
Four-and-a-Half LIM-Domain Protein 2 (FHL2) Induces Neuropeptide Y (NPY) in Macrophages in Visceral Adipose Tissue and Promotes Diet-Induced Obesity

 , ,
, ,  , ,
, ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
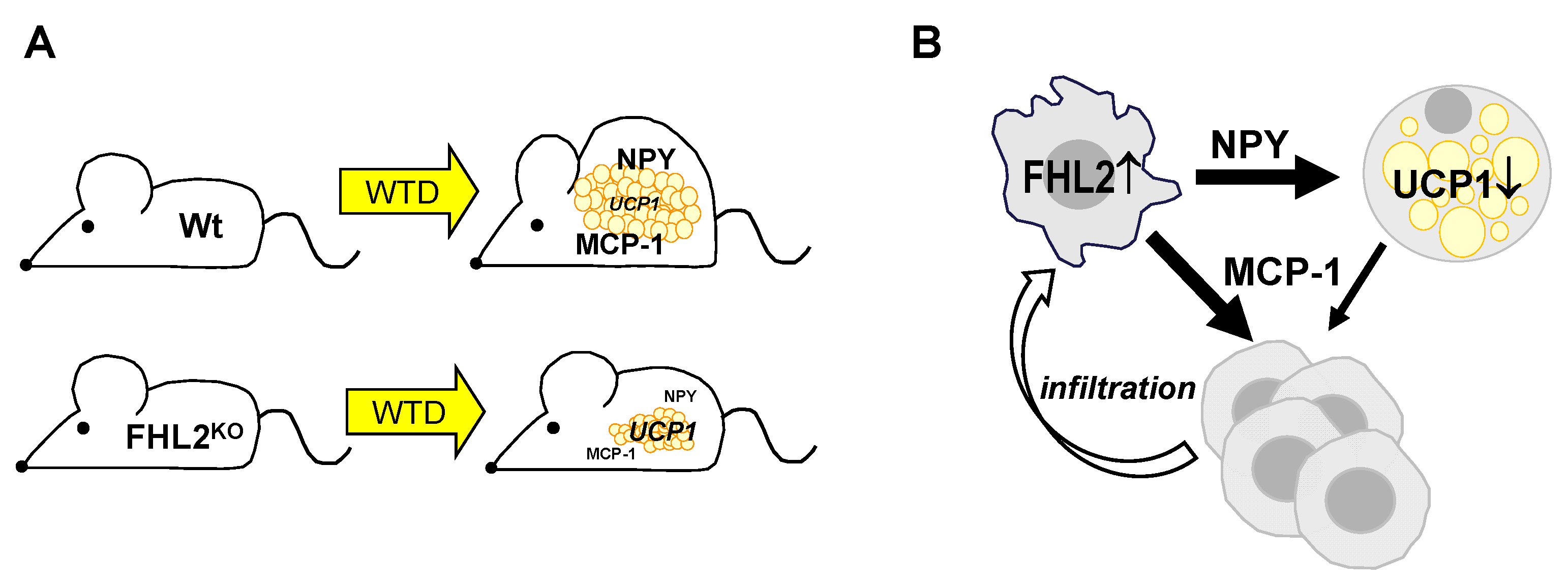
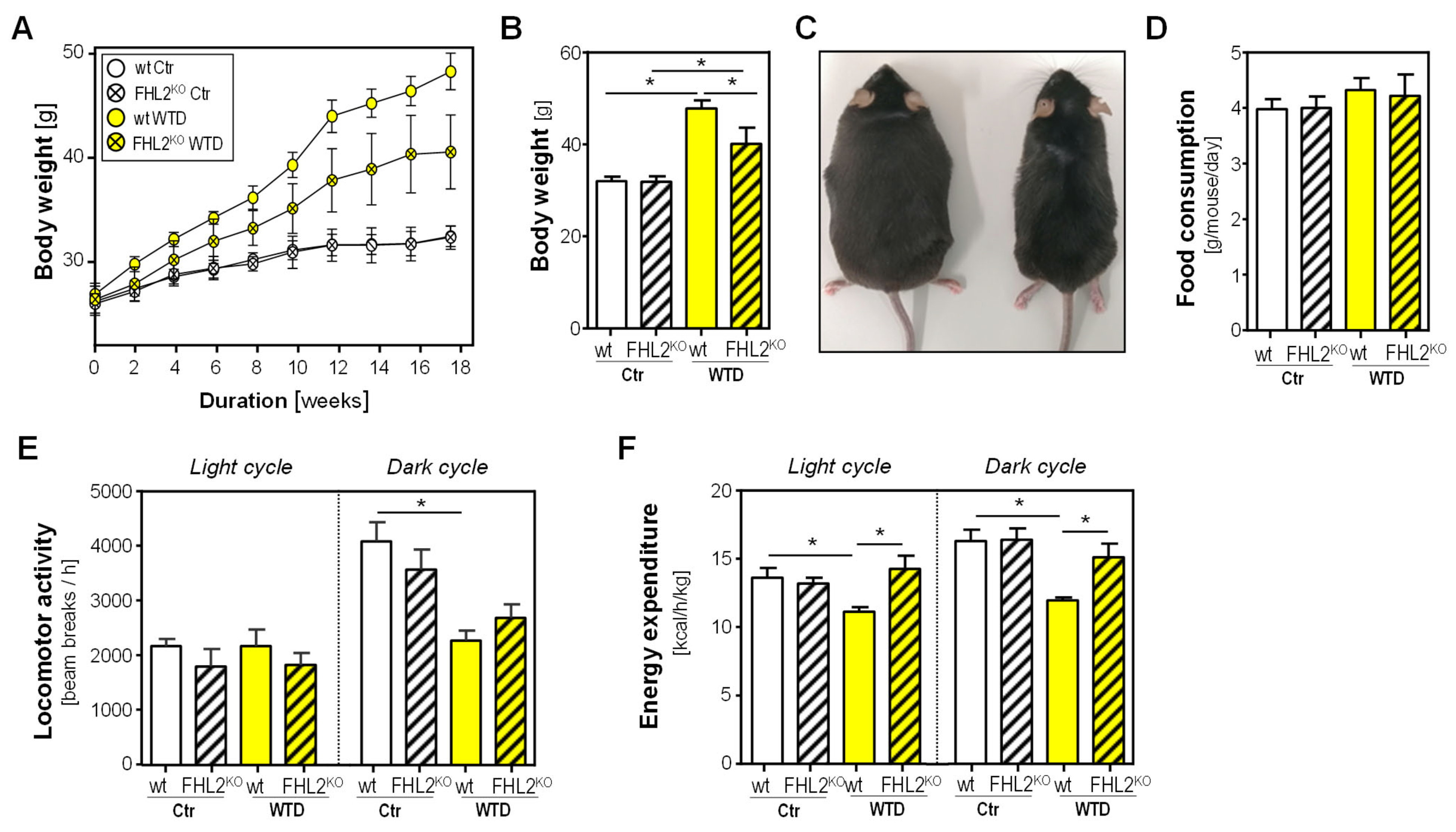
2.1. Effect of FHL2 Deficiency on Diet-Induced Obesity
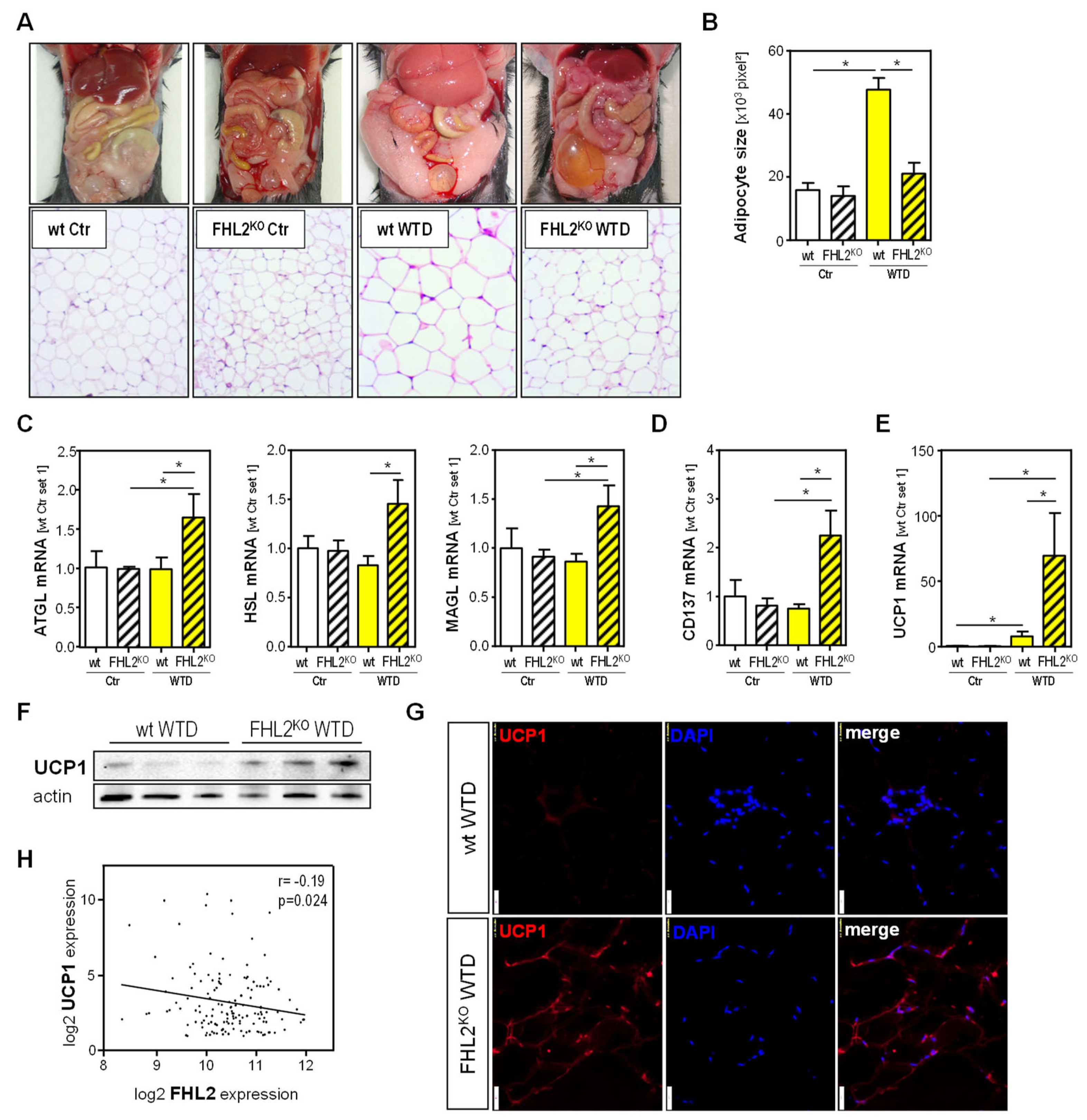
2.2. Effect of FHL2 Deficiency on Diet-Induced Expansion and Metabolic Activity of Visceral Adipose Tissue
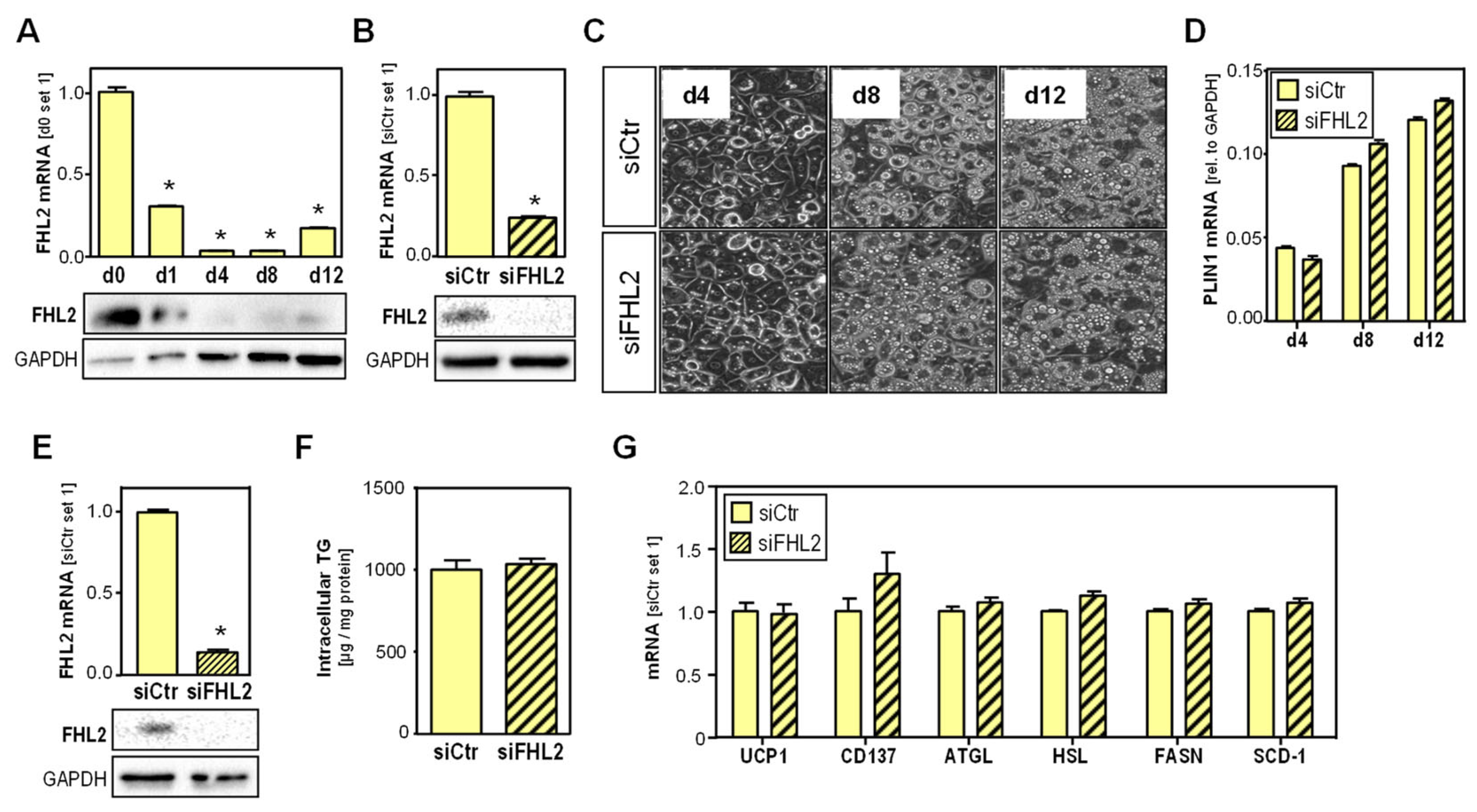
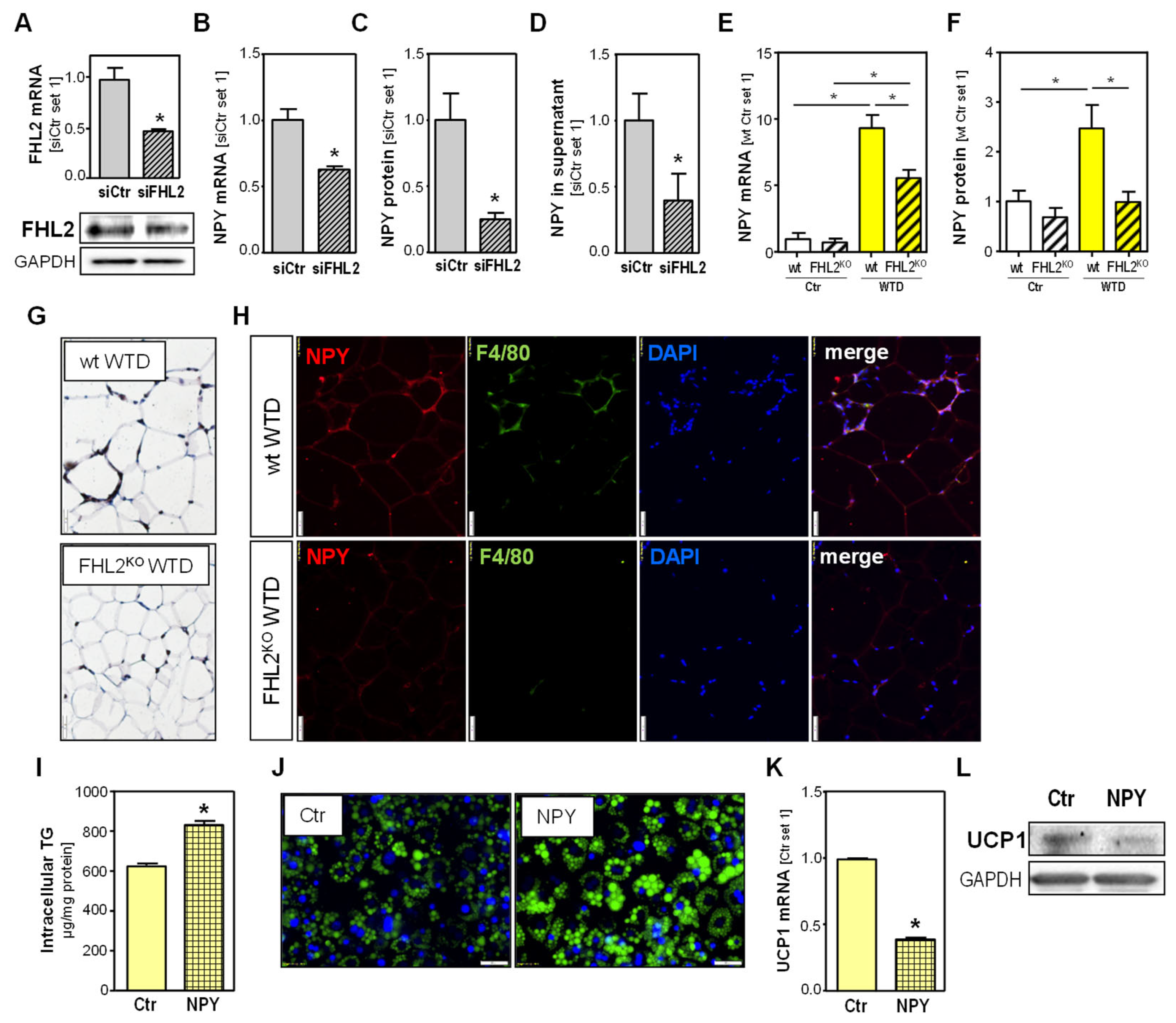
2.3. Effect of FHL2 Depletion in Adipocytes
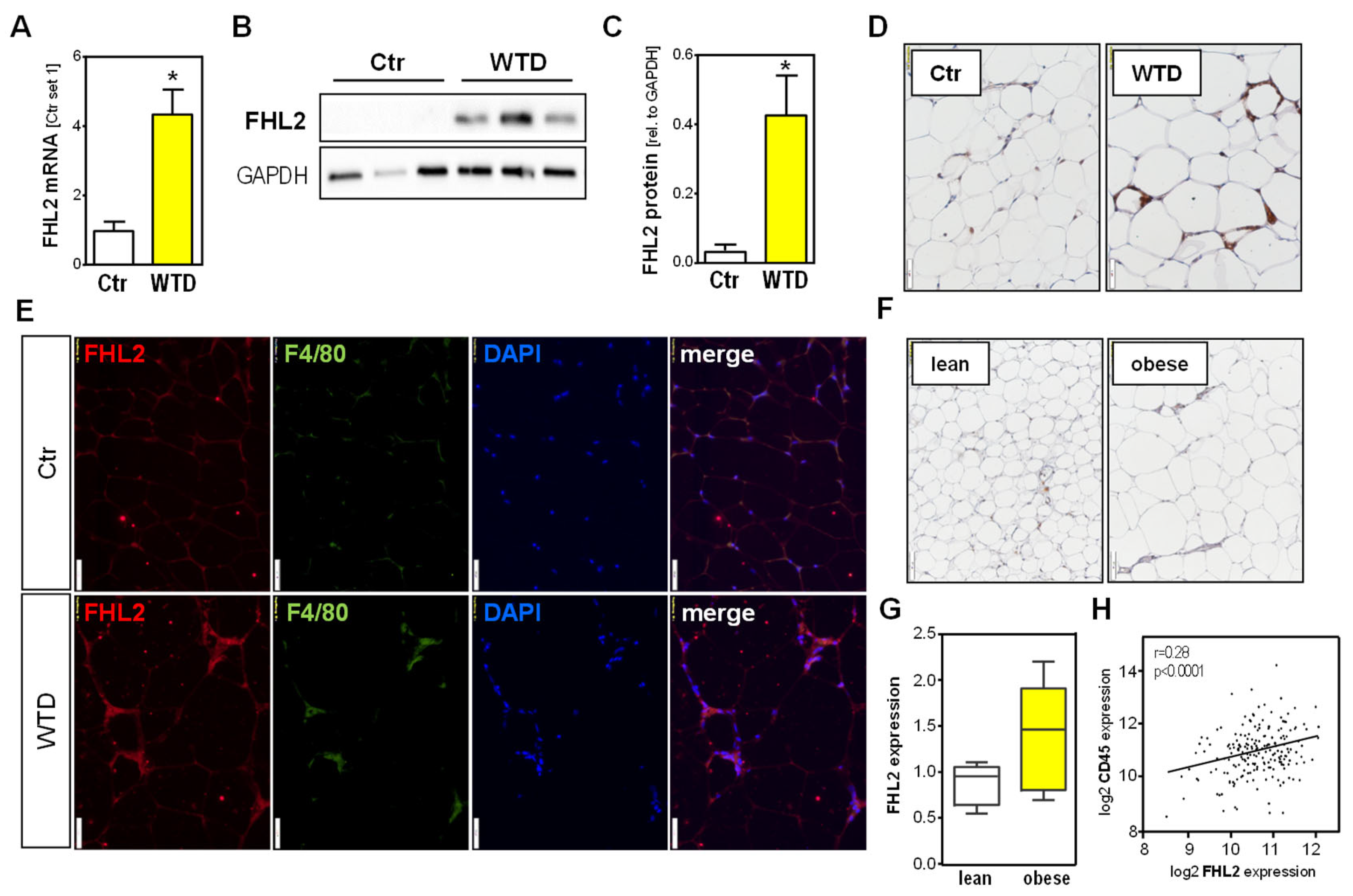
2.4. FHL2 Expression in Visceral Adipose Tissue
2.5. Effect of FHL2 Depletion in Macrophages
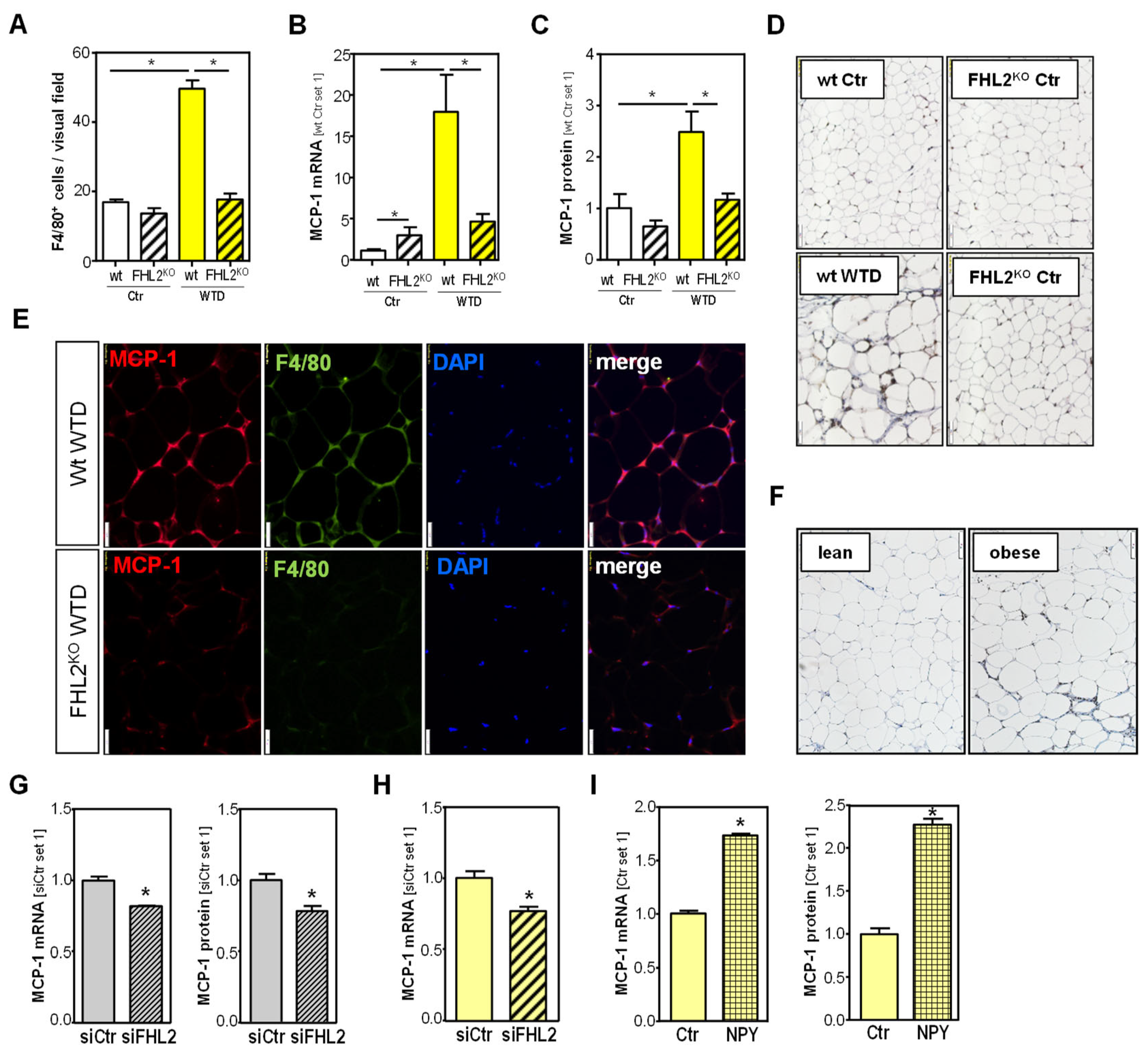
2.6. Effect of FHL2 Deficiency on Diet-Induced Inflammation of Visceral Adipose Tissue
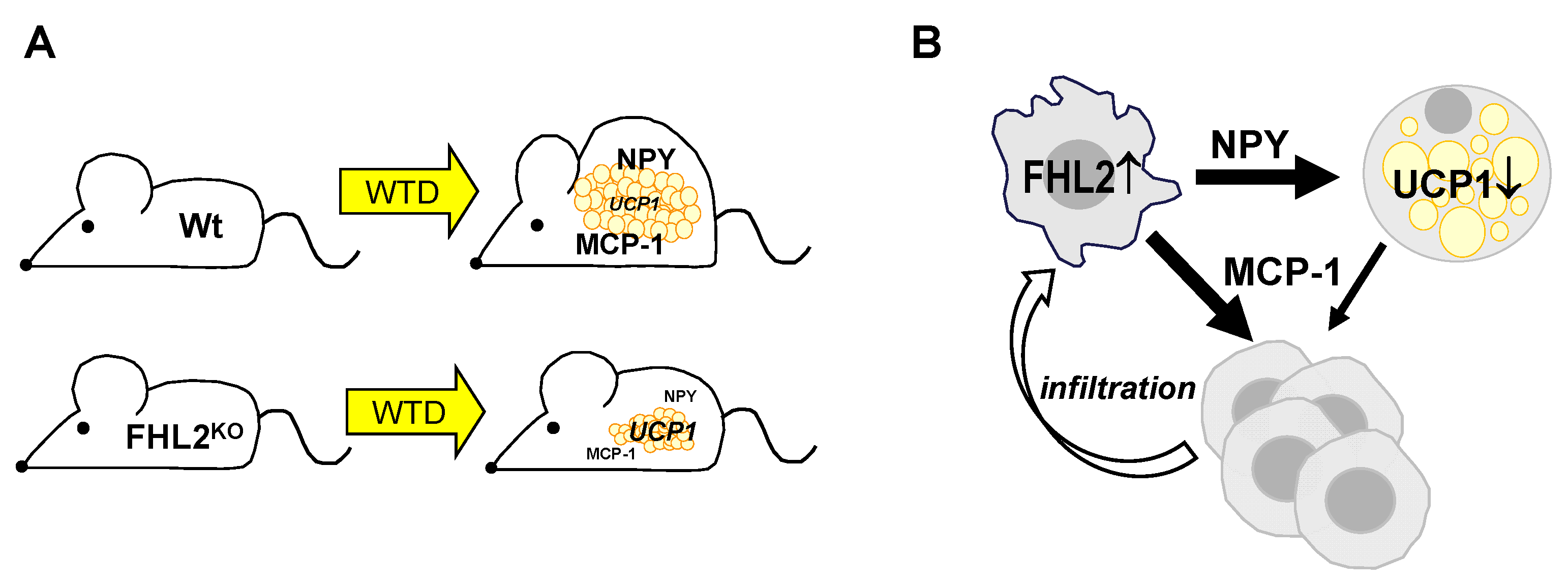
3. Discussion
4. Materials and Methods
4.1. Cells and Cell Culture
4.2. Animal Experiments
4.3. Human Visceral Adipose Tissue
4.4. Histological, Immunohistochemical, and Immunofluorescence Analysis
4.5. Analysis of Lipid Content
4.6. RNA Analysis
4.7. Protein Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loffler, M.C.; Betz, M.J.; Blondin, D.P.; Augustin, R.; Sharma, A.K.; Tseng, Y.H.; Scheele, C.; Zimdahl, H.; Mark, M.; Hennige, A.M.; et al. Challenges in Tackling Energy Expenditure as Obesity Therapy: From Preclinical Models to Clinical Application. Mol. Metab. 