
In Vitro Characterization of Sphingosine 1-Phosphate Receptor 1 (S1P1) Expression and Mediated Migration of Primary Human T and B Cells in the Context of Cenerimod, a Novel, Selective S1P1 Receptor Modulator

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
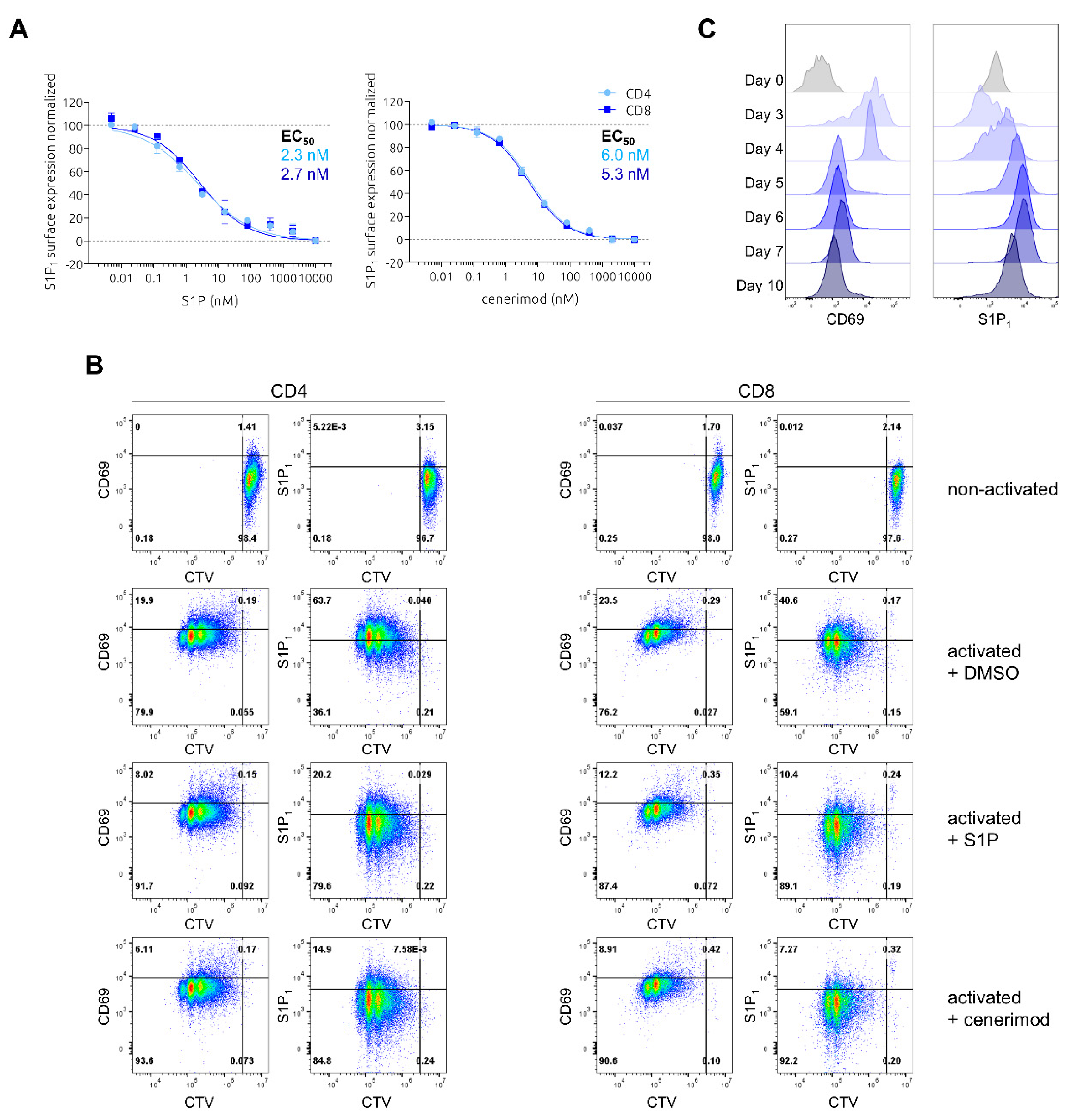
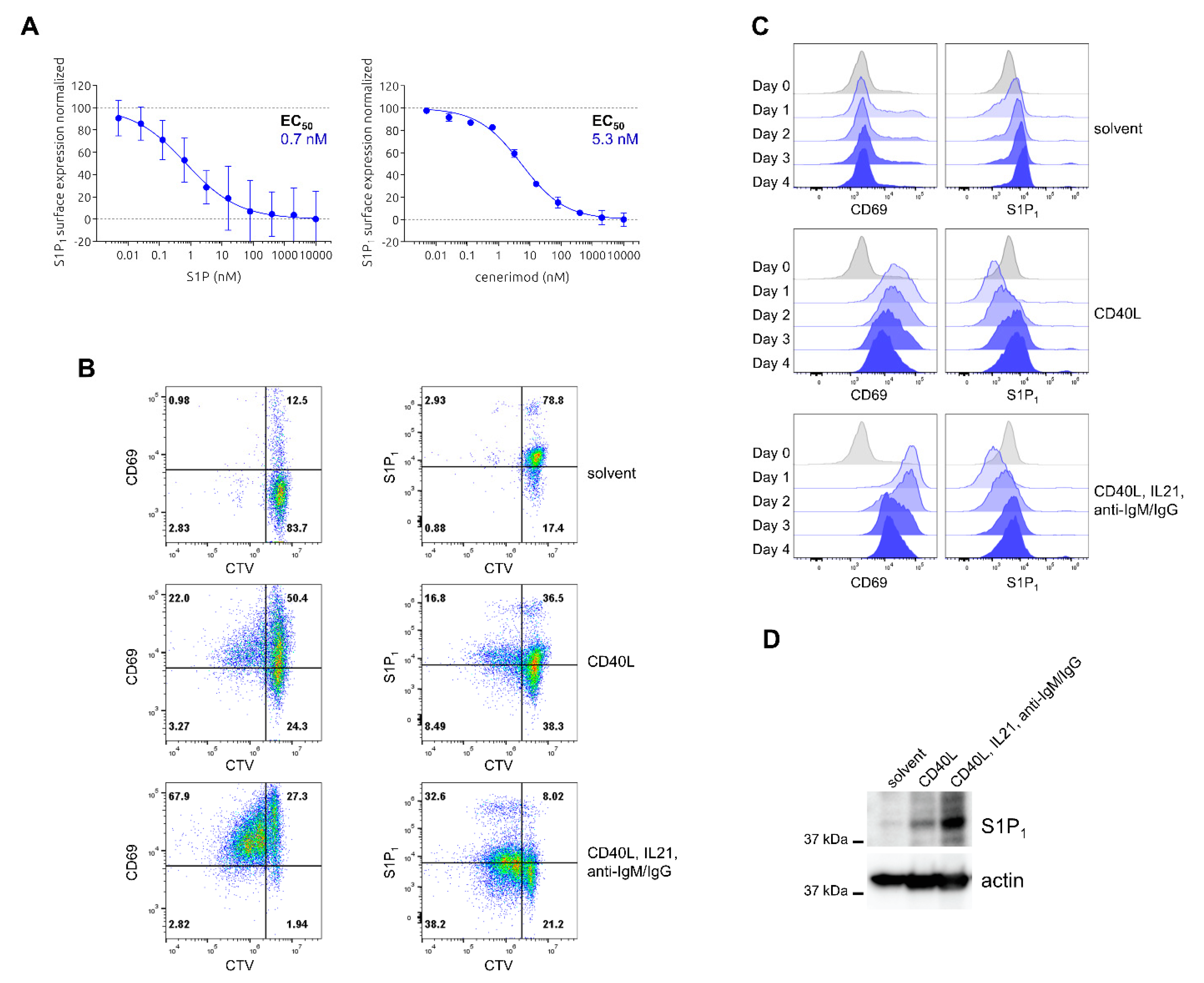
2.1. Characterization of Ligand-Mediated S1P1 Receptor Internalization and Reciprocal Expression of S1P1 Receptor and CD69 upon Activation in Primary Human T Lymphocytes
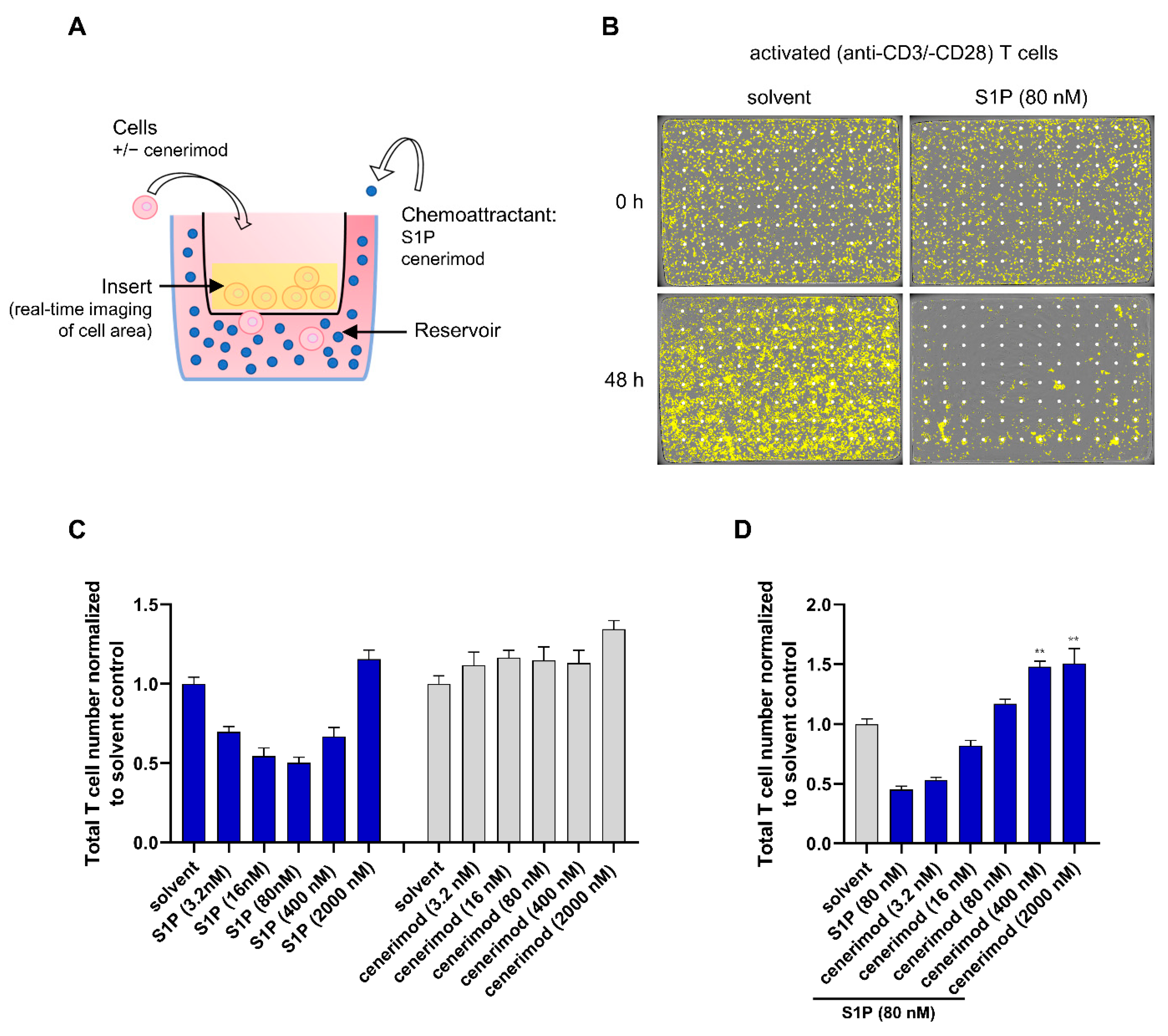
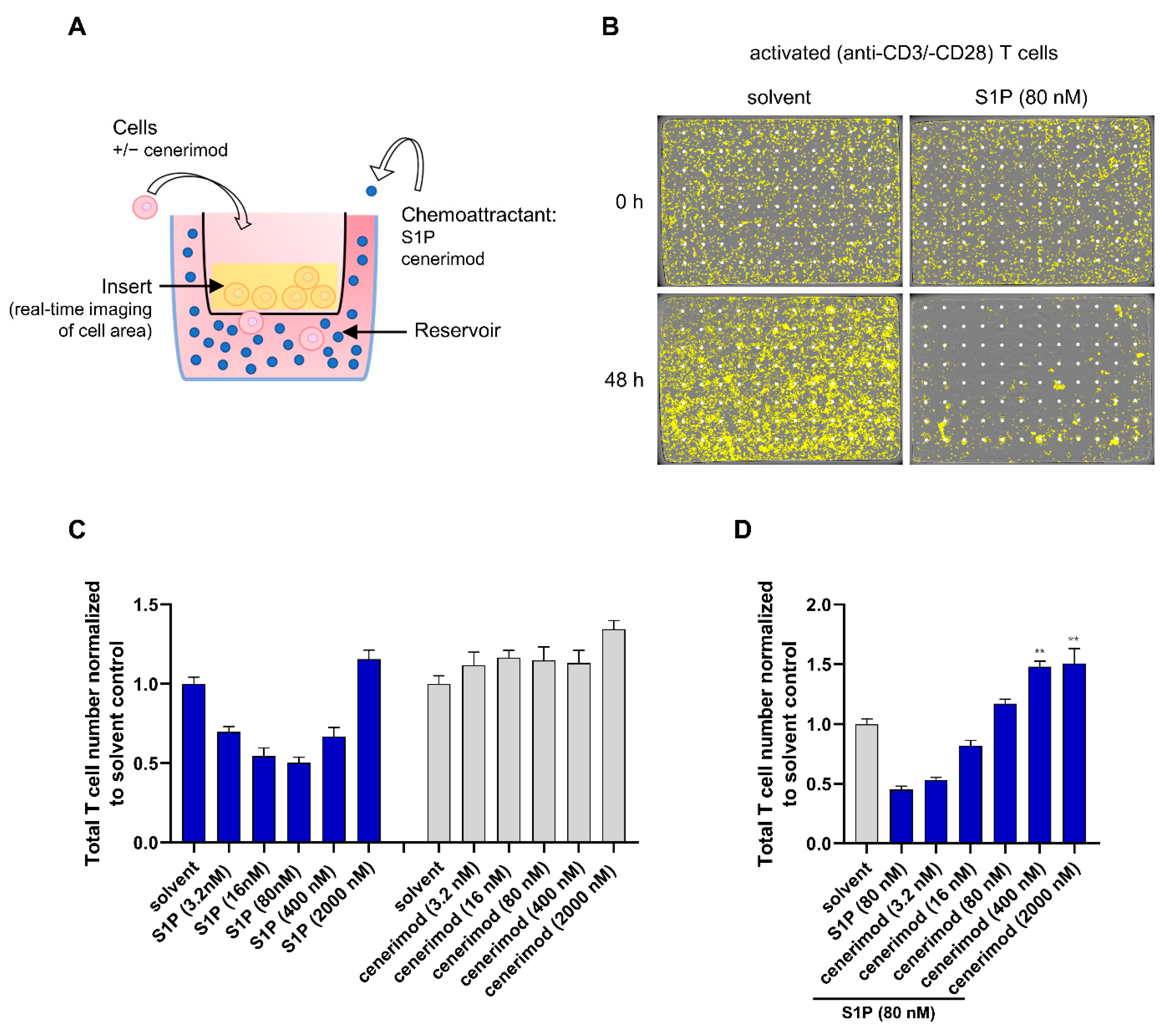
2.2. S1P1 Receptor Modulator Cenerimod Mediates Effective Prevention of T Cell Migration towards S1P
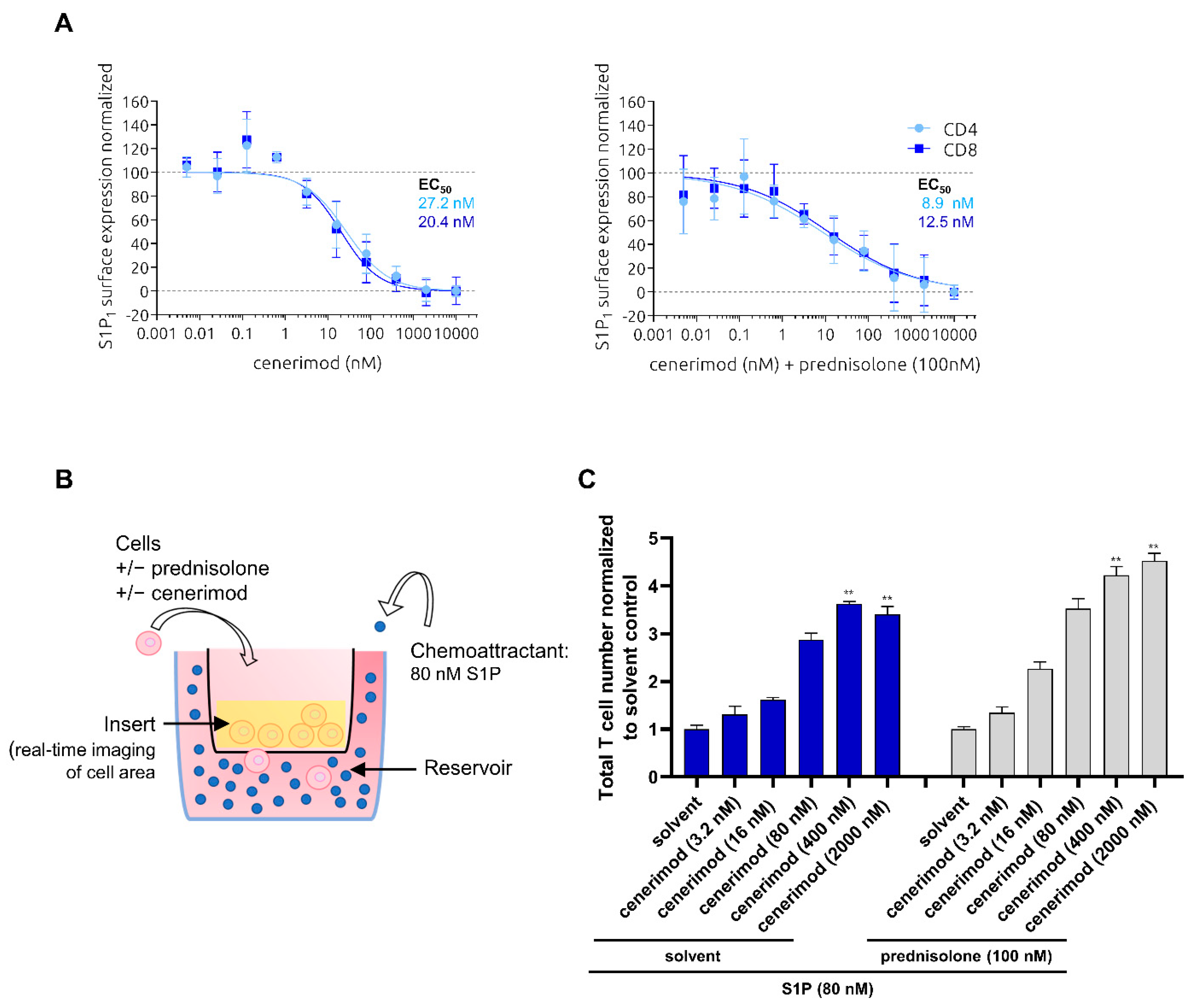
2.3. Co-Treatment with Prednisolone Has No Impact on S1P-Mediated Receptor Internalization and Migration in Primary Human T Lymphocytes
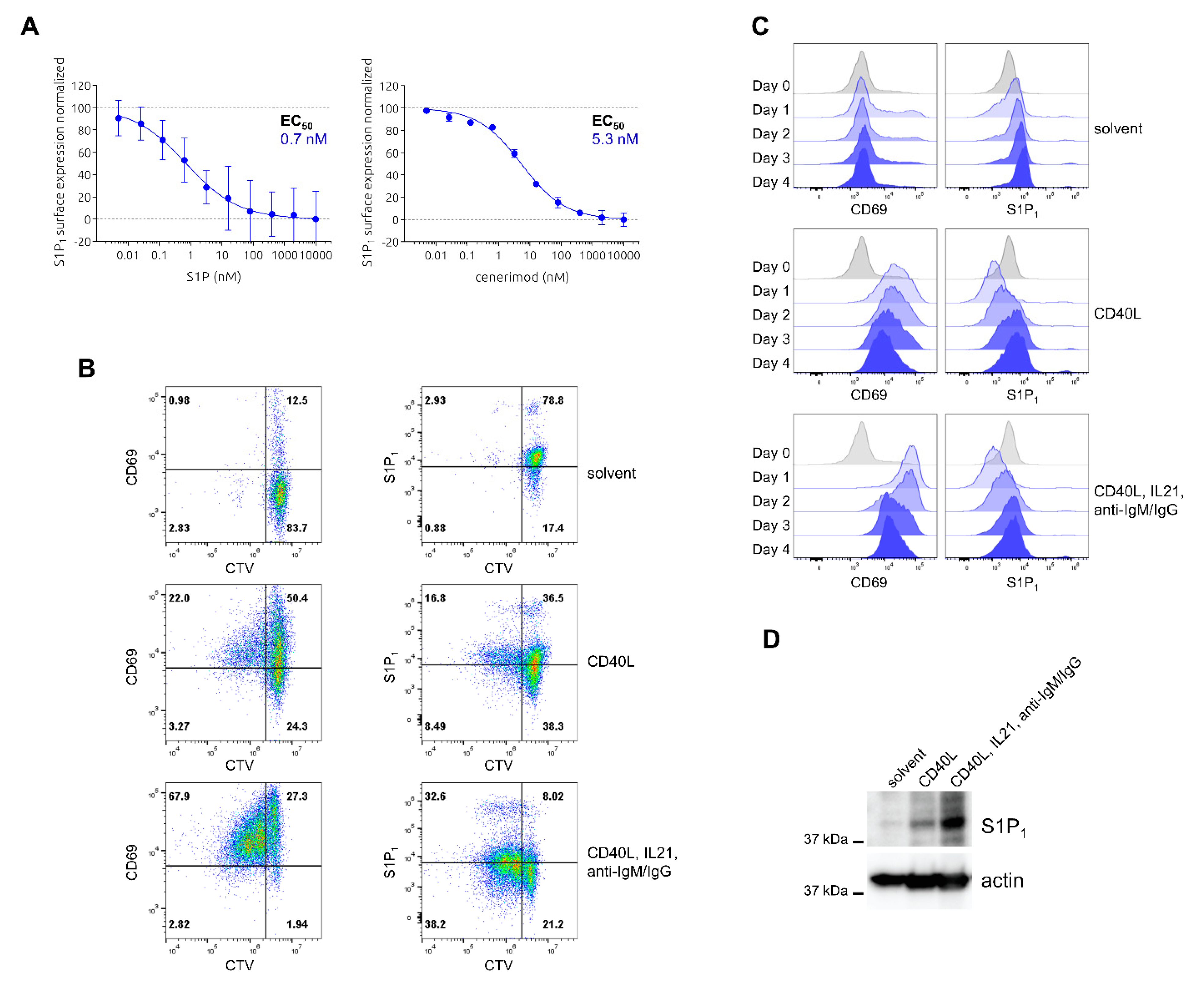
2.4. Characterization of Ligand-Mediated S1P1 Receptor Internalization and Reciprocal Expression of S1P1 Receptor and CD69 upon Activation in Primary Human B Lymphocytes
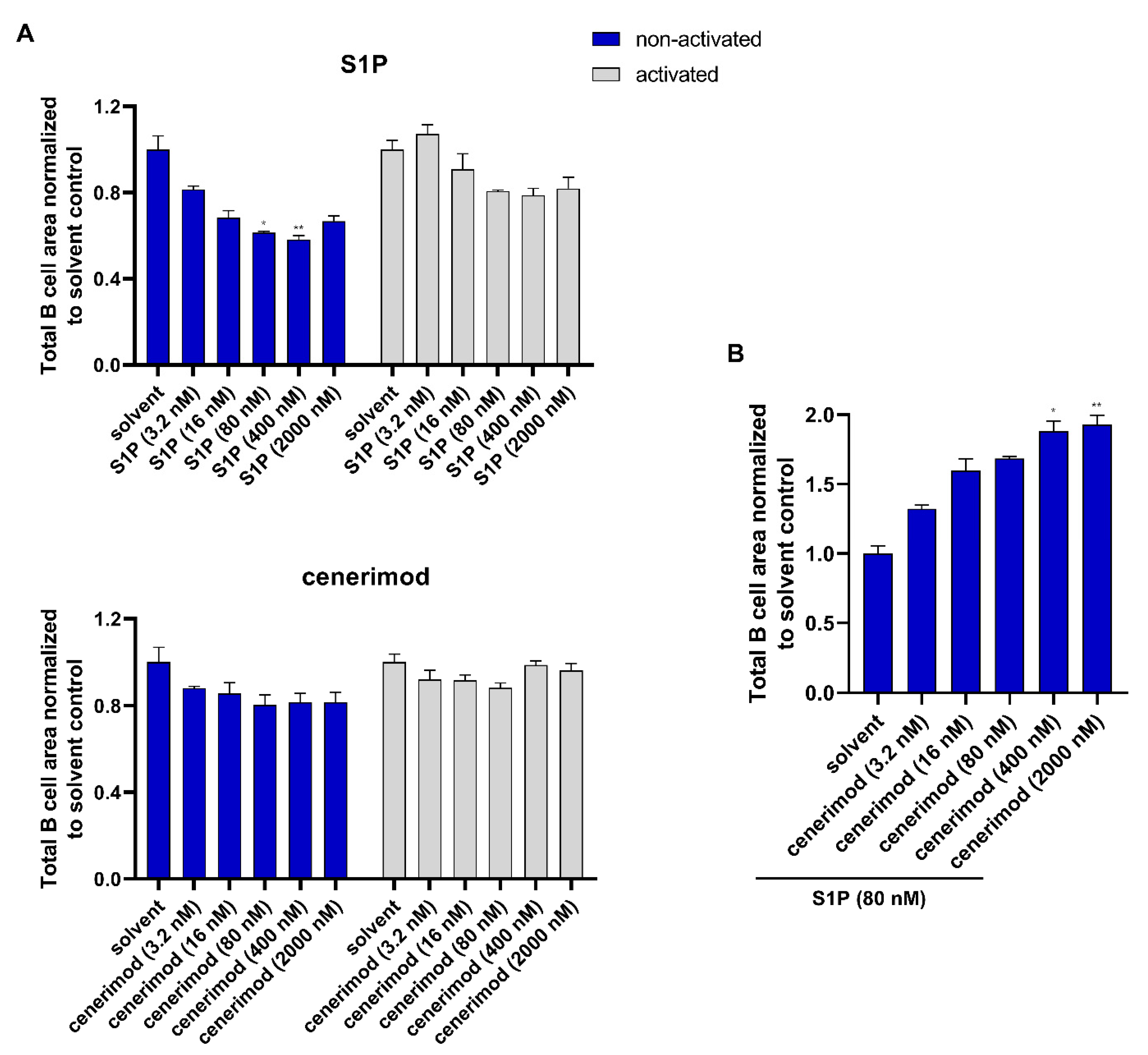
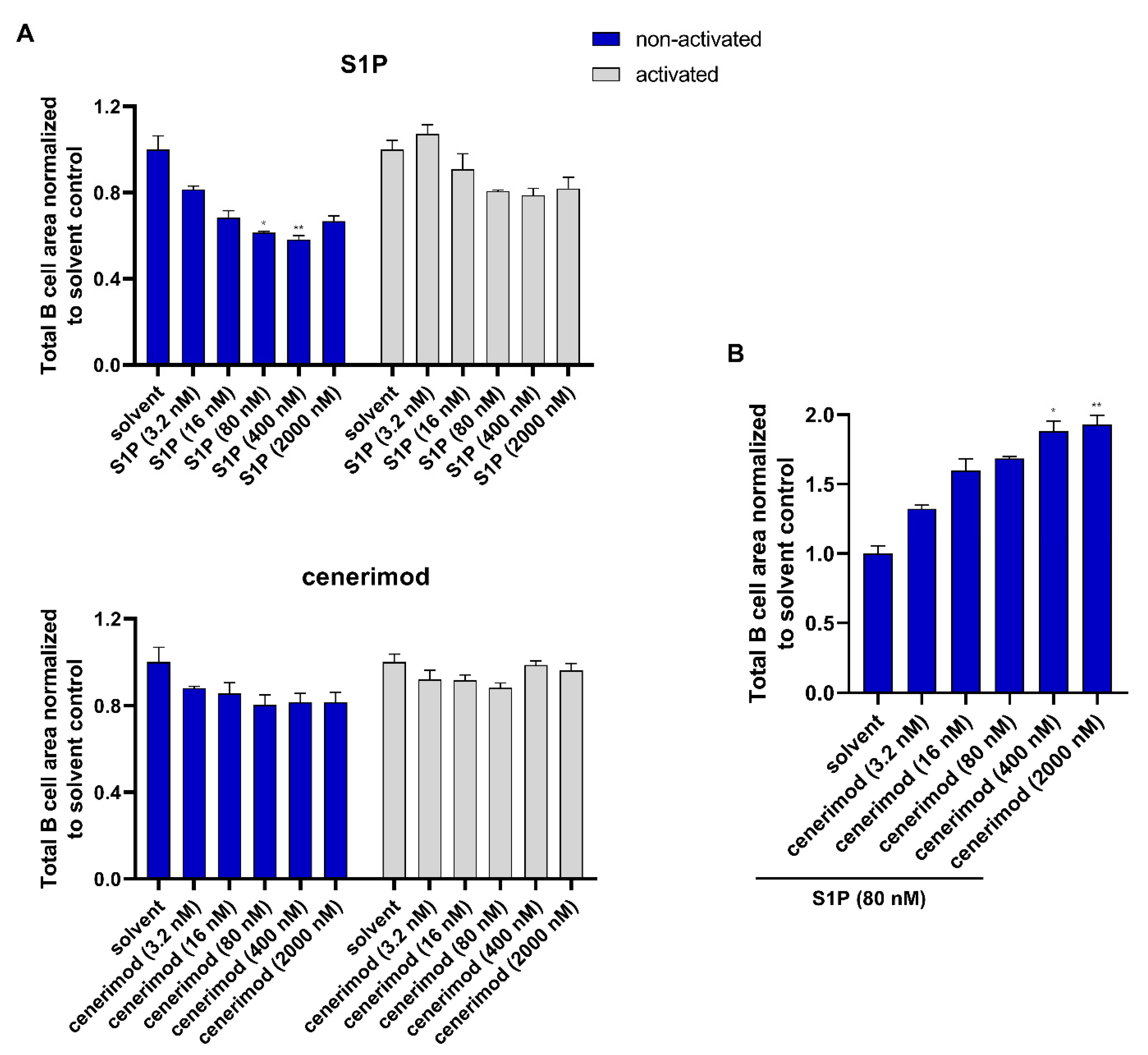
2.5. S1P1 Receptor Modulator Cenerimod Effectively Prevents B Cell Migration towards S1P
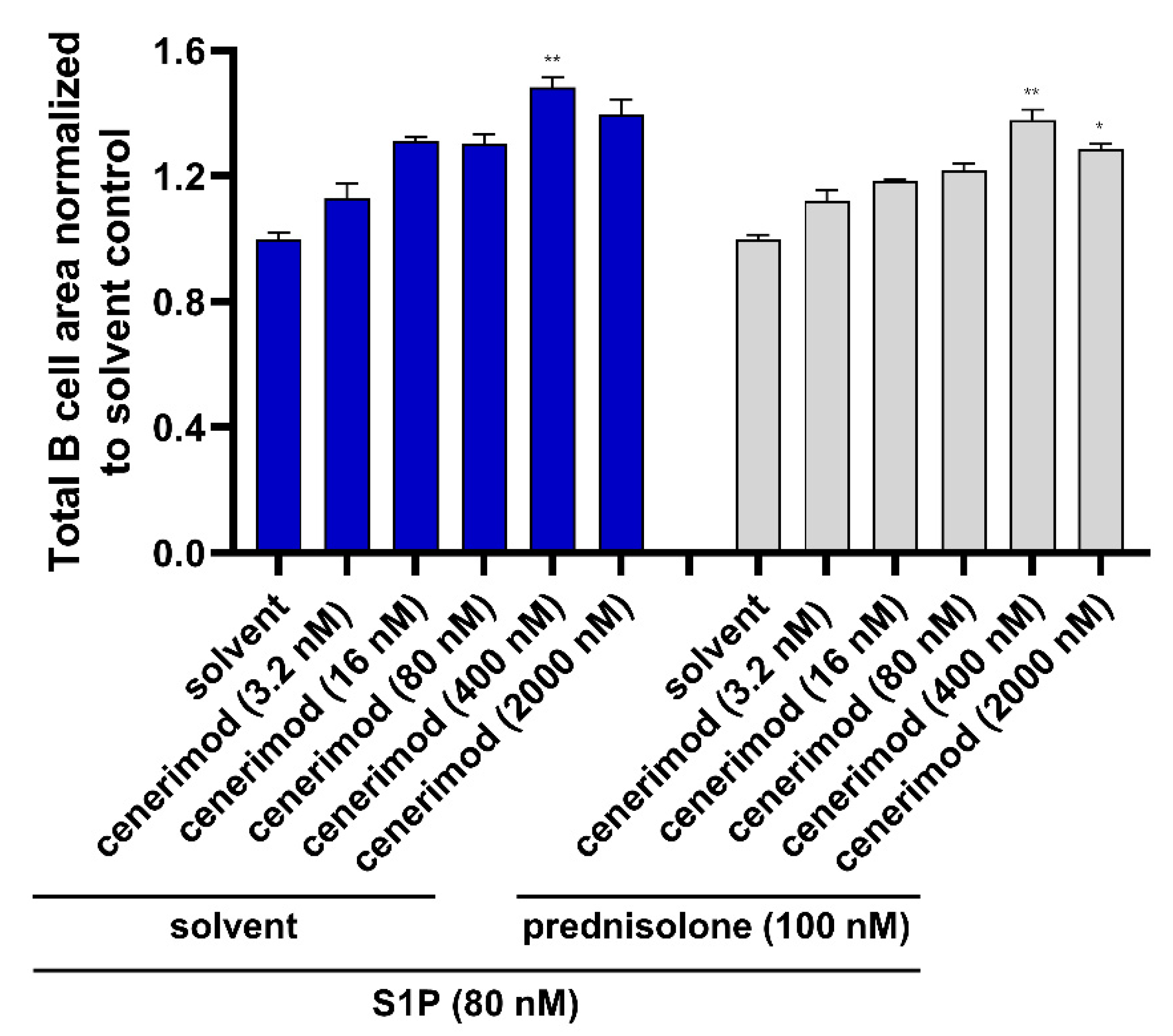
