Abstract
Glomerella leaf spot of apple, caused by Colletotrichum gloeosporioides, is a devastating disease that leads to severe defoliation and fruit spots. The Colletotrichum species secretes a series of effectors to manipulate the host’s immune response, facilitating its colonization in plants. However, the mechanism by which the effector of C. gloeosporioides inhibits the defenses of the host remains unclear. In this study, we reported a novel effector Sntf2 of C. gloeosporioides. The transient expression of SNTF2 inhibits BAX-induced cell death in tobacco plants. Sntf2 suppresses plant defense responses by reducing callose deposition and H2O2 accumulation. SNTF2 is upregulated during infection, and its deletion reduces virulence to the plant. Sntf2 is localized to the chloroplasts and interacts with Mdycf39 (a chloroplast PSII assembly factor) in apple leaves. The Mdycf39 overexpression line increases susceptibility to C. gloeosporioides, whereas the Mdycf39 transgenic silent line does not grow normally with pale white leaves, indicating that Sntf2 disturbs plant defense responses and growth by targeting Mdycf39.
1. Introduction
In agricultural and natural ecosystems, plants are exposed to numerous pathogens. Nevertheless, plants have complex defense systems to combat pathogen invasion [1]. When pathogen-associated molecular patterns (PAMPs) are recognized by pattern recognition receptors (PRRs) on the cell surface, the plant’s basal defense response is activated, termed PAMP-triggered immunity (PTI) [2]. Damage-associated molecular patterns (DAMPs)-triggered immunity (DTI) plays an important role in a plant’s basal defense response [3,4]. Plants have evolved intracellular nucleotide-binding leucine-rich-repeat receptors (NLRs) to recognize pathogen effectors directly or indirectly, leading to activation of the second line of defense, known as effector-triggered immunity (ETI) [2]. However, pathogens secrete a series of virulence effectors that interfere with the plant’s immune system, promoting pathogen invasion [5,6]. Hence, exploring the virulence mechanisms of effectors is important for revealing the infection strategies of plant pathogens.
The Colletotrichum species, belonging to Glomerellaceae of Ascomycota, develop penetration pegs from appressoria to invade the host [7]. Colletotrichum deploys distinct effectors at different infection phases to manipulate the host plant’s immune response [8]. Although various effectors have been identified [9,10], few have investigated their specific function mechanisms [11,12]. The interaction mechanism between effectors and host target proteins of Colletotrichum mainly focused on Colletotrichum higginsianum, Colletotrichum fruticola, and Colletotrichum orbiculare [13,14,15]. Each Colletotrichum species has evolved a set of effectors with unique strategies to adapt to its host plants [16]. In C. gloeosporioides, many studies focused on identifying the pathogenicity-related genes [17,18], but few focused on characterizing the mechanisms of effectors in the interactions between C. gloeosporioides and host plants.
C. gloeosporioides is a plant pathogenic fungus that infects various plants, including apple, mango, and poplar [19]. Glomerella leaf spot of apple (GLSA), caused by C. gloeosporioides, is a devastating disease that severely affects apple production [20,21]. Under favorable conditions, the latent period of this disease is as short as two days, and it spreads rapidly [19,22], causing necrotic fruit spots and severe defoliation in ‘Royal Gala’ and ‘Golden Delicious’ apples. Todate, the control of GLSA remains a challenge. Understanding the molecular interactions between C. gloeosporioides and apples is instrumental for sustaining effective disease control and developing disease-resistant varieties.
Chloroplasts not only play a role as photosynthetic organelles but also play a central role in plant defense by integrating environmental stimuli and the determinants of downstream defense responses [23,24,25]. Upon perception of pathogenic threat, chloroplast as the source of calcium, salicylic acid (SA), and reactive oxygen species (ROS) bursts communicates with the nucleus through retrograde signaling [24], mediates activation of plant immune signaling, and leads to the expression of defense-related genes [26,27]. To interfere with the function of chloroplasts in the interaction between the pathogens and the hosts, pathogens, including bacteria, viruses, fungi, and Oomycetes, have deployed effectors to target chloroplasts [6,23,27,28,29]. However, the underlying mechanisms with which effectors target chloroplasts to manipulate the host’s defenses have remained elusive in Colletotrichum.
In this study, we found that a novel effector Sntf2 of C. gloeosporioides played an essential role in suppressing the plant’s defense responses. Sntf2 is an extracellular-secreted protein including a signal peptide in its N-terminal, which plays a vital role in plant infection. Consistent with this, the deletion of SNTF2 triggered the plant’s defense responses, in which H2O2 accumulation and callose deposition increased in the apple leaves inoculated with the Δsntf2-1 mutant. We showed that Sntf2 could inhibit Bcl-2-associated X protein (BAX)-induced cell death, suppressing hypersensitive responses in plants. Our investigation demonstrated that Sntf2 could migrate into a host plant’s cells and interact with the Mdycf39 (a photosystem II assembly factor of Malus domestica). We also found that Mdycf39 played a vital role in plant resistance and development. Overall, our results show that Sntf2 perturbs the function of chloroplast, avoiding to trigger cell death and supporting pathogen colonization on live plants.
2. Results
2.1. Sntf2 Inhibits BAX-Induced Cell Death in Tobacco
BAX, a pro-apoptotic protein, stimulates cell death, which closely resembles the hypersensitive responsive (HR) in plants [30]. SNTF2 (CGGC5_13909) encodes a putative protein in C. gloeosporioides with 187 amino acid residues and contains a predicted signal peptide (SP; 1–20 aa) (Figure 1a). To investigate if Sntf2 inhibited a plant’s innate immunity, we performed transient expression of the gene in Nicotiana benthamiana. The Agrobacterium strain GV3101 carrying the potato virus X (PVX) vector pGR106-fused SNTF2 gene was infiltrated into the leaves of N. benthamiana, and the strain carrying pGR106-BAX was injected into the same sites on the leaves after 24 h post-infiltration. We found that Sntf2 suppressed the cell death triggered by BAX in the infiltrated leaves (Figure 1b). These data indicated that Sntf2 suppressed the HR response of the plant.
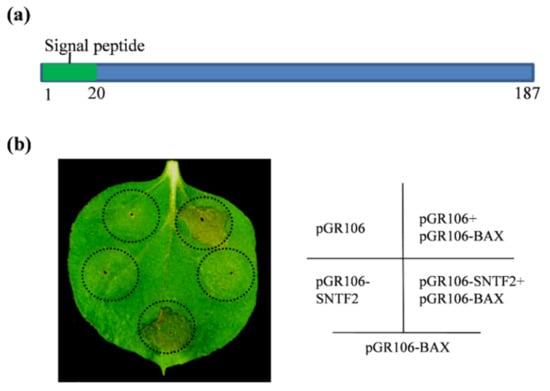
Figure 1.
Sntf2 suppressed BAX-triggered cell death in Nicotiana benthamiana. (a) The structure analysis of Sntf2. Sntf2 was predicted to contain an N-terminal signal peptide (1–20 aa). (b) The tobacco leaves infiltrated with Agrobacterium tumefaciens GV3101 harboring the pGR106 vector or the vector with SNTF2 inserted. The A. tumefaciens carrying the BAX gene were injected into the leaves after 24 h post-infiltration. The empty pGR106 vector was used as the control. The black dotted lines indicate the region of infiltration. The diagram on the right shows the transformant strains of A. tumefaciens (containing recombinant pGR106 vectors) that was injected inside the black dotted lines. Images were acquired at 7 days post-infiltration. The experiments were repeated six times.
