
Novel Toxicodynamic Model of Subcutaneous Envenomation to Characterize Snake Venom Coagulopathies and Assess the Efficacy of Site-Directed Inorganic Antivenoms


Abstract
:1. Introduction
2. Results
2.1. Effects of C. s. Scutulatus Envenomation on Whole Blood and Plasmatic Coagulation and Efficacy of Antivenom
2.2. Effects of B. moojeni Envenomation on Whole Blood Coagulation and Efficacy of Antivenom
2.3. Effects of C. rhodostoma Envenomation on Whole Blood Coagulation and Efficacy of Antivenom
3. Discussion
4. Materials and Methods
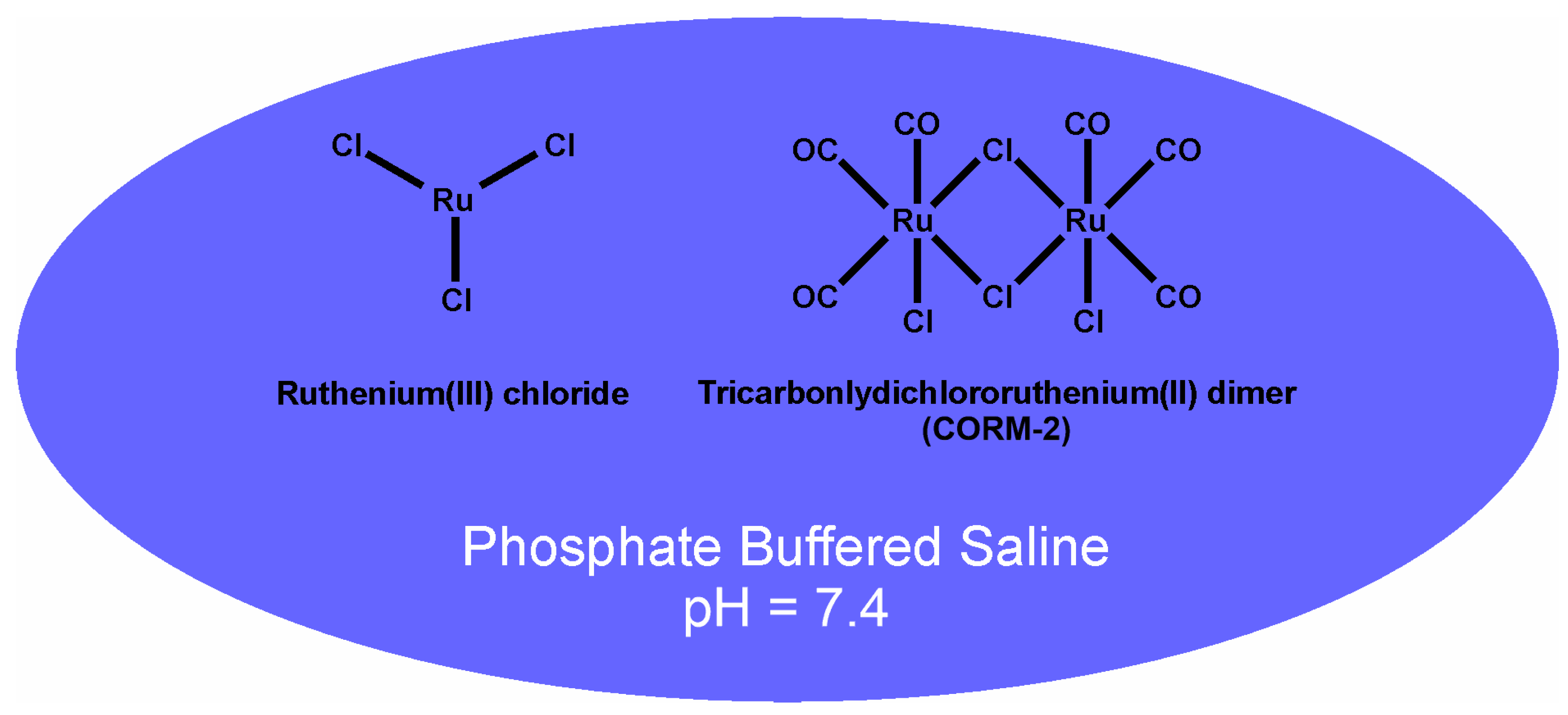
4.1. Chemicals and Venoms
4.2. Rabbit Model
4.3. Coagulation Monitoring
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.S.; Georgieva, D.; Genov, N.; Murakami, M.T.; Sinha, M.; Kumar, R.P.; Kaur, P.; Kumar, S.; Dey, S.; Sharma, S.; et al. Enzymatic toxins from snake venom: Structural characterization and mechanism of catalysis. FEBS J. 2011, 278, 4544–4576. [Google Scholar] [CrossRef] [PubMed]
- Osipov, A.; Utkin, Y. What Are the Neurotoxins in Hemotoxic Snake Venoms? Int. J. Mol. Sci. 2023, 24, 2919. [Google Scholar] [CrossRef]
- Teixeira, C.; Fernandes, C.M.; Leiguez, E.; Chudzinski-Tavassi, A.M. Inflammation Induced by Platelet-Activating Viperid Snake Venoms: Perspectives on Thromboinflammation. Front. Immunol. 2019, 10, 2082. [Google Scholar] [CrossRef]
- Ullah, A.; Masood, R.; Ali, I.; Ullah, K.; Ali, H.; Akbar, H.; Betzel, C. Thrombin-like enzymes from snake venom: Structural characterization and mechanism of action. Int. J. Biol. Macromol 2018, 114, 788–811. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Koh, C.Y. Metalloproteases Affecting Blood Coagulation, Fibrinolysis and Platelet Aggregation from Snake Venoms: Definition and Nomenclature of Interaction Sites. Toxins 2016, 8, 284. [Google Scholar] [CrossRef]
- Pinyachat, A. Comparative Studies of Structural and Functional Properties of Snake Venom Metalloproteinases. J. Med. Assoc. Thai 2016, 99 (Suppl. 1), S76–S88. [Google Scholar]
- Serrano, S.M. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef]
- Guo, C.; Liu, S.; Yao, Y.; Zhang, Q.; Sun, M.Z. Past decade study of snake venom L-amino acid oxidase. Toxicon 2012, 60, 302–311. [Google Scholar] [CrossRef]
- Del Brutto, O.H.; Del Brutto, V.J. Neurological complications of venomous snake bites: A review. Acta Neurol. Scand. 2012, 125, 363–372. [Google Scholar] [CrossRef]
