
Circulating miR-499a-5p Is a Potential Biomarker of MYH7—Associated Hypertrophic Cardiomyopathy

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
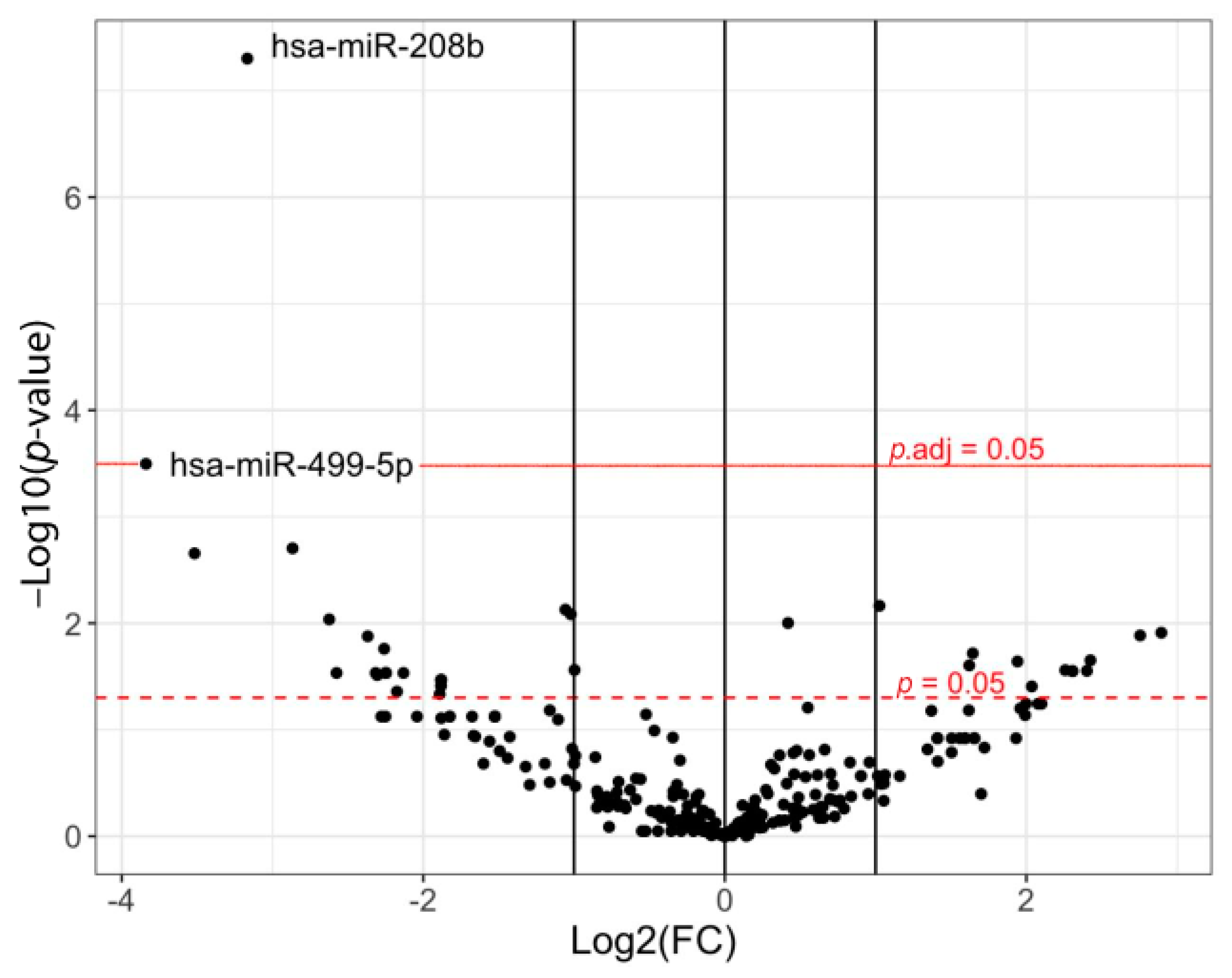
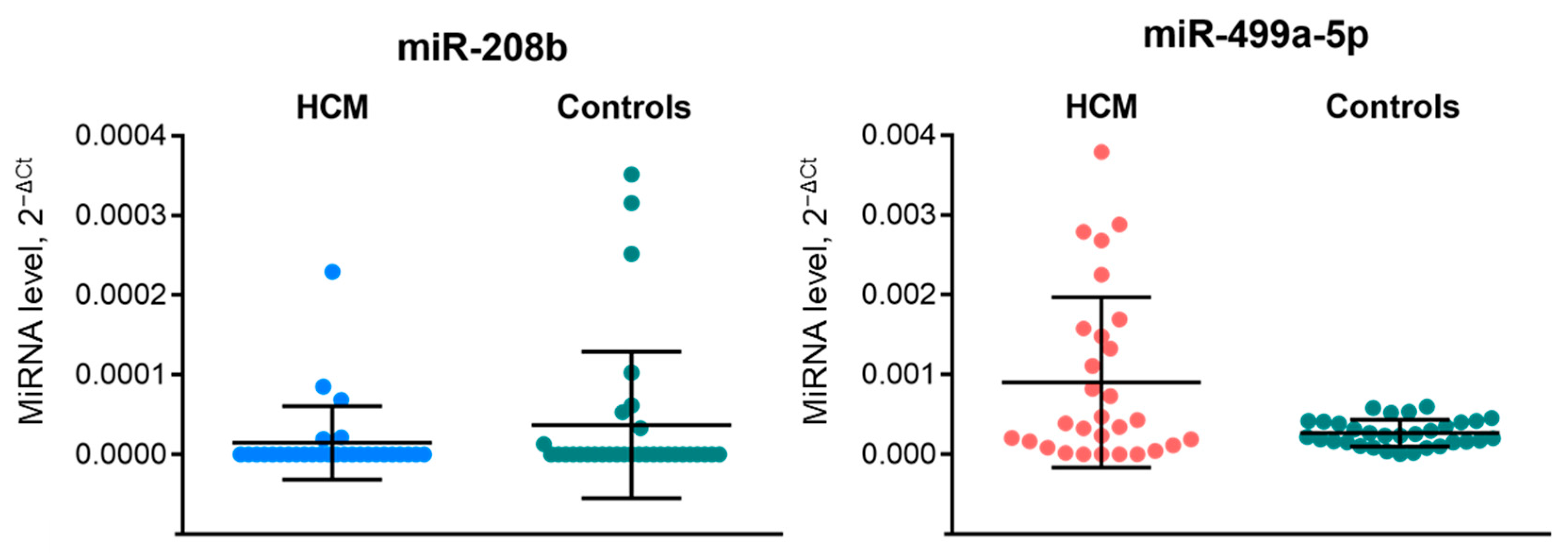
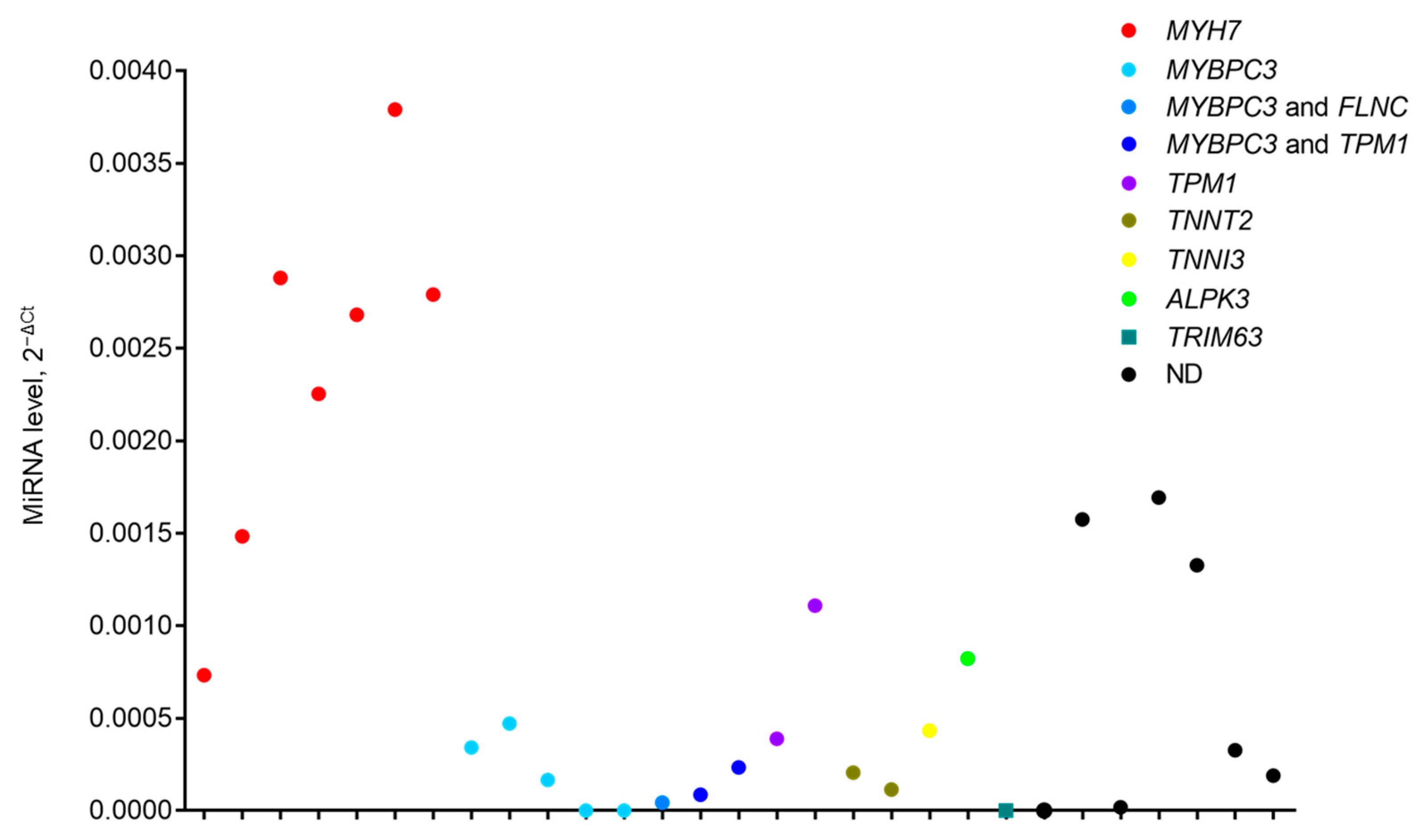
2. Results
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Plasma Collection and RNA Extraction
4.3. Small RNA Sequencing Libraries Preparation
4.4. Small RNA-seq Data Analysis
4.5. RT-qPCR
4.6. Statistical Analysis
4.7. ROC-Curve Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maron, B.J. Clinical Course and Management of Hypertrophic Cardiomyopathy. N. Engl. J. Med. 2018, 379, 655–668. [Google Scholar] [CrossRef]
- Cirino, A.L.; Ho, C. Hypertrophic Cardiomyopathy Overview. In GeneReviews®; Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Mirzaa, G.M., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Baulina, N.; Kiselev, I.; Chumakova, O.; Favorova, O. Hypertrophic Cardiomyopathy as an Oligogenic Disease: Transcriptomic Arguments. Mol. Biol. 2020, 54, 840–850. [Google Scholar] [CrossRef]
- Viswanathan, S.K.; Sanders, H.K.; McNamara, J.W.; Jagadeesan, A.; Jahangir, A.; Tajik, A.J.; Sadayappan, S. Hypertrophic Cardiomyopathy Clinical Phenotype Is Independent of Gene Mutation and Mutation Dosage. PLoS ONE 2017, 12, e0187948. [Google Scholar] [CrossRef] [Green Version]
- Weissler-Snir, A.; Hindieh, W.; Gruner, C.; Fourey, D.; Appelbaum, E.; Rowin, E.; Care, M.; Lesser, J.R.; Haas, T.S.; Udelson, J.E.; et al. Lack of Phenotypic Differences by Cardiovascular Magnetic Resonance Imaging in MYH7 (β-Myosin Heavy Chain)- Versus MYBPC3 (Myosin-Binding Protein C)-Related Hypertrophic Cardiomyopathy. Circ. Cardiovasc. Imaging 2017, 10, e005311. [Google Scholar] [CrossRef] [Green Version]
- Marian, A.J. Molecular Genetic Basis of Hypertrophic Cardiomyopathy. Circ. Res. 2021, 128, 1533–1553. [Google Scholar] [CrossRef]