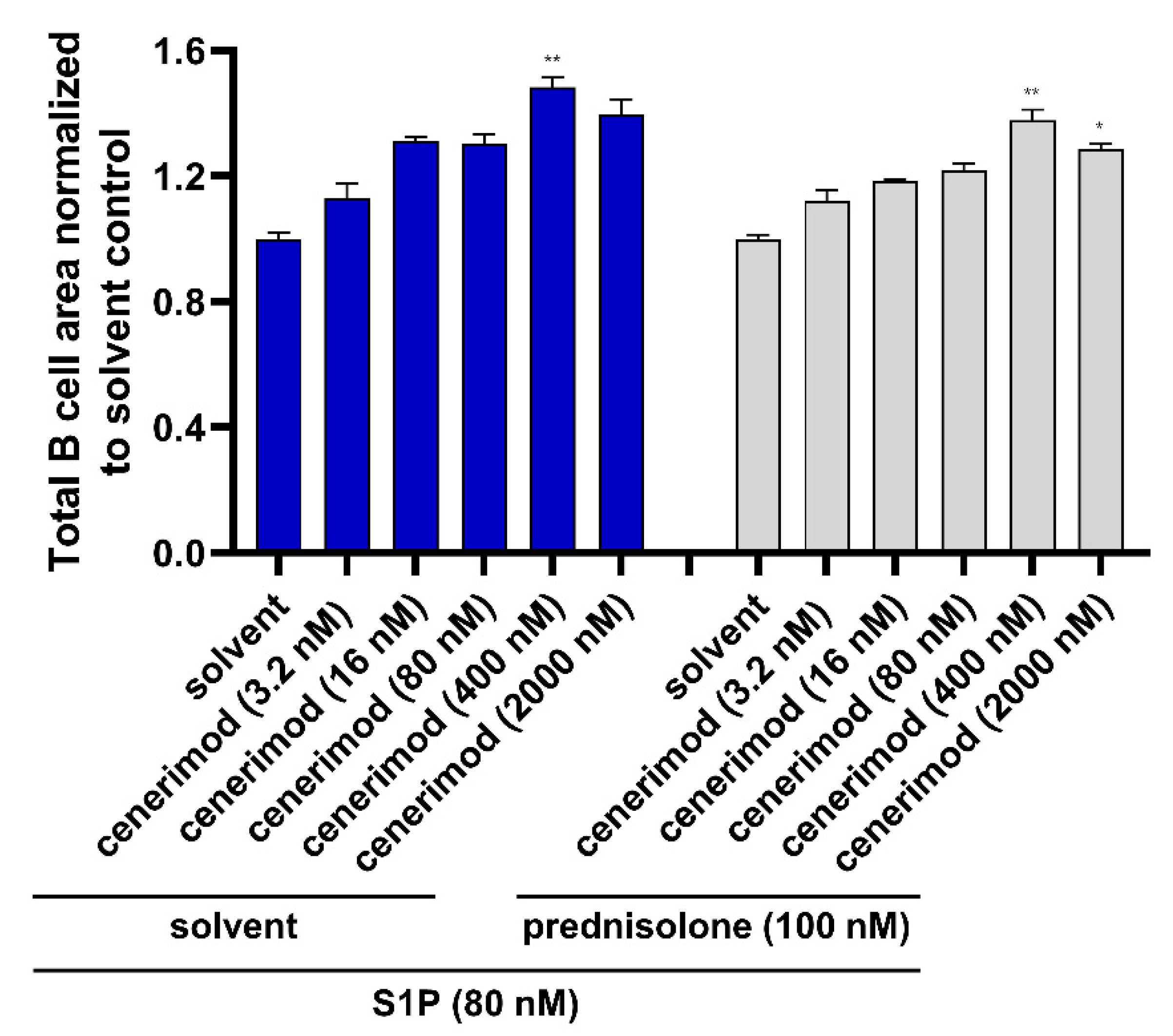
2.6. Co-Treatment with Prednisolone Has No Impact on the Ability of Cenerimod to Inhibit S1P-Mediated Migration in Primary Human B Lymphocytes
3. Discussion
4. Materials and Methods
4.1. Receptor Internalization Assay
4.2. CTV Proliferation Assay
4.3. Real-Time Migration Assay (IncuCyte™)
4.4. Western Blot Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hla, T.; Brinkmann, V. Sphingosine 1-Phosphate (S1P): Physiology and the Effects of S1P Receptor Modulation. Neurology 2011, 76, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jeldres, T.; Alvarez-Lobos, M.; Rivera-Nieves, J. Targeting Sphingosine-1-Phosphate Signaling in Immune-Mediated Diseases: Beyond Multiple Sclerosis. Drugs 2021, 81, 985–1002. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Giovannoni, G.; Hunter, S.F. Sphingosine 1-Phosphate Receptor Modulator Therapy for Multiple Sclerosis: Differential Downstream Receptor Signalling and Clinical Profile Effects. Drugs 2021, 81, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Yanagawa, Y.; Masubuchi, Y.; Kawaguchi, T.; Ohtsuki, M. FTY720, a Novel Immunosuppressant, Induces Sequestration of Circulating Mature Lymphocytes by Acceleration of Lymphocyte Homing in Rats. I. FTY720 Selectively Decreases the Number of Circulating Mature Lymphocytes by Acceleration of Lymphocyte Homing. J. Immunol. 1998, 160, 5037–5044. [Google Scholar]