2.2. SNTF2 Is Upregulated during Biotrophic Infection Phase
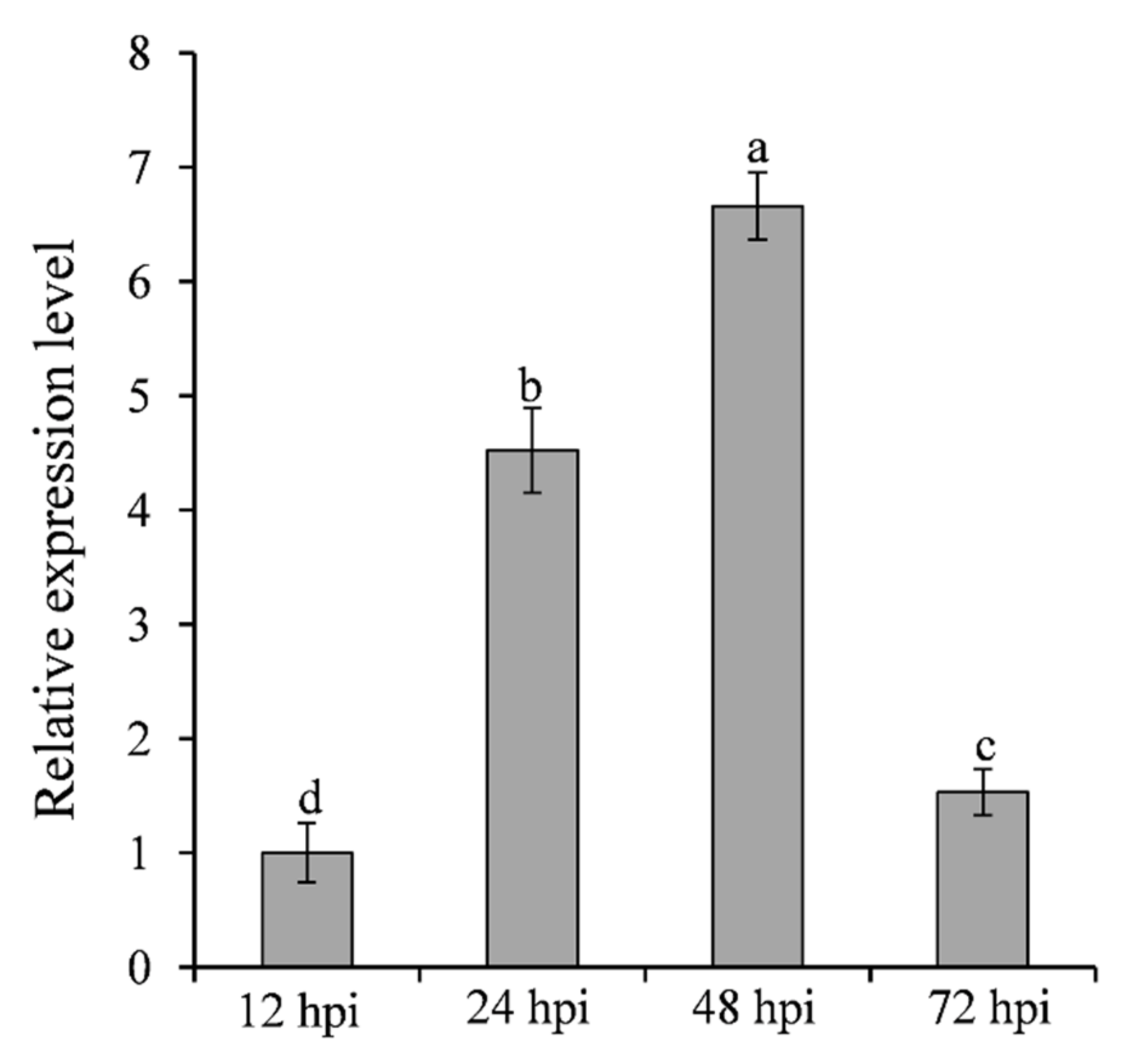
To analyze the expression pattern of SNTF2 at different infection phases, the transcripts of SNTF2 were examined in infected leaves at 12, 24, 48, and 72 h post-inoculation (hpi) using qRT-PCR. SNTF2 was upregulated at 24 hpi and 48 hpi (Figure 2). Appressoria formed at 12 hpi, and appressorium-mediated penetration occurred at 24 hpi (Figure S1). Fungal primary infection hyphae formed at 48 hpi (Figure S1). This meant that SNTF2 was highly expressed from appressorium-mediated penetration to infection-hyphae formation. In addition, fluorescence signal detection showed that Sntf2-eGFP was actually expressed during the infection in apple leaves (Figure S1). These results suggest that SNTF2 may play an important role in plant infection.
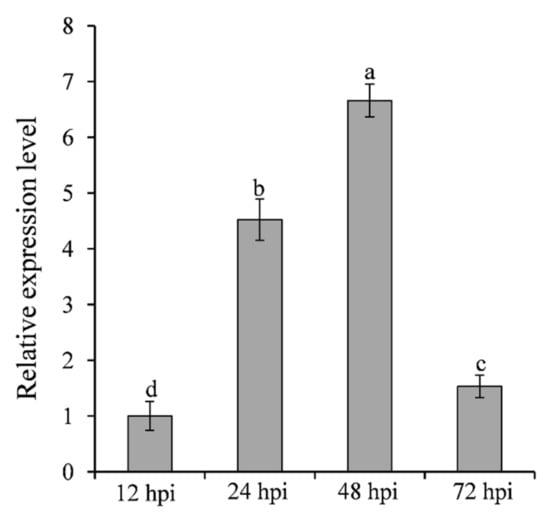
Figure 2.
Analysis of SNTF2 transcription at different infection phases by qRT-PCR. The cDNA was obtained from the apple leaves at 12, 24, 48, and 72 h post-inoculation (hpi) with Colletotrichum gloeosporioides. The M. domestica ubiquitin extension factor (MdUBQ) gene was used as the reference gene to normalize and analysis the transcription of SNTF2. Results were presented as the average fold values from three independent experiments compared with 12 hpi samples. Error bars represent standard deviations. Lowercase letters indicate significant differences (p < 0.01).
2.3. Sntf2 Is Required for the Pathogenicity of C. gloeosporioides
To determine the function of SNTF2 in the pathogenicity, SNTF2 deletion mutants were constructed using A. tumefaciens-mediated homologous recombination (Figure S2a). The Δsntf2 mutants were confirmed by PCR and Southern blot analysis (Figure S2b–d). The complementation strain Δsntf2-1/SNTF2 was generated and validated by PCR analysis (Figure S2c,d). The Δsntf2 mutant had a normal growth rate compared with the wild type, as well as conidial production, appressorial formation, and invasive pegs formation (Figure S3). The apple leaves inoculated with the Δsntf2 mutants showed tiny spots compared with the WT (Figure 3), indicating that Sntf2 was required for the pathogenicity of C. gloeosporioides.

Figure 3.
SNTF2 deletion reduced the pathogenicity of Colletotrichum gloeosporioides. The pathogenicity was evaluated based on the disease index. Each experiment was repeated three times with ten leaves used for each replicate. Error bars represent standard deviations. Lowercase letters indicate significant differences ( p< 0.01).
2.4. Sntf2 Suppresses Apple Defense Responses
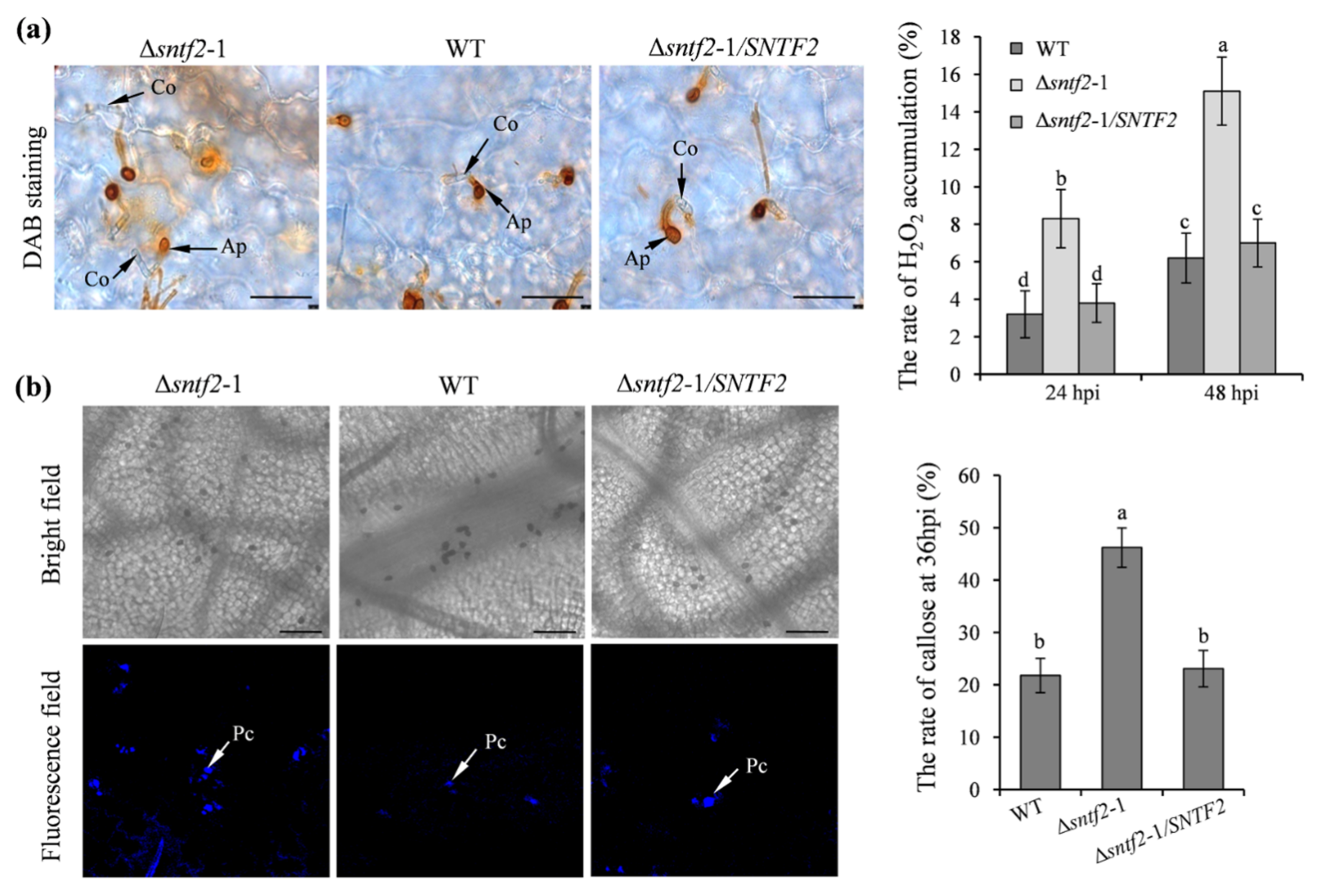
To evaluate if Sntf2 suppressed apple defense responses during the infection, we tested the generation of H2O2 and callose in apple leaves. The 3,3’-diaminobenzidine (DAB) staining result showed that H2O2 accumulation increased at the invasive sites when inoculated with the Δsntf2-1 mutant compared with the WT (Figure S4a). The H2O2 accumulation in the apple leaves inoculated with the complementation strain Δsntf2-1/SNTF2 was similar to the WT (Figure 4a). The rate of callose formation in apple leaves inoculated with the Δsntf2-1 mutant was 24.4% higher than the leaves inoculated with the WT (Figure 4b). Aniline blue staining also showed that callose deposition in leaves inoculated with the Δsntf2-1 mutant increased at 36 hpi compared with the WT (Figure S4b). These results indicated that the deletion of SNTF2 triggered apple defense responses.