- Walsh, R.; Thomson, K.L.; Ware, J.S.; Funke, B.H.; Woodley, J.; McGuire, K.J.; Mazzarotto, F.; Blair, E.; Seller, A.; Taylor, J.C.; et al. Reassessment of Mendelian Gene Pathogenicity Using 7,855 Cardiomyopathy Cases and 60,706 Reference Samples. Genet. Med. 2017, 19, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Van Driest, S.L.; Ackerman, M.J.; Ommen, S.R.; Shakur, R.; Will, M.L.; Nishimura, R.A.; Tajik, A.J.; Gersh, B.J. Prevalence and Severity of “Benign” Mutations in the β-Myosin Heavy Chain, Cardiac Troponin T, and α-Tropomyosin Genes in Hypertrophic Cardiomyopathy. Circulation 2002, 106, 3085–3090. [Google Scholar] [CrossRef] [Green Version]
- Oliva-Sandoval, M.J.; Ruiz-Espejo, F.; Monserrat, L.; Hermida-Prieto, M.; Sabater, M.; García-Molina, E.; Ortiz, M.; Rodríguez-García, M.I.; Núñez, L.; Gimeno, J.R.; et al. Insights into Genotype–Phenotype Correlation in Hypertrophic Cardiomyopathy. Findings from 18 Spanish Families with a Single Mutation in MYBPC3. Heart 2010, 96, 1980–1984. [Google Scholar] [CrossRef]
- Bagnall, R.D.; Ingles, J.; Dinger, M.E.; Cowley, M.J.; Ross, S.B.; Minoche, A.E.; Lal, S.; Turner, C.; Colley, A.; Rajagopalan, S.; et al. Whole Genome Sequencing Improves Outcomes of Genetic Testing in Patients with Hypertrophic Cardiomyopathy. J. Am. Coll. Cardiol. 2018, 72, 419–429. [Google Scholar] [CrossRef]
- Jansweijer, J.A.; van Spaendonck-Zwarts, K.Y.; Tanck, M.W.T.; van Tintelen, J.P.; Christiaans, I.; van der Smagt, J.; Vermeer, A.; Bos, J.M.; Moss, A.J.; Swan, H.; et al. Heritability in Genetic Heart Disease: The Role of Genetic Background. Open Heart 2019, 6, e000929. [Google Scholar] [CrossRef]
- Maron, B.J.; Maron, M.S.; Maron, B.A.; Loscalzo, J. Expanding Pathobiological Mechanisms in Hypertrophic Cardiomyopathy to Explain Heterogeneity: JACC Review Topic of the Week. J. Am. Coll. Cardiol. 2019, 73, 1978–1986. