
Anti-Cancer Activity of Phytochemicals Targeting Hypoxia-Inducible Factor-1 Alpha


Abstract
1. Introduction
2. Alkaloid and Organosulfur Compounds from Natural Sources
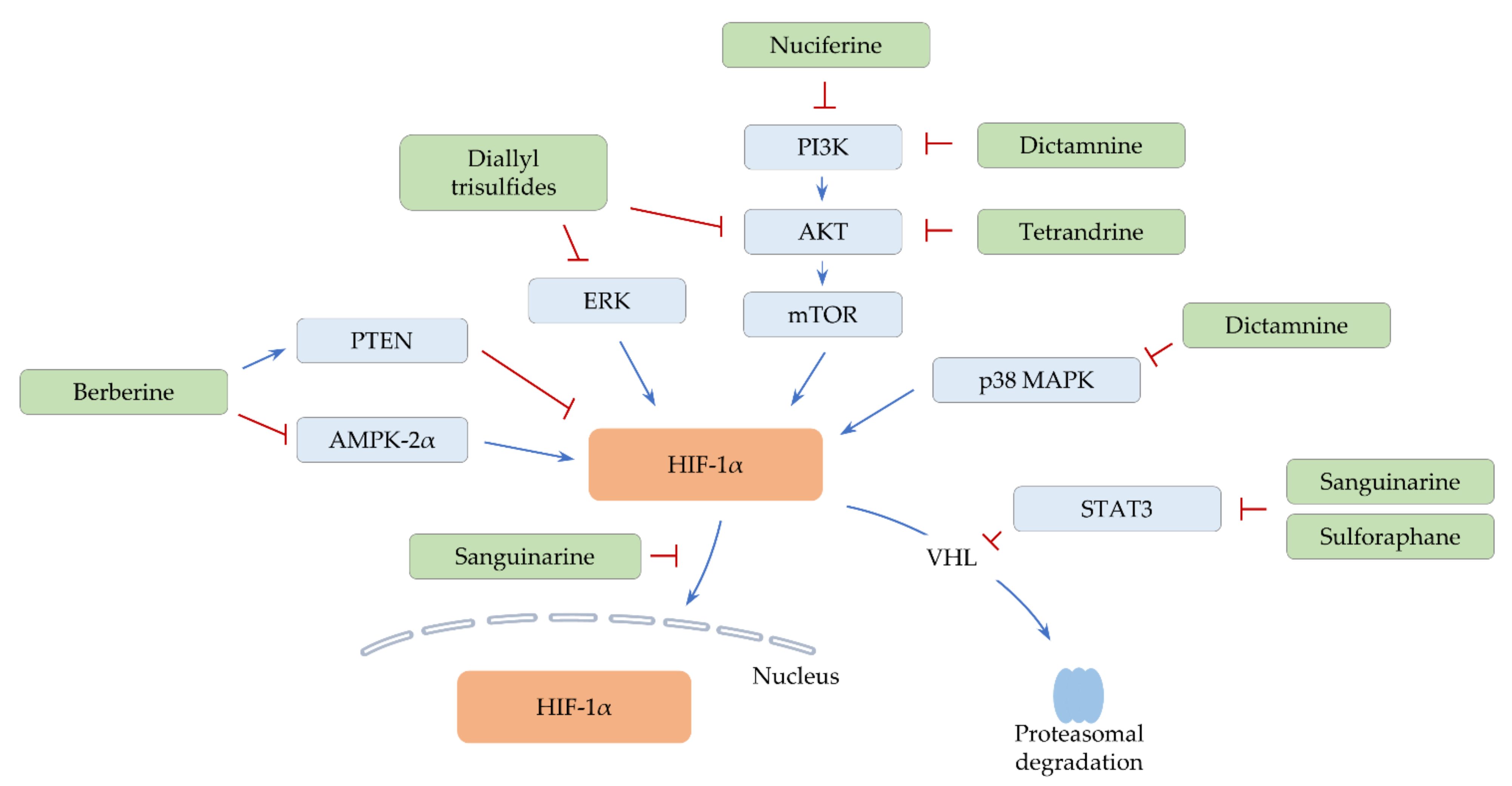
2.1. Alkaloids
2.1.1. Berberine

{kind=link}
{kind=link}
{kind=link}
| Compound | Compound Class | Type of Cancer | In Vitro Testing (Effective Concentrations, Cell Line, Culture Condition (Normoxia/Hypoxia)) | In Vivo Experiment Model (Dose and Administration Route) | Ref. |
|---|---|---|---|---|---|
| Alkaloids | |||||
| Berberine | Benzylisoquinoline | Breast cancer | 10–160 μM (human MCF-7 cell line), hypoxia (1% O2) | Subcutaneous injection of MCF-7 cells (5–200 mg/kg, oral) | [33] |
| Cyclopamine tartrate | Derivative of cyclopamine (veratrum alkaloid) | Lung cancer | 15–25 μM (human H1299 cell line), normoxia | Subcutaneous injection of H1299 cells (7.5 mg/kg, intravenous) | [36] |
| Dictamnine | Furanoquinoline alkaloid | Colorectal cancer | 10–100 μM (human HCT-116 cell line), hypoxia (1% O2) | Subcutaneous injection of HCT-116 cells (50–100 mg/kg, oral) | [37] |
| Nuciferine | Aporphine alkaloid | Colorectal cancer, Lung cancer | 4–48 μM (drug-sensitive or -resistant human HCT-8 and A549 cell lines), normoxia | Subcutaneous injection of drug-resistant A549 cells (7.5 mg/kg, intraperitoneal) | [38] |
| Sanguinarine | Benzophenanthridine alkaloid | Breast cancer | 2–4 μM (human MDA-MB-231 cell line), hypoxia (1% O2, cobalt (II) chloride (CoCl2)) | - | [39] |
| Breast cancer | 0.5–1 μM (human T47D and MDA-MB-231 cell lines), hypoxia (1% O2, CoCl2) | Subcutaneous injection of MDA-MB-231 cells (1.25–5 mg/kg, oral) | [40] | ||
| Hepatocellular carcinoma | 0.5–2 μM (human HepG2 and SMMC-7721 cell lines), hypoxia (1% O2, CoCl2) | Subcutaneous injection of HepG2 and SMMC-7721 cells (1.25–5 mg/kg, oral) | [41] | ||
| Tetrandrine | Benzylisoquinoline | Lung cancer | 5–10 μM (human A549 cell line), normoxia | - | [42] |
| Organosulfurs | |||||
| Diallyl trisulfide | Organic trisulfide | Breast cancer | 2.5–10 μM (human MDA-MB-231 cell line), hypoxia (1% O2) | Microinjection of MDA-MB-231 cells into perivitelline space of zebrafish embryos (2.5–10 μM), Tail vein or orthotopic injections of MDA-MB-231 cells (25–50 mg/kg, oral) | [43] |
| Sulforaphane | Isothiocyanate | Bladder cancer | 5–20 μM (human RT112 and RT4 cell lines), hypoxia (2% O2) | - | [44] |
| Hepatocellular carcinoma | 5–20 μM (human HepG2 cell line), hypoxia (CoCl2) | HepG2-bearing chick chorioallantoic membrane (CAM) assay (20 μM) | [45] | ||
2.1.2. Dictamnine
2.1.3. Nuciferine
2.1.4. Sanguinarine
2.1.5. Tetrandrine
2.2. Organosulfurs
2.2.1. Diallyl Trisulfides
2.2.2. Sulforaphane
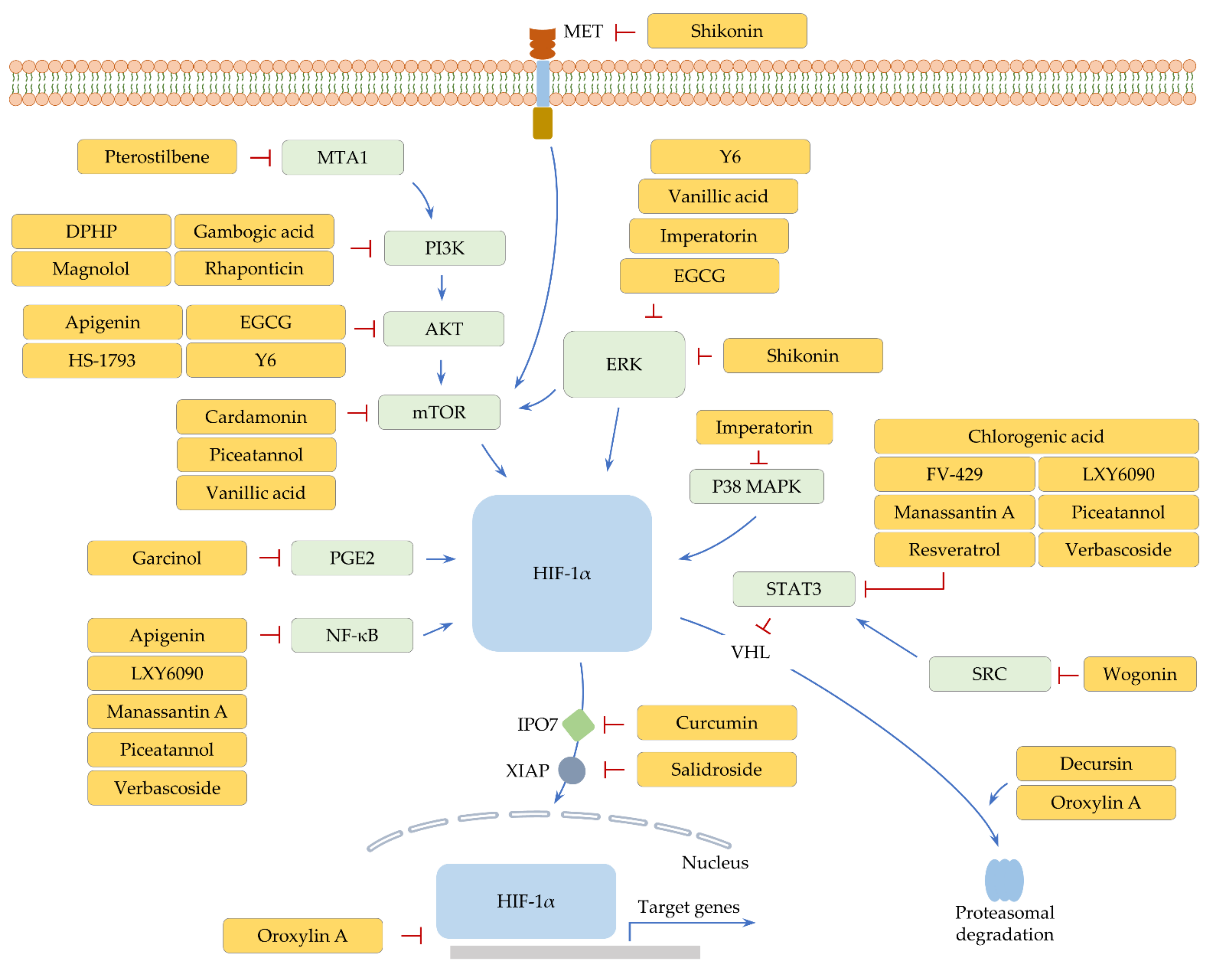
3. Natural Polyphenolic Compounds
3.1. Flavonoids
3.1.1. Apigenin
3.1.2. Cardamonin, Epigallocatechin-3-Gallate, and Y6
3.1.3. FV-429 and Wogonin
3.1.4. Oroxylin A
3.2. Lignans, Phenolic Acids, and Stilbenes
3.2.1. HS-1793 and Resveratrol
3.2.2. LXY6090 and Manassantin A
3.2.3. Magnolol
3.2.4. Piceatannol and Vanillic Acid
3.2.5. Pterostilbene
3.2.6. Rhaponticin
3.3. Other Polyphenols
3.3.1. Chlorogenic Acid
3.3.2. Curcumin
3.3.3. Decursin
3.3.4. DPHP, Garcinol, Imperatorin, Shikonin, and Verbascoside
3.3.5. Gambogic Acid
3.3.6. Salidroside
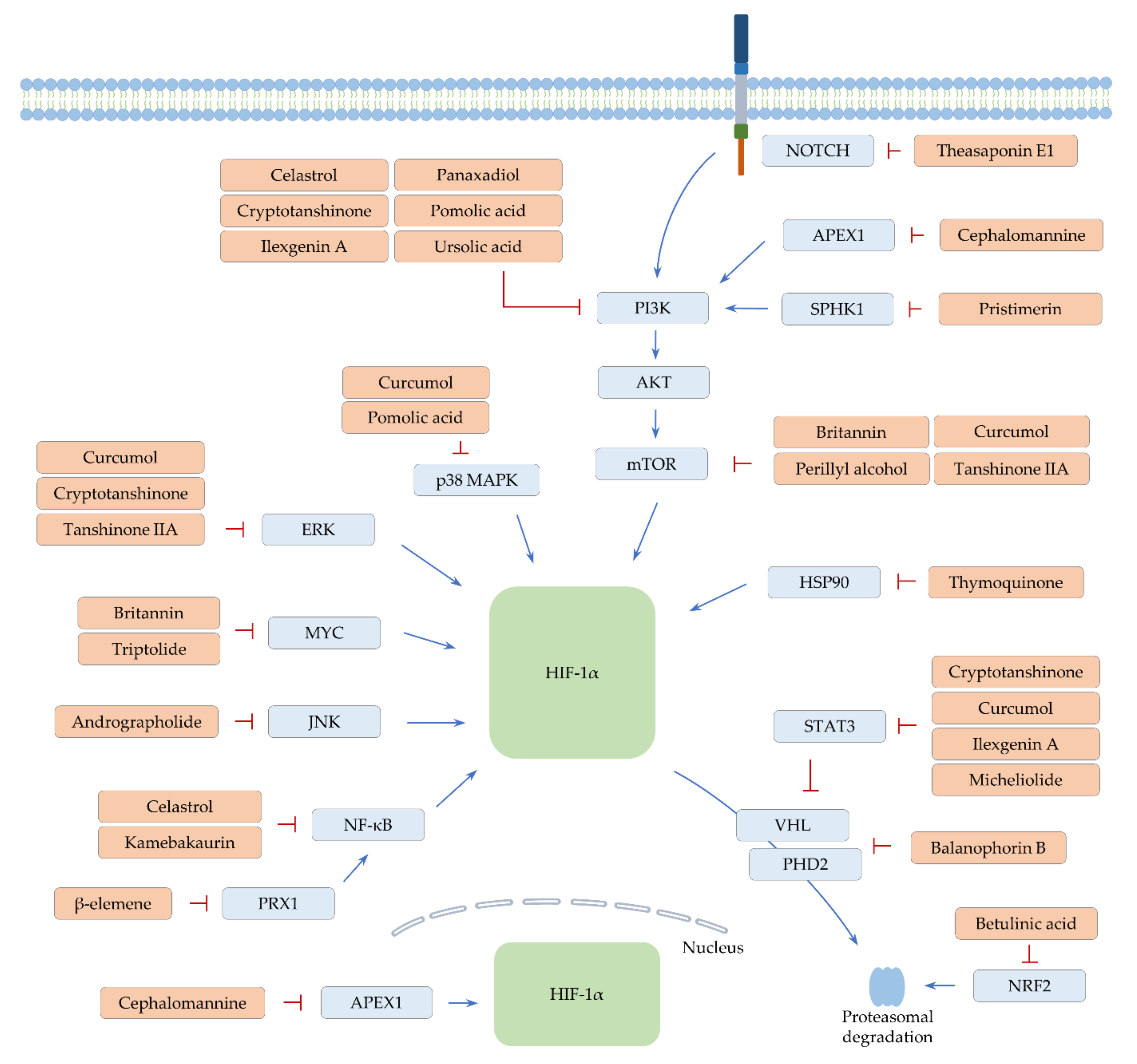
4. Terpene Phytochemicals
4.1. Monoterpenes
4.1.1. Perillyl Alcohol
4.1.2. Thymoquinone
4.2. Sesquiterpenes
4.2.1. β-Elemene and Micheliolide
4.2.2. Britannin
4.2.3. Curcumol
4.3. Diterpenes
4.3.1. Andrographolide
4.3.2. Cephalomannine
4.3.3. Cryptotanshinone and Kamebakaurin
4.3.4. Tanshinone IIA
4.3.5. Triptolide
4.4. Triterpenes
4.4.1. Balanophorin B
4.4.2. Betulinic Acid
4.4.3. Celastrol
4.4.4. Ilexgenin A and Panaxadiol
4.4.5. Pomolic Acid
4.4.6. Pristimerin
4.4.7. Theasaponin E1
4.4.8. Ursolic Acid
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tirpe, A.A.; Gulei, D.; Ciortea, S.M.; Crivii, C.; Berindan-Neagoe, I. Hypoxia: Overview on hypoxia-mediated mechanisms with a focus on the role of hif genes. Int. J. Mol. Sci. 2019, 20, 6140. [Google Scholar] [CrossRef]
- Liao, C.; Zhang, Q. Understanding the oxygen-sensing pathway and its therapeutic implications in diseases. Am. J. Pathol. 2020, 190, 1584–1595. [Google Scholar] [CrossRef]
- Joshi, S.; Singh, A.R.; Durden, D.L. Mdm2 regulates hypoxic hypoxia-inducible factor 1alpha stability in an e3 ligase, proteasome, and pten-phosphatidylinositol 3-kinase-akt-dependent manner. J. Biol. Chem 2014, 289, 22785–22797. [Google Scholar] [CrossRef]
- Pore, N.; Jiang, Z.; Shu, H.K.; Bernhard, E.; Kao, G.D.; Maity, A. Akt1 activation can augment hypoxia-inducible factor-1alpha expression by increasing protein translation through a mammalian target of rapamycin-independent pathway. Mol. Cancer Res. 2006, 4, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Eble, J.M.; Moon, E.; Yuan, H.; Weitzel, D.H.; Landon, C.D.; Nien, C.Y.; Hanna, G.; Rich, J.N.; Provenzale, J.M.; et al. Tumor cells upregulate normoxic hif-1alpha in response to doxorubicin. Cancer Res. 2013, 73, 6230–6242. [Google Scholar] [CrossRef] [PubMed]
- Moniz, S.; Bandarra, D.; Biddlestone, J.; Campbell, K.J.; Komander, D.; Bremm, A.; Rocha, S. Cezanne regulates e2f1-dependent hif2alpha expression. J. Cell Sci. 2015, 128, 3082–3093. [Google Scholar] [PubMed]
- Son, S.W.; Yun, B.D.; Song, M.G.; Lee, J.K.; Choi, S.Y.; Kuh, H.J.; Park, J.K. The hypoxia-long noncoding rna interaction in solid cancers. Int. J. Mol. Sci. 2021, 22, 7261. [Google Scholar] [CrossRef]
- Poon, E.; Harris, A.L.; Ashcroft, M. Targeting the hypoxia-inducible factor (hif) pathway in cancer. Expert Rev. Mol. Med. 2009, 11, e26. [Google Scholar] [CrossRef]
- Bertout, J.A.; Majmundar, A.J.; Gordan, J.D.; Lam, J.C.; Ditsworth, D.; Keith, B.; Brown, E.J.; Nathanson, K.L.; Simon, M.C. Hif2alpha inhibition promotes p53 pathway activity, tumor cell death, and radiation responses. Proc. Natl. Acad. Sci. USA 2009, 106, 14391–14396. [Google Scholar] [CrossRef]
- Nardinocchi, L.; Puca, R.; D’Orazi, G. Hif-1alpha antagonizes p53-mediated apoptosis by triggering hipk2 degradation. Aging 2011, 3, 33–43. [Google Scholar] [CrossRef]
- Wang, X.; Dong, J.; Jia, L.; Zhao, T.; Lang, M.; Li, Z.; Lan, C.; Li, X.; Hao, J.; Wang, H.; et al. Hif-2-dependent expression of stem cell factor promotes metastasis in hepatocellular carcinoma. Cancer Lett. 2017, 393, 113–124. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, G.; Li, X.; Zhang, Y.; Jiang, Y.; Shen, J.; Liu, J.; Wang, Q.; Zhu, J.; Feng, X.; et al. Hypoxia induces epithelial-mesenchymal transition via activation of snai1 by hypoxia-inducible factor -1alpha in hepatocellular carcinoma. BMC Cancer 2013, 13, 108. [Google Scholar] [CrossRef] [PubMed]
- Choueiri, T.K.; Kaelin, W.G., Jr. Targeting the hif2-vegf axis in renal cell carcinoma. Nat. Med. 2020, 26, 1519–1530. [Google Scholar] [CrossRef] [PubMed]
- Ravi, R.; Mookerjee, B.; Bhujwalla, Z.M.; Sutter, C.H.; Artemov, D.; Zeng, Q.; Dillehay, L.E.; Madan, A.; Semenza, G.L.; Bedi, A. Regulation of tumor angiogenesis by p53-induced degradation of hypoxia-inducible factor 1alpha. Genes Dev. 2000, 14, 34–44. [Google Scholar] [PubMed]
- Meijer, T.W.; Kaanders, J.H.; Span, P.N.; Bussink, J. Targeting hypoxia, hif-1, and tumor glucose metabolism to improve radiotherapy efficacy. Clin. Cancer Res. 2012, 18, 5585–5594. [Google Scholar] [CrossRef]
- Samanta, D.; Gilkes, D.M.; Chaturvedi, P.; Xiang, L.; Semenza, G.L. Hypoxia-inducible factors are required for chemotherapy resistance of breast cancer stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, E5429–E5438. [Google Scholar] [CrossRef] [PubMed]
- Noman, M.Z.; Desantis, G.; Janji, B.; Hasmim, M.; Karray, S.; Dessen, P.; Bronte, V.; Chouaib, S. Pd-l1 is a novel direct target of hif-1alpha, and its blockade under hypoxia enhanced mdsc-mediated t cell activation. J. Exp. Med. 2014, 211, 781–790. [Google Scholar] [CrossRef]
- Imtiyaz, H.Z.; Williams, E.P.; Hickey, M.M.; Patel, S.A.; Durham, A.C.; Yuan, L.J.; Hammond, R.; Gimotty, P.A.; Keith, B.; Simon, M.C. Hypoxia-inducible factor 2alpha regulates macrophage function in mouse models of acute and tumor inflammation. J. Clin. Investig. 2010, 120, 2699–2714. [Google Scholar] [CrossRef]
- Talks, K.L.; Turley, H.; Gatter, K.C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J.; Harris, A.L. The expression and distribution of the hypoxia-inducible factors hif-1alpha and hif-2alpha in normal human tissues, cancers, and tumor-associated macrophages. Am. J. Pathol. 2000, 157, 411–421. [Google Scholar] [CrossRef]
- Liu, Y.M.; Ying, S.P.; Huang, Y.R.; Pan, Y.; Chen, W.J.; Ni, L.Q.; Xu, J.Y.; Shen, Q.Y.; Liang, Y. Expression of hif-1alpha and hif-2alpha correlates to biological and clinical significance in papillary thyroid carcinoma. World J. Surg. Oncol. 2016, 14, 30. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Q.; Hu, J.; Chen, Y.; Liu, C.; Xu, C. Expression of hif-2alpha and vegf in cervical squamous cell carcinoma and its clinical significance. Biomed. Res. Int. 2016, 2016, 5631935. [Google Scholar]
- Moreno Roig, E.; Groot, A.J.; Yaromina, A.; Hendrickx, T.C.; Barbeau, L.M.O.; Giuranno, L.; Dams, G.; Ient, J.; Olivo Pimentel, V.; Van Gisbergen, M.W.; et al. Hif-1alpha and hif-2alpha differently regulate the radiation sensitivity of nsclc cells. Cells 2019, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Isono, T.; Chano, T.; Yoshida, T.; Kageyama, S.; Kawauchi, A.; Suzaki, M.; Yuasa, T. Hydroxyl-hif2-alpha is potential therapeutic target for renal cell carcinomas. Am. J. Cancer Res. 2016, 6, 2263–2276. [Google Scholar] [PubMed]
