hydrogen peroxide and superoxide mediation of copper-catalyzed oxidant production. J. Catal. 2013, 301, 54–64. [Google Scholar] [CrossRef]
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In Vitro Antioxidant versus Metal Ion Chelating Properties of Flavonoids: A Structure-Activity Investigation. PLoS ONE 2016, 11, e0165575. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, N.; Ohnishi, M.; Imamura, M.; Furuno, K. Differences in Antioxidative Efficiency of Catechins in Various Metal-Induced Lipid Peroxidations in Cultured Hepatocytes. J. Health Sci. 2001, 47, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Grinberg, L.N.; Newmark, H.; Kitrossky, N.; Rahamim, E.; Chevion, M.; Rachmilewitz, E.A. Protective effects of tea polyphenols against oxidative damage to red blood cells. Biochem. Pharmacol. 1997, 54, 973–978. [Google Scholar] [CrossRef]
- Andjelković, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Granato, D.; Grevink, R.; Zielinski, A.A.F.; Nunes, D.S.; van Ruth, S.M. Analytical Strategy Coupled with Response Surface Methodology to Maximize the Extraction of Antioxidants from Ternary Mixtures of Green, Yellow, and Red Teas (Camellia sinensis var. sinensis). J. Agric. Food Chem. 2014, 62, 10283–10296. [Google Scholar] [CrossRef] [PubMed]
- Carloni, P.; Tiano, L.; Padella, L.; Bacchetti, T.; Customu, C.; Kay, A.; Damiani, E. Antioxidant activity of white, green and black tea obtained from the same tea cultivar. Food Res. Int. 2013, 53, 900–908. [Google Scholar] [CrossRef]
- Zuo, X.; Tian, C.; Zhao, N.; Ren, W.; Meng, Y.; Jin, X.; Zhang, Y.; Ding, S.; Ying, C.; Ye, X. Tea polyphenols alleviate high fat and high glucose-induced endothelial hyperpermeability by attenuating ROS production via NADPH oxidase pathway. BMC Res. Notes 2014, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Rashidinejad, A.; Birch, E.J.; Everett, D.W. Green tea catechins suppress xanthine oxidase activity in dairy products: An improved HPLC analysis. J. Food Compost. Anal. 2016, 48, 120–127. [Google Scholar] [CrossRef]
- Lin, J.-K.; Chen, P.-C.; Ho, C.-T.; Lin-Shiau, S.-Y. Inhibition of Xanthine Oxidase and Suppression of Intracellular Reactive Oxygen Species in HL-60 Cells by Theaflavin-3,3′-digallate, (−)-Epigallocatechin-3-gallate, and Propyl Gallate. J. Agric. Food Chem. 2000, 48, 2736–2743. [Google Scholar] [CrossRef]
- Chan, M.M.-Y.; Fong, D.; Ho, C.-T.; Huang, H.-I. Inhibition of Inducible Nitric Oxide Synthase Gene Expression and Enzyme Activity by Epigallocatechin Gallate, a Natural Product from Green Tea. Biochem. Pharmacol. 1997, 54, 1281–1286. [Google Scholar] [CrossRef]
- Lin, Y.-L.; Lin, J.-K. (−)-Epigallocatechin-3-gallate Blocks the Induction of Nitric Oxide Synthase by Down-Regulating Lipopolysaccharide-Induced Activity of Transcription Factor Nuclear Factor-κB. Mol. Pharmacol. 1997, 52, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Tsai, S.-H.; Lin-Shiau, S.-Y.; Ho, C.-T.; Jen-Kun, L. Theaflavin-3,3′-digallate from black tea blocks the nitric oxide synthase by down-regulating the activation of NF-κB in macrophages. Eur. J. Pharmacol. 1999, 367, 379–388. [Google Scholar] [CrossRef]
