Abstract
Virus-specific proteins, including coat proteins, movement proteins, replication proteins, and suppressors of RNA interference are capable of triggering the hypersensitive response (HR), which is a type of cell death in plants. The main cell death signaling pathway involves direct interaction of HR-inducing proteins with nucleotide-binding leucine-rich repeats (NLR) proteins encoded by plant resistance genes. Singleton NLR proteins act as both sensor and helper. In other cases, NLR proteins form an activation network leading to their oligomerization and formation of membrane-associated resistosomes, similar to metazoan inflammasomes and apoptosomes. In resistosomes, coiled-coil domains of NLR proteins form Ca2+ channels, while toll-like/interleukin-1 receptor-type (TIR) domains form oligomers that display NAD+ glycohydrolase (NADase) activity. This review is intended to highlight the current knowledge on plant innate antiviral defense signaling pathways in an attempt to define common features of antiviral resistance across the kingdoms of life.
1. Introduction
Programmed cell death (PCD) is a fundamental aspect of functioning of multicellular organisms. At the morphological and biochemical levels, there are a number of basic similarities, even among representatives of different kingdoms of living organisms (plants and animals), although the molecular mechanisms of programmed cell death vary. For example, classical apoptosis is a characteristic exclusive to animals, and caspases that are responsible for the development of this type of programmed cell death are not found in living organisms outside the animal kingdom [1]. The similarity between the animal and plant kingdoms is manifested at the level of alternative death processes, such as autophagy and ferroptosis [2,3,4,5]. The caspase-like activity observed in plants is attributable to metacaspases, vacuolar processing enzymes (VPE), and the papain-like cysteine proteases [6,7,8]. Protein substrates that are cleaved during the development of PCD in plants and animals may be orthologs. DNA fragmentation is also characteristic of many types of PCD in organisms belonging to different kingdoms. PCD in plants can be associated with their development (developmental PCD, dPCD) or with the invasion of a pathogen (pathogen-related, pPCD) [9]. The latter is often termed the hypersensitive response (HR). Despite the considerable amount of existing experimental data, the mechanism of PCD in plants remains insufficiently studied [9]. The induction of cell death in plants may depend on pathogen proteins called avirulence factors (Avrs). Virus-specific proteins occupy a special place among these. Viral effectors are variable, relatively small, and—in many cases—well-studied proteins that usually accumulate in plants to high levels. Experimental infections allow rapid testing of wild-type/mutant effectors and HR sensors [10]. During HR, cells infected by a pathogen undergo PCD, forming necrotic lesions on the plant.
2. Virus-Specific Proteins Involved in the Hypersensitive Response
Avr recognition by resistance proteins (R proteins) leads to a strong defense response. As a rule, virus-specific avirulence factors interact with the coiled-coil nucleotide-binding leucine-rich repeats (LRR) protein CC-NB-LRR (CNL) class and toll-like receptor/Interleukin (IL)-1 receptor nucleotide-binding LRR protein TIR-NB-LRR (TNL) class [6,9]. There are two paths for triggering an antiviral defense response: Avrs can bind to an R protein with or without the participation of an intermediate host factor; it is also possible for Avrs to induce the synthesis of an R gene product and allow it to signal a response. Interaction between viral and host factors leads to a rapid response [10]. Information concerning numerous viral proteins involved in triggering HR is summarized in Table 1.

Table 1.
Viral effectors inducing hypersensitive response in plants a.
2.1. Virus RdRp Proteins
2.1.1. Tobacco Mosaic Virus (TMV) p50
The HR is most well studied for TMV infection. The N-gene product in Nicotiana glutinosa and replication-associated protein p50 (helicase domain of RNA-dependent RNA polymerase, RdRp) activate ATP-dependent HR induction [11,12,13,14,15,16,17].
TMV p50 interacts indirectly with toll-like/interleukin-1 receptor-type domain protein encoded by the N gene; in the Nicotiana benthamiana, such signaling requires an additional chloroplastic rhodanese sulfurtransferase, NRIP1 (Nuclear Receptor Interacting Protein 1), that recognizes p50 and inhibits the development of infection [18].
Squamosa Promoter Binding Protein Like 6 (SPL6) transcription factor also stimulates TMV resistance [19].
N Requirement Gene partner 1 (NRG1) participates in the plant response at the next step of signaling by forming a Ca2+ channel in the plasma membrane and promoting cation influx into the cell [20,21].
2.1.2. Potyviral RdRp (NIb)
A known function of the Pepper Mottle Virus (PepMoV) nuclear inclusion protein b (NIb) is activation of HR with the participation of CNL-type proteins Pvr4 or Pvr9. The coiled-coil domain of Pvr4 might be involved in specific Avr recognition [22]. These proteins are capable of causing cell death in Capsicum annuum and Capsicum chinense. The Nib proteins of other potyviruses, pepper severe mosaic virus (PeSMV), tobacco etch virus (TEV), and potato Y virus (PVY), should also be mentioned [22,23,24,25].
2.1.3. Plantago Asiatica Mosaic Virus (PlAMV) RdRp
The replicase of PlAMV is important for development of systemic necrosis. Infection of N. benthamiana with this potexvirus causes HR, while the PlAMV lily isolate Li6, in contrast to Li1, infects plants asymptomatically [26,27].
2.1.4. Tomato Yellow Leaf Curl Virus (TYLCV) Rep/C1
Replication-associated protein Rep/C1 of begomovirus TYLCV contains the avirulence motif for binding to the Ty-2 gene product in Solanum habrochaites. Ty-2 encodes the NB-LRR protein. Interaction between Rep/C1 and Ty-2 triggers HR [28,29]. Expression of the N-terminal 210 amino acid residues of Rep/C1 in tomato activates the genes responsible for suppression of cell death [30,31].
2.1.5. Cucumber Mosaic Virus (CMV) 2a
HR induced by CMV-replication-related protein 2a and TNL RS4-4 was observed in Phaseolus vulgaris. The efficiency of this interaction does not depend on the enzymatic activity of the replicase [32].
2.2. Virus Coat Proteins (CP)
2.2.1. Potato Virus X (PVX) CP
The potato Rx gene encoding a CNL-class protein is responsible for HR during PVX infection [33]. The PVX CP is required for efficient interaction with the Rx protein, which causes necrosis in potato and local lesions in Gomphrena globosa [34]. Ran GTPase Activating protein 2 (RanGAP2) as well as transcription factor Golden-like (NbGLK1) from N. benthamiana enforce the relocalization of Rx from the cytoplasm into the nucleus [35,36,37].
2.2.2. Turnip Crinkle Virus (TCV) CP
The coat protein of tombusvirus TCV interacts with the product of the HRT gene [38]. HRT-mediated resistance in the turnip is dependent on lipase-like protein Enhanced Disease Susceptibility 1 (EDS1) and its interacting lipase cofactor Phytoalexin Deficient 4 (PAD4), which form a complex with EDS1. A NB-LRR protein accomplishes downstream signaling [39,40]. The level of EDS1 and PAD4 synthesis is increased in the presence of sialic acid (SA), including exogenous treatment. In the latter case, HRT is stimulated via PAD4 [40,41,42,43].
2.2.3. Potato Virus Y (PVY) CP
Potyvirus PVY infection of potato activates HR connected with CP PVY and Ny gene products [44,45,46,47].
2.2.4. Cucumber Mosaic Virus (CMV) CP
The RCY1 (CNL-class) gene product from Arabidopsis thaliana interacts with the CMV coat protein resulting in HR. RCY1 and HRT belong to the same HRT/RP8 family and apparently diverged via recombination. CPs of CMV and TCV are not homologous, but recognize allelic gene products [48,49].
2.3. Virus Movement Proteins (MP)
2.3.1. Tomato Mosaic Virus (ToMV) MP
Movement proteins (MP) promote cell-to-cell viral transport through plasmodesmata. Resistance genes Tm2, Tm22 were found in tomato, Solanum peruvianum, Nicotiana tabacum, and Chenopodium amaranticolor. The Tm22 gene is MP-dependent, whereas the Tm1 gene is activated by RdRp. Tm22 codes for a CNL protein, but the mechanism of HR induction is not fully understood [50,51,52,53].
2.3.2. Tobacco Rattle Virus (TRV) MP
HR in tobacco and potato can be associated with the nucleus. TRV MP (29K) is the elicitor of HR response via the EDS1/PAD4 pathway. Overexpression of the 29K gene by agroinfiltration of leaves leads to cell death [46,54].
2.3.3. Tomato Spotted Wilt Virus (TSWV) MP
R genes belonging to the Sw-5 family from Solanum peruvianum code for CNL proteins. Tospovirus TSWV MP (NSM) induces cell death in tomato and N. benthamiana [55,56,57]. The RTSW gene product from Nicotiana alata also recognizes NSM in tobacco. RTSW domains required for HR triggering are different from Sw-5b; most likely, the proteins are not orthologs [58].
2.3.4. Cauliflower Mosaic Virus (CaMV) MP
The P6 protein of pararetrovirus CaMV induces systemic PCD in Nicotiana clevelandii and HR in Nicotiana edwardsonii [59]. P6 is also responsible for the non-necrotic defense response in A. thaliana, Nicotiana bigelovii, and N. glutinosa [59,60,61]. Transgene and transient expression of P6 in Arabidopsis and N. benthamiana changes the subcellular localization of transcription factor NPR1 (nonexpressor of pathogenesis-related genes 1) [62]. Introgression of CCD1 gene from N. bigelovii can selectively block systemic and local HR induced by CaMV infection in N. clevelandii and N. edwardsonii [63].
2.4. Other Viral Proteins Involved in the HR
2.4.1. Tomato Spotted Wilt Virus (TSWV) Silencing Suppressor Protein (NSs)
The NSs protein activates resistance of pepper and C. chinense against TSWV related to CNL Tsw resistance gene from tomato codes for a protein with unusually large LRR region (over 30 repeats) [64,65]. The NSs is a viral suppressor of RNA silencing [66] and a determinant for transmission by thrips (Thysanoptera) [67]. Changes at the N-terminus of NSs are crucial for interaction with Tsw sensor [68,69].
2.4.2. Tomato Leaf Curl New Delhi Virus (ToLCNDV) Silencing Suppressor Protein AC4
R-gene SISw5a product from Solanum lycopersicum is a homolog of the Sw-5a CNL protein. SlSw5a protein binds to AC4 from ToLCNDV, which inhibits RNA interference. A mutation at the C-terminus of the AC4 protein destroys this interaction, thus preventing PCD [70,71].