- Downes, N.L.; Laham-Karam, N.; Kaikkonen, M.U.; Yla-Herttuala, S. Differential but complementary hif1alpha and hif2alpha transcriptional regulation. Mol. Ther. 2018, 26, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- Loboda, A.; Jozkowicz, A.; Dulak, J. Hif-1 and hif-2 transcription factors--similar but not identical. Mol. Cells 2010, 29, 435–442. [Google Scholar] [CrossRef]
- Hoefflin, R.; Harlander, S.; Schafer, S.; Metzger, P.; Kuo, F.; Schonenberger, D.; Adlesic, M.; Peighambari, A.; Seidel, P.; Chen, C.Y.; et al. Hif-1alpha and hif-2alpha differently regulate tumour development and inflammation of clear cell renal cell carcinoma in mice. Nat. Commun. 2020, 11, 4111. [Google Scholar] [CrossRef]
- Son, S.W.; Lee, H.Y.; Moeng, S.; Kuh, H.J.; Choi, S.Y.; Park, J.K. Participation of micrornas in the treatment of cancer with phytochemicals. Molecules 2020, 25, 4701. [Google Scholar] [CrossRef] [PubMed]
- Montane, X.; Kowalczyk, O.; Reig-Vano, B.; Bajek, A.; Roszkowski, K.; Tomczyk, R.; Pawliszak, W.; Giamberini, M.; Mocek-Plociniak, A.; Tylkowski, B. Current perspectives of the applications of polyphenols and flavonoids in cancer therapy. Molecules 2020, 25, 3342. [Google Scholar] [CrossRef] [PubMed]
- Mitra, T.; Bhattacharya, R. Phytochemicals modulate cancer aggressiveness: A review depicting the anticancer efficacy of dietary polyphenols and their combinations. J. Cell Physiol. 2020, 235, 7696–7708. [Google Scholar] [CrossRef]
- Chen, Y.X.; Gao, Q.Y.; Zou, T.H.; Wang, B.M.; Liu, S.D.; Sheng, J.Q.; Ren, J.L.; Zou, X.P.; Liu, Z.J.; Song, Y.Y.; et al. Berberine versus placebo for the prevention of recurrence of colorectal adenoma: A multicentre, double-blinded, randomised controlled study. Lancet Gastroenterol. Hepatol. 2020, 5, 267–275. [Google Scholar] [CrossRef]
- Chen, Q.; Qin, R.; Fang, Y.; Li, H. Berberine sensitizes human ovarian cancer cells to cisplatin through mir-93/pten/akt signaling pathway. Cell Physiol. Biochem. 2015, 36, 956–965. [Google Scholar] [CrossRef]
- Qian, K.; Tang, C.Y.; Chen, L.Y.; Zheng, S.; Zhao, Y.; Ma, L.S.; Xu, L.; Fan, L.H.; Yu, J.D.; Tan, H.S.; et al. Berberine reverses breast cancer multidrug resistance based on fluorescence pharmacokinetics in vitro and in vivo. ACS Omega 2021, 6, 10645–10654. [Google Scholar] [CrossRef]
- Pan, Y.; Shao, D.; Zhao, Y.; Zhang, F.; Zheng, X.; Tan, Y.; He, K.; Li, J.; Chen, L. Berberine reverses hypoxia-induced chemoresistance in breast cancer through the inhibition of ampk- hif-1alpha. Int. J. Biol. Sci. 2017, 13, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Shi, Q.; Jin, Y.; Liu, Z.; Li, J.; Sun, W. Blockage of ampk-ulk1 pathway mediated autophagy promotes cell apoptosis to increase doxorubicin sensitivity in breast cancer (bc) cells: An in vitro study. BMC Cancer 2021, 21, 195. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K. Ampkalpha2 regulates hypoxia-inducible factor-1alpha stability and neutrophil survival to promote vascular repair after ischemia. Circ. Res. 2017, 120, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Kalainayakan, S.P.; Ghosh, P.; Dey, S.; Fitzgerald, K.E.; Sohoni, S.; Konduri, P.C.; Garrossian, M.; Liu, L.; Zhang, L. Cyclopamine tartrate, a modulator of hedgehog signaling and mitochondrial respiration, effectively arrests lung tumor growth and progression. Sci. Rep. 2019, 9, 1405. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wang, Z.; Li, M.Y.; Zhang, Z.; Mi, C.; Zuo, H.X.; Xing, Y.; Wu, Y.L.; Lian, L.H.; Xu, G.H.; et al. Dictamnine promotes apoptosis and inhibits epithelial-mesenchymal transition, migration, invasion and proliferation by downregulating the hif-1alpha and slug signaling pathways. Chem. Biol. Interact. 2018, 296, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.M.; Xu, P.; Chen, Q.; Feng, S.L.; Xie, Y. A multiple-targets alkaloid nuciferine overcomes paclitaxel-induced drug resistance in vitro and in vivo. Phytomedicine 2020, 79, 153342. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Wang, J.; Fan, M.; Ghauri, M.A.; Ullah, A.; Wang, B.; Dai, B.; Zhan, Y.; Zhang, D.; Zhang, Y. Sanguinarine disrupts the colocalization and interaction of hif-1alpha with tyrosine and serine phosphorylated-stat3 in breast cancer. J. Cell Mol. Med. 2020, 24, 3756–3761. [Google Scholar] [CrossRef]
- Su, Q.; Wang, J.; Wu, Q.; Ullah, A.; Ghauri, M.A.; Sarwar, A.; Chen, L.; Liu, F.; Zhang, Y. Sanguinarine combats hypoxia-induced activation of ephb4 and hif-1alpha pathways in breast cancer. Phytomedicine 2021, 84, 153503. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Fan, M.; Wang, J.; Ullah, A.; Ghauri, M.A.; Dai, B.; Zhan, Y.; Zhang, D.; Zhang, Y. Sanguinarine inhibits epithelial-mesenchymal transition via targeting hif-1alpha/tgf-beta feed-forward loop in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 939. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, L.; Zhao, F.; Yang, G.; Wang, J.J. Tetrandrine suppresses lung cancer growth and induces apoptosis, potentially via the vegf/hif-1alpha/icam-1 signaling pathway. Oncol. Lett. 2018, 15, 7433–7437. [Google Scholar] [PubMed]
- Wei, Z.; Shan, Y.; Tao, L.; Liu, Y.; Zhu, Z.; Liu, Z.; Wu, Y.; Chen, W.; Wang, A.; Lu, Y. Diallyl trisulfides, a natural histone deacetylase inhibitor, attenuate hif-1alpha synthesis, and decreases breast cancer metastasis. Mol. Carcinog. 2017, 56, 2317–2331. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Kang, T.W.; Jung, Y.D.; Zhang, C.; Lian, S. Sulforaphane inhibits nonmuscle invasive bladder cancer cells proliferation through suppression of hif-1alpha-mediated glycolysis in hypoxia. J. Agric. Food Chem. 2019, 67, 7844–7854. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Atkinson, S.J.; Akbareian, S.E.; Zhou, Z.; Munsterberg, A.; Robinson, S.D.; Bao, Y. Sulforaphane exerts anti-angiogenesis effects against hepatocellular carcinoma through inhibition of stat3/hif-1alpha/vegf signalling. Sci. Rep. 2017, 7, 12651. [Google Scholar] [CrossRef] [PubMed]
- Nys, K.; Van Laethem, A.; Michiels, C.; Rubio, N.; Piette, J.G.; Garmyn, M.; Agostinis, P. A p38(mapk)/hif-1 pathway initiated by uvb irradiation is required to induce noxa and apoptosis of human keratinocytes. J. Investig. Dermatol. 2010, 130, 2269–2276. [Google Scholar] [CrossRef] [PubMed]
- Khandrika, L.; Lieberman, R.; Koul, S.; Kumar, B.; Maroni, P.; Chandhoke, R.; Meacham, R.B.; Koul, H.K. Hypoxia-associated p38 mitogen-activated protein kinase-mediated androgen receptor activation and increased hif-1alpha levels contribute to emergence of an aggressive phenotype in prostate cancer. Oncogene 2009, 28, 1248–1260. [Google Scholar] [CrossRef]
- An, F.F.; Liu, Y.C.; Zhang, W.W.; Liang, L. Dihydroartemisinine enhances dictamnine-induced apoptosis via a caspase dependent pathway in human lung adenocarcinoma a549 cells. Asian Pac. J. Cancer Prev. 2013, 14, 5895–5900. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Y.; An, T.; Liu, P.; Zhu, J.; Yang, H.; Zhang, W.; Dong, T.; Jiang, J.; Zhang, Y.; et al. Nuciferine inhibits the progression of glioblastoma by suppressing the sox2-akt/stat3-slug signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 139. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Li, R.; Li, H.Y.; Cao, Y.B.; Bai, M.; Fan, X.J.; Wang, S.Y.; Zhang, B.; Li, S. Identification of the anti-tumor activity and mechanisms of nuciferine through a network pharmacology approach. Acta Pharmacol. Sin. 2016, 37, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yi, D.D.; Guo, J.L.; Xiang, Z.X.; Deng, L.F.; He, L. Nuciferine, extracted from nelumbo nucifera gaertn, inhibits tumor-promoting effect of nicotine involving wnt/beta-catenin signaling in non-small cell lung cancer. J. Ethnopharmacol. 2015, 165, 83–93. [Google Scholar] [CrossRef]
- Xia, X.; Wang, Q.; Ye, T.; Liu, Y.; Liu, D.; Song, S.; Zheng, C. Nrf2/abcb1-mediated efflux and parp1-mediated dampening of DNA damage contribute to doxorubicin resistance in chronic hypoxic hepg2 cells. Fundam Clin. Pharmacol. 2020, 34, 41–50. [Google Scholar] [CrossRef]
- Singh, A.; Wu, H.; Zhang, P.; Happel, C.; Ma, J.; Biswal, S. Expression of abcg2 (bcrp) is regulated by nrf2 in cancer cells that confers side population and chemoresistance phenotype. Mol. Cancer Ther. 2010, 9, 2365–2376. [Google Scholar] [CrossRef]
- He, X.; Wang, J.; Wei, W.; Shi, M.; Xin, B.; Zhang, T.; Shen, X. Hypoxia regulates abcg2 activity through the activivation of erk1/2/hif-1alpha and contributes to chemoresistance in pancreatic cancer cells. Cancer Biol. Ther. 2016, 17, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Wartenberg, M.; Ling, F.C.; Muschen, M.; Klein, F.; Acker, H.; Gassmann, M.; Petrat, K.; Putz, V.; Hescheler, J.; Sauer, H. Regulation of the multidrug resistance transporter p-glycoprotein in multicellular tumor spheroids by hypoxia-inducible factor (hif-1) and reactive oxygen species. FASEB J. 2003, 17, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.E.; Kim, H.S.; Lee, C.S.; Shin, Y.J.; Kim, Y.N.; Kang, G.H.; Kim, T.Y.; Juhnn, Y.S.; Kim, S.J.; Park, J.W.; et al. Stat3 inhibits the degradation of hif-1alpha by pvhl-mediated ubiquitination. Exp. Mol. Med. 2008, 40, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Pawlus, M.R.; Wang, L.; Hu, C.J. Stat3 and hif1alpha cooperatively activate hif1 target genes in mda-mb-231 and rcc4 cells. Oncogene 2014, 33, 1670–1679. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.Y.; Liu, Y.G.; Song, W.; Li, Y.Y.; Hou, D.S.; Luo, H.M.; Liu, P. Anticancer activity of tetrandrine by inducing pro-death apoptosis and autophagy in human gastric cancer cells. J. Pharm. Pharmacol. 2018, 70, 1048–1058. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, T.; Yu, M.; Li, K.; Li, W. The plant alkaloid tetrandrine inhibits metastasis via autophagy-dependent wnt/beta-catenin and metastatic tumor antigen 1 signaling in human liver cancer cells. J. Exp. Clin. Cancer Res. 2018, 37, 7. [Google Scholar] [CrossRef]