- Adhikary, B.; Yadav, S.K.; Chand, S.; Bandyopadhyay, S.K.; Chattopadhyay, S. Black tea and theaflavins suppress various inflammatory modulators and i-NOS mediated nitric oxide synthesis during gastric ulcer healing. Free Radic. Res. 2011, 45, 767–778. [Google Scholar] [CrossRef]
- Hong, J.; Smith, T.J.; Ho, C.-T.; August, D.A.; Yang, C.S. Effects of purified green and black tea polyphenols on cyclooxygenase- and lipoxygenase-dependent metabolism of arachidonic acid in human colon mucosa and colon tumor tissues. Biochem. Pharmacol. 2001, 62, 1175–1183. [Google Scholar] [CrossRef]
- Agarwal, R.; Katiyar, S.K.; Khan, S.G.; Mukhtar, H. Protection against ultraviolet B radiation-induced effects in the skin of SKH-1 hairless mice by a polyphenolic fraction isolated from green tea. Photochem. Photobiol. 1993, 58, 695–700. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Agarwal, R.; Wood, G.S.; Mukhtar, H. Inhibition of 12-O-Tetradecanoylphorbol-13-acetate-caused Tumor Promotion in 7,12-Dimethylbenz[a]anthracene-initiated SENCAR Mouse Skin by a Polyphenolic Fraction Isolated from Green Tea. Cancer Res. 1992, 52, 6890–6897. [Google Scholar]
- Katiyar, S.K.; Mukhtar, H. Inhibition of phorbol ester tumor promoter 12-O-tetradecanoylphorbol-13-acetate-caused inflammatory responses in SENCAR mouse skin by black tea polyphenols. Carcinogenesis 1997, 18, 1911–1916. [Google Scholar] [CrossRef] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Masella, R.; Di Benedetto, R.; Varì, R.; Filesi, C.; Giovannini, C. Novel mechanisms of natural antioxidant compounds in biological systems: Involvement of glutathione and glutathione-related enzymes. J. Nutr. Biochem. 2005, 16, 577–586. [Google Scholar] [CrossRef]
- Steele, V.E.; Kelloff, G.J.; Balentine, D.; Boone, C.W.; Mehta, R.; Bagheri, D.; Sigman, C.C.; Zhu, S.; Sharma, S. Comparative chemopreventive mechanisms of green tea, black tea and selected polyphenol extracts measured by in vitro bioassays. Carcinogenesis 2000, 21, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, X.; Jiao, H.; Zhao, B. Effect of Tea Catechins on the Change of Glutathione Levels Caused by Pb++ in PC12 Cells. Chem. Res. Toxicol. 2004, 17, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.J.; Hsieh, M.T.; Wu, C.R.; Wood, W.G.; Chen, Y.F. Green Tea Extract Ameliorates Learning and Memory Deficits in Ischemic Rats via Its Active Component Polyphenol Epigallocatechin-3-gallate by Modulation of Oxidative Stress and Neuroinflammation. Evid. Based Complement. Alternat. Med. 2012, 2012, 163106. [Google Scholar] [CrossRef] [PubMed]
- Oz, H.S.; McClain, C.J.; Nagasawa, H.T.; Ray, M.B.; de Villiers, W.J.S.; Chen, T.S. Diverse antioxidants protect against acetaminophen hepatotoxicity. J. Biochem. Mol. Toxicol. 2005, 18, 361–368. [Google Scholar] [CrossRef]
- Oz, H.S.; Chen, T.S. Green-tea Polyphenols Downregulate Cyclooxygenase and Bcl-2 Activity in Acetaminophen-induced Hepatotoxicity. Dig. Dis. Sci. 2008, 53, 2980–2988. [Google Scholar] [CrossRef]