2.4.3. Soybean Mosaic Virus (SMV) Proteins CI, P3 and HC-Pro
Potyvirus SMV encodes CI (cylindrical inclusion), P3 and HC-Pro (helper component proteinase) proteins that can function as Avrs interacting with NB-LRR proteins. Genetic mapping studies identified two resistance loci (Rsv and Rsc) that code for typical CNL proteins which provide resistance to SMV [72,73]. P3/HC-Pro act as Rsv1, Rsv2, and Rsv4 inductors, while CI acts as a trigger for Rsc4-3 [72,73,74,75]. The interaction between an avirulent SMV strain and a soybean resistance gene product belonging to the Rsv family induced siRNA accumulation followed by autophagy [74]. P3 cooperates with translation factors to trigger the unfolded protein response (UPR) [75,76].
3. NLR Signaling Leading to Hypersensitive Response in Plants and Formation of Resistosomes
3.1. Plant Cell Death Signal Transmission Network
Intracellular nucleotide-binding (NB) domain and LRR-containing proteins are an essential part of plant defense and innate immune systems. NB-LRR (NLR) proteins interact with effector proteins that are synthesized by pathogens. As a result, a protective reaction occurs, often involving rapid and limited cell death at the locus of infection called the hypersensitive response site. This type of immunity is termed Effector-Triggered Immunity (ETI). Plant NLR proteins are signal transduction ATPases with numerous domains (STAND superfamily) [77,78,79,80]. NLRs can be grouped into subclasses in accordance with their N-terminal domain. CNLs contain coiled-coil domain or RPW8-type (Resistance to Powdery Mildew 8) coiled-coil (CCR, RNL proteins), and TNLs contain toll/interleukin-1 receptor-type domain [80,81]. RPW8 is a membrane-associated protein from A. thaliana [81]. Typical coiled-coil domains incorporate the motif “EDVID”, while the RPW8-like coiled-coil CCR domains do not contain this motif [81]. Since in monocotyledonous plants, including cereals, NLR proteins incorporate only CC domains, one may assume that toll-like/interleukin-1 receptor-type domains were lost in the process of divergence from the common angiosperm ancestor [82]. In addition to ETI, there is another basic mechanism of plant resistance to pathogens, called pattern triggered immunity (PTI), that limits pathogen spreading but does not trigger cell death. This pathway requires participation of receptor-like kinases and/or receptor-like proteins that lack a protein kinase domain situated in the plasma membrane. There is very little evidence that PTI may be involved in viral resistance. None of the PTI receptors were shown to recognize virus particles [80].
Typical representatives of CNLs are HopZ-Activated Resistance1 (ZAR1) from Arabidopsis [83] and N. benthamiana [84] as well as Rx from potato [85]; for TNLs—N gene product from tobacco (N. tabacum) [11] and RPP1 from Arabidopsis [86]. Some NLRs act as singletons, combining functions of pathogen sensing and immune signaling, while others form a network of interactions involving at least two factors [87,88].
Activated singleton (for example, ZAR1) and helper NLR proteins oligomerize, forming circular “tunnel” structures termed resistosomes, consisting of several molecules of the corresponding protein or a complex of distinct proteins, and forming a Ca2+ channel in the plasma membrane (PM) of the cell [77,89,90,91,92]. The N-terminal helix α1 containing the MADA motif was found in about 20% of helper angiosperm CNLs capable of oligomerization [93]. In contrast, the NLR sensor proteins lack the MADA motif [93].
There are known examples in which overexpression of plant signal CC and TIR domains alone lead to the induction of HR. These include the TIR domains of factors RPS4, RPP1A, and At4g19530 in Arabidopsis [94], CCR (RPW8-like) domains of ADR1 and NRG1 proteins in Arabidopsis and N. benthamiana [95], as well as the CC domains from wheat proteins Sr33 and Sr50 in N. benthamiana [96]. The N-terminal fragment of the CC domain of factor AT1G12290 (100 aa) from Arabidopsis was sufficient to induce HR in N. benthamiana but not in Arabidopsis [97]. The CC-domain of barley protein MLA10, capable of forming a rod-shaped homodimer, was also capable of inducing cell death in N. benthamiana [98]. Most likely, a homodimer is also formed via self-association of the TIR domain from flax (Linum usitatissimum) L6 resistance protein [99]. On the contrary, the expression of the CC domain of the NLR protein Rx from potato in N. benthamiana leaves did not cause HR, unlike the expression of the NB domain [100]. For ADR1 and NRG1, it was shown that HR development caused by the expression of CCR domains occurred independently of SGT1, a cofactor of the chaperone HSP90 [95].
The NLR required for cell death (NRC) network in plants is comprised of at least two levels [101] and consists of sensor and helper proteins, forming pairs or genetically unlinked receptor networks [87,88,101,102]. For example, PVX CP triggers Rx activation [85] followed by signaling through any of the NRC2,3,4 CNL helper proteins and consequent cell death in N. benthamiana [87,103]. A similar pathway was described for the Bs2 sensor from wild pepper (C. chacoense) [104]. Sensor Rpi-blb2 from potato interacts with AVRblb2 effector from Phytophtora infestans [105] and activates NRC4, but not NRC2 [104,106]. It is important to note that the Rx sensor does not form stable complexes with either NRC2 or NRC4 and thus does not participate in their oligomerization. Activated homo-oligomers NRC2/NRC4 are localized to puncta that associate with the PM, while Rx remains in the cytosol [104]. N. benthamiana, in addition to genes encoding factors NRC2-NRC4, encodes atypical gene NRCX, defined as NLR modulator [107]. NRCX protein does not contain the usual N-terminal MADA motif and is not capable of oligomerization. When co-expressed, NRCX has a negative effect on NRC2 и NRC3, but not NRC4 [107]. Proteins belonging to the NRC network form an evolutionary superclade, estimated to be 100 million years old. In some cases, functionally related proteins are not phylogenetic relatives. In Solanaceae the NRC accounts for up to half of the NLR-ome [87]. It is assumed that, while performing the functions of pathogen perception and helper activation, in the course of evolution, sensor CNLs have lost the ability to oligomerize [104].
RNL proteins function downstream of pathogen detection proteins. For the effective manifestation of toll-like/interleukin-1 receptor-type signaling in plants lipase-like protein EDS1, other helper RNL proteins as well as Activated Disease Resistance gene 1 (ADR1) and NRG1 are needed [108,109]. Both proteins represent two subgroups of the RNL family. In Arabidopsis, NRG1 is required for cell death and ADR1 for transcriptional reprogramming of resistance genes [110,111,112]. In N. benthamiana, toll-like/interleukin-1 receptor-type (TIR) signals are transmitted primarily by NRG1 [113,114,115]. Two other lipases participating in this pathway are PAD4 and Senescence-Associated Gene 101 (SAG101). Either PAD4 or SAG101 forms functional heterodimeric subunits with EDS1 [116,117]. ADR1-EDS1-PAD4 module provides transcriptional defenses that restrict pathogen growth [118,119], whereas the NRG1-EDS1-SAG101 module is involved in both cell death and transcriptional regulation in N. benthamiana [112].
NLR-ID proteins contain the so-called integrated domain (ID), for example in the TNL pair RRS1/RPS4 from Arabidopsis, the sensor associates with a helper. The WRKY (defined by the conserved WRKYGQK motif and a zinc-finger region) transcription factor ID domain of RRS1 protein is located at the C-terminus. Interaction of the ID with the adjacent domain 4 (DOM4) retains the protein in an inactive form. Bacterial effector AvrRps4 from Pseudomonas syringae disrupts this interaction, leading to derepression of RRS1. NLR-IDs act in combination with the canonical NLR protein, which performs a helper function, inducing cell death [120]. Such pairs may also contain a CC domain instead of TIR, for example the pair of rice proteins RGA4/RGA5 [121].
Some NLR proteins can cause unwanted events in plants known as hybrid incompatibility, which is apparently due to the fact that incorrect interaction of sensors and helpers can cause an unintentional defense response [122]. For example, ADR1 and NRG1 proteins may recognize various “wrong” sensors, which increases the risk of autoimmune reactions [123]. The mechanisms that prevent aberrant TIR/TNL activity were reviewed recently [109]. To date, NLR genes are not classified as “lethal phenotype genes” necessary for the survival and development of plants [124].
3.2. Resistosomes in Plants
NLR proteins that combine the functions of sensors and helpers/executors are termed singleton NLRs. The mechanism of action of some of them is quite well studied. For example, according to cryo-electron microscopy, CNL protein ZAR1 forms a wheel-like activated pentameric membrane-associated complex termed the resistosome, which is an α-helical barrel interacting with the LRR and winged-helix domains [89,90]. In an inactive state, ZAR1 is precomplexed with the pseudokinase RKS1 (resistance-related kinase 1); during activation, the effector AvrAC from Xanthomonas campestris uridylates decoy kinase PBL2 (probable serine/threonine-protein kinase 2), which associates with the complex ZAR1-RKS1, causing its structural rearrangement and release of ADP from nucleotide-binding domain with subsequent oligomerization of the heterotrimer and its movement from the cytosol into the PM [90]. Structurally, the ZAR1 resistosome resembles the metazoan apoptosome APAF1 (apoptotic protease activating factor 1) [125,126,127] and inflammasome NAIP2-NLRC4 (NLR family apoptosis inhibitory protein 2—NLR family, CARD domain containing 4) [128,129,130]. Experiments involving in vitro reconstitution of ZAR1 resistosome in planar lipid bilayers showed that it is a calcium ion channel, which triggers Ca2+ influx in Arabidopsis protoplasts; it can be localized to the PM but not to the endoplasmic reticulum [131]. It was reported that activation of ZAR1 leads to perturbation of chloroplasts, abrupt disintegration of nuclei, and cell collapse [131]. CNL protein Sr35 from monocotyledonous wheat interacts with effector AvrSr35 of the wheat stem rust pathogen (fungus Puccinia graminis f. sp. tritici, Pgt). Activation of Sr35 occurs when its LRR domain interacts with AvrSr35. Furthermore, the Sr35-AvrSr35 complex oligomerizes, binding ATP, and forms a pentameric structure similar to “dicotyledonous” resistosome ZAR1, despite only 28.4% sequence conservation [92]. Complex Sr35-AvrSr35 was functional in N. benthamiana, inducing cell death. The Sr35 resistosome also exhibits Ca2+ channel activity in oocytes (Xenopus laevis) [92].
The putative pentameric resistosome NRG1 is formed from heterotrimeric subunits consisting of a helper RNL protein, interacting with cofactor lipases EDS1 и SAG101 [112]. Autoactive mutant NRG1 from Arabidopsis becomes localized to the PM and forms Ca2+-permeable cation channels [20]. Sensor TNL together with β-estradiol promotes oligomerization of NRG1-EDS1-SAG101 complex and directs it to the PM and nucleus [112].