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Z.; Zhang, W.; Zhang, L. MicroRNAs Play an Essential Role in Autophagy Regulation in Various Disease Phenotypes. BioFactors 2019, 45, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Chiti, E.; Di Paolo, M.; Turillazzi, E.; Rocchi, A. MicroRNAs in Hypertrophic, Arrhythmogenic and Dilated Cardiomyopathy. Diagnostics 2021, 11, 1720. [Google Scholar] [CrossRef] [PubMed]
- Palacín, M.; Coto, E.; Reguero, J.R.; Morís, C.; Alvarez, V. Profile of MicroRNAs in the Plasma of Hypertrophic Cardiomyopathy Patients Compared to Healthy Controls. Int. J. Cardiol. 2013, 167, 3075–3076. [Google Scholar] [CrossRef]
- Roncarati, R.; Viviani Anselmi, C.; Losi, M.A.; Papa, L.; Cavarretta, E.; Da Costa Martins, P.; Contaldi, C.; Saccani Jotti, G.; Franzone, A.; Galastri, L.; et al. Circulating MiR-29a, Among Other Up-Regulated MicroRNAs, Is the Only Biomarker for Both Hypertrophy and Fibrosis in Patients with Hypertrophic Cardiomyopathy. J. Am. Coll. Cardiol. 2014, 63, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Ellims, A.H.; Moore, X.; White, D.A.; Taylor, A.J.; Chin-Dusting, J.; Dart, A.M. Circulating MicroRNAs as Biomarkers for Diffuse Myocardial Fibrosis in Patients with Hypertrophic Cardiomyopathy. J. Transl. Med. 2015, 13, 314. [Google Scholar] [CrossRef]
- Miyamoto, S.; Ho, C.Y.; Korst, A.; Sucharov, C. Circulating microRNAs as a Biomarker of Pediatric Hypertrophic Cardiomyopathy. J. Am. Coll. Cardiol. 2019, 73, 680. [Google Scholar] [CrossRef]
- Ntelios, D.; Efthimiadis, G.; Zegkos, T.; Didagelos, M.; Katopodi, T.; Meditskou, S.; Parcharidou, D.; Karvounis, H.; Tzimagiorgis, G. Correlation of MiR-146a-5p Plasma Levels and Rs2910164 Polymorphism with Left Ventricle Outflow Tract Obstruction in Hypertrophic Cardiomyopathy. Hell. J. Cardiol. 2021, 62, 349–354. [Google Scholar] [CrossRef]