- Ye, L.Y.; Hu, S.; Xu, H.E.; Xu, R.R.; Kong, H.; Zeng, X.N.; Xie, W.P.; Wang, H. The effect of tetrandrine combined with cisplatin on proliferation and apoptosis of a549/ddp cells and a549 cells. Cancer Cell Int. 2017, 17, 40. [Google Scholar] [CrossRef]
- Zhang, H.; Wong, C.C.; Wei, H.; Gilkes, D.M.; Korangath, P.; Chaturvedi, P.; Schito, L.; Chen, J.; Krishnamachary, B.; Winnard, P.T., Jr.; et al. Hif-1-dependent expression of angiopoietin-like 4 and l1cam mediates vascular metastasis of hypoxic breast cancer cells to the lungs. Oncogene 2012, 31, 1757–1770. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Wang, Y.; Qiu, L.; Li, S.; Zhu, J.; Liang, Z.; Wan, Y.; Di, W. Hypoxia inducible factor 1alpha-mediated lox expression correlates with migration and invasion in epithelial ovarian cancer. Int. J. Oncol. 2013, 42, 1578–1588. [Google Scholar] [CrossRef]
- Wong, C.C.; Gilkes, D.M.; Zhang, H.; Chen, J.; Wei, H.; Chaturvedi, P.; Fraley, S.I.; Wong, C.M.; Khoo, U.S.; Ng, I.O.; et al. Hypoxia-inducible factor 1 is a master regulator of breast cancer metastatic niche formation. Proc. Natl. Acad. Sci. USA 2011, 108, 16369–16374. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Singh, S.V. Diallyl trisulfide, a constituent of processed garlic, inactivates akt to trigger mitochondrial translocation of bad and caspase-mediated apoptosis in human prostate cancer cells. Carcinogenesis 2006, 27, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Chen, B.; Liu, X.; Liu, P.; Zheng, G.; Ye, F.; Tang, H.; Xie, X. Diallyl disulfide suppresses src/ras/erk signaling-mediated proliferation and metastasis in human breast cancer by up-regulating mir-34a. PLoS ONE 2014, 9, e112720. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, W.; Zhang, Y.; Sun, W.; Yung, M.M.H.; Sun, J.; Li, J.; Chen, C.W.; Li, Z.; Meng, Y.; et al. Erk regulates hif1alpha-mediated platinum resistance by directly targeting phd2 in ovarian cancer. Clin. Cancer Res. 2019, 25, 5947–5960. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Xiao, D.; Lew, K.L.; Dhir, R.; Singh, S.V. Sulforaphane induces caspase-mediated apoptosis in cultured pc-3 human prostate cancer cells and retards growth of pc-3 xenografts in vivo. Carcinogenesis 2004, 25, 83–90. [Google Scholar] [CrossRef]
- Castro, N.P.; Rangel, M.C.; Merchant, A.S.; MacKinnon, G.; Cuttitta, F.; Salomon, D.S.; Kim, Y.S. Sulforaphane suppresses the growth of triple-negative breast cancer stem-like cells in vitro and in vivo. Cancer Prev Res. 2019, 12, 147–158. [Google Scholar] [CrossRef]
- Liu, C.M.; Peng, C.Y.; Liao, Y.W.; Lu, M.Y.; Tsai, M.L.; Yeh, J.C.; Yu, C.H.; Yu, C.C. Sulforaphane targets cancer stemness and tumor initiating properties in oral squamous cell carcinomas via mir-200c induction. J. Formos. Med. Assoc. 2017, 116, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Wu, M.J.; Yang, J.Y.; Camarillo, I.G.; Chang, C.J. Leptin-stat3-g9a signaling promotes obesity-mediated breast cancer progression. Cancer Res. 2015, 75, 2375–2386. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.; Choi, Y.C.; Jeong, Y.; Lee, G.; Yoon, S.; Jeong, Y.; Yoon, J.; Baek, K. Mir-200c downregulates hif-1alpha and inhibits migration of lung cancer cells. Cell Mol. Biol. Lett 2019, 24, 28. [Google Scholar] [CrossRef]
- Lin, C.C.; Chuang, Y.J.; Yu, C.C.; Yang, J.S.; Lu, C.C.; Chiang, J.H.; Lin, J.P.; Tang, N.Y.; Huang, A.C.; Chung, J.G. Apigenin induces apoptosis through mitochondrial dysfunction in u-2 os human osteosarcoma cells and inhibits osteosarcoma xenograft tumor growth in vivo. J. Agric. Food Chem. 2012, 60, 11395–11402. [Google Scholar] [CrossRef]
- Zhu, Y.; Mao, Y.; Chen, H.; Lin, Y.; Hu, Z.; Wu, J.; Xu, X.; Xu, X.; Qin, J.; Xie, L. Apigenin promotes apoptosis, inhibits invasion and induces cell cycle arrest of t24 human bladder cancer cells. Cancer Cell Int. 2013, 13, 54. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Van Wie, P.G.; Fai, L.Y.; Kim, D.; Wang, L.; Poyil, P.; Luo, J.; Zhang, Z. Downregulation of nedd9 by apigenin suppresses migration, invasion, and metastasis of colorectal cancer cells. Toxicol. Appl. Pharmacol. 2016, 311, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-Ayala, K.; Ortiz Rivera, I.; Alvarez, C.; Husain, K.; DeLoach, D.; Krystal, G.; Hibbs, M.L.; Jiang, K.; Ghansah, T. Apigenin increases ship-1 expression, promotes tumoricidal macrophages and anti-tumor immune responses in murine pancreatic cancer. Cancers 2020, 12, 3631. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Zhou, Q.; Liu, L.Z.; Xia, C.; Hu, X.; Shi, X.; Jiang, B.H. Apigenin inhibits tumor angiogenesis through decreasing hif-1alpha and vegf expression. Carcinogenesis 2007, 28, 858–864. [Google Scholar] [CrossRef]
- Ketkaew, Y.; Osathanon, T.; Pavasant, P.; Sooampon, S. Apigenin inhibited hypoxia induced stem cell marker expression in a head and neck squamous cell carcinoma cell line. Arch. Oral Biol. 2017, 74, 69–74. [Google Scholar] [CrossRef]
- Yoshida, T.; Hashimura, M.; Mastumoto, T.; Tazo, Y.; Inoue, H.; Kuwata, T.; Saegusa, M. Transcriptional upregulation of hif-1alpha by nf-kappab/p65 and its associations with beta-catenin/p300 complexes in endometrial carcinoma cells. Lab. Investig. 2013, 93, 1184–1193. [Google Scholar] [CrossRef]
- Zhou, J.; Callapina, M.; Goodall, G.J.; Brune, B. Functional integrity of nuclear factor kappab, phosphatidylinositol 3’-kinase, and mitogen-activated protein kinase signaling allows tumor necrosis factor alpha-evoked bcl-2 expression to provoke internal ribosome entry site-dependent translation of hypoxia-inducible factor 1alpha. Cancer Res. 2004, 64, 9041–9048. [Google Scholar] [PubMed]
- Tong, X.; Pelling, J.C. Targeting the pi3k/akt/mtor axis by apigenin for cancer prevention. Anticancer Agents Med. Chem. 2013, 13, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhao, D.; Zhou, H.G.; Wang, X.H.; Zhong, W.L.; Chen, S.; Gu, W.G.; Wang, W.; Zhang, C.H.; Liu, Y.R.; et al. Apigenin inhibits nf-kappab and snail signaling, emt and metastasis in human hepatocellular carcinoma. Oncotarget 2016, 7, 41421–41431. [Google Scholar] [CrossRef]
- Jin, J.; Qiu, S.; Wang, P.; Liang, X.; Huang, F.; Wu, H.; Zhang, B.; Zhang, W.; Tian, X.; Xu, R.; et al. Cardamonin inhibits breast cancer growth by repressing hif-1alpha-dependent metabolic reprogramming. J. Exp. Clin. Cancer Res. 2019, 38, 377. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.Q.; Xu, M.; Zhong, W.T.; Cui, Z.Y.; Liu, F.M.; Zhou, K.Y.; Li, X.Y. Egcg decreases the expression of hif-1alpha and vegf and cell growth in mcf-7 breast cancer cells. J. BUON 2014, 19, 435–439. [Google Scholar] [PubMed]
- Guo, Q.; Lu, L.; Liao, Y.; Wang, X.; Zhang, Y.; Liu, Y.; Huang, S.; Sun, H.; Li, Z.; Zhao, L. Influence of c-src on hypoxic resistance to paclitaxel in human ovarian cancer cells and reversal of fv-429. Cell Death Dis. 2018, 8, e3178. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Ma, R.; Huang, S.; Liao, Y.; Ding, Y.; Li, Z.; Guo, Q.; Tan, R.; Zhang, L.; Zhao, L. Oroxylin a increases the sensitivity of temozolomide on glioma cells by hypoxia-inducible factor 1alpha/hedgehog pathway under hypoxia. J. Cell Physiol. 2019, 234, 17392–17404. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Li, W.; Xu, Y.; Zhuo, Y.; Li, M.; He, Y.; Wang, X.; Guo, Q.; Zhao, L.; et al. Oroxylin a reverses hypoxia-induced cisplatin resistance through inhibiting hif-1alpha mediated xpc transcription. Oncogene 2020, 39, 6893–6905. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Zhao, J.K.; Ren, S.; Sun, W.W.; Zhang, W.J.; Zhang, J.N. Wogonin affects proliferation and the energy metabolism of sgc-7901 and a549 cells. Exp. Ther. Med. 2019, 17, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Zhao, R.Q.; Zhang, Y.K.; Gupta, P.; Fu, L.X.; Tang, A.Z.; Liu, B.M.; Chen, Z.S.; Yang, D.H.; Liang, G. Effect of y6, an epigallocatechin gallate derivative, on reversing doxorubicin drug resistance in human hepatocellular carcinoma cells. Oncotarget 2017, 8, 29760–29770. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.H.; Zhu, H.Q.; Chen, Y.Y.; Chen, R.L.; Fu, L.X.; Li, L.; Zhou, H.; Zhou, J.L.; Liang, G. The epigallocatechin gallate derivative y6 inhibits human hepatocellular carcinoma by inhibiting angiogenesis in mapk/erk1/2 and pi3k/akt/ hif-1alpha/vegf dependent pathways. J. Ethnopharmacol. 2020, 259, 112852. [Google Scholar] [CrossRef]
- Kim, D.H.; Sung, B.; Kim, J.A.; Kang, Y.J.; Hwang, S.Y.; Hwang, N.L.; Suh, H.; Choi, Y.H.; Im, E.; Chung, H.Y.; et al. Hs-1793, a resveratrol analogue, downregulates the expression of hypoxia-induced hif-1 and vegf and inhibits tumor growth of human breast cancer cells in a nude mouse xenograft model. Int. J. Oncol. 2017, 51, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Liu, Q.; Liu, X.; Ji, M.; Xie, P.; Chen, X. Lxy6090—A novel manassantin a derivative-limits breast cancer growth through hypoxia-inducible factor-1 inhibition. Onco. Targets Ther. 2016, 9, 3829–3840. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, M.C.; Lee, C.F.; Huang, W.H.; Chou, T.C. Magnolol suppresses hypoxia-induced angiogenesis via inhibition of hif-1alpha/vegf signaling pathway in human bladder cancer cells. Biochem. Pharmacol. 2013, 85, 1278–1287. [Google Scholar] [CrossRef]
- Kwak, S.H.; Stephenson, T.N.; Lee, H.E.; Ge, Y.; Lee, H.; Min, S.M.; Kim, J.H.; Kwon, D.Y.; Lee, Y.M.; Hong, J. Evaluation of manassantin a tetrahydrofuran core region analogues and cooperative therapeutic effects with egfr inhibition. J. Med. Chem. 2020, 63, 6821–6833. [Google Scholar] [CrossRef]