The resistosome of another resistance factor from Arabidopsis—TIR-containing protein RPP1, is tetrameric. Effector ATR1 from the oomycete pathogen Hyaloperonospora arabidopsidis activates RPP1 through LRR domain [91]. Unlike ZAR1, the RPP1 protein in the resistosome remains bound to ADP; the effector ATR1 retains interaction with LRR. This resistosome exhibits enzymatic activity and hydrolyzes nicotinamide adenine dinucleotide (NAD+). Resistosome RPP1 is a holoenzyme formed by two asymmetric homodimers, creating active sites for NAD+ hydrolysis. NAD+ glycohydrolase (NADase) activity requires the proximity of two TIR domains and is enhanced in the presence of polyethyleneglycol, Mg2+ ions (10 mM), and, to a lesser extent, Ca2+ [91,132]. TNL protein ROQ1 from N. benthamiana also forms a tetrameric resistosome in complex with the XopQ effector (Xanthomonas outer protein Q) from Xanthomonas euvesicatoria. The LRR domain has a horseshoe shape and wraps around XopQ. The nucleotide-binding domains of the oligomers of the resistor-activated ROQ1 interact with ATP and a Mg2+ ion. NADase activity was suggested, but not experimentally proven. The structure of the putative active center was similar to the resistosome RPP1 [77].
3.3. Metazoan Apoptosomes/Inflammasomes
Central nucleotide-binding domain (NOD, nucleotide-binding oligomerization domain), phylogenetically related to plant NB-ARC, was found in such proteins as human APAF-1 and its homolog CED-4 from Caenorhabditis elegans [133,134]. Unlike plant NLR proteins, the C-terminal region of APAF-1 contains WD40 repeats, also known as WD (terminating in a tryptophan-aspartic acid) or beta-transducin repeats. According to phylogenetic analysis, the domain NOD of APAF-1 is more similar to plant nucleotide-binding domains than to the NOD domains of other mammals [129,135]. The N-terminal region of APAF-1, like many other metazoan NLR proteins, contains a CARD domain (caspase recruitment domain). Often, a Pyrin (PYD, also known as PAAD/DAPIN) domain is found instead of CARD. Inflammasome NAIP2-NLRC4 and apoptosome APAF-1 activate caspase-1 and caspase-9, respectively [130,136,137]. CC domains are well-defined and form an α-helical barrel but largely buried in the tunnel-shaped structure of resistosome, whereas CARD domains are completely disordered in the absence of interacting partners and remain flexible in the oligomeric structure [89,90]. The TIR domains of the RPP1 resistosome also form a stable tunnel structure [91]. Structural differences between plant resistosomes, NAIP2-NLRC4 inflammasome and APAF-1 apoptosome suggest that transmission of the cell death signal using NLR proteins in plants and animals may differ significantly, primarily due to the action of the procaspase binding domain CARD. Perhaps, the ZAR1 resistosome affects the permeability of the PM or perturbs its integrity [89,90]. The NLRC4 inflammasome activates the protein Gasdermin D (GSDMD) via caspase-1, forming membrane pores and mediating a type of programmed cell death known as pyroptosis [138]. In contrast, the RPP1 resistosome acts as a holoenzyme for NAD+ hydrolysis [91], resembling the mechanism of action of metazoan CARD-containing holoenzymes responsible for recruitment of procaspases. It should be noted that the TIR domains of other plant NLR proteins are also capable of NADase activity, as well as some TIR-containing animal proteins devoid of NOD and C-terminal WD40/LRR domains [132,139]. NAD cleavage by TIR domains leads to the release of adenosine diphosphate ribose from NB-ARC domains, which, in turn, activates Ca2+ influx to the cytosol, subsequently activating the HR [77,92]. In addition, it is assumed that NADase activity of TNL resistosomes can produce cyclic-ADP-Ribose (vcADPR) and activate the formation of “downstream” RNL resistosomes, which, in turn, cause cell death via Ca2+ influx [112]. Interestingly, expression of wheat CNL Sr35 together with its pathogen effector AvrSr35 in Sf21 insect cells induced cell death in the absence of other plant proteins [92].
SARM-1 (sterile alpha and TIR motif containing 1) protein induces neuronal cell death and axon degeneration related to NADase activity of the protein dimer [140]. In addition to TIR, it contains other domains, including tandem sterile alpha motifs (SAM) responsible for its self-association. Plant and animal TIR domains are phylogenetically distant but structurally similar as demonstrated by crystallization of human octameric TIR from SARM-1 [132]. In vitro experiments show that animal TIR domains hydrolyze NAD+ much more rapidly than those of plants [132]. Surprisingly, expression of either human TIR domain [132] or an autoactive fragment of SARM-1 [139] in N. benthamiana caused HR [132,139], even in EDS1 knockout plants [132], which indicates a mechanism different than that of typical plant TNL signaling [132]. A fusion protein consisting of the human SAM domain responsible for oligomerization and the plant RPS4 TIR domain was functional and induced HR in N. tabacum [139].
The TIR domain of RPS4 from Arabidopsis was successfully fused to an animal NLRC4 protein incorporating an LRR domain, which, in particular, distinguishes it from WD-containing APAF-1. Expression in N. benthamiana and N. tabacum, together with combinations of assistant genes (NAIP5/flagellin FlaA from Legionella pneumophila or NAIP2/rod protein PrgJ from Salmonella typhimurium), led to the formation of stable immunoprecipitable “plant inflammasomes” capable of inducing HR [141].
A proton pump formed by bacterio-opsin (bO) and located in the plasma membrane of transgenic N. tabacum cells activates HR, manifested as a “lesion mimic phenotype” [142]. Protein HrpZ1 from plant pathogen P. syringae generates membrane-associated pore structures in parsley (Petroselinum crispum) cell suspensions and activates MAPK cascades, which are involved in PCD signaling [143].
4. Common Features of Antiviral Resistance across the Kingdoms
Virus-induced ETI in animals is insufficiently studied. The NKG2D receptor of natural killer cells interacts with viral effectors, which leads to cell activation. One of the most striking examples is the mouse cytomegalovirus, encoding the m18 effector, which affects the deacetylation of histones in host cells. Viral infections can affect the efficiency of protein synthesis in animals, which might induce ETI as well. For example, protein factors BCL-xL and MCL-1 triggered pyroptosis associated with gasdermin E in virus-infected barrier epithelial cells [144].
Other animal pathogens may interact with NLRs either directly or indirectly. Mouse NAIP proteins can serve as examples of the first option. NAIP5 binds the bacterial protein flagellin, whereas NAIP2 binds the inner rod component from type III secretion system (T3SS). In turn, both proteins are able to activate the NLRC4 inflammasome and remain members of the heterooligomeric complex as single subunits. The HD1 and HD2 motifs of the nucleotide-binding domain NACHT (found in NAIP, CIITA, HET-E, TP1, hence the name) play a key role in such recognition, and the flanking BIR1 and LRR domains are auxiliary [145]. Mutual replacement of NACHT domains in NAIP chimeric proteins changes the specificity of recognition of their corresponding partners. Synthetic recombinant protein NAIP 2/5 provides recognition of both bacterial Avrs [146]. A direct mechanism of intracellular perception between Avrs and sensors in plants, in particular, may imply interaction of the effector with the LRR domain (ATR1 and RPP1) [147] or C-JID domain (C-terminal jelly roll/Ig-like) located downstream of LRR (XopQ and ROQ1) [148]. One can also mention recognition of PVX CP by the C-terminal LRR domain of Rx from potato.
In addition to the well-studied example of ZAR1, involved in indirect ETI recognition in plants (Section 3.2) [89,90], RIN4 (RPM1-interacting protein 4) “guardee” protein from Arabidopsis forms complexes with NLR guard proteins RPS2 and RPM1. If the protein recognized by the effector has no other physiological function, it is called a “decoy” [78]. Protease AvrRpt2 from P. syringae cleaves RIN4 and releases the guard [149]. The AvrRpm1 effector performs (ADP)-ribosylation of RIN4, which is subsequently phosphorylated [150]. Indirect recognition in animals can be represented by NLRP1b protein with noncanonical organization of domains—the C terminal CARD is separated from LRR by the so-called function-to-find domain (FIIND). Protease LF (Lethal Factor) from Bacillus anthracis cleaves off the C-terminus of NLRP1b capable of oligomerization and caspase recruitment [78,151]. Such a model is often defined as “recognition with integrated domains (ID)” [78]. Plant TNL guardee RRS1 interacts with the TNL helper/executor RPS4 and inhibits its autoactivity (Section 3.1, Figure 1). RRS1 interacts, in particular, with the PopP2 effector from Ralstonia solanacearum via C-terminal ID domain WRKY, which leads to its acetylation [78,152].
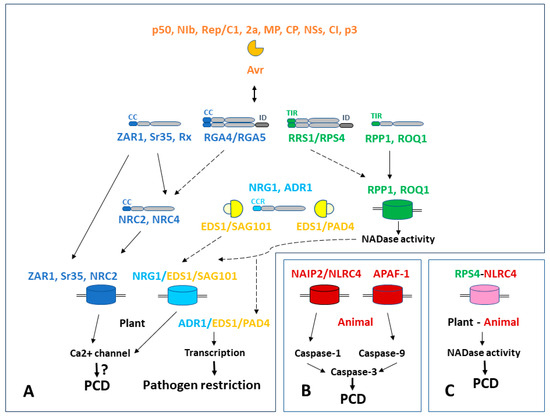
Figure 1.
Schematic representation of plant NLR signaling and resistosome formation alongside the animal apoptosome and inflammasome, and the main programmed cell death (PCD) pathways. (A) Plant signaling network. CNL, RNL, TNL pathways are shown in blue, pale blue, and green, respectively. Avr effectors are displayed in orange and helper lipases are displayed in yellow. (B) Animal apoptosome (APAF-1) and inflammasome (NAIP2/NLRC4) are shown in red. (C) Chimeric “plant inflammasome” generated with fusion protein consisting of plant TIR domain and animal NLRC4 subunit is shown in purple. Dotted lines demonstrate the intended pathways.
Recently, striking similarities between effector-triggered PCD pathways in plants and animals have been found. Foremostly, these concern the formation of similar membrane-associated structures, playing a key role in the induction of HR in plants and apoptosis/pyroptosis in animal cells. Such structures are called “resistosomes” [83,84,86]. Currently known plant resistosomes allow Ca2+ influx into the cytosol or demonstrate NADase activity [89,91].