- Derda, A.A.; Thum, S.; Lorenzen, J.M.; Bavendiek, U.; Heineke, J.; Keyser, B.; Stuhrmann, M.; Givens, R.C.; Kennel, P.J.; Schulze, P.C.; et al. Blood-Based MicroRNA Signatures Differentiate Various Forms of Cardiac Hypertrophy. Int. J. Cardiol. 2015, 196, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Ntelios, D.; Georgiou, E.; Alexouda, S.; Malousi, A.; Efthimiadis, G.; Tzimagiorgis, G. A Critical Approach for Successful Use of Circulating MicroRNAs as Biomarkers in Cardiovascular Diseases: The Case of Hypertrophic Cardiomyopathy. Heart Fail. Rev. 2022, 27, 281–294. [Google Scholar] [CrossRef]
- Gao, J.; Collyer, J.; Wang, M.; Sun, F.; Xu, F. Genetic Dissection of Hypertrophic Cardiomyopathy with Myocardial RNA-Seq. Int. J. Mol. Sci. 2020, 21, 3040. [Google Scholar] [CrossRef] [PubMed]
- Fichtlscherer, S.; De Rosa, S.; Fox, H.; Schwietz, T.; Fischer, A.; Liebetrau, C.; Weber, M.; Hamm, C.W.; Röxe, T.; Müller-Ardogan, M.; et al. Circulating MicroRNAs in Patients with Coronary Artery Disease. Circ. Res. 2010, 107, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenz, M.; Robbins, J. Impact of Beta-Myosin Heavy Chain Expression on Cardiac Function during Stress. J. Am. Coll. Cardiol. 2004, 44, 2390–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rundell, V.L.M.; Manaves, V.; Martin, A.F.; de Tombe, P.P. Impact of β-Myosin Heavy Chain Isoform Expression on Cross-Bridge Cycling Kinetics. Am. J. Physiol.-Heart Circ. Physiol. 2005, 288, H896–H903. [Google Scholar] [CrossRef] [Green Version]
- Broadwell, L.J.; Smallegan, M.J.; Rigby, K.M.; Navarro-Arriola, J.S.; Montgomery, R.L.; Rinn, J.L.; Leinwand, L.A. Myosin 7b Is a Regulatory Long Noncoding RNA (LncMYH7b) in the Human Heart. J. Biol. Chem. 2021, 296, 100694. [Google Scholar] [CrossRef]
- Desjardins, P.R.; Burkman, J.M.; Shrager, J.B.; Allmond, L.A.; Stedman, H.H. Evolutionary Implications of Three Novel Members of the Human Sarcomeric Myosin Heavy Chain Gene Family. Mol. Biol. Evol. 2002, 19, 375–393. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.A.; Broadwell, L.J.; Buvoli, M.; Leinwand, L.A. Nonproductive Splicing Prevents Expression of MYH7b Protein in the Mammalian Heart. J. Am. Heart Assoc. 2021, 10, e020965. [Google Scholar] [CrossRef]