- Hodgson, W.C.; Wickramaratna, J.C. In vitro neuromuscular activity of snake venoms. Clin. Exp. Pharmacol. Physiol. 2002, 29, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Bickler, P.E. Amplification of Snake Venom Toxicity by Endogenous Signaling Pathways. Toxins 2020, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Rowan, E.G. What does beta-bungarotoxin do at the neuromuscular junction? Toxicon 2001, 39, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Pretorius, E. Carbon monoxide: Anticoagulant or procoagulant? Thromb. Res. 2014, 133, 315–321. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Sánchez, E.E.; Redford, D.T. Characterization of the Rabbit as an In Vitro and In Vivo Model to Assess the Effects of Fibrinogenolytic Activity of Snake Venom on Coagulation. Basic Clin. Pharmacol. Toxicol. 2018, 122, 157–164. [Google Scholar] [CrossRef]
- Nielsen, V.G. Crotalus atrox Venom Exposed to Carbon Monoxide Has Decreased Fibrinogenolytic Activity In Vivo in Rabbits. Basic Clin. Pharmacol. Toxicol. 2018, 122, 82–86. [Google Scholar] [CrossRef]
- van Helden, D.F.; Thomas, P.A.; Dosen, P.J.; Imtiaz, M.S.; Laver, D.R.; Isbister, G.K. Pharmacological approaches that slow lymphatic flow as a snakebite first aid. PLoS Negl. Trop. Dis. 2014, 8, e2722. [Google Scholar] [CrossRef]
- Paniagua, D.; Vergara, I.; Román, R.; Romero, C.; Benard-Valle, M.; Calderón, A.; Jiménez, L.; Bernas, M.J.; Witte, M.H.; Boyer, L.V.; et al. Antivenom effect on lymphatic absorption and pharmacokinetics of coral snake venom using a large animal model. Clin. Toxicol. 2019, 57, 727–734. [Google Scholar] [CrossRef]
- van Helden, D.F.; Dosen, P.J.; O’Leary, M.A.; Isbister, G.K. Two pathways for venom toxin entry consequent to injection of an Australian elapid snake venom. Sci. Rep. 2019, 9, 8595. [Google Scholar] [CrossRef]
- Bachmann, S.B.; Proulx, S.T.; He, Y.; Ries, M.; Detmar, M. Differential effects of anaesthesia on the contractility of lymphatic vessels in vivo. J. Physiol. 2019, 597, 2841–2852. [Google Scholar] [CrossRef]
- Soto-Miranda, M.A.; Suami, H.; Chang, D.W. Mapping superficial lymphatic territories in the rabbit. Anat. Rec. 2013, 296, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.; Mora, R.; Lomonte, B.; Gutiérrez, J.M. Effects of Bothrops asper snake venom on lymphatic vessels: Insights into a hidden aspect of envenomation. PLoS Negl. Trop. Dis. 2008, 2, e318. [Google Scholar] [CrossRef] [PubMed]
- Fahmi, L.; Makran, B.; Boussadda, L.; Lkhider, M.; Ghalim, N. Haemostasis disorders caused by envenomation by Cerastes cerastes and Macrovipera mauritanica vipers. Toxicon 2016, 116, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, L.K.; Saroja, J.B.; Rajalingam, M.; John, V.; Valappil, M.P.; Sreelatha, H.V. Rabbit snakebite model to assess safety and efficacy of anti viper chicken antibodies (IgY). Am. J. Clin. Exp. Med. 2015, 3, 32–38. [Google Scholar] [CrossRef]
- Nielsen, V.G. Carbon monoxide inhibits the anticoagulant activity of Mojave rattlesnake venoms type A and B. J. Thromb. Thrombolysis. 2019, 48, 256–262. [Google Scholar] [CrossRef]