Viral effectors can interact with NLR sensors via various mechanisms, changing their conformation and thereby activating them. Avr/NLR sensor interaction can be mediated by and involve several assistant proteins, which, as a rule, subsequently retain interaction with the NLR protein undergoing oligomerization [89,90]. The NAIP2/NLRC4 inflammasome, like many plant resistosomes, contains auxiliary proteins [128,129,130]. “Assistant” and “helper” proteins should be distinguished; the former are connected to the periphery of the resistosome, while the latter are a core part. In addition, singleton CC-containing proteins are able to form resistosomes “unaided”, whereas the sensor CNLs transmit the signal to the next level and helper CNLs form resistosomes [89,90,92,104]. Both CNLs and TNLs can form “sensor pairs”, while the subsequent steps of signal transmission remain unclear [101]. For comparison, apoptosome APAF-1 formation requires the participation of cytochrome C and dATP, and the NAIP2/NLRC4 inflammasome requires involvement of pathogen-associated molecular patterns (PAMPs) [125,126,127,128,129,130]. TNL signaling is indirect. Oligomeric TNL resistosomes display NADase activity and activate helper RNL proteins with assistant lipase-like protein EDS1 in complex with either SAG101 or PAD4 lipases. To date, it is assumed that RNL NRG1 forms resistosomes that allow Ca2+ influx, while RNL ADR1 acts as a transcriptional regulator [112]. Interestingly, induction of cell death in plants using animal TIR domains dispenses with EDS1 assistance [132].
At the level of the PM (Figure 1), the main similarity between plant resistosomes and apoptosomes/inflammasomes of animals is primarily structural. Nevertheless, it is necessary to note the phylogenetic relationship between the nucleotide-binding plant domain NB-ARC and the animal domain NOD of APAF-1 [135]. TIR domains of plants and animals have no significant homology at the amino acid level, but are functionally interchangeable. For example, the chimeric “plant inflammasome” that includes the plant TIR domain together with a “complete” animal NLRC4 protein demonstrated NADase activity followed by HR induction. For its formation in plant cells, as well as in animal cells, auxiliary NAIP and PAMP proteins were needed [141]. From the point of view of modular organization of cell death pathways, inflammasome-inducing pathogen signal PAMP resembles the Sr35 singleton resistosome [92]. A fusion protein consisting of human oligomerization domain SAM and plant TIR was functional and provided activation of the hypersensitive response in plants [139]. CC and CCR domains are present only in plants. On the contrary, the CARD domain, which provides activation of procaspases, is absent in plant NLR proteins. The mechanism of activation of plant metacaspases and caspase-like proteins involving membrane Ca2+ channels need to be studied in more detail (Figure 1).
Plants and animals share proteins containing NBD and LRR. It is believed that these proteins evolved from a common prokaryotic ancestral ATPase. NBDs are highly conserved but, based on phylogenetic analysis, it is generally accepted that they diverged early from the NB-ARC (plants, partly animals) and NACHT (animals) lineages [78,135]. Fungi also possess both types of NBD but lack LRRs [153]. Either NB-ARC or NACHT domains probably served as “assembly centers”, which developed independently from each other, collecting sets of N- and C-terminal functional domains that differ significantly in plants and animals [78,154]. In both kingdoms, the system of triggering and NLR signaling for PCD pathways is very complex and flexible. Different types of Avr recognition, various guardee, decoy, helper, and assistant proteins together with homo- and heterooligomeric membrane protein complexes present a wide field for research. The structural features of NLRs allow obtaining functional fluorescent fusion proteins or separate domains. Additional domains such as animal BIR or plant integrated domains involved in Avr recognition as well as chimeric proteins including TIR [139,141] and NACHT [146] domains look like promising targets for NLR engineering. In our view, the number of similar or interchangeable elements in the organization of plant and animal innate immune systems seems sufficient to raise the question posed in the title of this review—is there a common mechanism of antiviral resistance conserved across kingdoms? The answer can only be obtained experimentally.
5. Conclusions
The development of PCD in plants and Metazoa differs significantly. However, recent structural studies of plant resistosomes have revealed many similarities with animal apoptosomes and inflammasomes. It should be recognized that there are obvious blind spots in our understanding of the relationship between the formation and functioning of resistosomes, the presence of caspase-like activity in plants, and the development of the HR. Nevertheless, it is necessary to mention that the progress made to date is considerable. The relative simplicity and diversity of virus-specific effectors of the hypersensitive response makes them easier to study in comparison with effectors of fungal and bacterial pathogens. In addition to the transient expression of effector genes using agroinfiltration, it is possible to quickly obtain stable transgenes (Arabidopsis) and infect plants with natural viruses or viral vectors. Special attention should be paid to the data on the induction of HR during the expression of individual domains of plant NLR proteins. Further study of such effects may have practical significance for agriculture and medicine.
6. Future Directions
The multi-domain organization of NLR signaling proteins in plants and their functional homologues in Metazoa, as well as the variety of corresponding pathways and a “modular” structure of interaction between effector, sensor, and helper proteins allows for the envisioning of various “cross-kingdom” studies. In this regard, we can mention modeling of cell death pathways in plants and animals, creation of chimeric plant/animal NLR proteins, and assembly of resistosome-like structures in vitro (in planar lipid bilayers) or in vivo in plant and animal cells, including heterogeneous plant/animal subunits.
Author Contributions
Conceptualization, P.A.I., T.V.G. and A.A.Z.J.; writing—original draft preparation, P.A.I., T.V.G. and M.N.R.; writing—review and editing, P.A.I., T.V.G. and A.A.Z.J.; visualization P.A.I., T.V.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Interdisciplinary Scientific and Educational School of Moscow University “Molecular Technologies of the Living Systems and Synthetic Biology”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kesavardhana, S.; Malireddi, R.K.S.; Kanneganti, T.D. Caspases in Cell Death, Inflammation, and Pyroptosis. Annu. Rev. Immunol. 2020, 38, 567–595. [Google Scholar] [CrossRef] [PubMed]
- Distéfano, A.M.; Martin, M.V.; Córdoba, J.P.; Bellido, A.M.; D’Ippólito, S.; Colman, S.L.; Soto, D.; Roldán, J.A.; Bartoli, C.G.; Zabaleta, E.J.; et al. Heat stress induces ferroptosis-like cell death in plants. J. Cell Biol. 2017, 216, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shen, Y.; Chen, C.; Sui, X.; Yang, J.; Wang, L.; Zhou, J. The crosstalk between autophagy and ferroptosis: What can we learn to target drug resistance in cancer? Cancer Biol. Med. 2019, 16, 630–646. [Google Scholar] [CrossRef] [PubMed]
- Distéfano, A.M.; López, G.A.; Setzes, N.; Marchetti, F.; Cainzos, M.; Cascallares, M.; Zabaleta, E.; Pagnussat, G.C. Ferroptosis in plants: Triggers, proposed mechanisms, and the role of iron in modulating cell death. J. Exp. Bot. 2021, 72, 2125–2135. [Google Scholar] [CrossRef]
- Wang, P.; Wang, T.; Han, J.; Li, M.; Zhao, Y.; Su, T.; Ma, C. Plant Autophagy: An Intricate Process Controlled by Various Signaling Pathways. Front. Plant Sci. 2021, 12, 754982. [Google Scholar] [CrossRef]
- Salguero-Linares, J.; Coll, N.S. Plant proteases in the control of the hypersensitive response. J. Exp. Bot. 2019, 70, 2087–2095. [Google Scholar] [CrossRef]
- Balakireva, A.V.; Zamyatnin, A.A., Jr. Cutting Out the Gaps Between Proteases and Programmed Cell Death. Front. Plant Sci. 2019, 10, 704. [Google Scholar] [CrossRef]
- Huh, S.U. Evolutionary Diversity and Function of Metacaspases in Plants: Similar to but Not Caspases. Int. J. Mol. Sci. 2022, 23, 4588. [Google Scholar] [CrossRef]
- Ebeed, H.T.; El-Helely, A.A. Programmed Cell Death in Plants: Insights into Developmental and Stress-Induced Cell Death. Curr. Protein Pept. Sci. 2021, 22, 873–889. [Google Scholar] [CrossRef]
- Huang, C. From Player to Pawn: Viral Avirulence Factors Involved in Plant Immunity. Viruses 2021, 13, 688. [Google Scholar] [CrossRef]
- Erickson, F.L.; Holzberg, S.; Calderon-Urrea, A.; Handley, V.; Axtell, M.; Corr, C.; Baker, B. The helicase domain of the TMV replicase proteins induces the N-mediated defence response in tobacco. Plant J. 1999, 18, 67–75. [Google Scholar] [CrossRef]