- Rossi, A.C.; Mammucari, C.; Argentini, C.; Reggiani, C.; Schiaffino, S. Two Novel/Ancient Myosins in Mammalian Skeletal Muscles: MYH14/7b and MYH15 Are Expressed in Extraocular Muscles and Muscle Spindles. J. Physiol. 2010, 588, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.L.; Buvoli, M.; Leinwand, L.A. Uncoupling of Expression of an Intronic MicroRNA and Its Myosin Host Gene by Exon Skipping. Mol. Cell. Biol. 2010, 30, 1937–1945. [Google Scholar] [CrossRef] [Green Version]
- Warkman, A.S.; Whitman, S.A.; Miller, M.K.; Garriock, R.J.; Schwach, C.M.; Gregorio, C.C.; Krieg, P.A. Developmental Expression and Cardiac Transcriptional Regulation of Myh7b, a Third Myosin Heavy Chain in the Vertebrate Heart. Cytoskeleton 2012, 69, 324–335. [Google Scholar] [CrossRef] [Green Version]
- Esposito, T.; Sampaolo, S.; Limongelli, G.; Varone, A.; Formicola, D.; Diodato, D.; Farina, O.; Napolitano, F.; Pacileo, G.; Gianfrancesco, F.; et al. Digenic Mutational Inheritance of the Integrin Alpha 7 and the Myosin Heavy Chain 7B Genes Causes Congenital Myopathy with Left Ventricular Non-Compact Cardiomyopathy. Orphanet J. Rare Dis. 2013, 8, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Li, Z.; Nie, J.; Wang, H.; Yu, B.; Wen, Z.; Sun, Y.; Shi, X.; Jin, L.; Wang, D.-W. MYH7B Variants Cause Hypertrophic Cardiomyopathy by Activating the CaMK-Signaling Pathway. Sci. China Life Sci. 2020, 63, 1347–1362. [Google Scholar] [CrossRef] [PubMed]
- Landrum, M.J.; Lee, J.M.; Benson, M.; Brown, G.R.; Chao, C.; Chitipiralla, S.; Gu, B.; Hart, J.; Hoffman, D.; Jang, W.; et al. ClinVar: Improving Access to Variant Interpretations and Supporting Evidence. Nucleic Acids Res. 2018, 46, D1062–D1067. [Google Scholar] [CrossRef] [Green Version]