- Nielsen, V.G. Modulation of Diverse Procoagulant Venom Activities by Combinations of Platinoid Compounds. Int. J. Mol. Sci. 2021, 22, 4612. [Google Scholar] [CrossRef]
- Carstairs, S.D.; Kreshak, A.A.; Tanen, D.A. Crotaline Fab antivenom reverses platelet dysfunction induced by Crotalus scutulatus venom: An in vitro study. Acad. Emerg. Med. 2013, 20, 522–525. [Google Scholar] [CrossRef]
- Corrigan, J.J., Jr.; Jeter, M.A. Mojave rattlesnake (Crotalus scutulatus scutulatus) venom: In vitro effect on platelets, fibrinolysis, and fibrinogen clotting. Vet. Hum. Toxicol. 1990, 32, 439–441. [Google Scholar]
- Aguiar, W.D.S.; Galizio, N.D.C.; Serino-Silva, C.; Sant’Anna, S.S.; Grego, K.F.; Tashima, A.K.; Nishiduka, E.S.; Morais-Zani, K.; Tanaka-Azevedo, A.M. Comparative compositional and functional analyses of Bothrops moojeni specimens reveal several individual variations. PLoS ONE 2019, 14, e0222206. [Google Scholar]
- Tang, E.L.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics of Calloselasma rhodostoma, the Malayan pit viper: A complex toxin arsenal unraveled. J. Proteom. 2016, 148, 44–56. [Google Scholar] [CrossRef]
- Glenn, J.L.; Straight, R. Mojave rattlesnake Crotalus scutulatus scutulatus venom: Variation in toxicity with geographical origin. Toxicon 1978, 16, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.F.; Maruyama, M.; Kamiguti, A.S.; Antonio, L.C. Comparative study of nine Bothrops snake venoms from adult female snakes and their offspring. Toxicon 1991, 29, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Pornmanee, P.; Sánchez, E.E.; López, G.; Petsom, A.; Khow, O.; Pakmanee, N.; Chanhome, L.; Sangvanich, P.; Pérez, J.C. Neutralization of lethality and proteolytic activities of Malayan pit viper (Calloselasma rhodostoma) venom with North American Virginia opossum (Didelphis virginiana) serum. Toxicon 2008, 52, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Chawla, N.; Mangla, D.; Gomes, S.B.; Arkebauer, M.R.; Wasko, K.A.; Sadacharam, K.; Vosseller, K. Carbon monoxide-releasing molecule-2 enhances coagulation in rabbit plasma and decreases bleeding time in clopidogrel/aspirin-treated rabbits. Blood Coagul. Fibrinolysis. 2011, 22, 756–759. [Google Scholar] [CrossRef]
- Uddin, M.J.; Jeong, J.; Pak, E.S.; Ha, H. CO-Releasing Molecule-2 Prevents Acute Kidney Injury through Suppression of ROS-Fyn-ER Stress Signaling in Mouse Model. Oxid. Med. Cell Longev. 2021, 2021, 9947772. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, Y.; Zhu, J.; Ma, J.; Niu, Q.; Wang, X. Carbon monoxide attenuates LPS-induced myocardial dysfunction in rats by regulating the mitochondrial dynamic equilibrium. Eur. J. Pharmacol. 2020, 889, 173726. [Google Scholar] [CrossRef]