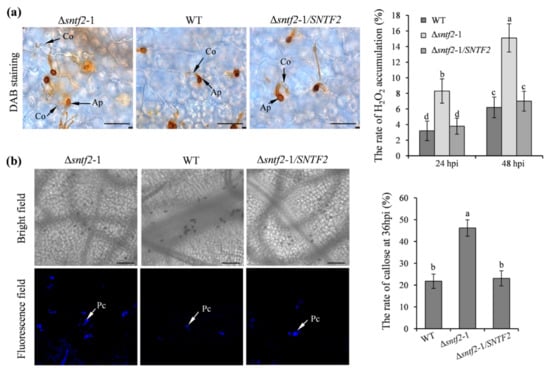
Figure 4.
The H2O2 accumulation and callose deposition in apple leaves during the infection. (a) DAB staining was performed to detect H2O2 accumulation on apple leaves. DAB oxidation led to brownish polymer formation that was deposited at the site of H2O2 accumulation. Co: conidia; Ap: appressorium; bar: 50 μm. (b) Aniline blue staining was to observe callose deposition on apple leaves at 36 hpi. Apple leaves were inoculated with WT, Δsntf2-1, Δsntf2-2, Δsntf2-3, or Δsntf2-1/SNTF2 strains. Pc: callose deposition; bar: 50 μm. The rate of H2O2 accumulation and callose formation was evaluated. Error bars represent standard deviations. Lowercase letters represent significant differences (p < 0.01).
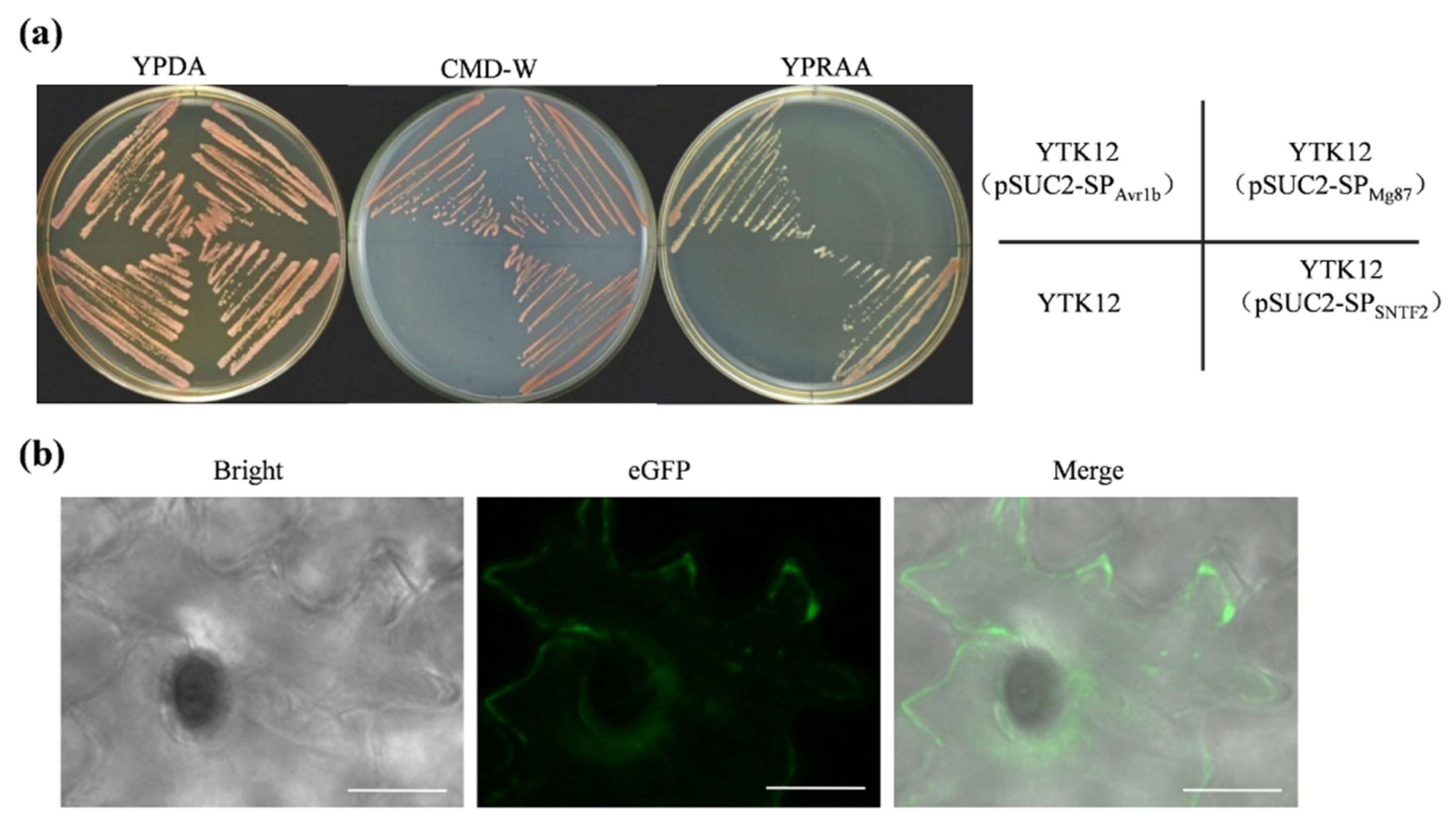
2.5. Sntf2 Is a Secretion Protein
SignalP 4.0 analysis revealed that a SP was present in Sntf2 (SP; 1–20 aa; Figure 1a). To verify the function of this SP, an expression vector harboring the yeast invertase sequence fused to the SP was constructed and transformed into yeast strain YTK12. The transformant containing pSUC2-SPSNTF2 could grow on YPRAA plates (Figure 5a), which was consistent with the positive Avr1b [31], indicating that the SP of Sntf2 was able to secrete invertase. The invertase secreted by the transformant was confirmed by using 2, 3, 5-triphenyltetrazolium chloride (TTC) assays (Figure S5b). To observe the extracellular secretion of Sntf2, we constructed a Sntf2-eGFP expression strain. We found that Sntf2-eGFP was secreted into plant cells (Figure 5b and Figure S1). These results revealed that the SP of Sntf2 was functional in mediating the extracellular secretion of Sntf2. Expectedly, the complementation mutant Δsntf2-1/SNTF2Δsp did not restore fully the pathogenicity of the Δsntf2-1 strain (Figure S5c,d), indicating that the SP of Sntf2 played a vital role in plant infection.
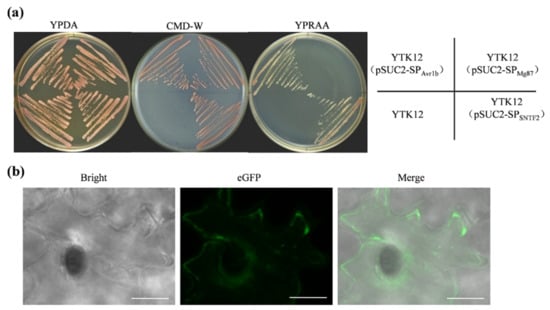
Figure 5.