- Thottakara, T.; Lund, N.; Krämer, E.; Kirchhof, P.; Carrier, L.; Patten, M. A Novel MiRNA Screen Identifies MiRNA-4454 as a Candidate Biomarker for Ventricular Fibrosis in Patients with Hypertrophic Cardiomyopathy. Biomolecules 2021, 11, 1718. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhou, Y.; Wang, C.-X. LncRNA-MIAT Regulates Fibrosis in Hypertrophic Cardiomyopathy (HCM) by Mediating the Expression of MiR-29a-3p. J. Cell Biochem. 2019, 120, 7265–7275. [Google Scholar] [CrossRef] [PubMed]
- van Rooij, E.; Quiat, D.; Johnson, B.A.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Kelm, R.J.; Olson, E.N. A Family of MicroRNAs Encoded by Myosin Genes Governs Myosin Expression and Muscle Performance. Dev. Cell 2009, 17, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Sadayappan, S.; Gulick, J.; Klevitsky, R.; Lorenz, J.N.; Sargent, M.; Molkentin, J.D.; Robbins, J. Cardiac Myosin Binding Protein-C Phosphorylation in a {beta}-Myosin Heavy Chain Background. Circulation 2009, 119, 1253–1262. [Google Scholar] [CrossRef]
- Nakao, K.; Minobe, W.; Roden, R.; Bristow, M.R.; Leinwand, L.A. Myosin Heavy Chain Gene Expression in Human Heart Failure. J. Clin. Investig. 1997, 100, 2362–2370. [Google Scholar] [CrossRef]
- Miyata, S.; Minobe, W.; Bristow, M.R.; Leinwand, L.A. Myosin Heavy Chain Isoform Expression in the Failing and Nonfailing Human Heart. Circ. Res. 2000, 86, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Lowes, B.D.; Gilbert, E.M.; Abraham, W.T.; Minobe, W.A.; Larrabee, P.; Ferguson, D.; Wolfel, E.E.; Lindenfeld, J.A.; Tsvetkova, T.; Robertson, A.D.; et al. Myocardial Gene Expression in Dilated Cardiomyopathy Treated with Beta-Blocking Agents. N. Engl. J. Med. 2002, 346, 1357–1365. [Google Scholar] [CrossRef]