- Kang, B.C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Annu. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Czymmek, K.; Tallóczy, Z.; Levine, B.; Dinesh-Kumar, S.P. Autophagy regulates programmed cell death during the plant innate immune response. Cell 2005, 121, 567–577. [Google Scholar] [CrossRef]
- Ueda, H.; Yamaguchi, Y.; Sano, H. Direct interaction between the tobacco mosaic virus helicase domain and the ATP-bound resistance protein, N factor during the hypersensitive response in tobacco plants. Plant Mol. Biol. 2006, 61, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Solovieva, A.D.; Frolova, O.Y.; Solovyev, A.G.; Morozov, S.Y.; Zamyatnin, A.A., Jr. Effect of mitochondria-targeted antioxidant SkQ1 on programmed cell death induced by viral proteins in tobacco plants. Biochemistry 2013, 78, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Hong, Q.; Li, Y.; Li, Q.; Wang, M. Autophagy contributes to regulate the ROS levels and PCD progress in TMV-infected tomatoes. Plant Sci. 2018, 269, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Konakalla, N.C.; Nitin, M.; Kaldis, A.; Masarapu, H.; Carpentier, S.; Voloudakis, A. dsRNA Molecules from the Tobacco Mosaic Virus p126 Gene Counteract TMV-Induced Proteome Changes at an Early Stage of Infection. Front. Plant Sci. 2021, 12, 663707. [Google Scholar] [CrossRef]
- Caplan, J.L.; Mamillapalli, P.; Burch-Smith, T.M.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplastic protein NRIP1 mediates innate immune receptor recognition of a viral effector. Cell 2008, 132, 449–462. [Google Scholar] [CrossRef]
- Padmanabhan, M.S.; Ma, S.; Burch-Smith, T.M.; Czymmek, K.; Huijser, P.; Dinesh-Kumar, S.P. Novel positive regulatory role for the SPL6 transcription factor in the N TIR-NB-LRR receptor-mediated plant innate immunity. PLoS Pathog. 2013, 9, e1003235. [Google Scholar] [CrossRef]
- Jacob, P.; Kim, N.H.; Wu, F.; El-Kasmi, F.; Chi, Y.; Walton, W.G.; Furzer, O.J.; Lietzan, A.D.; Sunil, S.; Kempthorn, K.; et al. Plant “helper” immune receptors are Ca2+-permeable nonselective cation channels. Science 2021, 373, 420–425. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.; Hong, Q.; Lin, Y.; Mao, W.; Zhou, S. Reactive oxygen species triggering systemic programmed cell death process via elevation of nuclear calcium ion level in tomatoes resisting tobacco mosaic virus. Plant Sci. 2018, 270, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.B.; Lee, H.Y.; Seo, S.; Lee, J.H.; Choi, D. RNA-dependent RNA polymerase (NIb) of the potyviruses is an avirulence factor for the broad-spectrum resistance gene Pvr4 in Capsicum annuum cv. CM334. PLoS ONE 2015, 10, e0119639. [Google Scholar] [CrossRef]
- Tran, P.T.; Choi, H.; Choi, D.; Kim, K.H. Molecular characterization of Pvr9 that confers a hypersensitive response to Pepper mottle virus (a potyvirus) in Nicotiana benthamiana. Virology 2015, 481, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Yu, J.; Kim, K.H. Pepper Mottle Virus and Its Host Interactions: Current State of Knowledge. Viruses 2021, 13, 1930. [Google Scholar] [CrossRef]
- Gong, Y.N.; Tang, R.Q.; Zhang, Y.; Peng, J.; Xian, O.; Zhang, Z.H.; Zhang, S.B.; Zhang, D.Y.; Liu, H.; Luo, X.W.; et al. The NIa-Protease Protein Encoded by the Pepper Mottle Virus Is a Pathogenicity Determinant and Releases DNA Methylation of Nicotiana benthamiana. Front Microbiol. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Ozeki, J.; Takahashi, S.; Komatsu, K.; Kagiwada, S.; Yamashita, K.; Mori, T.; Hirata, H.; Yamaji, Y.; Ugaki, M.; Namba, S. A single amino acid in the RNA-dependent RNA polymerase of Plantago asiatica mosaic virus contributes to systemic necrosis. Arch. Virol. 2006, 151, 2067–2075. [Google Scholar] [CrossRef]
- Komatsu, K.; Hammond, J. Plantago asiatica mosaic virus: An emerging plant virus causing necrosis in lilies and a new model RNA virus for molecular research. Mol. Plant Pathol. 2022, 23, 1401–1414. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Ohnishi, J.; Saito, A.; Ohyama, A.; Nunome, T.; Miyatake, K.; Fukuoka, H. An NB-LRR gene, TYNBS1, is responsible for resistance mediated by the Ty-2 Begomovirus resistance locus of tomato. Theor. Appl. Genet. 2018, 131, 1345–1362. [Google Scholar] [CrossRef]
- Shen, X.; Yan, Z.; Wang, X.; Wang, Y.; Arens, M.; Du, Y.; Visser, R.G.F.; Kormelink, R.; Bai, Y.; Wolters, A.A. The NLR Protein Encoded by the Resistance Gene Ty-2 Is Triggered by the Replication-Associated Protein Rep/C1 of Tomato Yellow Leaf Curl Virus. Front. Plant Sci. 2020, 11, 545306. [Google Scholar] [CrossRef]
- Lucioli, A.; Berardi, A.; Gatti, F.; Tavazza, R.; Pizzichini, D.; Tavazza, M. Tomato yellow leaf curl Sardinia virus-resistant tomato plants expressing the multifunctional N-terminal domain of the replication-associated protein show transcriptional changes resembling stress-related responses. Mol. Plant Pathol. 2014, 15, 31–43. [Google Scholar] [CrossRef]
- Lucioli, A.; Perla, C.; Berardi, A.; Gatti, F.; Spanò, L.; Tavazza, M. Transcriptomics of tomato plants infected with TYLCSV or expressing the central TYLCSV Rep protein domain uncover changes impacting pathogen response and senescence. Plant Physiol. Biochem. 2016, 103, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.S.; Rojas, M.R.; Lee, J.Y.; Lee, S.W.; Jeon, J.S.; Ronald, P.; Lucas, W.J.; Gilbertson, R.L. A viral resistance gene from common bean functions across plant families and is up-regulated in a non-virus-specific manner. Proc. Natl. Acad. Sci. USA 2006, 103, 11856–11861. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, A.; Köhn, B.A.; Dedi, C.; Baulcombe, D.C. The coat protein of potato virus X is a strain-specific elicitor of Rx1-mediated virus resistance in potato. Plant J. 1995, 8, 933–941. [Google Scholar] [CrossRef]
- Goulden, M.G.; Baulcombe, D.C. Functionally Homologous Host Components Recognize Potato Virus X in Gomphrena globosa and Potato. Plant Cell 1993, 5, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–792. [Google Scholar] [CrossRef]
- Tameling, W.I.; Baulcombe, D.C. Physical association of the NB-LRR resistance protein Rx with a Ran GTPase-activating protein is required for extreme resistance to Potato virus X. Plant Cell 2007, 19, 1682–1694. [Google Scholar] [CrossRef]
- Walling, L.L. Extreme resistance: The GLK-Rx1 alliance. J. Biol. Chem. 2018, 293, 3234–3235. [Google Scholar] [CrossRef]
- Zhao, Y.; DelGrosso, L.; Yigit, E.; Dempsey, D.A.; Klessig, D.F.; Wobbe, K.K. The amino terminus of the coat protein of Turnip crinkle virus is the AVR factor recognized by resistant arabidopsis. Mol. Plant Microbe Interact. 2000, 13, 1015–1018. [Google Scholar] [CrossRef]
- Cooley, M.B.; Pathirana, S.; Wu, H.J.; Kachroo, P.; Klessig, D.F. Members of the Arabidopsis HRT/RPP8 family of resistance genes confer resistance to both viral and oomycete pathogens. Plant Cell 2000, 12, 663–676. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.C.; Navarre, D.; Kachroo, A.; Kang, H.G.; Klessig, D.; Kachroo, P. Signaling requirements and role of salicylic acid in HRT- and rrt-mediated resistance to turnip crinkle virus in Arabidopsis. Plant J. 2004, 40, 47–59. [Google Scholar] [CrossRef]
- Falk, A.; Feys, B.J.; Frost, L.N.; Jones, J.D.; Daniels, M.J.; Parker, J.E. EDS1, an essential component of R gene-mediated disease resistance in Arabidopsis has homology to eukaryotic lipases. Proc. Natl. Acad. Sci. USA 1999, 96, 3292–3297. [Google Scholar] [CrossRef] [PubMed]
- Jirage, D.; Tootle, T.L.; Reuber, T.L.; Frost, L.N.; Feys, B.J.; Parker, J.E.; Ausubel, F.M.; Glazebrook, J. Arabidopsis thaliana PAD4 encodes a lipase-like gene that is important for salicylic acid signaling. Proc. Natl. Acad. Sci. USA 1999, 96, 13583–13588. [Google Scholar] [CrossRef] [PubMed]
- Feys, B.J.; Moisan, L.J.; Newman, M.A.; Parker, J.E. Direct interaction between the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4. EMBO J. 2001, 20, 5400–5411. [Google Scholar] [CrossRef] [PubMed]
- Valkonen, J.P. Elucidation of virus-host interactions to enhance resistance breeding for control of virus diseases in potato. Breed. Sci. 2015, 65, 69–76. [Google Scholar] [CrossRef]
- Grech-Baran, M.; Witek, K.; Szajko, K.; Witek, A.I.; Morgiewicz, K.; Wasilewicz-Flis, I.; Jakuczun, H.; Marczewski, W.; Jones, J.D.G.; Hennig, J. Extreme resistance to Potato virus Y in potato carrying the Rysto gene is mediated by a TIR-NLR immune receptor. Plant Biotechnol. J. 2020, 18, 655–667. [Google Scholar] [CrossRef]
- Ross, B.T.; Zidack, N.K.; Flenniken, M.L. Extreme Resistance to Viruses in Potato and Soybean. Front. Plant Sci. 2021, 12, 658981. [Google Scholar] [CrossRef]
- Baebler, Š.; Coll, A.; Gruden, K. Plant Molecular Responses to Potato Virus Y: A Continuum of Outcomes from Sensitivity and Tolerance to Resistance. Viruses 2020, 12, 217. [Google Scholar] [CrossRef]
- Takahashi, H.; Suzuki, M.; Natsuaki, K.; Shigyo, T.; Hino, K.; Teraoka, T.; Hosokawa, D.; Ehara, Y. Mapping the virus and host genes involved in the resistance response in cucumber mosaic virus-Infected Arabidopsis thaliana. Plant Cell Physiol. 2001, 42, 340–347. [Google Scholar] [CrossRef]