- Gatfield, J.; Monnier, L.; Studer, R.; Bolli, M.H.; Steiner, B.; Nayler, O. Sphingosine-1-Phosphate (S1P) Displays Sustained S1P1 Receptor Agonism and Signaling through S1P Lyase-Dependent Receptor Recycling. Cell. Signal. 2014, 26, 1576–1588. [Google Scholar] [CrossRef]
- Rahman, A. Systemic Lupus Erythematosus. N. Engl. J. Med. 2008, 358, 929–939. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Huang, T.; Chang, S.; Wang, Y.; Fan, W.; Ji, H.; Wang, J.; Yang, J.; Kang, J.; Zhou, Y. Role of Sphingosine-1-Phosphate Mediated Signalling in Systemic Lupus Erythematosus. Prostaglandins Other Lipid Mediat. 2021, 156, 106584. [Google Scholar] [CrossRef] [PubMed]
- Piali, L.; Birker-Robaczewska, M.; Lescop, C.; Froidevaux, S.; Schmitz, N.; Morrison, K.; Kohl, C.; Rey, M.; Studer, R.; Vezzali, E.; et al. Cenerimod, a Novel Selective S1P 1 Receptor Modulator with Unique Signaling Properties. Pharm. Res. Perspect. 2017, 5, e00370. [Google Scholar] [CrossRef] [Green Version]
- Strasser, D.S.; Froidevaux, S.; Sippel, V.; Gerossier, E.; Grieder, U.; Pierlot, G.M.; Kieninger-Graefitsch, A.; Vezzali, E.; Stalder, A.K.; Renault, B.; et al. Preclinical to Clinical Translation of Cenerimod, a Novel S1P 1 Receptor Modulator, in Systemic Lupus Erythematosus. RMD Open 2020, 6, e001261. [Google Scholar] [CrossRef]