- Fu, Y.; Zheng, S.; Lu, S.C.; Chen, A. Epigallocatechin-3-gallate Inhibits Growth of Activated Hepatic Stellate Cells by Enhancing the Capacity of Glutathione Synthesis. Mol. Pharmacol. 2008, 73, 1465–1473. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Betts, N.M.; Mulugeta, A.; Tong, C.; Newman, E.; Lyons, T.J. Green tea supplementation increases glutathione and plasma antioxidant capacity in adults with the metabolic syndrome. Nutr. Res. 2013, 33, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Katiyar, S.K.; Afaq, F.; Perez, A.; Mukhtar, H. Green tea polyphenol (–)-epigallocatechin-3-gallate treatment of human skin inhibits ultraviolet radiation-induced oxidative stress. Carcinogenesis 2001, 22, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Fang, Y.; Wei, S.M. Effect and mechanism of epigallocatechin-3-gallate (EGCG). against the hydrogen peroxide-induced oxidative damage in human dermal fibroblasts. J. Cosmet. Sci. 2013, 64, 35–44. [Google Scholar] [PubMed]
- Sabu, M.C.; Smitha, K.; Ramadasan, K. Anti-diabetic activity of green tea polyphenols and their role in reducing oxidative stress in experimental diabetes. J. Ethnopharmacol. 2002, 83, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Negishi, H.; Xu, J.-W.; Ikeda, K.; Njelekela, M.; Nara, Y.; Yamori, Y. Black and Green Tea Polyphenols Attenuate Blood Pressure Increases in Stroke-Prone Spontaneously Hypertensive Rats. J. Nutr. 2004, 134, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.A.; Radwan, N.M.; Aboul Ezz, H.S.; Salama, N.A. The antioxidant effect of Green Tea Mega EGCG against electromagnetic radiation-induced oxidative stress in the hippocampus and striatum of rats. Electromagn. Biol. Med. 2017, 36, 63–73. [Google Scholar] [CrossRef]
- Jeon, S.E.; Choi-Kwon, S.; Park, K.A.; Lee, H.J.; Park, M.S.; Lee, J.H.; Kwon, S.B.; Park, K.C. Dietary supplementation of (+)-catechin protects against UVB-induced skin damage by modulating antioxidant enzyme activities. Photodermatol. Photoimmunol. Photomed. 2003, 19, 235–241. [Google Scholar] [CrossRef]
- Zhang, L.; Gui, S.; Wang, J.; Chen, Q.; Zeng, J.; Liu, A.; Chen, Z.; Lu, X. Oral administration of green tea polyphenols (TP) improves ileal injury and intestinal flora disorder in mice with Salmonella typhimurium infection via resisting inflammation, enhancing antioxidant action and preserving tight junction. J. Funct. Foods 2020, 64, 103654. [Google Scholar] [CrossRef]
- El-Beshbishy, H.A.; Tork, O.M.; El-Bab, M.F.; Autifi, M.A. Antioxidant and antiapoptotic effects of green tea polyphenols against azathioprine-induced liver injury in rats. Pathophysiology 2011, 18, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Chandronitha, C.; Ananthi, S.; Ramakrishnan, G.; Lakshmisundaram, R.; Gayathri, V.; Vasanthi, H.R. Protective role of tannin-rich fraction of Camellia sinensis in tissue arsenic burden in Sprague Dawley rats. Hum. Exp. Toxicol. 2010, 29, 705–719. [Google Scholar] [CrossRef]
- Han, X.-D.; Zhang, Y.-Y.; Wang, K.-L.; Huang, Y.-P.; Yang, Z.-B.; Liu, Z. The involvement of Nrf2 in the protective effects of (−)-Epigallocatechin-3-gallate (EGCG) on NaAsO(2)-induced hepatotoxicity. Oncotarget 2017, 8, 65302–65312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugisawa, A.; Kimura, M.; Fenech, M.; Umegaki, K. Anti-genotoxic effects of tea catechins against reactive oxygen species in human lymphoblastoid cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2004, 559, 97–103. [Google Scholar] [CrossRef]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedesco, I.; Spagnuolo, C.; Russo, G.L.; Russo, M.; Cervellera, C.; Moccia, S. The Pro-Oxidant Activity of Red Wine Polyphenols Induces an Adaptive Antioxidant Response in Human Erythrocytes. Antioxidants 2021, 10, 800. [Google Scholar] [CrossRef]