- Montag, J.; Petersen, B.; Flögel, A.K.; Becker, E.; Lucas-Hahn, A.; Cost, G.J.; Mühlfeld, C.; Kraft, T.; Niemann, H.; Brenner, B. Successful Knock-in of Hypertrophic Cardiomyopathy-Mutation R723G into the MYH7 Gene Mimics HCM Pathology in Pigs. Sci. Rep. 2018, 8, 4786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montag, J.; Kowalski, K.; Makul, M.; Ernstberger, P.; Radocaj, A.; Beck, J.; Becker, E.; Tripathi, S.; Keyser, B.; Mühlfeld, C.; et al. Burst-Like Transcription of Mutant and Wildtype MYH7-Alleles as Possible Origin of Cell-to-Cell Contractile Imbalance in Hypertrophic Cardiomyopathy. Front. Physiol. 2018, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Elliott, P.M.; Anastasakis, A.; Borger, M.A.; Borggrefe, M.; Cecchi, F.; Charron, P.; Hagege, A.A.; Lafont, A.; Limongelli, G.; Mahrholdt, H.; et al. 2014 ESC Guidelines on Diagnosis and Management of Hypertrophic Cardiomyopathy: The Task Force for the Diagnosis and Management of Hypertrophic Cardiomyopathy of the European Society of Cardiology (ESC). Eur. Heart J. 2014, 35, 2733–2779. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential Expression Analysis for Sequence Count Data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential Expression Analysis of Multifactor RNA-Seq Experiments with Respect to Biological Variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Auer, P.L.; Doerge, R.W. Statistical Design and Analysis of RNA Sequencing Data. Genetics 2010, 185, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Discovery Group | Validation Group |
|---|---|---|
| Female, n (%) | 5 (55.5) | 12 (41) |
| Age, years | 50.2 ± 10.3 | 48.0 ± 12.6 |
| BMI, kg/m2 | 26.69 ± 4.73 | 26.95 ± 4.14 |
| Five-year HCM Risk-SCD (%) | 5.24 ± 2.94 | 3.87 ± 2.30 |
| Family history of HCM, n (%) | 4 (44.4) | 12 (41.4) |
| Family history of SCD, n (%) | 2 (22.2) | 5 (17.2) |
| Atrial fibrillation, n (%) | 1 (11.1) | 4 (13.8) |
| Ventricular tachycardia, n (%) | 4 (44.4) | 6 (20.7) |
| Arterial hypertension, n (%) | 3 (33.3) | 12 (41.4) |
| Coronary heart disease, n (%) | 1 (11.1) | 2 (6.9) |
| Diabetes mellitus, n (%) | 0 (0) | 1 (3.4) |
| Echocardiography | ||
| Maximal LVWT, mm | 25.50 ± 6.41 | 23.56 ± 5.54 |