- Hermann, V.; Batalov, A.; Smakotina, S.; Juif, P.-E.; Cornelisse, P. First Use of Cenerimod, a Selective S1P 1 Receptor Modulator, for the Treatment of SLE: A Double-Blind, Randomised, Placebo-Controlled, Proof-of-Concept Study. Lupus Sci. Med. 2019, 6, e000354. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Matsuoka, T.; Chihara, N.; Miyake, S.; Sato, W.; Araki, M.; Okamoto, T.; Lin, Y.; Ogawa, M.; Murata, M.; et al. Differential Effects of Fingolimod on B-Cell Populations in Multiple Sclerosis. Mult. Scler. 2014, 20, 1371–1380. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Yako, Y.; Fujioka, Y.; Kajita, M.; Kameyama, T.; Kon, S.; Ishikawa, S.; Ohba, Y.; Ohno, Y.; Kihara, A.; et al. A Role of the Sphingosine-1-Phosphate (S1P)–S1P Receptor 2 Pathway in Epithelial Defense against Cancer (EDAC). Mol. Biol. Cell 2016, 27, 491–499. [Google Scholar] [CrossRef]
- Edsall, L.C.; Spiegel, S. Enzymatic Measurement of Sphingosine 1-Phosphate. Anal. Biochem. 1999, 272, 80–86. [Google Scholar] [CrossRef]
- Garris, C.S.; Blaho, V.A.; Hla, T.; Han, M.H. Sphingosine-1-Phosphate Receptor 1 Signalling in T Cells: Trafficking and Beyond. Immunology 2014, 142, 347–353. [Google Scholar] [CrossRef]