The secretory function validation of Sntf2. (a) The secretory function test of the predicted signal peptide (SP) of Sntf2 was based on the yeast’s secretory system. The YTK12 strains containing pSUC2-SPAvr1b (SPAvr1b: the SP sequence of Avr1b from Phytophthora sojae) were used as a positive control, and those containing pSUC2-SPMg87 (SPMg87: the first 25 amino acids of Mg87 protein from Magnaporthe oryzae) were used as negative controls. (b) Sntf2-eGFP fusion protein was translocated into plant cells during apple leaf infection. The leaves were inoculated with Δsntf2-1/gpdAp:SNTF2:eGFP strain (gpdAp: the promoter of glyceraldehyde-3-phosphate dehydrogenase gene from Aspergillus nidulans). Bar: 10 μm.
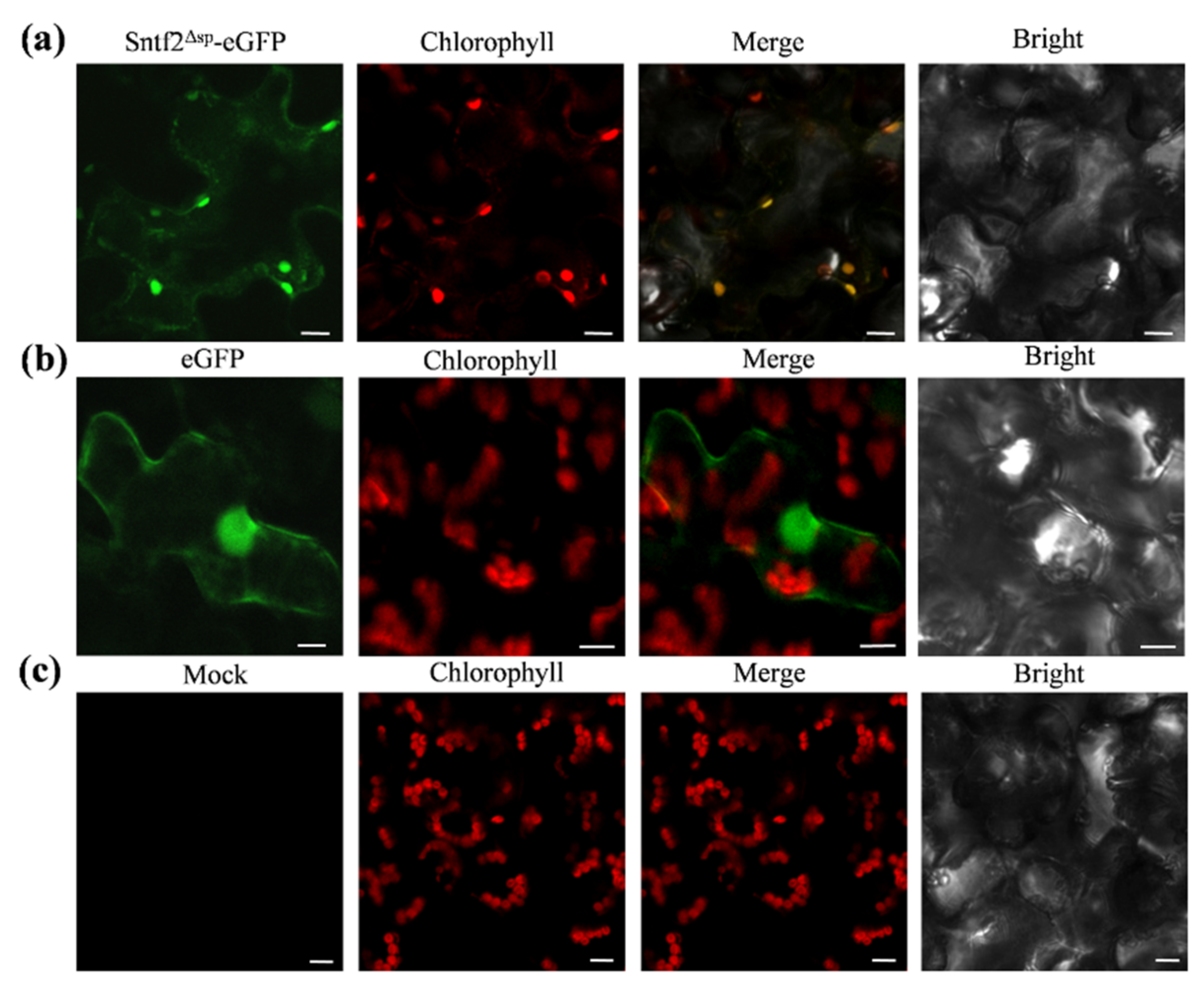
2.6. Sntf2 Is Localized to Plant Chloroplasts
To observe the subcellular localization of Sntf2, we transiently expressed the Sntf2Δsp-eGFP fusion protein using A. tumefaciens LBA4404 infiltration in N. benthamiana. We found that the fluorescence signal of Sntf2Δsp-eGFP fusion proteins overlapped with the chloroplast autofluorescence signal (Figure 6). The localization analysis showed that the Sntf2Δsp-eGFP was mainly presented in chloroplasts.
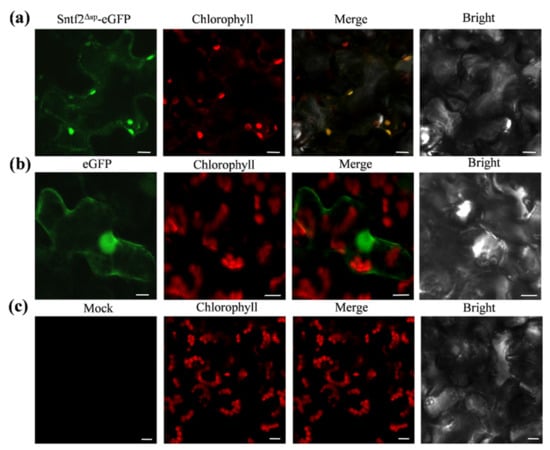
Figure 6.
Sntf2Δsp-eGFP was localized to chloroplasts in Nicotiana benthamiana. (a) Sntf2Δsp-eGFP fluorescence signal overlapped with the chloroplasts autofluorescence signal. (b) The GFP protein was used as a control. (c) Mock. Bar: 10 μm.
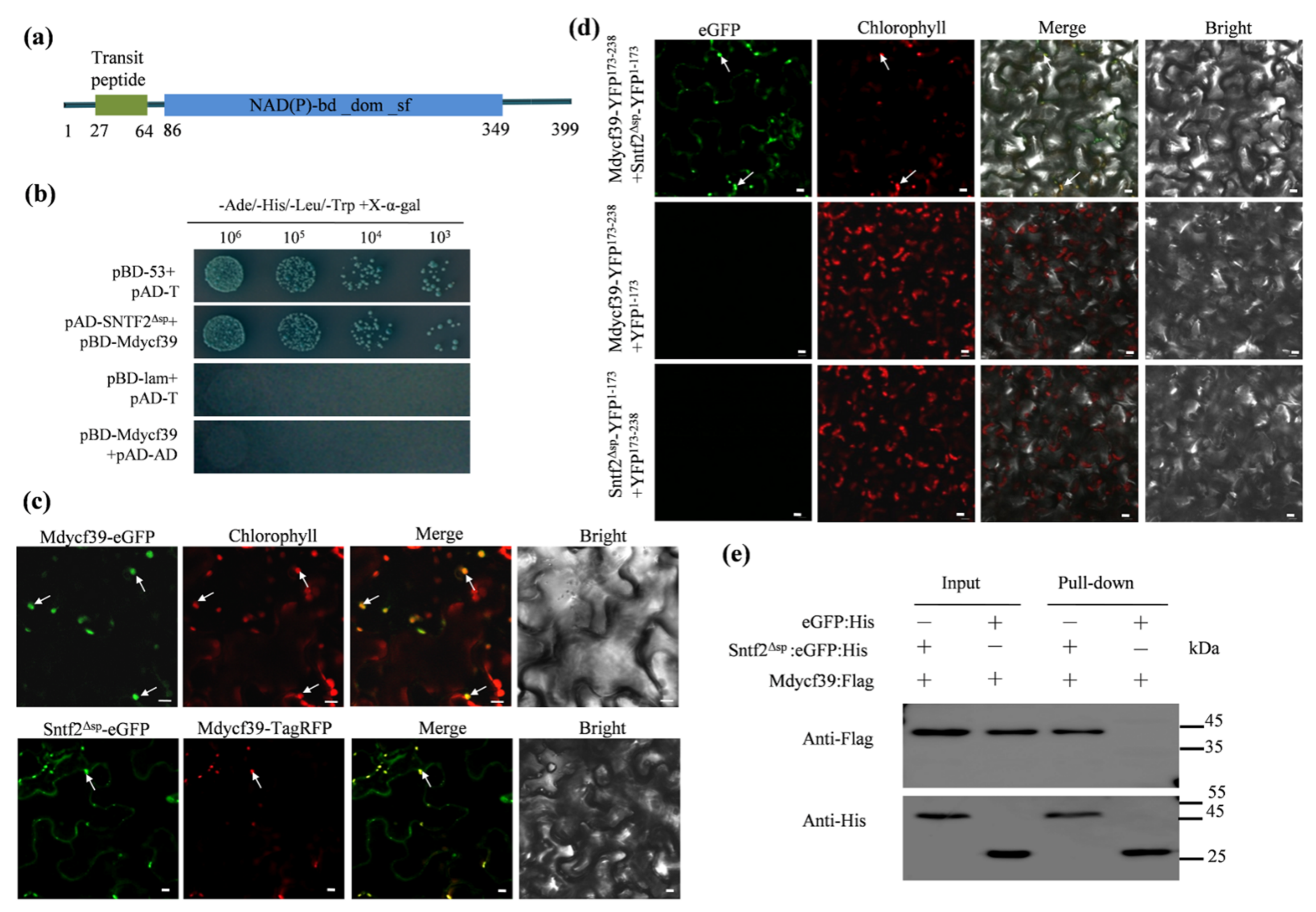
2.7. Sntf2 Interacts with the Photosystem II Assembly FactorMdycf39
To investigate Sntf2-interacting proteins, we performed a yeast two-hybrid (Y2H) screening assay. Twelve distinct putative interactors were confirmed via re-transformation into yeast (Table S1 and Figure S6). One of twelve positive genes encoded a chloroplast photosystem II assembly factor (MD05G1131800) (Table S1). We designated the gene as Mdycf39 since it was annotated as an ycf39-like protein in Malus domestica (NCBI databases). Mdycf39 is highly conserved with other ycf39-like proteins identified from plant species including Arabidopsis thaliana [32](80.76% identity to HCF244; Figure S7a). The Mdycf39 contained a predicted chloroplast-targeting sequence (cTP; 27–64 aa) and a NAD(P)-binding domain (Figure 7a).