- Aljabali, A.A.A.; Bakshi, H.A.; Hakkim, F.L.; Haggag, Y.A.; Al-Batanyeh, K.M.; Al Zoubi, M.S.; Al-Trad, B.; Nasef, M.M.; Satija, S.; Mehta, M.; et al. Albumin nano-encapsulation of piceatannol enhances its anticancer potential in colon cancer via downregulation of nuclear p65 and hif-1alpha. Cancers 2020, 12, 113. [Google Scholar] [CrossRef]
- Butt, N.A.; Kumar, A.; Dhar, S.; Rimando, A.M.; Akhtar, I.; Hancock, J.C.; Lage, J.M.; Pound, C.R.; Lewin, J.R.; Gomez, C.R.; et al. Targeting mta1/hif-1alpha signaling by pterostilbene in combination with histone deacetylase inhibitor attenuates prostate cancer progression. Cancer Med. 2017, 6, 2673–2685. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jia, R.; Lv, T.; Wang, M.; He, S.; Zhang, X. Resveratrol suppresses tumor progression via inhibiting stat3/hif-1alpha/vegf pathway in an orthotopic rat model of non-small-cell lung cancer (nsclc). Onco. Targets Ther. 2020, 13, 7057–7063. [Google Scholar] [CrossRef]
- Xu, Q.H.; Xiao, Y.; Li, X.Q.; Fan, L.; Zhou, C.C.; Cheng, L.; Jiang, Z.D.; Wang, G.H. Resveratrol counteracts hypoxia-induced gastric cancer invasion and emt through hedgehog pathway suppression. Anticancer Agents Med. Chem. 2020, 20, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Qin, T.; Sun, L.; Qian, W.; Li, J.; Duan, W.; Lei, J.; Wang, Z.; Ma, J.; Li, X.; et al. Resveratrol ameliorates the malignant progression of pancreatic cancer by inhibiting hypoxia-induced pancreatic stellate cell activation. Cell Transplant. 2020, 29, 963689720929987. [Google Scholar] [CrossRef]
- Kim, A.; Ma, J.Y. Rhaponticin decreases the metastatic and angiogenic abilities of cancer cells via suppression of the hif1alpha pathway. Int J. Oncol. 2018, 53, 1160–1170. [Google Scholar] [PubMed]
- Gong, J.; Zhou, S.; Yang, S. Vanillic acid suppresses hif-1alpha expression via inhibition of mtor/p70s6k/4e-bp1 and raf/mek/erk pathways in human colon cancer hct116 cells. Int. J. Mol. Sci. 2019, 20, 465. [Google Scholar] [CrossRef] [PubMed]
- Park, J.J.; Hwang, S.J.; Park, J.H.; Lee, H.J. Chlorogenic acid inhibits hypoxia-induced angiogenesis via down-regulation of the hif-1alpha/akt pathway. Cell Oncol 2015, 38, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, F.; Taverna, S.; Alessandro, R.; Fontana, S. Swath-ms based quantitative proteomics analysis reveals that curcumin alters the metabolic enzyme profile of cml cells by affecting the activity of mir-22/ipo7/hif-1alpha axis. J. Exp. Clin. Cancer Res. 2018, 37, 170. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Yoon, S.H.; Jang, H.; Jeong, J.H.; Lee, Y.M. Decursin promotes hif-1alpha proteasomal degradation and immune responses in hypoxic tumour microenvironment. Phytomedicine 2020, 78, 153318. [Google Scholar] [CrossRef]
- Velatooru, L.R.; Vakamullu, S.; Penugurti, V. Alpinoid c analog inhibits angiogenesis and induces apoptosis in colo205cell line. Chem. Biol. Interact. 2019, 308, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, W.; Guo, L.; Bao, W.; Jin, N.; Liu, R.; Liu, P.; Wang, Y.; Guo, Q.; Chen, B. Gambogic acid suppresses hypoxia-induced hypoxia-inducible factor-1alpha/vascular endothelial growth factor expression via inhibiting phosphatidylinositol 3-kinase/akt/mammalian target protein of rapamycin pathway in multiple myeloma cells. Cancer Sci. 2014, 105, 1063–1070. [Google Scholar] [CrossRef]
- Ranjbarnejad, T.; Saidijam, M.; Tafakh, M.S.; Pourjafar, M.; Talebzadeh, F.; Najafi, R. Garcinol exhibits anti-proliferative activities by targeting microsomal prostaglandin e synthase-1 in human colon cancer cells. Hum. Exp. Toxicol. 2017, 36, 692–700. [Google Scholar] [CrossRef]
- Mi, C.; Ma, J.; Wang, K.S.; Zuo, H.X.; Wang, Z.; Li, M.Y.; Piao, L.X.; Xu, G.H.; Li, X.; Quan, Z.S.; et al. Imperatorin suppresses proliferation and angiogenesis of human colon cancer cell by targeting hif-1alpha via the mtor/p70s6k/4e-bp1 and mapk pathways. J. Ethnopharmacol. 2017, 203, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Liu, H.J.; Li, M.; Zhai, D.H.; Tang, Y.H.; Yang, L.; Qiao, K.L.; Yang, J.H.; Zhong, W.L.; Zhang, Q.; et al. Salidroside improves the hypoxic tumor microenvironment and reverses the drug resistance of platinum drugs via hif-1alpha signaling pathway. EBioMedicine 2018, 38, 25–36. [Google Scholar] [CrossRef]
- Li, M.Y.; Mi, C.; Wang, K.S.; Wang, Z.; Zuo, H.X.; Piao, L.X.; Xu, G.H.; Li, X.; Ma, J.; Jin, X. Shikonin suppresses proliferation and induces cell cycle arrest through the inhibition of hypoxia-inducible factor-1alpha signaling. Chem. Biol. Interact. 2017, 274, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Seyfi, D.; Behzad, S.B.; Nabiuni, M.; Parivar, K.; Tahmaseb, M.; Amini, E. Verbascoside attenuates rac-1 and hif-1alpha signaling cascade in colorectal cancer cells. Anticancer Agents Med. Chem. 2018, 18, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Li, C.; Qi, Q.; Shen, J.; Chang, K. Cardamonin induces g2/m arrest and apoptosis via activation of the jnk-foxo3a pathway in breast cancer cells. Cell Biol. Int. 2019, 44, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Zan, L.; Chen, Q.; Zhang, L.; Li, X. Epigallocatechin gallate (egcg) suppresses growth and tumorigenicity in breast cancer cells by downregulation of mir-25. Bioengineered 2019, 10, 374–382. [Google Scholar] [CrossRef]
- Shrivastava, S.; Jeengar, M.K.; Thummuri, D.; Koval, A.; Katanaev, V.L.; Marepally, S.; Naidu, V.G.M. Cardamonin, a chalcone, inhibits human triple negative breast cancer cell invasiveness by downregulation of wnt/beta-catenin signaling cascades and reversal of epithelial-mesenchymal transition. Biofactors 2017, 43, 152–169. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Shi, W.; Guo, H.; Long, W.; Wang, Y.; Qi, J.; Liu, J.; Xu, Y. The inhibitory effect of (-)-epigallocatechin-3-gallate on breast cancer progression via reducing scube2 methylation and dnmt activity. Molecules 2019, 24, 2899. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.H.; Horng, C.T.; Lee, C.F.; Chiang, N.N.; Tsai, F.J.; Lu, C.C.; Chiang, J.H.; Hsu, Y.M.; Yang, J.S.; Chen, F.A. Epigallocatechin gallate sensitizes cisplatin-resistant oral cancer car cell apoptosis and autophagy through stimulating akt/stat3 pathway and suppressing multidrug resistance 1 signaling. Environ. Toxicol 2017, 32, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Liu, Z.; Li, J.; Zhang, Q.; Zhong, P.; Teng, T.; Chen, M.; Xie, Z.; Ji, A.; Li, Y. Epigallocatechin-3-gallate inhibits the growth and increases the apoptosis of human thyroid carcinoma cells through suppression of egfr/ras/raf/mek/erk signaling pathway. Cancer Cell Int. 2019, 19, 43. [Google Scholar] [CrossRef] [PubMed]
- Holczer, M.; Besze, B.; Zambo, V.; Csala, M.; Banhegyi, G.; Kapuy, O. Epigallocatechin-3-gallate (egcg) promotes autophagy-dependent survival via influencing the balance of mtor-ampk pathways upon endoplasmic reticulum stress. Oxid. Med. Cell Longev. 2018, 2018, 6721530. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Zhang, Y.; Yang, L.; Liu, Y.; Huang, S.; Lu, L.; Kong, L.; Li, Z.; Guo, Q.; et al. Wogonin reversed resistant human myelogenous leukemia cells via inhibiting nrf2 signaling by stat3/nf-kappab inactivation. Sci. Rep. 2017, 7, 39950. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yao, J.; Wu, X.P.; Zhao, L.; Zhou, Y.X.; Zhang, Y.; You, Q.D.; Guo, Q.L.; Lu, N. Wogonin suppresses human alveolar adenocarcinoma cell a549 migration in inflammatory microenvironment by modulating the il-6/stat3 signaling pathway. Mol. Carcinog 2015, 54 (Suppl. S1), E81–E93. [Google Scholar] [CrossRef] [PubMed]
- Bhuria, V.; Xing, J.; Scholta, T.; Bui, K.C.; Nguyen, M.L.T.; Malek, N.P.; Bozko, P.; Plentz, R.R. Hypoxia induced sonic hedgehog signaling regulates cancer stemness, epithelial-to-mesenchymal transition and invasion in cholangiocarcinoma. Exp. Cell Res. 2019, 385, 111671. [Google Scholar] [CrossRef] [PubMed]
- Po, A.; Citarella, A.; Catanzaro, G.; Besharat, Z.M.; Trocchianesi, S.; Gianno, F.; Sabato, C.; Moretti, M.; De Smaele, E.; Vacca, A.; et al. Hedgehog-gli signalling promotes chemoresistance through the regulation of abc transporters in colorectal cancer cells. Sci. Rep. 2020, 10, 13988. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.L.; Rodriguez-Cruz, V.; Walker, N.D.; Greco, S.J.; Rameshwar, P. Temozolomide resistance and tumor recurrence: Halting the hedgehog. Cancer Cell Microenviron. 2015, 2, e747. [Google Scholar]
- Kudo, K.; Gavin, E.; Das, S.; Amable, L.; Shevde, L.A.; Reed, E. Inhibition of gli1 results in altered c-jun activation, inhibition of cisplatin-induced upregulation of ercc1, xpd and xrcc1, and inhibition of platinum-DNA adduct repair. Oncogene 2012, 31, 4718–4724. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, M.J.; Sung, B.; Suh, H.; Jung, J.H.; Chung, H.Y.; Kim, N.D. Resveratrol analogue, hs-1793, induces apoptotic cell death and cell cycle arrest through downregulation of akt in human colon cancer cells. Oncol. Rep. 2017, 37, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, B.Y.; Kim, K.S.; Nam, J.B.; Hong, Y.S.; Lee, J.J. Suppression of rela/p65 transactivation activity by a lignoid manassantin isolated from saururus chinensis. Biochem. Pharmacol. 2003, 66, 1925–1933. [Google Scholar] [CrossRef]
- Chang, J.S.; Lee, S.W.; Kim, M.S.; Yun, B.R.; Park, M.H.; Lee, S.G.; Park, S.J.; Lee, W.S.; Rho, M.C. Manassantin a and b from saururus chinensis inhibit interleukin-6-induced signal transducer and activator of transcription 3 activation in hep3b cells. J. Pharmacol. Sci. 2011, 115, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D.; Oh, J.; Park, H.J.; Bae, K.; Lee, S.K. Magnolol inhibits angiogenesis by regulating ros-mediated apoptosis and the pi3k/akt/mtor signaling pathway in mes/eb-derived endothelial-like cells. Int. J. Oncol. 2013, 43, 600–610. [Google Scholar] [CrossRef]