- Shiow, L.R.; Rosen, D.B.; Brdičková, N.; Xu, Y.; An, J.; Lanier, L.L.; Cyster, J.G.; Matloubian, M. CD69 Acts Downstream of Interferon-α/β to Inhibit S1P1 and Lymphocyte Egress from Lymphoid Organs. Nature 2006, 440, 540–544. [Google Scholar] [CrossRef]
- Bankovich, A.J.; Shiow, L.R.; Cyster, J.G. CD69 Suppresses Sphingosine 1-Phosophate Receptor-1 (S1P1) Function through Interaction with Membrane Helix 4. J. Biol. Chem. 2010, 285, 22328–22337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benechet, A.P.; Menon, M.; Xu, D.; Samji, T.; Maher, L.; Murooka, T.T.; Mempel, T.R.; Sheridan, B.S.; Lemoine, F.M.; Khanna, K.M. T Cell-Intrinsic S1PR1 Regulates Endogenous Effector T-Cell Egress Dynamics from Lymph Nodes during Infection. Proc. Natl. Acad. Sci. USA 2016, 113, 2182–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matloubian, M.; Lo, C.G.; Cinamon, G.; Lesneski, M.J.; Xu, Y.; Brinkmann, V.; Allende, M.L.; Proia, R.L.; Cyster, J.G. Lymphocyte Egress from Thymus and Peripheral Lymphoid Organs Is Dependent on S1P Receptor 1. Nature 2004, 427, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Matsuyuki, H.; Shimano, K.; Kataoka, H.; Sugahara, K.; Chiba, K. Migration of CD4 T Cells and Dendritic Cells toward Sphingosine 1-Phosphate (S1P) Is Mediated by Different Receptor Subtypes: S1P Regulates the Functions of Murine Mature Dendritic Cells via S1P Receptor Type 3. J. Immunol. 