- Raza, H.; John, A. Green tea polyphenol epigallocatechin-3-gallate differentially modulates oxidative stress in PC12 cell compartments. Toxicol. Appl. Pharmacol. 2005, 207, 212–220. [Google Scholar] [CrossRef]
- Schuck, A.G.; Ausubel, M.B.; Zuckerbraun, H.L.; Babich, H. Theaflavin-3,3′-digallate, a component of black tea: An inducer of oxidative stress and apoptosis. Toxicol. Vitr. 2008, 22, 598–609. [Google Scholar] [CrossRef]
- Babich, H.; Schuck, A.G.; Weisburg, J.H.; Zuckerbraun, H.L. Research Strategies in the Study of the Pro-Oxidant Nature of Polyphenol Nutraceuticals. J. Toxicol. 2011, 2011, 467305. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.-S.; Zhou, B.; Yang, L.; Wu, L.-M.; Liu, Z.-L. Antioxidant synergism of tea polyphenols and α-tocopherol against free radical induced peroxidation of linoleic acid in solution. J. Chem. Soc. Perkin Trans. 1998, 4, 911–916. [Google Scholar] [CrossRef]
- Zhou, B.; Jia, Z.-S.; Chen, Z.-H.; Yang, L.; Wu, L.-M.; Liu, Z.-L. Synergistic antioxidant effect of green tea polyphenols with α-tocopherol on free radical initiated peroxidation of linoleic acid in micelles. J. Chem. Soc. Perkin Trans. 2000, 4, 785–791. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Ma, L.-P.; Zhou, B.; Yang, L.; Liu, Z.-L. Antioxidative effects of green tea polyphenols on free radical initiated and photosensitized peroxidation of human low density lipoprotein. Chem. Phys. Lipids 2000, 106, 53–63. [Google Scholar] [CrossRef]
- Zhou, B.; Chen, Z.; Chen, Y.; Jia, Z.; Jia, Y.; Zeng, L.; Wu, L.; Yang, L.; Liu, Z.L. Kinetic EPR studies on bio-antioxidants. Appl. Magn. Reson. 2000, 18, 397–406. [Google Scholar] [CrossRef]
- Zhou, B.; Wu, L.-M.; Yang, L.; Liu, Z.-L. Evidence for α-tocopherol regeneration reaction of green tea polyphenols in SDS micelles. Free Radic. Biol. Med. 2005, 38, 78–84. [Google Scholar] [CrossRef]
- Dai, F.; Chen, W.-F.; Zhou, B. Antioxidant synergism of green tea polyphenols with α-tocopherol and l-ascorbic acid in SDS micelles. Biochimie 2008, 90, 1499–1505. [Google Scholar] [CrossRef]
- Keum, Y.-S.; Jeong, W.-S.; Tony Kong, A.N. Chemoprevention by isothiocyanates and their underlying molecular signaling mechanisms. Mutat. Res. 2004, 555, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.; Singer, P.; Halpern, Z.; Bruck, R. Polyphenols in the treatment of inflammatory bowel disease and acute pancreatitis. Gut 2007, 56, 426–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, H.-K.; Surh, Y.-J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Vomund, S.; Schäfer, A.; Parnham, M.J.; Brüne, B.; von Knethen, A. Nrf2, the Master Regulator of Anti-Oxidative Responses. Int. J. Mol. Sci. 2017, 18, 2772. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, E.H.; Suzuki, T.; Funayama, R.; Nagashima, T.; Hayashi, M.; Sekine, H.; Tanaka, N.; Moriguchi, T.; Motohashi, H.; Nakayama, K.; et al. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat. Commun. 2016, 7, 11624. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, P.; Suchalatha, S.; Babu, P.V.A.; Devi, R.S.; Narayan, S.; Sabitha, K.E.; Shyamala Devi, C.S. Chemopreventive and therapeutic modulation of green tea polyphenols on drug metabolizing enzymes in 4-Nitroquinoline 1-oxide induced oral cancer. Chem. Biol. Interact. 2008, 172, 224–234. [Google Scholar] [CrossRef]