- Seyed, M.A.; Jantan, I.; Bukhari, S.N.; Vijayaraghavan, K. A comprehensive review on the chemotherapeutic potential of piceatannol for cancer treatment, with mechanistic insights. J. Agric. Food Chem. 2016, 64, 725–737. [Google Scholar] [CrossRef]
- Farrand, L.; Byun, S.; Kim, J.Y.; Im-Aram, A.; Lee, J.; Lim, S.; Lee, K.W.; Suh, J.Y.; Lee, H.J.; Tsang, B.K. Piceatannol enhances cisplatin sensitivity in ovarian cancer via modulation of p53, x-linked inhibitor of apoptosis protein (xiap), and mitochondrial fission. J. Biol. Chem. 2013, 288, 23740–23750. [Google Scholar] [CrossRef]
- Siedlecka-Kroplewska, K.; Slebioda, T.; Kmiec, Z. Induction of autophagy, apoptosis and aquisition of resistance in response to piceatannol toxicity in molt-4 human leukemia cells. Toxicol. Vitro 2019, 59, 12–25. [Google Scholar] [CrossRef]
- Reddy, S.D.; Pakala, S.B.; Molli, P.R.; Sahni, N.; Karanam, N.K.; Mudvari, P.; Kumar, R. Metastasis-associated protein 1/histone deacetylase 4-nucleosome remodeling and deacetylase complex regulates phosphatase and tensin homolog gene expression and function. J. Biol. Chem. 2012, 287, 27843–27850. [Google Scholar] [CrossRef]
- Yoo, Y.G.; Kong, G.; Lee, M.O. Metastasis-associated protein 1 enhances stability of hypoxia-inducible factor-1alpha protein by recruiting histone deacetylase 1. EMBO J. 2006, 25, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.Y.; Liu, Z.S.; Yan, H.J.; Yuan, Y.F.; Levenson, A.S.; Li, K. Pterostilbene inhibits mta1/hdac1 complex leading to pten acetylation in hepatocellular carcinoma. Biomed. Pharmacother. 2018, 101, 852–859. [Google Scholar] [CrossRef]
- Czop, M.; Bogucka-Kocka, A.; Kubrak, T.; Knap-Czop, K.; Makuch-Kocka, A.; Galkowski, D.; Wawer, J.; Kocki, T.; Kocki, J. Imaging flow cytometric analysis of stilbene-dependent apoptosis in drug resistant human leukemic cell lines. Molecules 2019, 24, 1896. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Tian, W.; Wang, X.; Ma, X. Inhibitory effect of desoxyrhaponticin and rhaponticin, two natural stilbene glycosides from the tibetan nutritional food rheum tanguticum maxim. Ex balf., on fatty acid synthase and human breast cancer cells. Food Funct. 2014, 5, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Mickymaray, S.; Alfaiz, F.A.; Paramasivam, A.; Veeraraghavan, V.P.; Periadurai, N.D.; Surapaneni, K.M.; Niu, G. Rhaponticin suppresses osteosarcoma through the inhibition of pi3k-akt-mtor pathway. Saudi J. Biol. Sci. 2021, 28, 3641–3649. [Google Scholar] [CrossRef]
- Zhang, F.; Yin, G.; Han, X.; Jiang, X.; Bao, Z. Chlorogenic acid inhibits osteosarcoma carcinogenesis via suppressing the stat3/snail pathway. J. Cell Biochem. 2019, 120, 10342–10350. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Kim, S.; Lee, S.; Jiang, H.L.; Kim, S.B.; Hong, S.H.; Cho, M.H. Knockdown of importin 7 inhibits lung tumorigenesis in k-ras(la1) lung cancer mice. Anticancer Res. 2017, 37, 2381–2386. [Google Scholar] [CrossRef] [PubMed]
- Li, S.R.; Gyselman, V.G.; Dorudi, S.; Bustin, S.A. Elevated levels of ranbp7 mrna in colorectal carcinoma are associated with increased proliferation and are similar to the transcription pattern of the proto-oncogene c-myc. Biochem. Biophys. Res. Commun. 2000, 271, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Golomb, L.; Bublik, D.R.; Wilder, S.; Nevo, R.; Kiss, V.; Grabusic, K.; Volarevic, S.; Oren, M. Importin 7 and exportin 1 link c-myc and p53 to regulation of ribosomal biogenesis. Mol. Cell 2012, 45, 222–232. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, A.; Tan, C.; Wu, Y.; Lee, H.T.; Li, W.; Xie, K.; Huang, S. Forkhead box m1 is essential for nuclear localization of glioma-associated oncogene homolog 1 in glioblastoma multiforme cells by promoting importin-7 expression. J. Biol. Chem. 2015, 290, 18662–18670. [Google Scholar] [CrossRef] [PubMed]
- Chachami, G.; Paraskeva, E.; Mingot, J.M.; Braliou, G.G.; Gorlich, D.; Simos, G. Transport of hypoxia-inducible factor hif-1alpha into the nucleus involves importins 4 and 7. Biochem. Biophys. Res. Commun. 2009, 390, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.H.; Kirschenbaum, A.; Lu, M.; Yao, S.; Dosoretz, A.; Holland, J.F.; Levine, A.C. Prostaglandin e2 induces hypoxia-inducible factor-1alpha stabilization and nuclear localization in a human prostate cancer cell line. J. Biol. Chem. 2002, 277, 50081–50086. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Barbero, A.; Dorado, F.; Velasco, S.; Pandiella, A.; Banas, B.; Lopez-Novoa, J.M. Tgf-beta1 induces cox-2 expression and pge2 synthesis through mapk and pi3k pathways in human mesangial cells. Kidney Int. 2006, 70, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Grabarska, A.; Skalicka-Wozniak, K.; Kielbus, M.; Dmoszynska-Graniczka, M.; Miziak, P.; Szumilo, J.; Nowosadzka, E.; Kowalczuk, K.; Khalifa, S.; Smok-Kalwat, J.; et al. Imperatorin as a promising chemotherapeutic agent against human larynx cancer and rhabdomyosarcoma cells. Molecules 2020, 25, 2046. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Yang, Y.; Gao, B. Imperatorin targets mcl-1 to sensitize cd133+ lung cancer cells to gammadelta-t cell-mediated cytotoxicity. Cell Physiol. Biochem. 2018, 49, 235–244. [Google Scholar] [CrossRef]
- Xu, W.W.; Huang, Z.H.; Liao, L.; Zhang, Q.H.; Li, J.Q.; Zheng, C.C.; He, Y.; Luo, T.T.; Wang, Y.; Hu, H.F.; et al. Direct targeting of creb1 with imperatorin inhibits tgfbeta2-erk signaling to suppress esophageal cancer metastasis. Adv. Sci. 2020, 7, 2000925. [Google Scholar] [CrossRef] [PubMed]
- Gwon, S.Y.; Ahn, J.Y.; Jung, C.H.; Moon, B.K.; Ha, T.Y. Shikonin suppresses erk 1/2 phosphorylation during the early stages of adipocyte differentiation in 3t3-l1 cells. BMC Complement. Altern. Med. 2013, 13, 207. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Liao, C.H.; Chen, W.S.; Pai, J.T.; Weng, M.S. Shikonin inhibited migration and invasion of human lung cancer cells via suppression of c-met-mediated epithelial-to-mesenchymal transition. J. Cell Biochem. 2017, 118, 4639–4651. [Google Scholar] [CrossRef]
- Memmott, R.M.; Dennis, P.A. Akt-dependent and -independent mechanisms of mtor regulation in cancer. Cell Signal. 2009, 21, 656–664. [Google Scholar] [CrossRef]
- Huang, X.; Xu, X.; Wang, X.; Tang, T.; Li, E.; Zhang, X.; Xu, J.; Shen, H.; Guo, C.; Xu, T.; et al. The akt-independent met-v-atpase-mtor axis suppresses liver cancer vaccination. Signal. Transduct. Target. Ther. 2020, 5, 122. [Google Scholar] [CrossRef]
- Zhang, Y.; Yuan, Y.; Wu, H.; Xie, Z.; Wu, Y.; Song, X.; Wang, J.; Shu, W.; Xu, J.; Liu, B.; et al. Effect of verbascoside on apoptosis and metastasis in human oral squamous cell carcinoma. Int. J. Cancer 2018, 143, 980–991. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.Q.; Wang, Z.T.; Zou, M.M.; Lin, J.H.; Li, Y.H.; Zhang, L.; Xu, R.X. Verbascoside inhibits glioblastoma cell proliferation, migration and invasion while promoting apoptosis through upregulation of protein tyrosine phosphatase shp-1 and inhibition of stat3 phosphorylation. Cell Physiol. Biochem. 2018, 47, 1871–1882. [Google Scholar] [CrossRef]
- Wu, C.H.; Chen, C.H.; Hsieh, P.F.; Lee, Y.H.; Kuo, W.W.; Wu, R.C.; Hung, C.H.; Yang, Y.L.; Lin, V.C. Verbascoside inhibits the epithelial-mesenchymal transition of prostate cancer cells through high-mobility group box 1/receptor for advanced glycation end-products/tgf-beta pathway. Environ. Toxicol. 2021, 36, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.S.; Kang, M.J.; Lee, J.H.; Ryu, B.K.; Lee, M.G.; Her, N.G.; Ha, T.K.; Han, J.; Kim, Y.K.; Chi, S.G. Opposite functions of hif-alpha isoforms in vegf induction by tgf-beta1 under non-hypoxic conditions. Oncogene 2011, 30, 1213–1228. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, M.; Khan, M.A.; Sharma, K.; Sharma, G.; Dutta, R.K.; Majumdar, S. Gambogic acid induced oxidative stress dependent caspase activation regulates both apoptosis and autophagy by targeting various key molecules (nf-kappab, beclin-1, p62 and nbr1) in human bladder cancer cells. Biochim. Biophys. Acta 2014, 1840, 3374–3384. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Zhang, Q.; Wang, S.; Gao, Y.; Chen, X.; Zhao, Y.; Qian, X. Gambogic acid impairs tumor angiogenesis by targeting yap/stat3 signaling axis. Phytother. Res. 2019, 33, 1579–1591. [Google Scholar] [CrossRef]
- Yu, J.; Wang, W.; Yao, W.; Yang, Z.; Gao, P.; Liu, M.; Wang, H.; Chen, S.; Wang, D.; Wang, W.; et al. Gambogic acid affects escc progression through regulation of pi3k/akt/mtor signal pathway. J. Cancer 2020, 11, 5568–5577. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Zhu, Y.; Zhang, J.; Zhu, Z.; Ning, Z.; Chen, H.; Liu, L.; Chen, Z.; Meng, Z. Mir-122 targets x-linked inhibitor of apoptosis protein to sensitize oxaliplatin-resistant colorectal cancer cells to oxaliplatin-mediated cytotoxicity. Cell Physiol. Biochem. 2018, 51, 2148–2159. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Ma, J.; Deng, G.; Qu, Y.; Yin, L.; Li, Y.; Han, Y.; Cai, C.; Shen, H.; Zeng, S. Zeb1 promotes oxaliplatin resistance through the induction of epithelial-mesenchymal transition in colon cancer cells. J. Cancer 2017, 8, 3555–3566. [Google Scholar] [CrossRef]
- Xu, K.; Zhan, Y.; Yuan, Z.; Qiu, Y.; Wang, H.; Fan, G.; Wang, J.; Li, W.; Cao, Y.; Shen, X.; et al. Hypoxia induces drug resistance in colorectal cancer through the hif-1alpha/mir-338-5p/il-6 feedback loop. Mol. Ther. 2019, 27, 1810–1824. [Google Scholar] [CrossRef]
- Park, C.V.; Ivanova, I.G.; Kenneth, N.S. Xiap upregulates expression of hif target genes by targeting hif1alpha for lys63-linked polyubiquitination. Nucleic Acids Res. 2017, 45, 9336–9347. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Li, N.; Zhao, Y.; Wang, W.; Feng, X.L. Salidroside induces apoptosis in human ovarian cancer skov3 and a2780 cells through the p53 signaling pathway. Oncol. Lett. 2018, 15, 6513–6518. [Google Scholar] [CrossRef]