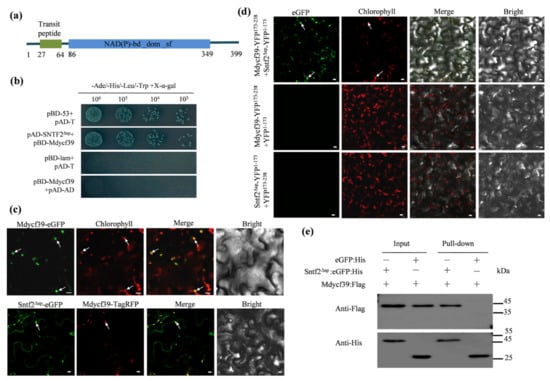
Figure 7.
Sntf2 interacted with Mdycf39 in chloroplasts. (a) Mdycf39 was predicted to contain a chloroplast-targeting transit peptide and a NAD(P)-binding domain by the LOCALIZER program and InterPro 85.0. (b) Confirmation of the Sntf2–Mdycf39 interaction using yeast two-hybrid assays. (c,d) Detection of the Sntf2–Mdycf39 interaction using co-localization and bimolecular fluorescence complementation assays. The arrows refer to the chloroplasts. Bar: 10 μm. (e) Confirmation of Sntf2–Mdycf39 interaction using pull-down assays. Western blots for the analysis of Sntf2:eGFP:His and Mdycf39:Flag purified proteins from Escherichia coli M15 and proteins from the His purification column using anti-His and anti-Flag antibodies. The Sntf2Δsp:eGFP:His and Mdycf39:Flag bands were 46 and 45 kDa, respectively. The protein marker is labeled on the right.
The yeast two-hybrid assay revealed that Mdycf39 interacted with Sntf2ΔSP (lacking SP of Sntf2) (Figure 7b). We then transiently co-expressed the fuse proteins Sntf2Δsp-eGFP and Mdycf39-TagRFP in N. benthamiana. The Mdycf39-eGFP signal was detected in the chloroplasts, and the Mdycf39-TagRFP signal was co-localized with the signal of Sntf2Δsp-eGFP (Figure 7c). The bimolecular fluorescence complementation (BiFC) assay revealed that the fluorescence signal of the interaction between Sntf2-YFP1−173 and Mdycf39-YFP173−238 was present in chloroplasts (Figure 7d). The pull-down analysis also revealed that Sntf2ΔSP interacted with Mdycf39 (Figure 7e). These results showed that Sntf2 interacted with the PS II assembly factor Mdycf39 in chloroplasts.
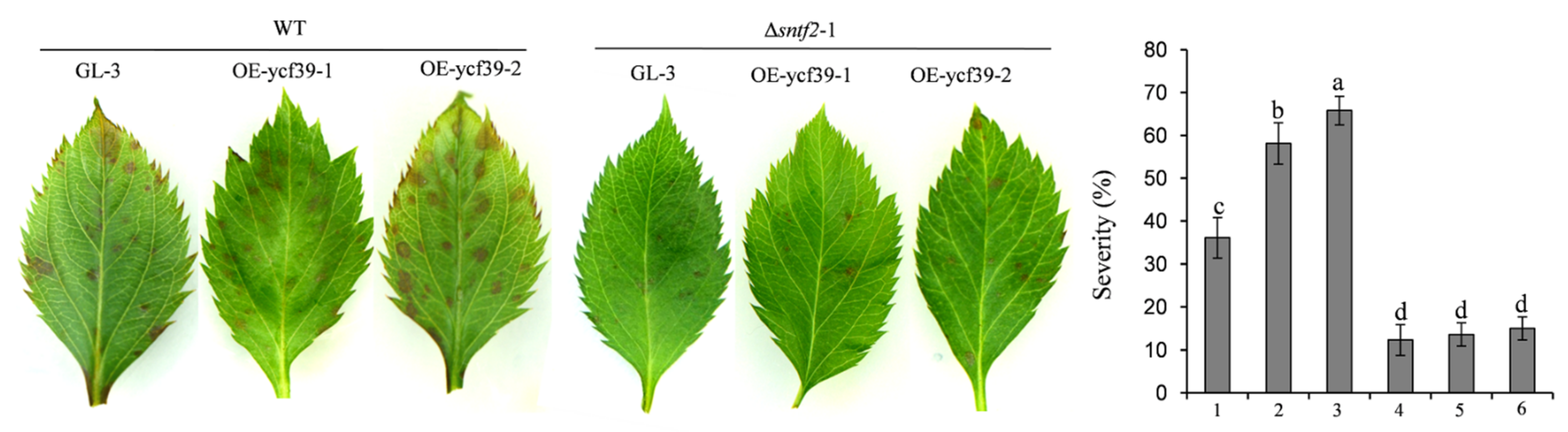
2.8. Mdycf39 Overexpression Increases Susceptibility to C. gloeosporioides in Apple
To study the function of Mdycf39 in plant infection by targeting Sntf2, we generated two Mdycf39 overexpression transgenic lines (OE-ycf39-1 and OE-ycf39-2) and a Mdycf39 transgenic silent line (Ri-ycf39) (Figure S7b,c). The OE-ycf39 lines showed no difference in plant growth compared with GL-3 (cv. Gala, a line) (Figure S7b), but they showed increased susceptibility to C. gloeosporioides compared with GL-3 (Figure S7d). However, the disease severity of the OE-ycf39 lines was similar to that of GL-3 when inoculated with Δsntf2-1 (Figure 8). The result indicated that Sntf2 may suppress apple defenses responses by targeting Mdycf39. Unfortunately, the Ri-ycf39 line did not grow normally with palewhite leaves (Figure S7c), which was consistent with the phenotype of hcf244 mutant in Arabidopsis [32], resulting in the abortion of the Ri-ycf39 line in the pathogenicity test. These results indicated that Mdycf39 played a vital role in plant resistance and development.

Figure 8.
Analysis of the susceptibility of OE-ycf39 lines. The susceptibility was evaluated at three days after inoculation with WT and Δsntf2-1 based on the disease index. 1–3: GL-3, OE-ycf39-1, and OE-ycf39-2 lines were inoculated with WT; 4–5: GL-3, OE-ycf39-1, and OE-ycf39-2 lines were inoculated with Δsntf2-1. Three replicates were performed for each experiment, with six leaves for each replicate. Error bars represent standard deviations. Lowercase letters represent significant differences (p < 0.01).
3. Discussion
The effector plays a vital role in manipulating the host’s immune responses and contributing to plant colonization by the pathogen [8,13,15]. In this study, we identified a novel effector Sntf2, which was required for the pathogenicity of C. gloeosporioides. SNTF2 was highly expressed during the biotrophic infection phase. Effectors were induced to express and secreted to plant cells at different infection phases of Colletotrichum [33,34]. During the early biotrophic infection phase, effectors secreted to the plant cells interfered with immunity of the host, promoting the infection of the pathogen [34,35,36]. Indeed, in this study, we found that Sntf2 inhibited BAX-induced cell death, that the deletion of SNTF2 induced callose deposition, and that H2O2 accumulation increased in infected leaves. These results indicated that Sntf2 played a role in suppressing plant defense and promoting biotrophic infection.