2007, 178, 3437–3446. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ribeiro, B.; Li, H.; Myer, L.; Chase, P.; Surti, N.; Lippy, J.; Zhang, L.; Cvijic, M.E. Leveraging the IncuCyte Technology for Higher-Throughput and Automated Chemotaxis Assays for Target Validation and Compound Characterization. SLAS Discov. 2018, 23, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, S.; Danza, A.; Arias Saavedra, M.; Carlomagno, A.; Goizueta, M.C.; Vivero, F.; Ruiz-Irastorza, G. Glucocorticoids in Systemic Lupus Erythematosus. Ten Questions and Some Issues. J. Clin. Med. 2020, 9, 2709. [Google Scholar] [CrossRef] [PubMed]
- Lateef, A.; Petri, M. Unmet Medical Needs in Systemic Lupus Erythematosus. Arthritis Res. 2012, 14, S4. [Google Scholar] [CrossRef] [Green Version]
- Fanouriakis, A.; Kostopoulou, M.; Alunno, A.; Aringer, M.; Bajema, I.; Boletis, J.N.; Cervera, R.; Doria, A.; Gordon, C.; Govoni, M.; et al. 2019 Update of the EULAR Recommendations for the Management of Systemic Lupus Erythematosus. Ann. Rheum. Dis. 2019, 78, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Winkler, J.; Derendorf, H. A Pharmacokinetic/Pharmacodynamic Approach to Predict Total Prednisolone Concentrations in Human Plasma. J. Pharm. Pharm. 2007, 34, 355–372. [Google Scholar] [CrossRef] [PubMed]