2021, 51, 101237. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose Tissue Remodeling and Obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef]
- Kuo, L.E.; Kitlinska, J.B.; Tilan, J.U.; Li, L.; Baker, S.B.; Johnson, M.D.; Lee, E.W.; Burnett, M.S.; Fricke, S.T.; Kvetnansky, R.; et al. Neuropeptide Y Acts Directly in the Periphery on Fat Tissue and Mediates Stress-Induced Obesity and Metabolic Syndrome. Nat. Med. 2007, 13, 803–811. [Google Scholar] [CrossRef]
- Ruohonen, S.T.; Pesonen, U.; Moritz, N.; Kaipio, K.; Roytta, M.; Koulu, M.; Savontaus, E. Transgenic Mice Overexpressing Neuropeptide Y in Noradrenergic Neurons: A Novel Model of Increased Adiposity and Impaired Glucose Tolerance. Diabetes 2008, 57, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Ruohonen, S.T.; Vähätalo, L.H.; Savontaus, E. Diet-Induced Obesity in Mice Overexpressing Neuropeptide Y in Noradrenergic Neurons. Int. J. Pept. 2012, 2012, 452524. [Google Scholar] [CrossRef]
- Park, S.; Fujishita, C.; Komatsu, T.; Kim, S.E.; Chiba, T.; Mori, R.; Shimokawa, I. Npy Antagonism Reduces Adiposity and Attenuates Age-Related Imbalance of Adipose Tissue Metabolism. FASEB J. 2014, 28, 5337–5348. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Komatsu, T.; Kim, S.E.; Tanaka, K.; Hayashi, H.; Mori, R.; Shimokawa, I. Neuropeptide Y Resists Excess Loss of Fat by Lipolysis in Calorie-Restricted Mice: A Trait Potential for the Life-Extending Effect of Calorie Restriction. Aging Cell 2017, 16, 339–348. [Google Scholar] [CrossRef]