In this study, we found that Sntf2 interacted with Mdycf39 (a PSII assembly factor of Malus domestica) in chloroplasts. Mdycf39 is highly homologous to HCF244, a conserved PSII assembly factor in plants [32]. Mdycf39 and HCF244 are homologous to Ycf39 of Synechocystis [37], which was demonstrated to participate in synthesis of the D1 (a subunit of PSII reaction centre) and assembly of the PSII in chloroplasts [32,37]. The synthesis of the D1 was necessary to maintain the structure and function of the PSII [38]. These suggested that Mdycf39 may also be involved in the assembly of the PSII in chloroplasts of apples. In this study, the apple Mdycf39-RNAi line did not grow normally on MS, consistent with the phenotype of hcf244 mutant in Arabidopsis [32]. These results indicated that Mdycf39 may affect the stability of PSII and play a role in plant growth and development.
Different suborganelles in the host often become targets of pathogens [23,39]. Many effectors target different components of the PSII in the host, which interferes with electron transport and inhibits ROS production, interfering with plant defense [6,28,40]. The effector RXLR31154 of Plasmopara viticola targets PsbP (a factor of the oxygen-evolving complex of PSII) to inhibit H2O2 production in a grapevine and enhance its susceptibility [6]. In our study, we found that the effector Sntf2 of C. gloeosporioides migrated into the host’s chloroplasts and interacted with Mdycf39. The transgenic overexpression lines of Mdycf39 in GL-3 showed an increased susceptibility to C. gloeosporioides. Our study indicated that Mdycf39 plays an important role in plant photosynthesis and defense responses. Mdycf39 may affect the stability of PSII and the production of H2O2 in chloroplasts.The defense response of plants is usually associated with ROS generation and callose depositions [41]. ROS has a direct anti-microbial role and is also a retrograde signaling molecule entering the nucleus to regulate the expression of defense-related genes, resulting in hypersensitive cell death [42,43,44]. In this study, the deletion of SNTF2 increased the accumulation of H2O2 and the deposition of callose during infection. We speculated that Sntf2 perturbs the function of chloroplast by targeting Mdycf39, inhibiting the apple’s defense response.
In conclusion, we identified a novel effector Sntf2 of C. gloeosporioides. Sntf2 is required for pathogenicity and plays a vital role in plant infection. The deletion of SNTF2 triggers plant defense responses. Sntf2 secreted into the host cells was located in the chloroplast. Sntf2 interacted with Mdycf39 (a photosystem II assembly factor) in chloroplasts. We demonstrated that Sntf2 perturbs the function of chloroplasts by targeting Mdycf39, avoiding hypersensitive cell death, and supporting the colonization of C. gloeosporioides in apple leaves.
4. Materials and Methods
4.1. Strains and Plant Materials
The C. gloeosporioides strain W16 was used as the wild type (WT) strain [19]. The fungal strains were cultured on PDA plates at 26 °C as described previously [19]. For transformant selection, G-418 sulphate or hygromycin B was added to PDA infinal concentrations of 500 μg/mL or 100 μg/mL, respectively [45]. The tissue-cultured ‘GL-3′ (cultivar ‘Royal Gala’) plants [46] and the transgenic plants were cultivated in MS medium [47] in a climate-controlled culture room at 25 ± 1 °C with a 16/8 h light/dark photoperiod as described by Dai et al. [46]. N. benthamiana seedlings were cultured in a greenhouse at 22–25 °C. The healthy leaves were obtained from 2-year-old seedlings of the ‘Golden Delicious’ variety of apples (Malus domestica) (Institute of Pomology of Chinese Academy of Agricultural Sciences, CAAS, Xingcheng, Liaoning Province, China).
4.2. Agrobacterium tumefaciens Infiltration Assays
The coding sequence of SNTF2 (CGGC5_13909) was amplified and ligated into the potato virus X (PVX) vector pGR106 to generate pGR106-SNTF2. The primers used in this assay are listed in Table S2. The recombinant vector was transformed into A. tumefaciens GV3101. For transient expression in N. benthamiana, the transformant strains of GV3101 were infiltrated into the leaves. The experiment was performed as described by Shang et al. [15].
4.3. Vector Construction and Fungal Transformation
The gene deletion construction and transformation of C. gloeosporioides were carried out using the protocols described previously [45]. The gene complementation construction and the fungal transformation were performed as described previously [48]. The primers used in this assay are listed in Table S2. A detailed description is shown in Table S2. The putative gene knockout mutants were identified by PCR and Southern blot analysis. The complementation strains were confirmed based on PCR analysis.
4.4. Phenotype Assays
The hyphal growth rate and conidial production were assessed according to the method described by Zhou et al. [45]. The formation rates of appressorium and invasive pegs were observed and calculated as described by Shang et al. [15]. For plant inoculation, fresh conidial suspensions (1 × 105 conidia/mL) were sprayed onto the apple leaves according to the method described by Zhou et al. [45]. The inoculated apple seedlings (2-year-oldseedlings of the ‘Golden Delicious’ variety as shown in Section 4.1) were cultured at 28 °C and 75% humidity. For the pathogenicity test, the disease lesions were examined 3 days post-inoculation. The severity of the GLSA on each leaf was estimated using a diagrammatic scale [49].
4.5. RNA Extraction and qRT-PCR Analysis
The total RNA was extracted using an RNAprep Pure Plant Kit (TianGen Biotech, China Beijing). For the qRT-PCR analysis of SNTF2, the RNA samples of the infestation phase were obtained from ‘Golden Delicious’ leaves (2-year-old seedlings of the ‘Golden Delicious’ variety, inoculated as shown in Section 4.4) at 12, 24, 48, and 72 hpi. For the qRT-PCR analysis of Mdycf39, the apple RNA samples were extracted from the transgenic overexpression lines of Mdycf39 and tissue-cultured ‘GL-3′ plants. The qRT-PCR was performedas described by Tan et al. [17]. The M. domestica ubiquitin extension (MdUBQ) was used as the endogenous reference gene [15].
4.6. Signal Peptide Activity Assay
The SP sequence of SNTF2 was amplified and ligated into pSUC2 vector to generate pSUC2-SPSNTF2. Recombinant vectors were transformed into the yeast strain YTK12, which lacks a secreted invertase [50]. All transformants were cultured on YPDA plates at 30 °C and cultured on CMD-W and YPRAA plates to assess if the invertase secreted. The enzyme activity of the invertase was evaluated based on the reduction of TTC to the insoluble red compound 1, 3, 5-triphenylformazan. The experiment was performed as described by Xu et al. [28].
4.7. Histochemical Assays
The H2O2 accumulation in plants was assessed using DAB staining as described by Chen et al. [51]. The samples were from ‘GL-3′ and transgenic leaves inoculated with the WT, Δsntf2-1, and Δsntf2-1/SNTF2 strains. DAB oxidation leads to brownish polymer formation and deposition at the site of ROS accumulation. The callose deposition was observed using aniline blue staining. The samples were decolourized by boiling in 96% ethanol for 5 min and then immersed in chloral hydrate overnight. The transparent leaf segments were stained with 0.05% aniline blue in 0.067 M K2HPO4 (pH 9.6) [28]. Then processed samples were preserved in 30% glycerol for microscopic analysis (Leica DM5000 B, Leica, Wetzlar, Germany). For every time point, either ten leaf discs were processed using DAB staining or aniline blue staining was obtained for ten leaf discs, and every experiment was performed three times. For every leaf disc, 100 appressoria were observed, and the percentage of ROS accumulation or callose deposition was calculated along with the means and standard deviations.
4.8. Yeast Two-Hybrid Assay
The coding sequences of SNTF2 (without SP) were cloned into pGBKT7-BD as the bait, and the Matchmaker GAL4 system (OE Biotech, Shanghai, China) was used to screen a cDNA library constructed from RNA and isolated from different infection phases of the ‘Golden Delicious’ apple leaves. The screening was performed according to the manufacturer’s instructions (OE Biotech, Shanghai, China). To confirm the interaction, the prey vector pAD-Sntf2Δsp and bait vector pBD-Mdycf39 (constructed using Mdycf39 from ‘Golden Delicious’) were co-transformed into Y2Hgold yeast strains. The transformed yeast strains were grown on a medium (SD/-Leu/-Trp and SD/-Leu/-Trp/-His/-Ade) at 30 °C.