- Kleinwort, A.; Lührs, F.; Heidecke, C.-D.; Lipp, M.; Schulze, T. S1P Signalling Differentially Affects Migration of Peritoneal B Cell Populations In Vitro and Influences the Production of Intestinal IgA In Vivo. Int. J. Mol. Sci. 2018, 19, 391. [Google Scholar] [CrossRef] [Green Version]
- Kunisawa, J.; Gohda, M.; Kurashima, Y.; Ishikawa, I.; Higuchi, M.; Kiyono, H. Sphingosine 1-Phosphate–Dependent Trafficking of Peritoneal B Cells Requires Functional NFκB-Inducing Kinase in Stromal Cells. Blood 2008, 111, 7. [Google Scholar] [CrossRef]
- De Wit, R.H.; de Munnik, S.M.; Leurs, R.; Vischer, H.F.; Smit, M.J. Molecular Pharmacology of Chemokine Receptors. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2016; Volume 570, pp. 457–515. ISBN 978-0-12-802171-2. [Google Scholar]
- Graeler, M.; Goetzl, E.J. Activation-regulated Expression and Chemotactic Function of Sphingosine 1-phosphate Receptors in Mouse Splenic T Cells. FASEB J. 2002, 16, 1874–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlicher, L.; Kulig, P.; von Münchow, A.; Murphy, M.J.; Keller, M.P. In Vitro Characterization of Sphingosine 1-Phosphate Receptor 1 (S1P1) Expression and Mediated Migration of Primary Human T and B Cells in the Context of Cenerimod, a Novel, Selective S1P1 Receptor Modulator. Int. J. Mol. Sci. 2022, 23, 1191. https://doi.org/10.3390/ijms23031191