- Tran, M.K.; Kurakula, K.; Koenis, D.S.; de Vries, C.J. Protein-Protein Interactions of the Lim-Only Protein Fhl2 and Functional Implication of the Interactions Relevant in Cardiovascular Disease. Biochim. Biophys. Acta 2016, 1863, 219–228. [Google Scholar] [CrossRef]
- Wixler, V. The Role of Fhl2 in Wound Healing and Inflammation. FASEB J. 2019, 33, 7799–7809. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Olivo, M.P.; Habibe, J.J.; Vos, M.; Ottenhoff, R.; Jongejan, A.; Herrema, H.; Zelcer, N.; Kooijman, S.; Rensen, P.C.; van Raalte, D.H.; et al. Four-and-a-Half Lim Domain Protein 2 (Fhl2) Deficiency Protects Mice from Diet-Induced Obesity and High Fhl2 Expression Marks Human Obesity. Metab. Clin. Exp. 2021, 121, 154815. [Google Scholar] [CrossRef]
- Engin, A.B. Adipocyte-Macrophage Cross-Talk in Obesity. Adv. Exp. Med. Biol. 2017, 960, 327–343. [Google Scholar] [PubMed]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef]
- Honecker, J.; Ruschke, S.; Seeliger, C.; Laber, S.; Strobel, S.; Pröll, P.; Nellaker, C.; Lindgren, C.M.; Kulozik, U.; Ecker, J.; et al. Transcriptome and Fatty-Acid Signatures of Adipocyte Hypertrophy and Its Non-Invasive Mr-Based Characterization in Human Adipose Tissue. EBioMedicine 2022, 79, 104020. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, K.; Rockstroh, D.; Wagner, I.V.; Weise, S.; Tauscher, R.; Schwartze, J.T.; Löffler, D.; Bühligen, U.; Wojan, M.; Till, H.; et al. Evidence of Early Alterations in Adipose Tissue Biology and Function and Its Association with Obesity-Related Inflammation and Insulin Resistance in Children. Diabetes 2015, 64, 1249–1261. [Google Scholar] [CrossRef]
- Sanchez-Gurmaches, J.; Hung, C.M.; Guertin, D.A. Guertin. Emerging Complexities in Adipocyte Origins and Identity. Trends Cell Biol. 2016, 26, 313–326. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, L.; Li, B.; Jiang, H.; Duan, Y.; Xie, Z.; Shuai, L.; Li, J.; Li, J. Amp-Activated Protein Kinase (Ampk) Regulates Energy Metabolism through Modulating Thermogenesis in Adipose Tissue. Front. Physiol. 2018, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Craft, B.; Hastie, M.; Repečka, K.; McDade, F.; Kamath, A.; Banerjee, A.; Luo, Y.; Rogers, D.; Brooks, A.N.; et al. Visualizing and Interpreting Cancer Genomics Data Via the Xena Platform. Nat. Biotechnol. 2020, 38, 675–678. [Google Scholar] [CrossRef]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte Death Defines Macrophage Localization and Function in Adipose Tissue of Obese Mice and Humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef]