4.9. Transient Expression Analysis in N. benthamiana
For the subcellular localization assay, SNTF2 (without SP) and Mdycf39 were cloned into pGR35s-eGFP or pGR35s-TagRFP and transformed into A. tumefaciens LBA4404 to express Sntf2Δsp-eGFP, Mdycf39-eGFP, and Mdycf39-TagRFP fusion proteins, respectively. The transient expression in N. benthamiana was performed as described by Xu et al. [28]. The N. benthamiana leaves were observed using a confocal microscope (Leica TCS SP8, Leica, Wetzlar, Germany) after 2 days of infiltration. For the BiFC assay, SNTF2 and Mdycf39 were ligated into the vectors pGR35s-YFP1−173 and pGR35s-YFP173−238. The Mdycf39-YFP173−238 and Sntf2Δsp-YFP1−173 fusion proteins were co-expressed by A. tumefaciens infiltration in N. benthamiana. The experiment was performed as described by Xu et al. [28]. GFP fluorescence was excited using a 488 nm laser, and emission was collected between 505 and 535 nm. YFP was excited using a 514 nm laser, and emission was collected between 530 and 560 nm. TagRFP was excited using a 555 nm laser, and emission was collected between 578 and 610 nm. For autofluorescent chloroplast detection, the excitation was 630 nm, and the collection range of emitted light was set at 650–681 nm [52].
4.10. Protein Extraction and Immunoblotting
For the pull-down assay, SNTF2 and Mdycf39 were cloned into pQE30-eGFP and pQE30-Flag and transformed into Escherichia coli M15 cells. Protein extraction was performed according to Dominguez-Martin et al. [53]. The induced proteins were observed using SDS-PAGE and Coomassie Brilliant Blue staining. The purified proteins were co-incubated on ice for 3 h and were purified using Ni-NTA beads (CW0894S, CWBIO, Beijing, China). For immune detection, the purified proteins were transferred to a polyvinylidene fluoride membrane. The corresponding mouse anti-His or mouse anti-Flag (1:1000; cat. no. HT201, TransGenBiotech, Beijing, China) antibody was used as the primary antibody, respectively. The HRP-labelled goat anti-mouse IgG (1:2000; cat. no. A0216, Beyotime, Shanghai, China) was used as the secondary antibody. The membrane was treated with a BeyoECL Star kit (Beyotime, Shanghai, China) for 2 min. Images were acquired using a BIO-RAD ChemiDoc™ Imaging System.
4.11. Generation of Transgenic ‘GL-3’Plants with Mdycf39 Overexpression or RNA-Interference
Mdycf39 was amplified and inserted into the pRNAi and pRPHA vectors to generate the RNAi transgenic silenced line Ri-ycf39 as well as the transgenic overexpression lines OE-ycf39. The Agrobacterium-mediated transformation of ‘GL-3’ was performed as described by Dai et al. [46]. A. tumefaciens LBA4404 was used for the stable transformations.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ijms23126379/s1.
Author Contributions
Conceptualization, Z.Z.; methodology, M.W., J.X., X.Z. and Z.Z.; software, M.W., J.X. and Z.J.; validation, M.W., Z.J. and H.Y.; formal analysis, M.W. and Z.J.; investigation, M.W., H.Y. and X.Z.; resources, Z.Z. and Z.J.; data curation, M.W.; writing—original draft preparation, M.W.; writing—review and editing, Z.Z.; visualization, M.W.; supervision, Z.Z. and Z.J.; project administration, Z.Z.; funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by The Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-2016-RIP), China Agriculture Research System of MOF and MARA (CARS-27), and Special Funds for Basic Scientific Research Operation of Central-level Public Welfare Scientific Research Institutes (1610182019030).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Acknowledgments
The authors would like to thank Zhang Zhi-hong, (Shenyang Agricultural University), for providing tissue-cultured ‘GL-3’ (cultivar ‘Royal Gala’) plants. Thank you, Dou Dao-long, (College of Plant Protection, Nanjing Agricultural University), for the pGR106 vector.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marco, Z.; Massimiliano, C.; Antonio, K.M.; Antonielle, M.; Silvia, M.; Irma, M.; Sylvie, J.; de Ortiz, G.M.; Christian, H.; Mathilde, F.; et al. LPMO-oxidized cellulose oligosaccharides evoke immunity in Arabidopsis conferring resistance towards necrotrophic fungus B. cinerea. Commun. Biol. 2021, 4, 4224. [Google Scholar]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant Cell Wall–Degrading Enzymes and Their Secretion in Plant-Pathogenic Fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Dou, D.; Zhou, J.M. Phytopathogen effectors subverting host immunity: Different foes, similar battleground. Cell Host Microbe 2012, 12, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Chen, T.; Yin, X.; Xiang, G.; Peng, J.; Fu, Q.; Xu, Y. A Plasmopara viticola RxLR effector targets a chloroplast protein PsbP to inhibit ROS production in grapevine. Plant J. 2021, 106, 1557–1570. [Google Scholar] [CrossRef]
- Perfect, S.E.; Hughes, H.B.; O’Connell, R.J.; Green, J.R. Colletotrichum: A Model Genus for Studies on Pathology and Fungal–Plant Interactions. Fungal Genet. Biol. 1999, 27, 186–198. [Google Scholar] [CrossRef]
- Bhadauria, V.; Banniza, S.; Vandenberg, A.; Selvaraj, G.; Wei, Y. Overexpression of a novel biotrophy-specific Colletotrichum truncatum effector, CtNUDIX, in hemibiotrophic fungal phytopathogens causes incompatibility with their host plants. Eukaryot. Cell 2013, 12, 2–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshino, K.; Irieda, H.; Sugimoto, F.; Yoshioka, H.; Okuno, T.; Takano, Y. Cell death of Nicotiana benthamiana is induced by secreted protein NIS1 of Colletotrichum orbiculare and is suppressed by a homologue of CgDN3. Mol. Plant-Microbe Interact. MPMI 2012, 25, 625. [Google Scholar] [CrossRef] [Green Version]
- Takahara, H.; Hacquard, S.; Kombrink, A.; Hughes, H.B.; Halder, V.; Robin, G.P.; Hiruma, K.; Neumann, U.; Shinya, T.; Kombrink, E. Colletotrichum higginsianum extracellular LysM proteins play dual roles in appressorial function and suppression of chitin-triggered plant immunity. New Phytol. 2016, 211, 1323–1337. [Google Scholar] [CrossRef] [Green Version]
- Takahara, H.; Yamaguchi, S.; Omura, N.; Nakajima, S.; Otoku, K.; Tanaka, S.; Ogura, K.; Kleemann, J.; O’Connell, R. The Colletotrichum higginsianum secreted effector protein ChEC91 induces plant cell death. J. Gen. Plant Pathol. 2021, 87, 344–353. [Google Scholar] [CrossRef]
- Ayako, T.; Mari, N.; Pamela, G.; Naoyoshi, K.; Ryoko, H.; Naoki, K.; Shunji, T.; Yoshitaka, T.; Yoshihiro, N.; Ken, S. The conserved Colletotrichum spp. effector candidate CEC3 induces nuclear expansion and cell death in plants. Front. Microbiol. 2021, 12, 682155. [Google Scholar]
- Kleemann, J.; Rinconrivera, L.J.; Takahara, H.; Neumann, U.; van Themaat, E.V.L.; Hc, V.D.D.; Hacquard, S.; Stüber, K.; Will, I.; Schmalenbach, W. Correction: Sequential delivery of host-induced virulence effectors by appressoria and intracellular hyphae of the phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Irieda, H.; Maeda, H.; Akiyama, K.; Hagiwara, A.; Saitoh, H.; Uemura, A.; Terauchi, R.; Takano, Y. Colletotrichum orbiculare secretes virulence effectors to a biotrophic interface at the primary hyphal neck via exocytosis coupled with SEC22-mediated traffic. Plant Cell 2014, 26, 2265–2281. [Google Scholar] [CrossRef] [Green Version]
- Shang, S.; Wang, B.; Zhang, S.; Liu, G.; Liang, X.; Zhang, R.; Gleason, M.L.; Sun, G. A novel effector CfEC92 of Colletotrichum fructicola contributes to glomerella leaf spot virulence by suppressing plant defences at the early infection phase. Mol. Plant Pathol. 2020, 21, 936–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouch, J.A.; O’Connell, R.; Gan, P.; Buiate, E.; Torres, M.F.; Beirn, L.; Shirasu, K.; Vaillancourt, L. The Genomics of Colletotrichum. In Genomics of Plant-Associated Fungi: Monocot Pathogens; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Tan, Q.; Zhao, X.; He, H.; Zhang, J.; Yi, T. Carbamoyl phosphate synthetase subunit Cpa1 interacting with Dut1, controls development, arginine biosynthesis, and pathogenicity of Colletotrichum gloeosporioides. Fungal Biol. 2020, 125, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Yakoby, N.; Beno-Moualem, D.; Keen, N.T.; Dinoor, A.; Pines, O.; Prusky, D. Colletotrichum gloeosporioides pelB Is an Important Virulence Factor in Avocado Fruit-Fungus Interaction. Mol Plant Microbe Interact 2001, 14, 988–995. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ji, Z.; Wang, N.; Chi, F.; Xu, C.; Zhou, Z.; Zhang, J. Identification of conidiogenesis-associated genes in Colletotrichum gloeosporioides by agrobacterium tumefaciens-mediated transformation. Curr. Microbiol. 2016, 73, 802–810. [Google Scholar] [CrossRef]