| Left atrial diameter, mm | 42.11 ± 5.30 | 44.00 (41.00–47.00) |
| LA ESV index, mL/m2 | 45.84 ± 13.45 | 44.39 ± 10.47 |
| LV EDD index, mm/m2 | 22.30 (21.80–24.85) | 23.57 ± 2.20 |
| LV ESD index, mm/m2 | 15.10 (9.70–16.50) | 13.90 (10.04–16.00) |
| LV EDV index, mL/m2 | 47.88 ± 13.97 | 44.41 ± 11.78 |
| LV ESV index, mL/m2 | 16.99 ± 8.66 | 13.10 (10.80–17.60) |
| LVOT obstruction, n (%) | 5 (55.5) | 9 (31.0) |
| LVEF (%) | 65.89 ± 8.25 | 67.86 ± 6.77 |
| Mitral E-e’ ratio | 14.15 ± 9.50 | 9.30 (6.35–14.3) |
| Apical form of HCM, n (%) | 0 (0) | 5 (17.2) |
| Electrocardiography | ||
| Pathological Q Waves, n (%) | 4 (44.4) | 7 (24.1) |
| T-Wave inversion, n (%) | 8 (88.9) | 22 (75.9) |
| Sokolow-Lyon index, mm | 32.11 ± 16.87 | 31.00 (25.00–41.00) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baulina, N.; Pisklova, M.; Kiselev, I.; Chumakova, O.; Zateyshchikov, D.; Favorova, O. Circulating miR-499a-5p Is a Potential Biomarker of MYH7—Associated Hypertrophic Cardiomyopathy. Int. J. Mol. Sci. 2022, 23, 3791. https://doi.org/10.3390/ijms23073791
Baulina N, Pisklova M, Kiselev I, Chumakova O, Zateyshchikov D, Favorova O. Circulating miR-499a-5p Is a Potential Biomarker of MYH7—Associated Hypertrophic Cardiomyopathy. International Journal of Molecular Sciences. 2022; 23(7):3791. https://doi.org/10.3390/ijms23073791
Chicago/Turabian StyleBaulina, Natalia, Maria Pisklova, Ivan Kiselev, Olga Chumakova, Dmitry Zateyshchikov, and Olga Favorova. 2022. "Circulating miR-499a-5p Is a Potential Biomarker of MYH7—Associated Hypertrophic Cardiomyopathy" International Journal of Molecular Sciences 23, no. 7: 3791. https://doi.org/10.3390/ijms23073791
APA StyleBaulina, N., Pisklova, M., Kiselev, I., Chumakova, O., Zateyshchikov, D., & Favorova, O. (2022). Circulating miR-499a-5p Is a Potential Biomarker of MYH7—Associated Hypertrophic Cardiomyopathy. International Journal of Molecular Sciences, 23(7), 3791. https://doi.org/10.3390/ijms23073791