- Takahashi, H.; Miller, J.; Nozaki, Y.; Takeda, M.; Shah, J.; Hase, S.; Ikegami, M.; Ehara, Y.; Dinesh-Kumar, S.P. RCY1, an Arabidopsis thaliana RPP8/HRT family resistance gene, conferring resistance to cucumber mosaic virus requires salicylic acid, ethylene and a novel signal transduction mechanism. Plant J. 2002, 32, 655–667. [Google Scholar] [CrossRef]
- Lanfermeijer, F.C.; Dijkhuis, J.; Sturre, M.J.; de Haan, P.; Hille, J. Cloning and characterization of the durable tomato mosaic virus resistance gene Tm-22 from Lycopersicon esculentum. Plant Mol. Biol. 2003, 52, 1037–1049. [Google Scholar] [CrossRef]
- Chen, T.; Liu, D.; Niu, X.; Wang, J.; Qian, L.; Han, L.; Liu, N.; Zhao, J.; Hong, Y.; Liu, Y. Antiviral Resistance Protein Tm-22 Functions on the Plasma Membrane. Plant Physiol. 2017, 173, 2399–2410. [Google Scholar] [CrossRef]
- Ishibashi, K.; Kezuka, Y.; Kobayashi, C.; Kato, M.; Inoue, T.; Nonaka, T.; Ishikawa, M.; Matsumura, H.; Katoh, E. Structural basis for the recognition-evasion arms race between Tomato mosaic virus and the resistance gene Tm-1. Proc. Natl. Acad. Sci. USA 2014, 111, E3486–E3495. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.Y.; Ma, H.Y.; Wang, L.; Tettey, C.; Zhao, M.S.; Geng, C.; Tian, Y.P.; Li, X.D. Identification of genetic determinants of tomato brown rugose fruit virus that enable infection of plants harbouring the Tm-22 resistance gene. Mol. Plant Pathol. 2021, 22, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Ghazala, W.; Varrelmann, M. Tobacco rattle virus 29K movement protein is the elicitor of extreme and hypersensitive-like resistance in two cultivars of Solanum tuberosum. Mol. Plant Microbe Interact. 2007, 20, 1396–1405. [Google Scholar] [CrossRef]
- Peiró, A.; Cañizares, M.C.; Rubio, L.; López, C.; Moriones, E.; Aramburu, J.; Sánchez-Navarro, J. The movement protein (NSm) of Tomato spotted wilt virus is the avirulence determinant in the tomato Sw-5 gene-based resistance. Mol. Plant Pathol. 2014, 15, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Hallwass, M.; de Oliveira, A.S.; de Campos Dianese, E.; Lohuis, D.; Boiteux, L.S.; Inoue-Nagata, A.K.; Resende, R.O.; Kormelink, R. The Tomato spotted wilt virus cell-to-cell movement protein (NSM) triggers a hypersensitive response in Sw-5-containing resistant tomato lines and in Nicotiana benthamiana transformed with the functional Sw-5b resistance gene copy. Mol. Plant Pathol. 2014, 15, 871–880. [Google Scholar] [CrossRef]
- Zhu, M.; Jiang, L.; Bai, B.; Zhao, W.; Chen, X.; Li, J.; Liu, Y.; Chen, Z.; Wang, B.; Wang, C.; et al. The intracellular immune receptor Sw-5b confers broad-spectrum resistance to Tospoviruses through recognition of a conserved 21-amino acid viral effector epitope. Plant Cell 2017, 29, 2214–2232. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Liu, Y.; Yu, H.; Yuan, C.; Zeng, J.; Zhao, L.; Tong, Z.; Tao, X. Non-Structural Protein NSm of Tomato Spotted Wilt Virus Is an Avirulence Factor Recognized by Resistance Genes of Tobacco and Tomato via Different Elicitor Active Sites. Viruses 2018, 10, 660. [Google Scholar] [CrossRef]
- Palanichelvam, K.; Cole, A.B.; Shababi, M.; Schoelz, J.E. Agroinfiltration of Cauliflower mosaic virus gene VI elicits hypersensitive response in Nicotiana species. Mol. Plant Microbe Interact. 2000, 13, 1275–1279. [Google Scholar] [CrossRef]
- Cole, A.B.; Király, L.; Ross, K.; Schoelz, J.E. Uncoupling resistance from cell death in the hypersensitive response of Nicotiana species to cauliflower mosaic virus infection. Mol. Plant Microbe Interact. 2001, 14, 31–41. [Google Scholar] [CrossRef]
- Harries, P.A.; Palanichelvam, K.; Yu, W.; Schoelz, J.E.; Nelson, R.S. The cauliflower mosaic virus protein P6 forms motile inclusions that traffic along actin microfilaments and stabilize microtubules. Plant Physiol. 2009, 149, 1005–1016. [Google Scholar] [CrossRef]
- Love, A.J.; Geri, C.; Laird, J.; Carr, C.; Yun, B.; Loake, G.J.; Tada, Y.; Sadanandom, A.; Milner, J.J. Cauliflower mosaic virus protein P6 inhibits signaling responses to salicylic acid and regulates innate immunity. PLoS ONE 2012, 7, e47535. [Google Scholar] [CrossRef]
- Cawly, J.; Cole, A.B.; Király, L.; Qiu, W.; Schoelz, J.E. The plant gene CCD1 selectively blocks cell death during the hypersensitive response to Cauliflower mosaic virus infection. Mol. Plant Microbe Interact. 2005, 18, 212–219. [Google Scholar] [CrossRef]
- de Ronde, D.; Butterbach, P.; Lohuis, D.; Hedil, M.; van Lent, J.W.; Kormelink, R. Tsw gene-based resistance is triggered by a functional RNA silencing suppressor protein of the Tomato spotted wilt virus. Mol. Plant Pathol. 2013, 14, 405–415. [Google Scholar] [CrossRef]
- van Grinsven, I.L.; Martin, E.C.; Petrescu, A.J.; Kormelink, R. Tsw—A case study on structure-function puzzles in plant NLRs with unusually large LRR domains. Front. Plant Sci. 2022, 13, 983693. [Google Scholar] [CrossRef]
- Schnettler, E.; Hemmes, H.; Huismann, R.; Goldbach, R.; Prins, M.; Kormelink, R. Diverging affinity of tospovirus RNA silencing suppressor proteins, NSs, for various RNA duplex molecules. J. Virol. 2010, 84, 11542–11554. [Google Scholar] [CrossRef]
- Margaria, P.; Bosco, L.; Vallino, M.; Ciuffo, M.; Mautino, G.C.; Tavella, L.; Turina, M. The NSs protein of tomato spotted wilt virus is required for persistent infection and transmission by Frankliniella occidentalis. J. Virol. 2014, 88, 5788–5802. [Google Scholar] [CrossRef]
- de Ronde, D.; Pasquier, A.; Ying, S.; Butterbach, P.; Lohuis, D.; Kormelink, R. Analysis of Tomato spotted wilt virus NSs protein indicates the importance of the N-terminal domain for avirulence and RNA silencing suppression. Mol. Plant Pathol. 2014, 15, 185–195. [Google Scholar] [CrossRef]
- Almási, A.; Nemes, K.; Csömör, Z.; Tóbiás, I.; Palkovics, L.; Salánki, K. A single point mutation in Tomato spotted wilt virus NSs protein is sufficient to overcome Tsw-gene-mediated resistance in pepper. J. Gen. Virol. 2017, 98, 1521–1525. [Google Scholar] [CrossRef]
- Sharma, N.; Sahu, P.P.; Prasad, A.; Muthamilarasan, M.; Waseem, M.; Khan, Y.; Thakur, J.K.; Chakraborty, S.; Prasad, M. The Sw5a gene confers resistance to ToLCNDV and triggers an HR response after direct AC4 effector recognition. Proc. Natl. Acad. Sci. USA 2021, 118, e2101833118. [Google Scholar] [CrossRef]
- Moriones, E.; Praveen, S.; Chakraborty, S. Tomato Leaf Curl New Delhi Virus: An Emerging Virus Complex Threatening Vegetable and Fiber Crops. Viruses 2017, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, L.; Jin, T.; Nie, Y.; Liu, H.; Qiu, Y.; Yang, Y.; Li, B.; Zhang, J.; Wang, D.; et al. A cell wall-localized NLR confers resistance to Soybean mosaic virus by recognizing viral-encoded cylindrical inclusion protein. Mol. Plant. 2021, 14, 1881–1900. [Google Scholar] [CrossRef] [PubMed]
- Eggenberger, A.L.; Hajimorad, M.R.; Hill, J.H. Gain of virulence on Rsv1-genotype soybean by an avirulent Soybean mosaic virus requires concurrent mutations in both P3 and HC-Pro. Mol. Plant Microbe Interact. 2008, 21, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.K.; Lee, S.H.; Kim, K.H. Strain-specific cylindrical inclusion protein of soybean mosaic virus elicits extreme resistance and a lethal systemic hypersensitive response in two resistant soybean cultivars. Mol. Plant Microbe Interact. 2009, 22, 1151–1159. [Google Scholar] [CrossRef]
- Chowda-Reddy, R.V.; Sun, H.; Chen, H.; Poysa, V.; Ling, H.; Gijzen, M.; Wang, A. Mutations in the P3 protein of Soybean mosaic virus G2 isolates determine virulence on Rsv4-genotype soybean. Mol. Plant Microbe Interact. 2011, 24, 37–43. [Google Scholar] [CrossRef]
- Khatabi, B.; Wen, R.H.; Hajimorad, M.R. Fitness penalty in susceptible host is associated with virulence of Soybean mosaic virus on Rsv1-genotype soybean: A consequence of perturbation of HC-Pro and not P3. Mol. Plant Pathol. 2013, 14, 885–897. [Google Scholar] [CrossRef]
- Martin, R.; Qi, T.; Zhang, H.; Liu, F.; King, M.; Toth, C.; Nogales, E.; Staskawicz, B.J. Structure of the activated ROQ1 resistosome directly recognizing the pathogen effector XopQ. Science 2020, 370, eabd9993. [Google Scholar] [CrossRef]
- Duxbury, Z.; Wu, C.H.; Ding, P. A Comparative Overview of the Intracellular Guardians of Plants and Animals: NLRs in Innate Immunity and Beyond. Annu. Rev. Plant Biol. 2021, 72, 155–184. [Google Scholar] [CrossRef]
- Ramírez-Zavaleta, C.Y.; García-Barrera, L.J.; Rodríguez-Verástegui, L.L.; Arrieta-Flores, D.; Gregorio-Jorge, J. An Overview of PRR- and NLR-Mediated Immunities: Conserved Signaling Components across the Plant Kingdom That Communicate Both Pathways. Int. J. Mol. Sci. 2022, 23, 12974. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ding, P.; Jones, J.D.G. Thirty years of resistance: Zig-zag through the plant immune system. Plant Cell 2022, 34, 1447–1478. [Google Scholar] [CrossRef]