Schlicher L, Kulig P, von Münchow A, Murphy MJ, Keller MP. In Vitro Characterization of Sphingosine 1-Phosphate Receptor 1 (S1P1) Expression and Mediated Migration of Primary Human T and B Cells in the Context of Cenerimod, a Novel, Selective S1P1 Receptor Modulator. International Journal of Molecular Sciences. 2022; 23(3):1191. https://doi.org/10.3390/ijms23031191
Chicago/Turabian StyleSchlicher, Lisa, Paulina Kulig, Audrey von Münchow, Mark J. Murphy, and Marcel P. Keller. 2022. "In Vitro Characterization of Sphingosine 1-Phosphate Receptor 1 (S1P1) Expression and Mediated Migration of Primary Human T and B Cells in the Context of Cenerimod, a Novel, Selective S1P1 Receptor Modulator" International Journal of Molecular Sciences 23, no. 3: 1191. https://doi.org/10.3390/ijms23031191
APA StyleSchlicher, L., Kulig, P., von Münchow, A., Murphy, M. J., & Keller, M. P. (2022). In Vitro Characterization of Sphingosine 1-Phosphate Receptor 1 (S1P1) Expression and Mediated Migration of Primary Human T and B Cells in the Context of Cenerimod, a Novel, Selective S1P1 Receptor Modulator. International Journal of Molecular Sciences, 23(3), 1191. https://doi.org/10.3390/ijms23031191