- Chandra Mohan, K.V.P.; Hara, Y.; Abraham, S.K.; Nagini, S. Comparative evaluation of the chemopreventive efficacy of green and black tea polyphenols in the hamster buccal pouch carcinogenesis model. Clin. Biochem. 2005, 38, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sun, C.-K.; Han, G.-Z.; Peng, J.-Y.; Li, Y.; Liu, Y.-X.; Lv, Y.-Y.; Liu, K.-X.; Zhou, Q.; Sun, H.-J. Protective effect of tea polyphenols against paracetamol-induced hepatotoxicity in mice is significantly correlated with cytochrome P450 suppression. World J. Gastroenterol. 2009, 15, 1829–1835. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Hao, S.; Chen, X.; Li, Y.; Yin, Z.; Zou, Y.; Song, X.; Li, L.; Ye, G.; Zhao, L.; et al. Epigallocatechin-3-gallate protects immunity and liver drug-metabolism function in mice loaded with restraint stress. Biomed. Pharmacother. 2020, 129, 110418. [Google Scholar] [CrossRef]
- Krishnan, R.; Raghunathan, R.; Maru, G.B. Effect of polymeric black tea polyphenols on benzo(a)pyrene [B(a)P]-induced cytochrome P4501A1 and 1A2 in mice. Xenobiotica 2005, 35, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Wang, H.; Li, G.X.; Yang, Z.; Guan, F.; Jin, H. Cancer prevention by tea: Evidence from laboratory studies. Pharmacol. Res. 2011, 64, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Jiao, J.J.; Duh, J.L.; Gudehithlu, K.; Tan, T.H.; Kong, A.N. Activation of mitogen-activated protein kinases by green tea polyphenols: Potential signaling pathways in the regulation of antioxidant-responsive element-mediated phase II enzyme gene expression. Carcinogenesis 1997, 18, 451–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Yu, R.; Owuor, E.D.; Kong, A.N. Activation of antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch. Pharm. Res. 2000, 23, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Na, H.-K.; Kim, E.-H.; Jung, J.-H.; Lee, H.-H.; Hyun, J.-W.; Surh, Y.-J. (−)-Epigallocatechin gallate induces Nrf2-mediated antioxidant enzyme expression via activation of PI3K and ERK in human mammary epithelial cells. Arch. Biochem. Biophys. 2008, 476, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-P.; Wilson, K.; Kawa, A.; Raner, G.M. Effects of green tea extracts on gene expression in HepG2 and Cal-27 cells. Food Chem. Toxicol. 2006, 44, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.; Xu, C.; Hu, R.; Jain, M.R.; Nair, S.; Lin, W.; Yang, C.S.; Chan, J.Y.; Kong, A.N.T. Comparison of (−)-Epigallocatechin-3-Gallate Elicited Liver and Small Intestine Gene Expression Profiles Between C57BL/6J Mice and C57BL/6J/Nrf2 (−/−) Mice. Pharm. Res. 2005, 22, 1805–1820. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Mi, Y.; Wang, Y.; Li, R.; Huang, S.; Li, X.; Liu, X. Neuroprotective action of tea polyphenols on oxidative stress-induced apoptosis through the activation of the TrkB/CREB/BDNF pathway and Keap1/Nrf2 signaling pathway in SH-SY5Y cells and mice brain. Food Funct. 2017, 8, 4421–4432. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.; Mi, Y.; Fan, R.; Li, R.; Wang, Y.; Li, X.; Huang, S.; Liu, X. Tea polyphenols ameliorate hydrogen peroxide- and constant darkness-triggered oxidative stress via modulating the Keap1/Nrf2 transcriptional signaling pathway in HepG2 cells and mice liver. RSC Adv. 2017, 7, 32198–32208. [Google Scholar] [CrossRef] [Green Version]