- Wróblewski, T.; Spiridon, L.; Martin, E.C.; Petrescu, A.J.; Cavanaugh, K.; Truco, M.J.; Xu, H.; Gozdowski, D.; Pawłowski, K.; Michelmore, R.W.; et al. Genome-wide functional analyses of plant coiled-coil NLR-type pathogen receptors reveal essential roles of their N-terminal domain in oligomerization, networking, and immunity. PLoS Biol. 2018, 16, e2005821. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Pennill, L.A.; Ning, J.; Lee, S.W.; Ramalingam, J.; Webb, C.A.; Zhao, B.; Sun, Q.; Nelson, J.C.; Leach, J.E.; et al. Diversity in nucleotide binding site-leucine-rich repeat genes in cereals. Genome Res. 2002, 12, 1871–1884. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Wu, R.; Guttman, D.S.; Desveaux, D. Allele-specific virulence attenuation of the Pseudomonas syringae HopZ1a type III effector via the Arabidopsis ZAR1 resistance protein. PLoS Genet. 2010, 6, e1000894. [Google Scholar] [CrossRef]
- Schultink, A.; Qi, T.; Bally, J.; Staskawicz, B. Using forward genetics in Nicotiana benthamiana to uncover the immune signaling pathway mediating recognition of the Xanthomonas perforans effector XopJ4. New Phytol. 2019, 221, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Moffett, P.; Farnham, G.; Peart, J.; Baulcombe, D.C. Interaction between domains of a plant NBS-LRR protein in disease resistance-related cell death. EMBO J. 2002, 21, 4511–4519. [Google Scholar] [CrossRef]
- Botella, M.A.; Parker, J.E.; Frost, L.N.; Bittner-Eddy, P.D.; Beynon, J.L.; Daniels, M.J.; Holub, E.B.; Jones, J.D. Three genes of the Arabidopsis RPP1 complex resistance locus recognize distinct Peronospora parasitica avirulence determinants. Plant Cell 1998, 10, 1847–1860. [Google Scholar] [CrossRef]
- Wu, C.H.; Abd-El-Haliem, A.; Bozkurt, T.O.; Belhaj, K.; Terauchi, R.; Vossen, J.H.; Kamoun, S. NLR network mediates immunity to diverse plant pathogens. Proc. Natl. Acad. Sci. USA 2017, 114, 8113–8118. [Google Scholar] [CrossRef]
- Wu, C.H.; Derevnina, L.; Kamoun, S. Receptor networks underpin plant immunity. Science 2018, 360, 1300–1301. [Google Scholar] [CrossRef]
- Wang, J.; Hu, M.; Wang, J.; Qi, J.; Han, Z.; Wang, G.; Qi, Y.; Wang, H.W.; Zhou, J.M.; Chai, J. Reconstitution and structure of a plant NLR resistosome conferring immunity. Science 2019, 364, eaav5870. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.; Hu, M.; Wu, S.; Qi, J.; Wang, G.; Han, Z.; Qi, Y.; Gao, N.; Wang, H.W.; et al. Ligand-triggered allosteric ADP release primes a plant NLR complex. Science 2019, 364, eaav5868. [Google Scholar] [CrossRef]
- Ma, S.; Lapin, D.; Liu, L.; Sun, Y.; Song, W.; Zhang, X.; Logemann, E.; Yu, D.; Wang, J.; Jirschitzka, J.; et al. Direct pathogen-induced assembly of an NLR immune receptor complex to form a holoenzyme. Science 2020, 370, eabe3069. [Google Scholar] [CrossRef] [PubMed]
- Förderer, A.; Li, E.; Lawson, A.W.; Deng, Y.N.; Sun, Y.; Logemann, E.; Zhang, X.; Wen, J.; Han, Z.; Chang, J.; et al. A wheat resistosome defines common principles of immune receptor channels. Nature 2022, 610, 532–539. [Google Scholar] [CrossRef]
- Adachi, H.; Contreras, M.P.; Harant, A.; Wu, C.H.; Derevnina, L.; Sakai, T.; Duggan, C.; Moratto, E.; Bozkurt, T.O.; Maqbool, A.; et al. N-terminal motif in NLR immune receptors is functionally conserved across distantly related plant species. eLife 2019, 8, e49956. [Google Scholar] [CrossRef] [PubMed]
- Swiderski, M.R.; Birker, D.; Jones, J.D. The TIR domain of TIR-NB-LRR resistance proteins is a signaling domain involved in cell death induction. Mol. Plant Microbe Interact. 2009, 22, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Collier, S.M.; Hamel, L.P.; Moffett, P. Cell death mediated by the N-terminal domains of a unique and highly conserved class of NB-LRR protein. Mol. Plant Microbe Interact. 2011, 24, 918–931. [Google Scholar] [CrossRef]
- Cesari, S.; Moore, J.; Chen, C.; Webb, D.; Periyannan, S.; Mago, R.; Bernoux, M.; Lagudah, E.S.; Dodds, P.N. Cytosolic activation of cell death and stem rust resistance by cereal MLA-family CC-NLR proteins. Proc. Natl. Acad. Sci. USA 2016, 113, 10204–10209. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, X.; Sun, K.; Gao, Z. Structure and function analysis of a CC-NBS-LRR protein AT1G12290. Biochem. Biophys. Res. Commun. 2021, 534, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Cheng, W.; Spiridon, L.N.; Töller, A.; Lukasik, E.; Saijo, Y.; Liu, P.; Shen, Q.H.; Micluta, M.A.; Somssich, I.E.; et al. Coiled-coil domain-dependent homodimerization of intracellular barley immune receptors defines a minimal functional module for triggering cell death. Cell Host Microbe 2011, 9, 187–199. [Google Scholar] [CrossRef]
- Bernoux, M.; Ve, T.; Williams, S.; Warren, C.; Hatters, D.; Valkov, E.; Zhang, X.; Ellis, J.G.; Kobe, B.; Dodds, P.N. Structural and functional analysis of a plant resistance protein TIR domain reveals interfaces for self-association, signaling, and autoregulation. Cell Host Microbe 2011, 9, 200–211. [Google Scholar] [CrossRef]
- Rairdan, G.J.; Collier, S.M.; Sacco, M.A.; Baldwin, T.T.; Boettrich, T.; Moffett, P. The coiled-coil and nucleotide binding domains of the Potato Rx disease resistance protein function in pathogen recognition and signaling. Plant Cell 2008, 20, 739–751. [Google Scholar] [CrossRef]
- Feehan, J.M.; Castel, B.; Bentham, A.R.; Jones, J.D. Plant NLRs get by with a little help from their friends. Curr. Opin. Plant Biol. 2020, 56, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Adachi, H.; Derevnina, L.; Kamoun, S. NLR singletons, pairs, and networks: Evolution, assembly, and regulation of the intracellular immunoreceptor circuitry of plants. Curr. Opin. Plant Biol. 2019, 50, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Derevnina, L.; Contreras, M.P.; Adachi, H.; Upson, J.; Vergara Cruces, A.; Xie, R.; Skłenar, J.; Menke, F.L.H.; Mugford, S.T.; MacLean, D.; et al. Plant pathogens convergently evolved to counteract redundant nodes of an NLR immune receptor network. PLoS Biol. 2021, 19, e3001136. [Google Scholar] [CrossRef]
- Contreras, M.P.; Pai, H.; Tumtas, Y.; Duggan, C.; Yuen, E.L.H.; Cruces, A.V.; Kourelis, J.; Ahn, H.K.; Lee, K.T.; Wu, C.H.; et al. Sensor NLR immune proteins activate oligomerization of their NRC helpers in response to plant pathogens. EMBO J. 2023, 42, e111519. [Google Scholar] [CrossRef]
- Oh, S.K.; Kwon, S.Y.; Choi, D. Rpi-blb2-Mediated Hypersensitive Cell Death Caused by Phytophthora infestans AVRblb2 Requires SGT1, but not EDS1, NDR1, Salicylic Acid-, Jasmonic Acid-, or Ethylene-Mediated Signaling. Plant Pathol. J. 2014, 30, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Duggan, C.; Moratto, E.; Savage, Z.; Hamilton, E.; Adachi, H.; Wu, C.H.; Leary, A.Y.; Tumtas, Y.; Rothery, S.M.; Maqbool, A.; et al. Dynamic localization of a helper NLR at the plant-pathogen interface underpins pathogen recognition. Proc. Natl. Acad. Sci. USA 2021, 118, e2104997118. [Google Scholar] [CrossRef]
- Adachi, H.; Sakai, T.; Harant, A.; Pai, H.; Honda, K.; Toghani, A.; Claeys, J.; Duggan, C.; Bozkurt, T.O.; Wu, C.H.; et al. An atypical NLR protein modulates the NRC immune receptor network in Nicotiana benthamiana. PLoS Genet. 2023, 19, e1010500. [Google Scholar] [CrossRef] [PubMed]
- Lapin, D.; Bhandari, D.D.; Parker, J.E. Origins and Immunity Networking Functions of EDS1 Family Proteins. Annu. Rev. Phytopathol. 2020, 58, 253–276. [Google Scholar] [CrossRef] [PubMed]
- Lapin, D.; Johanndrees, O.; Wu, Z.; Li, X.; Parker, J.E. Molecular innovations in plant TIR-based immunity signaling. Plant Cell 2022, 34, 1479–1496. [Google Scholar] [CrossRef]
- Saile, S.C.; Jacob, P.; Castel, B.; Jubic, L.M.; Salas-Gonzáles, I.; Bäcker, M.; Jones, J.D.G.; Dangl, J.L.; El Kasmi, F. Two unequally redundant “helper” immune receptor families mediate Arabidopsis thaliana intracellular “sensor” immune receptor functions. PLoS Biol. 2020, 18, e3000783. [Google Scholar] [CrossRef]
- Sun, X.; Lapin, D.; Feehan, J.M.; Stolze, S.C.; Kramer, K.; Dongus, J.A.; Rzemieniewski, J.; Blanvillain-Baufumé, S.; Harzen, A.; Bautor, J.; et al. Pathogen effector recognition-dependent association of NRG1 with EDS1 and SAG101 in TNL receptor immunity. Nat. Commun. 2021, 12, 3335. [Google Scholar] [CrossRef] [PubMed]
- Feehan, J.M.; Wang, J.; Sun, X.; Choi, J.; Ahn, H.K.; Ngou, B.P.M.; Parker, J.E.; Jones, J.D.G. Oligomerization of a plant helper NLR requires cell-surface and intracellular immune receptor activation. Proc. Natl. Acad. Sci. USA 2023, 120, e2210406120. [Google Scholar] [CrossRef] [PubMed]
- Peart, J.R.; Mestre, P.; Lu, R.; Malcuit, I.; Baulcombe, D.C. NRG1, a CC-NB-LRR protein, together with N, a TIR-NB-LRR protein, mediates resistance against tobacco mosaic virus. Curr. Biol. 2005, 15, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Seong, K.; Thomazella, D.P.T.; Kim, J.R.; Pham, J.; Seo, E.; Cho, M.J.; Schultink, A.; Staskawicz, B.J. NRG1 functions downstream of EDS1 to regulate TIR-NLR-mediated plant immunity in Nicotiana benthamiana. Proc. Natl. Acad. Sci. USA 2018, 115, E10979–E10987. [Google Scholar] [CrossRef] [PubMed]
- Lapin, D.; Kovacova, V.; Sun, X.; Dongus, J.A.; Bhandari, D.; von Born, P.; Bautor, J.; Guarneri, N.; Rzemieniewski, J.; Stuttmann, J.; et al. A Coevolved EDS1-SAG101-NRG1 Module Mediates Cell Death Signaling by TIR-Domain Immune Receptors. Plant Cell 2019, 31, 2430–2455. [Google Scholar] [CrossRef]