- Zhang, W.; Cline, M.A.; Gilbert, E.R. Hypothalamus-Adipose Tissue Crosstalk: Neuropeptide Y and the Regulation of Energy Metabolism. Nutr. Metab. 2014, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Guan, H.; Arany, E.; Hill, D.J.; Cao, X. Neuropeptide Y Is Produced in Visceral Adipose Tissue and Promotes Proliferation of Adipocyte Precursor Cells Via the Y1 Receptor. FASEB J. 2008, 22, 2452–2464. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.I.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. Mcp-1 Contributes to Macrophage Infiltration into Adipose Tissue, Insulin Resistance, and Hepatic Steatosis in Obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef]
- Gerhardt, C.C.; Romero, I.A.; Cancello, R.; Camoin, L.; Strosberg, A.D. Chemokines Control Fat Accumulation and Leptin Secretion by Cultured Human Adipocytes. Mol. Cell. Endocrinol. 2001, 175, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Sartipy, P.; Loskutoff, D.J. Monocyte Chemoattractant Protein 1 in Obesity and Insulin Resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 7265–7270. [Google Scholar] [CrossRef]
- van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-Activated Brown Adipose Tissue in Healthy Men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef]
- Chao, P.T.; Yang, L.; Aja, S.; Moran, T.H.; Bi, S. Knockdown of Npy Expression in the Dorsomedial Hypothalamus Promotes Development of Brown Adipocytes and Prevents Diet-Induced Obesity. Cell Metab. 2011, 13, 573–583. [Google Scholar] [CrossRef]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef]