- Joshi, H.P.; Kumar, H.; Choi, U.Y.; Lim, Y.C.; Choi, H.; Kim, J.; Kyung, J.W.; Sohn, S.; Kim, K.T.; Kim, J.K.; et al. CORM-2-Solid Lipid Nanoparticles Maintain Integrity of Blood-Spinal Cord Barrier After Spinal Cord Injury in Rats. Mol. Neurobiol. 2020, 57, 2671–2689. [Google Scholar] [CrossRef]
- Hervera, A.; Gou, G.; Leánez, S.; Pol, O. Effects of treatment with a carbon monoxide-releasing molecule and a heme oxygenase 1 inducer in the antinociceptive effects of morphine in different models of acute and chronic pain in mice. Psychopharmacology 2013, 228, 463–477. [Google Scholar] [CrossRef]
- Choi, E.K.; Park, H.J.; Sul, O.J.; Rajasekaran, M.; Yu, R.; Choi, H.S. Carbon monoxide reverses adipose tissue inflammation and insulin resistance upon loss of ovarian function. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E621–E630. [Google Scholar] [CrossRef]
- Magierowska, K.; Magierowski, M.; Hubalewska-Mazgaj, M.; Adamski, J.; Surmiak, M.; Sliwowski, Z.; Kwiecien, S.; Brzozowski, T. Carbon Monoxide (CO) Released from Tricarbonyldichlororuthenium (II) Dimer (CORM-2) in Gastroprotection against Experimental Ethanol-Induced Gastric Damage. PLoS ONE 2015, 10, e0140493. [Google Scholar] [CrossRef]
- Feger, M.; Fajol, A.; Lebedeva, A.; Meissner, A.; Michael, D.; Voelkl, J.; Alesutan, I.; Schleicher, E.; Reichetzeder, C.; Hocher, B.; et al. Effect of carbon monoxide donor CORM-2 on vitamin D3 metabolism. Kidney Blood Press Res. 2013, 37, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, A.A.; Prawez, S.; Kumar, D.; Prasad, R.; Amarpal; Tandan, S.K.; Kumar, D. Wound healing activity of carbon monoxide liberated from CO-releasing molecule (CO-RM). Naunyn. Schmiedebergs. Arch. Pharmacol. 2011, 384, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Castany, S.; Carcolé, M.; Leánez, S.; Pol, O. The role of carbon monoxide on the anti-nociceptive effects and expression of cannabinoid 2 receptors during painful diabetic neuropathy in mice. Psychopharmacology 2016, 233, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Jurga, A.M.; Piotrowska, A.; Starnowska, J.; Rojewska, E.; Makuch, W.; Mika, J. Treatment with a carbon monoxide-releasing molecule (CORM-2) inhibits neuropathic pain and enhances opioid effectiveness in rats. Pharmacol. Rep. 2016, 68, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, A.; Bernardes, G.J.; Shimanovich, U.; Sárria, M.P.; Nogueira, E.; Preto, A.; Gomes, A.C.; Cavaco-Paulo, A. Folic acid-tagged protein nanoemulsions loaded with CORM-2 enhance the survival of mice bearing subcutaneous A20 lymphoma tumors. Nanomedicine 2015, 11, 1077–1083. [Google Scholar] [CrossRef]