- Voss, M.; Toelzer, C.; Bhandari, D.D.; Parker, J.E.; Niefind, K. Arabidopsis immunity regulator EDS1 in a PAD4/SAG101-unbound form is a monomer with an inherently inactive conformation. J. Struct. Biol. 2019, 208, 107390. [Google Scholar] [CrossRef]
- Dongus, J.A.; Parker, J.E. EDS1 signalling: At the nexus of intracellular and surface receptor immunity. Curr. Opin. Plant Biol. 2021, 62, 102039. [Google Scholar] [CrossRef]
- Wu, Z.; Tian, L.; Liu, X.; Zhang, Y.; Li, X. TIR signal promotes interactions between lipase-like proteins and ADR1-L1 receptor and ADR1-L1 oligomerization. Plant Physiol. 2021, 187, 681–686. [Google Scholar] [CrossRef]
- Huang, S.; Jia, A.; Song, W.; Hessler, G.; Meng, Y.; Sun, Y.; Xu, L.; Laessle, H.; Jirschitzka, J.; Ma, S.; et al. Identification and receptor mechanism of TIR-catalyzed small molecules in plant immunity. Science 2022, 377, eabq3297. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, H.; Hu, L.; Martinez, P.P.; Moschou, P.N.; Cevik, V.; Ding, P.; Duxbury, Z.; Sarris, P.F.; Jones, J.D.G. Distinct modes of derepression of an Arabidopsis immune receptor complex by two different bacterial effectors. Proc. Natl. Acad. Sci. USA 2018, 115, 10218–10227. [Google Scholar] [CrossRef]
- Césari, S.; Kanzaki, H.; Fujiwara, T.; Bernoux, M.; Chalvon, V.; Kawano, Y.; Shimamoto, K.; Dodds, P.; Terauchi, R.; Kroj, T. The NB-LRR proteins RGA4 and RGA5 interact functionally and physically to confer disease resistance. EMBO J. 2014, 33, 1941–1959. [Google Scholar] [CrossRef] [PubMed]
- Wan, W.L.; Kim, S.T.; Castel, B.; Charoennit, N.; Chae, E. Genetics of autoimmunity in plants: An evolutionary genetics perspective. New Phytol. 2021, 229, 1215–1233. [Google Scholar] [CrossRef] [PubMed]
- Kourelis, J.; Adachi, H. Activation and Regulation of NLR Immune Receptor Networks. Plant Cell Physiol. 2022, 63, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.P.; Seddon, A.E.; Moghe, G.D.; Simenc, M.C.; Shiu, S.H. Characteristics of Plant Essential Genes Allow for within- and between-Species Prediction of Lethal Mutant Phenotypes. Plant Cell 2015, 27, 2133–2147. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.J.; Li, W.; Chao, Y.; Schwarzenbacher, R.; Shi, Y. Structure of the apoptotic protease-activating factor 1 bound to ADP. Nature 2005, 434, 926–933. [Google Scholar] [CrossRef]
- Reubold, T.F.; Wohlgemuth, S.; Eschenburg, S. Crystal structure of full-length Apaf-1: How the death signal is relayed in the mitochondrial pathway of apoptosis. Structure 2011, 19, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Li, Y.; Hu, Q.; Bai, X.C.; Huang, W.; Yan, C.; Scheres, S.H.; Shi, Y. Atomic structure of the apoptosome: Mechanism of cytochrome c- and dATP-mediated activation of Apaf-1. Genes. Dev. 2015, 29, 2349–2361. [Google Scholar] [CrossRef]
- Hu, Z.; Yan, C.; Liu, P.; Huang, Z.; Ma, R.; Zhang, C.; Wang, R.; Zhang, Y.; Martinon, F.; Miao, D.; et al. Crystal structure of NLRC4 reveals its autoinhibition mechanism. Science 2013, 341, 172–175. [Google Scholar] [CrossRef]
- Maekawa, S.; Ohto, U.; Shibata, T.; Miyake, K.; Shimizu, T. Crystal structure of NOD2 and its implications in human disease. Nat. Commun. 2016, 7, 11813. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, S.; Ruan, J.; Wu, J.; Tong, A.B.; Yin, Q.; Li, Y.; David, L.; Lu, A.; Wang, W.L.; et al. Cryo-EM structure of the activated NAIP2-NLRC4 inflammasome reveals nucleated polymerization. Science 2015, 350, 404–409. [Google Scholar] [CrossRef]
- Bi, G.; Su, M.; Li, N.; Liang, Y.; Dang, S.; Xu, J.; Hu, M.; Wang, J.; Zou, M.; Deng, Y.; et al. The ZAR1 resistosome is a calcium-permeable channel triggering plant immune signaling. Cell 2021, 184, 3528–3541.e12. [Google Scholar] [CrossRef]
- Horsefield, S.; Burdett, H.; Zhang, X.; Manik, M.K.; Shi, Y.; Chen, J.; Qi, T.; Gilley, J.; Lai, J.S.; Rank, M.X.; et al. NAD+ cleavage activity by animal and plant TIR domains in cell death pathways. Science 2019, 365, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Lukasik, E.; Takken, F.L. STANDing strong, resistance proteins instigators of plant defence. Curr. Opin. Plant Biol. 2009, 12, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Pang, Y.; Hu, Q.; Liu, Q.; Li, H.; Zhou, Y.; He, T.; Liang, Q.; Liu, Y.; Yuan, X.; et al. Crystal structure of the Caenorhabditis elegans apoptosome reveals an octameric assembly of CED-4. Cell 2010, 141, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Urbach, J.M.; Ausubel, F.M. The NBS-LRR architectures of plant R-proteins and metazoan NLRs evolved in independent events. Proc. Natl. Acad. Sci. USA 2017, 114, 1063–1068. [Google Scholar] [CrossRef]
- Hu, Z.; Chai, J. Structural Mechanisms in NLR Inflammasome Assembly and Signaling. Curr. Top. Microbiol. Immunol. 2016, 397, 23–42. [Google Scholar]
- Li, Y.; Zhou, M.; Hu, Q.; Bai, X.C.; Huang, W.; Scheres, S.H.; Shi, Y. Mechanistic insights into caspase-9 activation by the structure of the apoptosome holoenzyme. Proc. Natl. Acad. Sci. USA 2017, 114, 1542–1547. [Google Scholar] [CrossRef]
- Wang, L.; Wu, H. Keeping the Death Protein in Check. Immunity 2019, 51, 1–2. [Google Scholar] [CrossRef]
- Wan, L.; Essuman, K.; Anderson, R.G.; Sasaki, Y.; Monteiro, F.; Chung, E.H.; Osborne Nishimura, E.; DiAntonio, A.; Milbrandt, J.; Dangl, J.L.; et al. TIR domains of plant immune receptors are NAD+-cleaving enzymes that promote cell death. Science 2019, 365, 799–803. [Google Scholar] [CrossRef]
- Gerdts, J.; Brace, E.J.; Sasaki, Y.; DiAntonio, A.; Milbrandt, J. SARM1 activation triggers axon degeneration locally via NAD⁺ destruction. Science 2015, 348, 453–457. [Google Scholar] [CrossRef]
- Duxbury, Z.; Wang, S.; MacKenzie, C.I.; Tenthorey, J.L.; Zhang, X.; Huh, S.U.; Hu, L.; Hill, L.; Ngou, P.M.; Ding, P.; et al. Induced proximity of a TIR signaling domain on a plant-mammalian NLR chimera activates defense in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 18832–18839. [Google Scholar] [CrossRef] [PubMed]
- Pontier, D.; Mittler, R.; Lam, E. Mechanism of cell death and disease resistance induction by transgenic expression of bacterio-opsin. Plant J. 2002, 30, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, S.; Lee, J.; Gäbler, Y.; Kemmerling, B.; Haapalainen, M.L.; Li, C.M.; Wei, Z.; Keller, H.; Joosten, M.; Taira, S.; et al. Separable roles of the Pseudomonas syringae pv. phaseolicola accessory protein HrpZ1 in ion-conducting pore formation and activation of plant immunity. Plant J. 2009, 57, 706–717. [Google Scholar] [CrossRef]
- Remick, B.C.; Gaidt, M.M.; Vance, R.E. Effector-Triggered Immunity. Annu. Rev. Immunol. 2023, 41, 453–481. [Google Scholar] [CrossRef] [PubMed]
- Tenthorey, J.L.; Haloupek, N.; López-Blanco, J.R.; Grob, P.; Adamson, E.; Hartenian, E.; Lind, N.A.; Bourgeois, N.M.; Chacón, P.; Nogales, E.; et al. The structural basis of flagellin detection by NAIP5: A strategy to limit pathogen immune evasion. Science 2017, 358, 888–893. [Google Scholar] [CrossRef] [PubMed]
- Tenthorey, J.L.; Kofoed, E.M.; Daugherty, M.D.; Malik, H.S.; Vance, R.E. Molecular basis for specific recognition of bacterial ligands by NAIP/NLRC4 inflammasomes. Mol. Cell 2014, 54, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Krasileva, K.V.; Dahlbeck, D.; Staskawicz, B.J. Activation of an Arabidopsis resistance protein is specified by the in planta association of its leucine-rich repeat domain with the cognate oomycete effector. Plant Cell 2010, 22, 2444–2458. [Google Scholar] [CrossRef]
- Schultink, A.; Qi, T.; Lee, A.; Steinbrenner, A.D.; Staskawicz, B. Roq1 mediates recognition of the Xanthomonas and Pseudomonas effector proteins XopQ and HopQ1. Plant J. 2017, 92, 787–795. [Google Scholar] [CrossRef]
- Mackey, D.; Belkhadir, Y.; Alonso, J.M.; Ecker, J.R.; Dangl, J.L. Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance. Cell 2003, 112, 379–389. [Google Scholar] [CrossRef]
- Redditt, T.J.; Chung, E.H.; Karimi, H.Z.; Rodibaugh, N.; Zhang, Y.; Trinidad, J.C.; Kim, J.H.; Zhou, Q.; Shen, M.; Dangl, J.L.; et al. AvrRpm1 Functions as an ADP-Ribosyl Transferase to Modify NOI Domain-Containing Proteins, Including Arabidopsis and Soybean RPM1-Interacting Protein4. Plant Cell 2019, 31, 2664–2681. [Google Scholar]
- Xu, H.; Shi, J.; Gao, H.; Liu, Y.; Yang, Z.; Shao, F.; Dong, N. The N-end rule ubiquitin ligase UBR2 mediates NLRP1B inflammasome activation by anthrax lethal toxin. EMBO J. 2019, 38, e101996. [Google Scholar] [CrossRef] [PubMed]
- Sarris, P.F.; Duxbury, Z.; Huh, S.U.; Ma, Y.; Segonzac, C.; Sklenar, J.; Derbyshire, P.; Cevik, V.; Rallapalli, G.; Saucet, S.B.; et al. A Plant Immune Receptor Detects Pathogen Effectors that Target WRKY Transcription Factors. Cell 2015, 161, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Dyrka, W.; Lamacchia, M.; Durrens, P.; Kobe, B.; Daskalov, A.; Paoletti, M.; Sherman, D.J.; Saupe, S.J. Diversity and variability of NOD-like receptors in fungi. Genome Biol. Evol. 2014, 6, 3137–3158. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Vance, R.E.; Dangl, J.L. Intracellular innate immune surveillance devices in plants and animals. Science 2016, 354, aaf6395. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).