- Kolb, H. Obese Visceral Fat Tissue Inflammation: From Protective to Detrimental? BMC Med. 2022, 20, 494. [Google Scholar] [CrossRef]
- Gaculenko, A.; Gregoric, G.; Popp, V.; Seyler, L.; Ringer, M.; Kachler, K.; Wu, Z.; Kisel, W.; Hofbauer, C.; Hofbauer, L.C.; et al. Systemic Pparγ Antagonism Reduces Metastatic Tumor Progression in Adipocyte-Rich Bone in Excess Weight Male Rodents. J. Bone Miner. Res. 2021, 36, 2440–2452. [Google Scholar] [CrossRef]
- Bauer, S.; Wanninger, J.; Schmidhofer, S.; Weigert, J.; Neumeier, M.; Dorn, C.; Hellerbrand, C.; Zimara, N.; Schaffler, A.; Aslanidis, C.; et al. Sterol Regulatory Element-Binding Protein 2 (Srebp2) Activation after Excess Triglyceride Storage Induces Chemerin in Hypertrophic Adipocytes. Endocrinology 2011, 152, 26–35. [Google Scholar] [CrossRef]
- Sommer, J.; Dorn, C.; Gäbele, E.; Bataille, F.; Freese, K.; Seitz, T.; Thasler, W.E.; Büttner, R.; Weiskirchen, R.; Bosserhoff, A.; et al. Four-and-a-Half Lim-Domain Protein 2 (Fhl2) Deficiency Aggravates Cholestatic Liver Injury. Cells 2020, 9, 248. [Google Scholar] [CrossRef]
- Wixler, V.; Hirner, S.; Muller, J.M.; Gullotti, L.; Will, C.; Kirfel, J.; Gunther, T.; Schneider, H.; Bosserhoff, A.; Schorle, H.; et al. Deficiency in the Lim-Only Protein Fhl2 Impairs Skin Wound Healing. J. Cell Biol. 2007, 177, 163–172. [Google Scholar] [CrossRef]