- Surh, Y.-J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer. 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Stein, B.; Baldwin, A.S., Jr.; Ballard, D.W.; Greene, W.C.; Angel, P.; Herrlich, P. Cross-coupling of the NF-kappa B p65 and Fos/Jun transcription factors produces potentiated biological function. EMBO J. 1993, 12, 3879–3891. [Google Scholar] [CrossRef]
- Chinenov, Y.; Kerppola, T.K. Close encounters of many kinds: Fos-Jun interactions that mediate transcription regulatory specificity. Oncogene 2001, 20, 2438–2452. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Oz, H.; Barve, S.; DeVilliers, W.; McClain, C.; Varilek, G. The green tea polyphenol EGCG blocks nuclear factor-kappa B activation by inhibiting I kappa B kinase activity in the intestinal epithelial cell line IEC-6. Mol. Pharmacol. 2001, 60, 528–533. [Google Scholar]
- Hafeez, B.B.; Ahmed, S.; Wang, N.; Gupta, S.; Zhang, A.; Haqqi, T.M. Green tea polyphenols-induced apoptosis in human osteosarcoma SAOS-2 cells involves a caspase-dependent mechanism with downregulation of nuclear factor-κB. Toxicol. Appl. Pharmacol. 2006, 216, 11–19. [Google Scholar] [CrossRef]
- Rasheed, Z.; Anbazhagan, A.N.; Akhtar, N.; Ramamurthy, S.; Voss, F.R.; Haqqi, T.M. Green tea polyphenol epigallocatechin-3-gallate inhibits advanced glycation end product-induced expression of tumor necrosis factor-α and matrix metalloproteinase-13 in human chondrocytes. Arthritis Res. Ther. 2009, 11, R71. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, D.S.; Catravas, J.D.; Odoms, K.; Denenberg, A.; Malhotra, V.; Wong, H.R. Epigallocatechin-3-gallate, a Green Tea–Derived Polyphenol, Inhibits IL-1β-Dependent Proinflammatory Signal Transduction in Cultured Respiratory Epithelial Cells. J. Nutr. 2004, 134, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.Y.; Kim, S.H.; Jeong, H.J.; Kim, S.Y.; Shin, T.Y.; Um, J.Y.; Hong, S.H.; Kim, H.M. Epigallocatechin-3-Gallate Inhibits Secretion of TNF-α, IL-6 and IL-8 through the Attenuation of ERK and NF-κB in HMC-1 Cells. Int. Arch. Allergy Immunol. 2007, 142, 335–344. [Google Scholar] [CrossRef]
- Singh, R.; Akhtar, N.; Haqqi, T.M. Green tea polyphenol epigallocatechi3-gallate: Inflammation and arthritis. Life Sci. 2010, 86, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.Y.; Huang, C.; Meng, X.; Dong, Z.; Yang, C.S. Inhibition of Activator Protein 1 Activity and Cell Growth by Purified Green Tea and Black Tea Polyphenols in H-ras-transformed Cells. Cancer Res. 1999, 59, 4610–4617. [Google Scholar] [PubMed]