- Ma, J.; Li, J.; Wang, K.S.; Mi, C.; Piao, L.X.; Xu, G.H.; Li, X.; Lee, J.J.; Jin, X. Perillyl alcohol efficiently scavenges activity of cellular ros and inhibits the translational expression of hypoxia-inducible factor-1alpha via mtor/4e-bp1 signaling pathways. Int. Immunopharmacol. 2016, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Hu, Z.; Zhao, Q.; Zhang, X.; Zhang, H.; Wang, H.; Xue, W.; Yu, L.; Duan, G. Down-regulation of hypoxia-inducible factor-1alpha and downstream glucose transporter protein-1 gene by beta-elemene enhancing the radiosensitivity of lung adenocarcinoma transplanted tumor. Onco. Targets Ther. 2020, 13, 11627–11635. [Google Scholar] [CrossRef]
- Yu, X.; Li, Z.; Zhang, Y.; Xu, M.; Che, Y.; Tian, X.; Wang, R.; Zou, K.; Zou, L. Beta-elemene inhibits radiation and hypoxia-induced macrophages infiltration via prx-1/nf-kappab/hif-1alpha signaling pathway. Onco. Targets Ther. 2019, 12, 4203–4211. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, G.; Zheng, Z.; Lu, B.; Ji, L. Andrographolide reduced vegfa expression in hepatoma cancer cells by inactivating hif-1alpha: The involvement of jnk and mta1/hdca. Chem. Biol. Interact. 2017, 273, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Li, L.; Qi, W.; Liu, B.; Jiang, Z.; Song, J.; Hua, H. Balanophorin b inhibited glycolysis with the involvement of hif-1alpha. Life Sci. 2021, 267, 118910. [Google Scholar] [CrossRef]
- Kim, H.J.; Cho, H.S.; Ban, H.S.; Nakamura, H. Suppression of hif-1alpha accumulation by betulinic acid through proteasome activation in hypoxic cervical cancer. Biochem. Biophys. Res. Commun. 2020, 523, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.F.; Zhang, Z.H.; Li, M.Y.; Wang, J.Y.; Xing, Y.; Ri, M.; Jin, C.H.; Xu, G.H.; Piao, L.X.; Zuo, H.X.; et al. Britannin stabilizes t cell activity and inhibits proliferation and angiogenesis by targeting pd-l1 via abrogation of the crosstalk between myc and hif-1alpha in cancer. Phytomedicine 2021, 81, 153425. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, X.; Zhao, P.; Zhao, H.; Gao, W.; Wang, L. Celastrol suppresses glioma vasculogenic mimicry formation and angiogenesis by blocking the pi3k/akt/mtor signaling pathway. Front. Pharmacol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Leong, S.W.; Wang, J.; Wu, Q.; Ghauri, M.A.; Sarwar, A.; Su, Q.; Zhang, Y. Cephalomannine inhibits hypoxia-induced cellular function via the suppression of apex1/hif-1alpha interaction in lung cancer. Cell Death Dis. 2021, 12, 490. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, C.; Duanmu, J.; Wu, Y.; Tao, J.; Yang, A.; Yin, X.; Xiong, B.; Gu, J.; Li, C.; et al. Cryptotanshinone inhibits the growth and invasion of colon cancer by suppressing inflammation and tumor angiogenesis through modulating mmp/timp system, pi3k/akt/mtor signaling and hif-1alpha nuclear translocation. Int. Immunopharmacol. 2018, 65, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.X.; Jin, Y.; Wang, Z.; Li, M.Y.; Zhang, Z.H.; Wang, J.Y.; Xing, Y.; Ri, M.H.; Jin, C.H.; Xu, G.H.; et al. Curcumol inhibits the expression of programmed cell death-ligand 1 through crosstalk between hypoxia-inducible factor-1alpha and stat3 (t705) signaling pathways in hepatic cancer. J. Ethnopharmacol. 2020, 257, 112835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qiao, X.; Chen, M.; Li, P.; Wen, X.; Sun, M.; Ma, X.; Hou, Y.; Yang, J. Ilexgenin a prevents early colonic carcinogenesis and reprogramed lipid metabolism through hif1alpha/srebp-1. Phytomedicine 2019, 63, 153011. [Google Scholar] [CrossRef]
- Wang, K.S.; Ma, J.; Mi, C.; Li, J.; Lee, J.J.; Jin, X. Kamebakaurin inhibits the expression of hypoxia-inducible factor-1alpha and its target genes to confer antitumor activity. Oncol. Rep. 2016, 35, 2045–2052. [Google Scholar] [CrossRef][Green Version]
- Kong, P.; Yu, K.N.; Yang, M.; Almahi, W.A.; Nie, L.; Chen, G.; Han, W. Micheliolide enhances radiosensitivities of p53-deficient non-small-cell lung cancer via promoting hif-1alpha degradation. Int. J. Mol. Sci. 2020, 21, 3392. [Google Scholar] [CrossRef]
- Wang, Z.; Li, M.Y.; Zhang, Z.H.; Zuo, H.X.; Wang, J.Y.; Xing, Y.; Ri, M.; Jin, H.L.; Jin, C.H.; Xu, G.H.; et al. Panaxadiol inhibits programmed cell death-ligand 1 expression and tumour proliferation via hypoxia-inducible factor (hif)-1alpha and stat3 in human colon cancer cells. Pharmacol. Res. 2020, 155, 104727. [Google Scholar] [CrossRef]
- Park, J.H.; Yoon, J.; Park, B. Pomolic acid suppresses hif1alpha/vegf-mediated angiogenesis by targeting p38-mapk and mtor signaling cascades. Phytomedicine 2016, 23, 1716–1726. [Google Scholar] [CrossRef]
- Lee, S.O.; Kim, J.S.; Lee, M.S.; Lee, H.J. Anti-cancer effect of pristimerin by inhibition of hif-1alpha involves the sphk-1 pathway in hypoxic prostate cancer cells. BMC Cancer 2016, 16, 701. [Google Scholar] [CrossRef]
- Li, G.; Shan, C.; Liu, L.; Zhou, T.; Zhou, J.; Hu, X.; Chen, Y.; Cui, H.; Gao, N. Tanshinone iia inhibits hif-1alpha and vegf expression in breast cancer cells via mtor/p70s6k/rps6/4e-bp1 signaling pathway. PLoS ONE 2015, 10, e0117440. [Google Scholar]
- Li, B.; Tong, T.; Ren, N.; Rankin, G.O.; Rojanasakul, Y.; Tu, Y.; Chen, Y.C. Theasaponin e1 inhibits platinum-resistant ovarian cancer cells through activating apoptosis and suppressing angiogenesis. Molecules 2021, 26, 1681. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kim, G.H.; Park, E.J.; Oh, T.I.; Lee, S.; Kan, S.Y.; Kang, H.; Kim, B.M.; Kim, J.H.; Lim, J.H. Thymoquinone selectively kills hypoxic renal cancer cells by suppressing hif-1alpha-mediated glycolysis. Int. J. Mol. Sci. 2019, 20, 1092. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhou, X.; Jiang, B.; Zhao, Q.; Zhou, G. Triptolide suppresses proliferation, hypoxia-inducible factor-1alpha and c-myc expression in pancreatic cancer cells. Mol. Med. Rep. 2015, 12, 4508–4513. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.Z.; Xuan, Y.Y.; Zhang, Q.; Huang, J.J. Ursolic acid sensitized colon cancer cells to chemotherapy under hypoxia by inhibiting mdr1 through hif-1alpha. J. Zhejiang Univ. Sci. B 2016, 17, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhang, Q.; Yu, M.; Qi, X.; Wang, G.; Xiao, L.; Yi, Q.; Jin, W. Ursolic acid sensitizes radioresistant nsclc cells expressing hif-1alpha through reducing endogenous gsh and inhibiting hif-1alpha. Oncol. Lett. 2017, 13, 754–762. [Google Scholar] [CrossRef]
- Wang, W.J.; Sui, H.; Qi, C.; Li, Q.; Zhang, J.; Wu, S.F.; Mei, M.Z.; Lu, Y.Y.; Wan, Y.T.; Chang, H.; et al. Ursolic acid inhibits proliferation and reverses drug resistance of ovarian cancer stem cells by downregulating abcg2 through suppressing the expression of hypoxia-inducible factor-1alpha in vitro. Oncol. Rep. 2016, 36, 428–440. [Google Scholar] [CrossRef]
- Isaacs, J.S.; Jung, Y.J.; Mimnaugh, E.G.; Martinez, A.; Cuttitta, F.; Neckers, L.M. Hsp90 regulates a von hippel lindau-independent hypoxia-inducible factor-1 alpha-degradative pathway. J. Biol. Chem. 2002, 277, 29936–29944. [Google Scholar] [CrossRef]
- Tang, X.; Ding, Q.; Chen, C.; Chen, F.; Zhou, X.; Hong, C.J.; Pan, W. Micheliolide inhibits gastric cancer growth in vitro and in vivo via blockade of the il-6/stat3 pathway. Pharmazie 2019, 74, 175–178. [Google Scholar]
- Antonangeli, F.; Natalini, A.; Garassino, M.C.; Sica, A.; Santoni, A.; Di Rosa, F. Regulation of pd-l1 expression by nf-kappab in cancer. Front. Immunol. 2020, 11, 584626. [Google Scholar] [CrossRef]
- Lim, S.O.; Li, C.W.; Xia, W.; Cha, J.H.; Chan, L.C.; Wu, Y.; Chang, S.S.; Lin, W.C.; Hsu, J.M.; Hsu, Y.H.; et al. Deubiquitination and stabilization of pd-l1 by csn5. Cancer Cell 2016, 30, 925–939. [Google Scholar] [CrossRef]
- Jung, Y.; Isaacs, J.S.; Lee, S.; Trepel, J.; Liu, Z.G.; Neckers, L. Hypoxia-inducible factor induction by tumour necrosis factor in normoxic cells requires receptor-interacting protein-dependent nuclear factor kappa b activation. Biochem. J. 2003, 370, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Doe, M.R.; Ascano, J.M.; Kaur, M.; Cole, M.D. Myc posttranscriptionally induces hif1 protein and target gene expression in normal and cancer cells. Cancer Res. 2012, 72, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lin, J.; Yang, X.; Long, J.; Bai, Y.; Yang, X.; Mao, Y.; Sang, X.; Seery, S.; Zhao, H. Combination regimens with pd-1/pd-l1 immune checkpoint inhibitors for gastrointestinal malignancies. J. Hematol. Oncol. 2019, 12, 42. [Google Scholar] [CrossRef]
- Yu, C.C.; Chen, C.A.; Fu, S.L.; Lin, H.Y.; Lee, M.S.; Chiou, W.Y.; Su, Y.C.; Hung, S.K. Andrographolide enhances the anti-metastatic effect of radiation in ras-transformed cells via suppression of erk-mediated mmp-2 activity. PLoS ONE 2018, 13, e0205666. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, Y.; Tang, N.; Sun, D.; Lan, Y.; Yu, Z.; Zhao, X.; Feng, L.; Zhang, B.; Jin, L.; et al. Andrographolide inhibits breast cancer through suppressing cox-2 expression and angiogenesis via inactivation of p300 signaling and vegf pathway. J. Exp. Clin. Cancer Res. 2018, 37, 248. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.A.; Huang, D.Y.; Lin, W.W. Beclin-1-independent autophagy positively regulates internal ribosomal entry site-dependent translation of hypoxia-inducible factor 1alpha under nutrient deprivation. Oncotarget 2014, 5, 7525–7539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Costa, M.; Gao, J.; Huang, C. Jnk1 mediates degradation hif-1alpha by a vhl-independent mechanism that involves the chaperones hsp90/hsp70. Cancer Res. 2010, 70, 813–823. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, H.B.; Yoon, S.P.; Lim, S.C.; Cha, M.J.; Jeon, Y.J.; Park, S.G.; Chang, I.Y.; You, H.J. Colon cancer progression is driven by apex1-mediated upregulation of jagged. J. Clin. Investig. 2013, 123, 3211–3230. [Google Scholar] [CrossRef]