- Shen, W.C.; Wang, X.; Qin, W.T.; Qiu, X.F.; Sun, B.W. Exogenous carbon monoxide suppresses Escherichia coli vitality and improves survival in an Escherichia coli-induced murine sepsis model. Acta Pharmacol. Sin. 2014, 35, 1566–1576. [Google Scholar] [CrossRef]
- Yin, H.; Fang, J.; Liao, L.; Nakamura, H.; Maeda, H. Styrene-maleic acid copolymer-encapsulated CORM2, a water-soluble carbon monoxide (CO) donor with a constant CO-releasing property, exhibits therapeutic potential for inflammatory bowel disease. J. Control. Release 2014, 187, 14–21. [Google Scholar] [CrossRef]
- Hervera, A.; Leánez, S.; Motterlini, R.; Pol, O. Treatment with carbon monoxide-releasing molecules and an HO-1 inducer enhances the effects and expression of µ-opioid receptors during neuropathic pain. Anesthesiology 2013, 118, 1180–1197. [Google Scholar] [CrossRef]
- Hervera, A.; Leánez, S.; Negrete, R.; Motterlini, R.; Pol, O. Carbon monoxide reduces neuropathic pain and spinal microglial activation by inhibiting nitric oxide synthesis in mice. PLoS ONE 2012, 7, 43693. [Google Scholar] [CrossRef]
- Czibik, G.; Gravning, J.; Martinov, V.; Ishaq, B.; Knudsen, E.; Attramadal, H.; Valen, G. Gene therapy with hypoxia-inducible factor 1 alpha in skeletal muscle is cardioprotective in vivo. Life Sci. 2011, 88, 543–550. [Google Scholar] [CrossRef]
- Hidaka, A.; Azuma, Y.T.; Nakajima, H.; Takeuchi, T. Nitric oxide and carbon monoxide act as inhibitory neurotransmitters in the longitudinal muscle of C57BL/6J mouse distal colon. J. Pharmacol. Sci. 2010, 112, 231–241. [Google Scholar] [CrossRef]
- Lee, T.S.; Tsai, H.L.; Chau, L.Y. Induction of heme oxygenase-1 expression in murine macrophages is essential for the anti-inflammatory effect of low dose 15-deoxy-Delta 12,14-prostaglandin J2. J. Biol. Chem. 2003, 278, 19325–19330. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G. Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms. Int. J. Mol. Sci. 2020, 21, 2970. [Google Scholar] [CrossRef] [PubMed]
- Pe, T.; Khin Aung Cho, K.A. Amount of venom injected by Russell’s viper (Vipera russelli). Toxicon 1986, 24, 730–733. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Geary, B.T.; Baird, M.S. Evaluation of the contribution of platelets to clot strength by thromboelastography in rabbits: The role of tissue factor and cytochalasin D. Anesth. Analg. 2000, 91, 35–39. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | BSL | 15 | 30 | 45 | 60 | 75 | 90 | 105 | 120 | 135 | 150 | 165 | 180 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HR | 192 ± 39 | 185 ± 33 | 183 ± 40 | 185 ± 33 | 176 ± 37 | 171 ± 12 | 168 ± 14 | 169 ± 8 | 188 ± 35 | 177 ± 19 | 172 ± 17 | 178 ± 13 | 168 ± 22 |
| SpO2 | 98 ± 1 | 98 ± 1 | 98 ± 1 | 98 ± 2 | 96 ± 3 | 97 ± 2 | 98 ± 1 | 98 ± 1 | 98 ± 1 | 98 ± 1 | 97 ± 2 | 95 ± 2 | 98 ± 2 |