- Chiou, Y.-S.; Huang, Q.; Ho, C.-T.; Wang, Y.-J.; Pan, M.-H. Directly interact with Keap1 and LPS is involved in the anti-inflammatory mechanisms of (−)-epicatechin-3-gallate in LPS-induced macrophages and endotoxemia. Free Radic. Biol. Med. 2016, 94, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-W.; Zhang, Y.; Zhang, L.; Li, X.; Yu, J.-B.; Zhang, H.-T.; Tan, B.-B.; Jiang, L.-H.; Wang, Y.-X.; Liang, Y.; et al. Protective effect of tea polyphenols on renal ischemia/reperfusion injury via suppressing the activation of TLR4/NF-κB p65 signal pathway. Gene 2014, 542, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.; Rosenstiel, P.; Hampe, J.; Nikolaus, S.; Groessner, B.; Schottelius, A.; Kühbacher, T.; Hämling, J.; Fölsch, U.R.; Seegert, D. Activation of signal transducer and activator of transcription (STAT) 1 in human chronic inflammatory bowel disease. Gut 2002, 51, 379–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.Y. STAT3 in immune responses and inflammatory bowel diseases. Cell Res. 2006, 16, 214–219. [Google Scholar] [CrossRef] [Green Version]
- Zykova, T.A.; Zhang, Y.; Zhu, F.; Bode, A.M.; Dong, Z. The signal transduction networks required for phosphorylation of STAT1 at Ser727 in mouse epidermal JB6 cells in the UVB response and inhibitory mechanisms of tea polyphenols. Carcinogenesis 2005, 26, 331–342. [Google Scholar] [CrossRef]
- Tedeschi, E.; Suzuki, H.; Menegazzi, M. Antiinflammatory Action of EGCG, the Main Component of Green Tea, through STAT-1 Inhibition. Ann. N. Y. Acad. Sci. 2002, 973, 435–437. [Google Scholar] [CrossRef]
- Watson, J.L.; Ansari, S.; Cameron, H.; Wang, A.; Akhtar, M.; McKay, D.M. Green tea polyphenol (−)-epigallocatechin gallate blocks epithelial barrier dysfunction provoked by IFN-γ but not by IL-4. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, 954–961. [Google Scholar] [CrossRef]
- Cheng, C.-W.; Shieh, P.-C.; Lin, Y.-C.; Chen, Y.-J.; Lin, Y.-H.; Kuo, D.-H.; Liu, J.-Y.; Kao, J.-Y.; Kao, M.-C.; Way, T.-D. Indoleamine 2,3-Dioxygenase, an Immunomodulatory Protein, Is Suppressed by (−)-Epigallocatechin-3-gallate via Blocking of γ-Interferon-Induced JAK-PKC-δ-STAT1 Signaling in Human Oral Cancer Cells. J. Agric. Food Chem. 2010, 58, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Darra, E.; Shoji, K.; Mariotto, S.; Suzuki, H. Protective effect of epigallocatechin-3-gallate on ischemia/reperfusion-induced injuries in the heart: STAT1 silencing flavonoid. Genes Nutr. 2007, 2, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Di Paola, R.; Mazzon, E.; Muià, C.; Genovese, T.; Menegazzi, M.; Zaffini, R.; Suzuki, H.; Cuzzocrea, S. Green tea polyphenol extract attenuates lung injury in experimental model of carrageenan-induced pleurisy in mice. Respir. Res. 2005, 6, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.; Hochstetter, D.; Yao, L.; Zhao, Y.; Zhou, J.; Wang, Y.; Xu, P. Green Tea Polyphenol (−)-Epigallocatechin Gallate (EGCG) Attenuates Neuroinflammation in Palmitic Acid-Stimulated BV-2 Microglia and High-Fat Diet-Induced Obese Mice. Int. J. Mol. Sci. 2019, 20, 5081. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong, V.-L.; Jeong, W.-S. Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship. Int. J. Mol. Sci. 2021, 22, 9109. https://doi.org/10.3390/ijms22179109
Truong V-L, Jeong W-S. Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship. International Journal of Molecular Sciences. 2021; 22(17):9109. https://doi.org/10.3390/ijms22179109
Chicago/Turabian StyleTruong, Van-Long, and Woo-Sik Jeong. 2021. "Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship" International Journal of Molecular Sciences 22, no. 17: 9109. https://doi.org/10.3390/ijms22179109