- Dorn, C.; Engelmann, J.C.; Saugspier, M.; Koch, A.; Hartmann, A.; Müller, M.; Spang, R.; Bosserhoff, A.; Hellerbrand, C. Increased Expression of C-Jun in Nonalcoholic Fatty Liver Disease. Lab. Investig. 2014, 94, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Urbach, Y.K.; Raber, K.A.; Canneva, F.; Plank, A.C.; Andreasson, T.; Ponten, H.; Kullingsjo, J.; Nguyen, H.P.; Riess, O.; von Horsten, S. Automated Phenotyping and Advanced Data Mining Exemplified in Rats Transgenic for Huntington’s Disease. J. Neurosci. Methods 2014, 234, 38–53. [Google Scholar] [CrossRef] [PubMed]
- Parlee, S.D.; Lentz, S.I.; Mori, H.; MacDougald, O.A. Quantifying Size and Number of Adipocytes in Adipose Tissue. Methods Enzymol. 2014, 537, 93–122. [Google Scholar] [PubMed]
- Wobser, H.; Dorn, C.; Weiss, T.S.; Amann, T.; Bollheimer, C.; Büttner, R.; Schölmerich, J.; Hellerbrand, C. Lipid Accumulation in Hepatocytes Induces Fibrogenic Activation of Hepatic Stellate Cells. Cell Res. 2009, 19, 996–1005. [Google Scholar] [CrossRef]
- Targett-Adams, P.; Chambers, D.; Gledhill, S.; Hope, R.G.; Coy, J.F.; Girod, A.; McLauchlan, J. Live Cell Analysis and Targeting of the Lipid Droplet-Binding Adipocyte Differentiation-Related Protein. J. Biol. Chem. 2003, 278, 15998–16007. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| ATGL | CAACGCCACTCACATCTACGGA | CAGGTTGAAGGAGGGATGCAGA |
| β-actin | AGGCCAACCGTGAAAAGAT | GGCGTGAGGGAGAGCATA |
| CD137 | CTCGCTGCCCTGAGATCGAA | TCGGCTGTCCACCTATGCTG |
| FASN | ACAATGGACCCCCAGCTTCG | CAGACGCCAGTGTTCGTTCC |
| FHL2 | ACTGCCTGACCTGCTTCTGT | TTGCCTGGTTATGAAAGAAAA |