- Deng, X.; Zhen, P.; Niu, X.; Dai, Y.; Wang, Y.; Zhou, M. Ape1 promotes proliferation and migration of cutaneous squamous cell carcinoma. J. Dermatol. Sci. 2020, 100, 67–74. [Google Scholar] [CrossRef]
- Logsdon, D.P.; Grimard, M.; Luo, M.; Shahda, S.; Jiang, Y.; Tong, Y.; Yu, Z.; Zyromski, N.; Schipani, E.; Carta, F.; et al. Regulation of hif1alpha under hypoxia by ape1/ref-1 impacts ca9 expression: Dual targeting in patient-derived 3d pancreatic cancer models. Mol. Cancer Ther. 2016, 15, 2722–2732. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.S.; Kim, H.N.; Shin, K.D.; Yoon, Y.J.; Kim, S.J.; Han, D.C.; Kwon, B.M. Cryptotanshinone inhibits constitutive signal transducer and activator of transcription 3 function through blocking the dimerization in du145 prostate cancer cells. Cancer Res. 2009, 69, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, L.; Luo, Y.; Odaka, Y.; Awate, S.; Zhou, H.; Shen, T.; Zheng, S.; Lu, Y.; Huang, S. Cryptotanshinone activates p38/jnk and inhibits erk1/2 leading to caspase-independent cell death in tumor cells. Cancer Prev. Res. 2012, 5, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Koo, T.H.; Hwang, B.Y.; Lee, J.J. Kaurane diterpene, kamebakaurin, inhibits nf-kappa b by directly targeting the DNA-binding activity of p50 and blocks the expression of antiapoptotic nf-kappa b target genes. J. Biol. Chem. 2002, 277, 18411–18420. [Google Scholar] [CrossRef]
- Guan, Z.; Chen, J.; Li, X.; Dong, N. Tanshinone iia induces ferroptosis in gastric cancer cells through p53-mediated slc7a11 down-regulation. Biosci. Rep. 2020, 40, BSR20201807. [Google Scholar] [CrossRef]
- Lv, C.; Zeng, H.W.; Wang, J.X.; Yuan, X.; Zhang, C.; Fang, T.; Yang, P.M.; Wu, T.; Zhou, Y.D.; Nagle, D.G.; et al. The antitumor natural product tanshinone iia inhibits protein kinase c and acts synergistically with 17-aag. Cell Death Dis. 2018, 9, 165. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.A.; Dudeja, V.; McCarroll, J.A.; Borja-Cacho, D.; Dawra, R.K.; Grizzle, W.E.; Vickers, S.M.; Saluja, A.K. Triptolide induces pancreatic cancer cell death via inhibition of heat shock protein 70. Cancer Res. 2007, 67, 9407–9416. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, X.; Tian, X.; Shen, C.; Zhang, Q.; Zhang, Y.; Wang, Z.; Wang, F.; Tao, Y. Triptolide inhibits tumor growth by induction of cellular senescence. Oncol. Rep. 2017, 37, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.D.; Lei, X.P.; Zhong, Y.H.; Chen, M.S.; Ke, Y.Y.; Li, Z.; Chen, J.; Huang, L.J.; Zhang, Y.; Liang, L.; et al. Triptolide suppresses the growth and metastasis of non-small cell lung cancer by inhibiting beta-catenin-mediated epithelial-mesenchymal transition. Acta Pharmacol. Sin. 2021, 42, 1486–1497. [Google Scholar] [CrossRef]
- Zeng, A.; Hua, H.; Liu, L.; Zhao, J. Betulinic acid induces apoptosis and inhibits metastasis of human colorectal cancer cells in vitro and in vivo. Bioorg. Med. Chem. 2019, 27, 2546–2552. [Google Scholar] [CrossRef]
- Kim, S.Y.; Hwangbo, H.; Kim, M.Y.; Ji, S.Y.; Kim, D.H.; Lee, H.; Kim, G.Y.; Moon, S.K.; Leem, S.H.; Yun, S.J.; et al. Betulinic acid restricts human bladder cancer cell proliferation in vitro by inducing caspase-dependent cell death and cell cycle arrest, and decreasing metastatic potential. Molecules 2021, 26, 1381. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Cao, J.; Chen, K.; Cheng, L.; Zhou, C.; Yan, B.; Qian, W.; Li, J.; Duan, W.; Ma, J.; et al. Betulinic acid inhibits stemness and emt of pancreatic cancer cells via activation of ampk signaling. Int. J. Oncol. 2019, 54, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.K.; Li, J.L.; Zhang, S.; Xing, P.Y.; Xia, M.F. Betulinic acid exerts potent antitumor effects on paclitaxel-resistant human lung carcinoma cells (h460) via g2/m phase cell cycle arrest and induction of mitochondrial apoptosis. Oncol. Lett. 2018, 16, 3628–3634. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Zhou, J.; Lv, H.; Yu, Q.; Peng, L.; Hua, S. Betulin exhibits anti-inflammatory activity in lps-stimulated macrophages and endotoxin-shocked mice through an ampk/akt/nrf2-dependent mechanism. Cell Death Dis. 2017, 8, e2798. [Google Scholar] [CrossRef]
- Aiken, C.T.; Kaake, R.M.; Wang, X.; Huang, L. Oxidative stress-mediated regulation of proteasome complexes. Mol. Cell Proteom. 2011, 10, R110.006924. [Google Scholar] [CrossRef] [PubMed]
- Kannaiyan, R.; Shanmugam, M.K.; Sethi, G. Molecular targets of celastrol derived from thunder of god vine: Potential role in the treatment of inflammatory disorders and cancer. Cancer Lett. 2011, 303, 9–20. [Google Scholar] [CrossRef]
- Shi, J.; Li, J.; Xu, Z.; Chen, L.; Luo, R.; Zhang, C.; Gao, F.; Zhang, J.; Fu, C. Celastrol: A review of useful strategies overcoming its limitation in anticancer application. Front. Pharmacol. 2020, 11, 558741. [Google Scholar] [CrossRef]
- Vihanto, M.M.; Plock, J.; Erni, D.; Frey, B.M.; Frey, F.J.; Huynh-Do, U. Hypoxia up-regulates expression of eph receptors and ephrins in mouse skin. FASEB J. 2005, 19, 1689–1691. [Google Scholar] [CrossRef]
- Tang, N.N.; Zhu, H.; Zhang, H.J.; Zhang, W.F.; Jin, H.L.; Wang, L.; Wang, P.; He, G.J.; Hao, B.; Shi, R.H. Hif-1alpha induces ve-cadherin expression and modulates vasculogenic mimicry in esophageal carcinoma cells. World J. Gastroenterol. 2014, 20, 17894–17904. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, C.; Zhang, Y.Q.; Ge, L.T.; Chen, J.; Jia, X.Q.; Gu, R.X.; Sun, Y.; Sun, W.D. Ilexgenin a induces b16-f10 melanoma cell g1/s arrest in vitro and reduces tumor growth in vivo. Int. Immunopharmacol. 2015, 24, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, J.; Fan, J.H.; Zhang, Y.Q.; Zhao, J.X.; Dai, X.J.; Liu, Q.; Shen, Y.J.; Liu, C.; Sun, W.D.; et al. Ilexgenin a exerts anti-inflammation and anti-angiogenesis effects through inhibition of stat3 and pi3k pathways and exhibits synergistic effects with sorafenib on hepatoma growth. Toxicol. Appl. Pharmacol. 2017, 315, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, L.; Rocha, G.D.G.; Queiroz, R.M.; Martins, C.A.; Takiya, C.M.; Gattass, C.R. Pomolic acid induces apoptosis and inhibits multidrug resistance protein mrp1 and migration in glioblastoma cells. Oncol. Rep. 2017, 38, 2525–2534. [Google Scholar] [CrossRef][Green Version]
- Kim, B.; Kim, Y.C.; Park, B. Pomolic acid inhibits metastasis of her2 overexpressing breast cancer cells through inactivation of the erk pathway. Int. J. Oncol. 2016, 49, 744–752. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, B.; Kim, J.H.; Park, B. Pomolic acid inhibits invasion of breast cancer cells through the suppression of cxc chemokine receptor type 4 expression. J. Cell Biochem. 2016, 117, 1296–1307. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.A.; Rocha, G.D.G.; Gattass, C.R.; Takiya, C.M. Pomolic acid exhibits anticancer potential against a docetaxelresistant pc3 prostate cell line. Oncol. Rep. 2019, 42, 328–338. [Google Scholar] [PubMed]
- Ader, I.; Brizuela, L.; Bouquerel, P.; Malavaud, B.; Cuvillier, O. Sphingosine kinase 1: A new modulator of hypoxia inducible factor 1alpha during hypoxia in human cancer cells. Cancer Res. 2008, 68, 8635–8642. [Google Scholar] [CrossRef]
- Bouquerel, P.; Gstalder, C.; Muller, D.; Laurent, J.; Brizuela, L.; Sabbadini, R.A.; Malavaud, B.; Pyronnet, S.; Martineau, Y.; Ader, I.; et al. Essential role for sphk1/s1p signaling to regulate hypoxia-inducible factor 2alpha expression and activity in cancer. Oncogenesis 2016, 5, e209. [Google Scholar] [CrossRef]
- Meurette, O.; Stylianou, S.; Rock, R.; Collu, G.M.; Gilmore, A.P.; Brennan, K. Notch activation induces akt signaling via an autocrine loop to prevent apoptosis in breast epithelial cells. Cancer Res. 2009, 69, 5015–5022. [Google Scholar] [CrossRef]
- Wang, L.; Zi, H.; Luo, Y.; Liu, T.; Zheng, H.; Xie, C.; Wang, X.; Huang, X. Inhibition of notch pathway enhances the anti-tumor effect of docetaxel in prostate cancer stem-like cells. Stem Cell Res. Ther. 2020, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.D.; Chaudhary, N.; Seo, H.J.; Kim, M.Y.; Shin, T.S. Theasaponin e(1) as an effective ingredient for anti-angiogenesis and anti-obesity effects. Biosci. Biotechnol. Biochem. 2014, 78, 279–287. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, L.; Shi, H.; Chen, H.; Tao, J.; Shen, R.; Wang, T. Ursolic acid enhances the therapeutic effects of oxaliplatin in colorectal cancer by inhibition of drug resistance. Cancer Sci. 2018, 109, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Xiang, F.; Fan, Y.; Ni, Z.; Liu, Q.; Zhu, Z.; Chen, Z.; Hao, W.; Yue, H.; Wu, R.; Kang, X. Ursolic acid reverses the chemoresistance of breast cancer cells to paclitaxel by targeting mirna-149-5p/myd88. Front. Oncol 2019, 9, 501. [Google Scholar] [CrossRef] [PubMed]



| Compound | Compound Class | Type of Cancer | In Vitro Testing (Effective Concentrations, Cell Line, Culture Condition (Normoxia/Hypoxia)) | In Vivo Experiment Model (Dose and Administration Route) | Ref. |
|---|---|---|---|---|---|
| Polyphenols (Flavonoids) | |||||
| Apigenin | Trihydroxyflavone | Prostate, Ovarian, Colon, and Breast cancer | 10–40 μM (human PC-3, DU145, MCF-7, HCT-8, LNCaP cell lines), hypoxia (1% O2) | PC-3 and OVCAR-3-bearing CAM assay (7.5–20 μM), Matrigel plug assays (15–20 μM) | [76] |
| Head and Neck cancer | 20–40 μM (human HN-30 cell line), hypoxia (0.5–1% O2) | - | [77] | ||
| Cardamonin | Chalcone | Breast cancer | 20–40 μM (human MDA-MB-231 cell line), hypoxia (CoCl2) | Subcutaneous injection of MDA-MB-231 cells (3 mg/kg, intra-peritoneal) | [82] |