- Gonzalez, E.; Sutton, T.B. First report of Glomerella leaf spot (Glomerella cingulata) of apple in the United States. Plant Dis. 1999, 83, 1074. [Google Scholar] [CrossRef]
- Wang, C.X.; Zhang, Z.F.; Li, B.H.; Wang, H.Y.; Dong, X.L. First report of Glomerella Leaf spot of apple caused by Glomerella cingulata in China. Plant Dis. 2012, 96, 912. [Google Scholar] [CrossRef]
- González, E.; Sutton, T.B.; Correll, J.C. Clarification of the Etiology of Glomerella Leaf Spot and Bitter Rot of Apple Caused by Colletotrichum spp. Based on Morphology and Genetic, Molecular, and Pathogenicity Tests. Phytopathology 2006, 96, 982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Torres Zabala, M.; Littlejohn, G.; Jayaraman, S.; Studholme, D.; Bailey, T.; Lawson, T.; Tillich, M.; Licht, D.; Bölter, B.; Delfino, L.; et al. Chloroplasts play a central role in plant defence and are targeted by pathogen effectors. Nat. Plants 2015, 1, 15074. [Google Scholar] [CrossRef]
- Chan, K.X.; Phua, S.Y.; Crisp, P.; McQuinn, R.; Pogson, B.J. Learning the languages of the chloroplast: Retrograde signaling and beyond. Annu. Rev. Plant Biol. 2016, 67, 25–53. [Google Scholar] [CrossRef]
- Medina-Puche, L.; Tan, H.; Dogra, V.; Wu, M.; Lozano-Duran, R. A defense pathway linking plasma membrane and chloroplasts and co-opted by pathogens. Cell 2020, 182, 1109–1124. [Google Scholar] [CrossRef]
- Serrano, I.; Audran, C.; Rivas, S. Chloroplasts at work during plant innate immunity. J. Exp. Bot. 2016, 67, 3845–3854. [Google Scholar] [CrossRef] [PubMed]
- Nomura, H.; Komori, T.; Uemura, S.; Kanda, Y.; Shimotani, K.; Nakai, K.; Furuichi, T.; Takebayashi, K.; Sugimoto, T.; Sano, S.; et al. Chloroplast-mediated activation of plant immune signalling in Arabidopsis. Nat. Commun. 2012, 3, 926. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Diaz, T.; Zhang, D.; Fan, P.; Wang, L.; Ding, X.; Jiang, Y.; Jimenez-Gon-gora, T.; Medina-Puche, L.; Zhao, X.; Feng, Z. A virus-targeted plant receptor-like kinase promotes cell-to-cell spread of RNAi. Proc. Natl. Acad. Sci. USA 2018, 115, 1388–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacomme, C.; Santa, C.S. Bax-induced cell death in tobacco is similar to the hypersensitive response. Proc. Natl. Acad. Sci. USA 1999, 96, 7956–7961. [Google Scholar] [CrossRef] [Green Version]
- Weixiao, Y.; Yufu, W.; Tao, C.; Yang, L.; Chaoxi, L. Functional Evaluation of the Signal Peptides of Secreted Proteins. Bio-Protoc. 2018, 8, e2839. [Google Scholar]
- Link, S.; Engelmann, K.; Meierhoff, K.; Westhoff, P. The atypical short-chain dehydrogenases HCF173 and HCF244 are jointly involved in translational initiation of the psbA mRNA of Arabidopsis. Plant Physiol. 2012, 160, 2202–2218. [Google Scholar] [CrossRef] [Green Version]
- De Queiroz, C.B.; Correia, H.L.N.; Santana, M.F. The repertoire of effector candidates in Colletotrichum lindemuthianum reveals important information about Colletotrichum genus lifestyle. Appl. Microbiol. Biotechnol. 2019, 103, 2295–2309. [Google Scholar] [CrossRef]
- Gan, P.; Ikeda, K.; Irieda, H.; Narusaka, M.; O’Connell, R.J.; Narusaka, Y.; Takano, Y.; Kubo, Y.; Shirasu, K. Comparative genomic and transcriptomic analyses reveal the hemibiotrophic stage shift of Colletotrichum fungi. New Phytol. 2013, 197, 1236–1249. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Shang, S.; Dong, Q.; Wang, B.; Zhang, R.; Mark, L.G.; Sun, G. Transcriptomic analysis reveals candidate genes regulating development and host interactions of Colletotrichum fructicola. Bmc. Genom. 2018, 19, 557. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Knoppova, J.; Sobotka, R.; Tichy, M.; Yu, J.; Konik, P.; Halada, P.; Nixon, P.J.; Komenda, J. Discovery of a chlorophyll binding protein complex involved in the early steps of photosystem II assembly in Synechocystis. Plant Cell 2014, 26, 1200–1212. [Google Scholar] [CrossRef] [Green Version]
- Andersson, B.; Aro, E.M. Photodamage and D1 Protein Turnover in Photosystem II. Regul. Photosynth. 2001, 11, 377–393. [Google Scholar]
- Robin, G.P.; Kleemann, J.; Neumann, U.; Cabre, L.; Dallery, J.F.; Lapalu, N.; O’Connell, R.J. Subcellular Localization Screening of Colletotrichum higginsianum Effector Candidates Identifies Fungal Proteins Targeted to Plant Peroxisomes, Golgi Bodies, and Microtubules. Front. Plant Sci. 2018, 9, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herva, J.J.R.; Melendi, P.G.; Lanza, R.C.; Lamas, M.A.; Alvarez, I.R.; Li, Z.; Torrejón, G.L.; Díaz, I.; de Pozo, J.C.; Chakravarthy, S.; et al. A bacterial cysteine protease effector protein interferes with photosynthesis to suppress plant innate immune responses. Cell. Microbiol. 2012, 14, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Couto, D.; Zipfel, C. Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef]
- Torres, M.A. ROS in biotic interactions. Physiol. Plant. 2010, 138, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Galvez-Valdivieso, G.; Mullineaux, P.M. The role of reactive oxygen species in signalling from chloroplasts to the nucleus. Physiol Plant 2010, 138, 430–439. [Google Scholar] [CrossRef]
- Quan, L.J.; Zhang, B.; Shi, W.-W.; Li, H.-Y. Hydrogen Peroxide in Plants:a Versatile Molecule of the Reactive Oxygen Species Network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef]
- Zhou, Z.; Wu, J.; Wang, M.; Zhang, J. ABC protein CgABCF2 is required for asexual and sexual development, appressorial formation and plant infection in Colletotrichum gloeosporioides. Microb. Pathog. 2017, 110, 85–92. [Google Scholar] [CrossRef]
- Dai, H.; Li, W.; Han, G.; Yi, Y.; Zhang, Z. Development of a seedling clone with high regeneration capacity and susceptibility to Agrobacterium in apple. Sci. Hortic. 2013, 164, 202–208. [Google Scholar] [CrossRef]
- Srinivasan, C.; Liu, Z.; Scorza, R. Ectopic expression of class 1 KNOX genes induce adventitious shoot regeneration and alter growth and development of tobacco (Nicotiana tabacum L.) and European plum (Prunus domestica L.). Plant Cell Rep. 2011, 30, 655–664. [Google Scholar] [CrossRef]
- Zhao, X.; Tang, B.; Xu, J.; Wang, N.; Zhou, Z.; Zhang, J. A SET domain-containing protein involved in cell wall integrity signaling and peroxisome biogenesis is essential for appressorium formation and pathogenicity of Colletotrichum gloeosporioides. Fungal Genet. Biol. 2020, 145, 103474. [Google Scholar] [CrossRef]
- Bogo, A.; Casa, T.R.; Rufato, L.; Goncalves, J.M. The effect of hail protection nets on Glomerella leaf spot in ‘Royal Gala’ apple. Crop Prot. 2012, 31, 40–44. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H.; Willems, A.R.; Woods, R.A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 1995, 11, 355–360. [Google Scholar] [CrossRef]
- Chen, M.; Zeng, H.; Qiu, D.; Guo, L.; Yang, X.; Shi, H.; Zhou, T.; Zhao, J. Purification and characterization of a novel hypersensitive response-inducing elicitor from Magnaporthe oryzae that triggers defense response in rice. PLoS ONE 2012, 7, e37654. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Wang, W.; Li, H.; Pan, X. Autofluorescence of chloroplasts measured by a laser scanning confocal microscope. J. Tsinghua Univ. (Sci. Technol.) 2017, 57, 651–654. [Google Scholar]
- Dominguez-Martin, M.A.; Polívka, T.; Sutter, M.; Ferlez, B.; Lechno-Yossef, S.; Montgomery, B.L.; Kerfeld, C.A. Structural and spectroscopic characterization of HCP2. Biochim. Biophys. Acta Bioenerg. 2019, 1860, 414–424. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).