| Time | BSL | 15 | 30 | 45 | 60 | 75 | 90 | 105 | 120 | 135 | 150 | 165 | 180 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | 218 ± 21 | 222 ± 18 | 208 ± 36 | 194 ± 35 | 214 ± 29 | 204 ± 34 | 196 ± 39 | 209 ± 29 | 214 ± 25 | 212 ± 25 | 218 ± 22 | 216 ± 38 | 219 ± 22 |
| A + V | 194 ± 41 | 222 ± 32 | 242 ± 15 | 203 ± 43 | 218 ± 26 | 226 ± 29 | 228 ± 28 | 217 ± 33 | 213 ± 29 | 221 ± 20 | 220 ± 14 | 221 ± 19 | 228 ± 18 |
| Time | BSL | 15 | 30 | 45 | 60 | 75 | 90 | 105 | 120 | 135 | 150 | 165 | 180 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | 98 ± 2 | 97 ± 1 | 96 ± 3 | 98 ± 2 | 96 ± 3 | 96 ± 3 | 98 ± 2 | 98 ± 1 | 99 ± 1 | 98 ± 2 | 98 ± 2 | 98 ± 2 | 98 ± 1 |
| A + V | 98 ± 2 | 96 ± 2 | 96 ± 3 | 97 ± 2 | 98 ± 1 | 98 ± 2 | 95 ± 3 | 97 ± 2 | 97 ± 2 | 97 ± 2 | 98 ± 2 | 97 ± 2 | 97 ± 2 |
| Time | BSL | 15 | 30 | 45 | 60 | 75 | 90 | 105 | 120 | 135 | 150 | 165 | 180 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | 214 ± 20 | 224 ± 28 | 220 ± 32 | 216 ± 19 | 232 ± 21 | 231 ± 28 | 231 ± 15 | 236 ± 13 | 223 ± 22 | 227 ± 19 | 232 ± 23 | 242 ± 22 | 239 ± 19 |
| A + V | 239 ± 2 * | 246 ± 6 | 237 ± 11 | 226 ± 16 | 216 ± 14 | 219 ± 9 | 218 ± 13 | 217 ± 9 | 218 ± 14 | 220 ± 13 | 219 ± 7 | 224 ± 9 | 223 ± 15 |
| Time | BSL | 15 | 30 | 45 | 60 | 75 | 90 | 105 | 120 | 135 | 150 | 165 | 180 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V | 97 ± 2 | 98 ± 2 | 98 ± 1 | 98 ± 1 | 97 ± 2 | 97 ± 2 | 98 ± 2 | 98 ± 2 | 98 ± 1 | 98 ± 2 | 98 ± 1 | 98 ± 1 | 99 ± 1 |
| A + V | 99 ± 1 | 97 ± 2 | 96 ± 2 | 98 ± 1 | 98 ± 1 | 99 ± 1 | 99 ± 1 | 98 ± 1 | 98 ± 1 | 98 ± 2 | 99 ± 1 | 98 ± 2 | 99 ± 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, V.G. Novel Toxicodynamic Model of Subcutaneous Envenomation to Characterize Snake Venom Coagulopathies and Assess the Efficacy of Site-Directed Inorganic Antivenoms. Int. J. Mol. Sci. 2023, 24, 13939. https://doi.org/10.3390/ijms241813939
Nielsen VG. Novel Toxicodynamic Model of Subcutaneous Envenomation to Characterize Snake Venom Coagulopathies and Assess the Efficacy of Site-Directed Inorganic Antivenoms. International Journal of Molecular Sciences. 2023; 24(18):13939. https://doi.org/10.3390/ijms241813939
Chicago/Turabian StyleNielsen, Vance G. 2023. "Novel Toxicodynamic Model of Subcutaneous Envenomation to Characterize Snake Venom Coagulopathies and Assess the Efficacy of Site-Directed Inorganic Antivenoms" International Journal of Molecular Sciences 24, no. 18: 13939. https://doi.org/10.3390/ijms241813939
APA StyleNielsen, V. G. (2023). Novel Toxicodynamic Model of Subcutaneous Envenomation to Characterize Snake Venom Coagulopathies and Assess the Efficacy of Site-Directed Inorganic Antivenoms. International Journal of Molecular Sciences, 24(18), 13939. https://doi.org/10.3390/ijms241813939