| Epigallocatechin-3-gallate (EGCG) | Flavanol | Breast cancer | 5–20 μM (human MCF-7 cell line), normoxia | - | [83] |
| FV-429 | Wogonin derivative | Ovarian cancer | 5–20 μM (human SK-OV-3 and A2780 cell lines), hypoxia (1% O2) | Subcutaneous injection of A2780 cells (10 mg/kg) | [84] |
| Oroxylin A | Dihydroxyflavone | Glioblastoma | 25–20 μM (human U251 cell line), 10–20 μM (rat C6 cell line), hypoxia (1% O2) | Intracranial transplantations of U251 cells or subcutaneous injections of mouse GL261 cell line (300 mg/kg, oral) | [85] |
| Lung cancer | 50 μM (human H460, A549, 95D, PC9, HCC827 and H1975 cell lines), hypoxia (1% O2) | Subcutaneous injections of H460 cells (50 mg/kg, oral) | [86] | ||
| Wogonin | Hydroxyflavone | Gastric cancer | 20–100 μM (human SGC-7901 cell line), normoxia | - | [87] |
| Y6 | EGCG derivative | Hepatocellular carcinoma | 10–15 μM (doxorubicin-resistant human BEL-7404 cell line), normoxia | - | [88] |
| Hepatocellular carcinoma | 10 μg/mL (human SMMC-7721 cell line), hypoxia (1% O2) | CAM model for angiogenesis assay (200–500 μg/mL), Subcutaneous injections of HepG2 cells (55 mg/kg, oral) | [89] | ||
| Polyphenols (Lignans, Phenolic Acids, and Stilbenes) | |||||
| HS-1793 | Resveratrol analogue | Breast cancer | 12.5–50 μM (human MCF-7 and MDA-MB-231 cell lines), hypoxia (1% O2) | Subcutaneous injections of MDA-MB-231 cells (5–10 mg/kg, intraperitoneal) | [90] |
| LXY6090 | Manassantin A derivative | Breast cancer | 0.4–10 nM (human T47D, MCF-7, and MX-1 cell lines), hypoxia (1% O2) | Subcutaneous injections of MX-1 cells (25–100 mg/kg, oral) | [91] |
| Magnolol | Lignan | Bladder cancer | 1–10 μM (human T24 cell line), hypoxia (1% O2) | CAM model of T24 cells (1–10 μM), Matrigel plug assays (25–75 μg), Subcutaneous injections of T24 cells (2–10 mg/kg, intraperitoneal) | [92] |
| Manassantin A | Lignan | Lung cancer | 0.01–10 μM (luciferase-reporter assay using human embryonic kidney 293T cells), hypoxia (1% O2) | Lewis lung carcinoma allografts (5 mg/kg, intraperitoneal) | [93] |
| Piceatannol | Stilbene | Colorectal cancer | 7.5 μg/mL of piceatannol-loaded nanoparticles (PNs) (human CaCo-2 and HT-29 cell lines), normoxia | Colitis-associated colorectal cancer mouse model (40 mg/kg) | [94] |
| Pterostilbene | Stilbene | Prostate cancer | 50 μM (human LNCaP and PC3M cell lines), normoxia | Pten-null mouse model (10 mg/kg, intraperitoneal) | [95] |
| Resveratrol | Stilbene | Lung cancer | - | Orthotopic injections of A549 cells (250 mg/kg, intragastric) | [96] |
| Gastric cancer | 12.5–100 μM (human SGC-7901 cell line), hypoxia (3% O2) | - | [97] | ||
| Pancreatic cancer | 50 μM (human pancreatic stellate cells from normal tissues), hypoxia (3% O2) | KPC mouse model of pancreatic cancer (50 mg/kg) | [98] | ||
| Rhaponticin | Stilbene | Fibrosarcoma | 25–100 μM (human HT1080 cell line), hypoxia (CoCl2) | - | [99] |
| Vanillic acid | Phenolic acid | Colorectal cancer | 3–30 μM (human HCT-116 cell line), hypoxia (1% O2, CoCl2) | Subcutaneous injections of HCT-116 cells (10–30 mg/kg, oral) | [100] |
| Other Polyphenols | |||||
| Chlorogenic acid | Tannin | Lung cancer | 2–10 μM (human A549 cell line), hypoxia (1% O2, CoCl2) | Matrigel plug assays (10 μM) | [101] |
| Curcumin | Diarylheptanoid | Chronic myelogenous leukemia | 20 μM (human K526 cell line), normoxia | - | [102] |
| Decursin | Pyranocoumarin | Lung and Colorectal cancer | 10–50 μM (human A549 and HCT-116 cell lines), hypoxia (1% O2, CoCl2) | Lewis lung carcinoma allografts (10 mg/kg, intraperitoneal) | [103] |
| DPHP | Alpinoid c (diarylheptanoid) derivative | Colorectal cancer | 3.5–14 μM (human COLO205 cell line), hypoxia (CoCl2) | CAM model for angiogenesis assay (3.5–14 μM) | [104] |
| Gambogic acid | Xanthone | Multiple myeloma | 0.1–0.2 μM (human U266 cell line), hypoxia (1% O2) | Subcutaneous injections of U266 cells (2–4 mg/kg, intravenous) | [105] |
| Garcinol | Polyisoprenylated benzophenone | Colorectal cancer | 20–60 μM (human HT-29 cell line), normoxia | - | [106] |
| Imperatorin | Furanocoumarin | Colorectal cancer | 50–150 μM (human HCT-116 cell line), hypoxia (1% O2, CoCl2) | Subcutaneous injections of HCT-116 cells (50–100 mg/kg, oral) | [107] |
| Salidroside | Phenylethanoid | Hepatocellular carcinoma | 100 μM (human PLC/PRF/5, SMMC-7721, and HepG2 cell lines), hypoxia (1% O2) | Subcutaneous or orthotopic injections of PLC/PRF/5 cells (60 mg/kg, intragastric) | [108] |
| Shikonin | Naphthoquinone | Colorectal cancer | 1–10 μM (human SW-620 and HCT-116 cell lines), hypoxia (1% O2) | Subcutaneous injections of HCT-116 cells (2–10 mg/kg, oral) | [109] |
| Verbascoside | Phenylethanoid glycoside | Colorectal cancer | 50–150 μM (human HT-29 cell line), normoxia | - | [110] |
| Compound | Compound Class | Type of Cancer | In Vitro Testing (Effective Concentrations, Cell Line, Culture Condition (Normoxia/Hypoxia) | In Vivo Experiment Model (Dose and Administration Route) | Ref. |
|---|---|---|---|---|---|
| β-elemene | Sesquiterpene | Lung cancer | - | Subcutaneous injections of A549 cells (45 mg/kg) | [165] |
| Lung cancer | - | Lewis lung carcinoma allografts (45 mg/kg, intraperitoneal) | [166] | ||
| Andrographolide | Diterpene | Hepatocellular carcinoma | 25–50 μM (human Hep3B and HepG2 cell lines), normoxia | Subcutaneous injections of Hep3B cells (10 mg/kg, intraperitoneal) | [167] |
| Balanophorin B | Triterpene | Hepatocellular carcinoma | 25–50 μM (human Huh-7 and HepG2 cell lines), hypoxia (1% O2) | Subcutaneous injections of HepG2 cells (50–100 mg/kg, oral) | [168] |
| Betulinic acid | Triterpene | Cervical cancer | 3–30 μM (human HeLa cell line), hypoxia (1% O2) | - | [169] |
| Britannin | Sesquiterpene | Colorectal cancer | 1–10 μM (human HCT-116 cell line), normoxia | Subcutaneous injections of HCT-116 cells (5–15 mg/kg, oral) | [170] |
| Celastrol | Triterpene | Glioblastoma | 0.25–1 μM (human U87 and U251 cell lines), normoxia | Orthotopic injections of U87 cells (0.5–2 mg/kg, intraperitoneal) | [171] |
| Cephalomannine | Diterpene | Lung cancer | 0.025–0.1 μM (human A549 and H460 cell lines), hypoxia (1% O2) | Subcutaneous injections of H460 cells (0.4 mg/kg, intraperitoneal) | [172] |
| Cryptotanshinone | Diterpene | Colorectal cancer | 5–20 μM (mouse CT26 cell line), normoxia | Subcutaneous injections of CT26 cells (20–80 mg/kg, oral) | [173] |
| Curcumol | Sesquiterpene | Hepatocellular carcinoma | 3–30 μM (human Hep3B cell line), hypoxia (1% O2) | Subcutaneous injections of Hep3B cells (3–30 mg/kg, oral) | [174] |
| Ilexgenin A | Triterpene | Colorectal cancer | 25–50 μM (human HT-29 and HCT-116 cell lines), hypoxia (1% O2) | Colitis-associated colorectal cancer mouse model (20 mg/kg) | [175] |
| Kamebakaurin | Diterpene | Colorectal cancer | 10–30 μM (human HCT-116 cell line), hypoxia (CoCl2) | Subcutaneous injections of HCT-116 cells (15–50 mg/kg, oral) | [176] |
| Micheliolide | Sesquiterpene | Lung cancer | 5–20 μM (human H1299 and Calu-1 cell lines), hypoxia (1% O2) | - | [177] |
| Panaxadiol | Triterpene | Colorectal cancer | 10 μM (human HCT-116 cell line), hypoxia (1% O2) | Subcutaneous injections of HCT-116 cells (10–30 mg/kg, oral) | [178] |
| Perillyl alcohol | Monoterpene | Cervical and Colorectal cancer, Hepatocellular carcinoma | 50–200 μM (human HCT-116, HeLa, and SK-Hep1 cell lines), hypoxia (1% O2, CoCl2) | Subcutaneous injections of HCT-116 cells (50–100 mg/kg, oral) | [164] |
| Pomolic acid | Triterpene | Breast cancer | 1–10 μM (human MCF-7 and MDA-MB-231 cell lines), hypoxia (CoCl2) | - | [179] |
| Pristimerin | Triterpene | Prostate cancer | 1 μM (human PC-3, DU145, and LNCaP cell lines), hypoxia (1% O2) | - | [180] |
| Tanshinone IIA | Diterpene | Breast cancer | 2.5–20 μM (human MCF-7 and MDA-MB-231 cell lines), hypoxia (1% O2) | Subcutaneous injections of MDA-MB-231 cells (50 mg/kg, intraperitoneal) | [181] |
| Theasaponin E1 | Triterpene | Ovarian cancer | 1–5 μM (human OVCAR-3 and A2780/CP70 cell lines), normoxia | CAM model of OVCAR-3 cells (4 μM) | [182] |
| Thymoquinone | Monoterpene | Renal cancer | 5–10 μM (human Caki-1, Caki-2, and A498 cell lines), hypoxia (1% O2) | - | [183] |
| Triptolide | Diterpene | Pancreatic cancer | 55–140 μM (human SW1990 cell line), normoxia | Subcutaneous injections of SW1990 cells (0.2–0.8 mg/kg, intraperitoneal) | [184] |
| Ursolic acid | Triterpene | Colorectal cancer | 20–40 μM (human RKO, LoVo, and SW480 cell lines), hypoxia (1% O2) | - | [185] |
| Lung cancer | 50–80 μM (human H1299 cell line), normoxia | - | [186] | ||
| Ovarian cancer | 6.5–65 μM (spheroid cultures of human SKOV3 cell line), hypoxia (1% O2) | - | [187] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, B.D.; Son, S.W.; Choi, S.Y.; Kuh, H.J.; Oh, T.-J.; Park, J.K. Anti-Cancer Activity of Phytochemicals Targeting Hypoxia-Inducible Factor-1 Alpha. Int. J. Mol. Sci. 2021, 22, 9819. https://doi.org/10.3390/ijms22189819
Yun BD, Son SW, Choi SY, Kuh HJ, Oh T-J, Park JK. Anti-Cancer Activity of Phytochemicals Targeting Hypoxia-Inducible Factor-1 Alpha. International Journal of Molecular Sciences. 2021; 22(18):9819. https://doi.org/10.3390/ijms22189819
Chicago/Turabian StyleYun, Ba Da, Seung Wan Son, Soo Young Choi, Hyo Jeong Kuh, Tae-Jin Oh, and Jong Kook Park. 2021. "Anti-Cancer Activity of Phytochemicals Targeting Hypoxia-Inducible Factor-1 Alpha" International Journal of Molecular Sciences 22, no. 18: 9819. https://doi.org/10.3390/ijms22189819
APA StyleYun, B. D., Son, S. W., Choi, S. Y., Kuh, H. J., Oh, T.-J., & Park, J. K. (2021). Anti-Cancer Activity of Phytochemicals Targeting Hypoxia-Inducible Factor-1 Alpha. International Journal of Molecular Sciences, 22(18), 9819. https://doi.org/10.3390/ijms22189819