| GAPDH | AGCCTCGTCCCGTAGACAAA | TGATGGGCTTCCCGTTGATG |
| HSL | TGGCAGTGGTGTGTAACTAGGATT | TCGTGCGTAAATCCATGCTGTG |
| MAGL | CTGTGGCGGTAGTGGAATGC | CCCAGCTCCATGGGACACAA |
| MCP-1 | TGCAGGTCCCTGTCATGCTTC | TGGACCCATTCCTTCTTGGGGT |
| NPY | TGGCCAGATACTACTCCGCT | GCAGACTGGTTTCAGGGGAT |
| PLIN1 | CTTGGGCGTCTGCCTTACCTA | TTGCTGGCACCCTGTACACC |
| SCD-1 | AACAGTGCCGCGCATCTCTA | GAAGCCCAAAGCTCAGCTACTC |
| UCP1 | ACACCAAGGAAGGACCGACG | ATGACGTTCCAGGACCCGAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sommer, J.; Ehnis, H.; Seitz, T.; Schneider, J.; Wild, A.B.; Moceri, S.; Buechler, C.; Bozec, A.; Weber, G.F.; Merkel, S.; et al. Four-and-a-Half LIM-Domain Protein 2 (FHL2) Induces Neuropeptide Y (NPY) in Macrophages in Visceral Adipose Tissue and Promotes Diet-Induced Obesity. Int. J. Mol. Sci. 2023, 24, 14943. https://doi.org/10.3390/ijms241914943
Sommer J, Ehnis H, Seitz T, Schneider J, Wild AB, Moceri S, Buechler C, Bozec A, Weber GF, Merkel S, et al. Four-and-a-Half LIM-Domain Protein 2 (FHL2) Induces Neuropeptide Y (NPY) in Macrophages in Visceral Adipose Tissue and Promotes Diet-Induced Obesity. International Journal of Molecular Sciences. 2023; 24(19):14943. https://doi.org/10.3390/ijms241914943
Chicago/Turabian StyleSommer, Judith, Hanna Ehnis, Tatjana Seitz, Julia Schneider, Andreas B. Wild, Sandra Moceri, Christa Buechler, Aline Bozec, Georg F. Weber, Susanne Merkel, and et al. 2023. "Four-and-a-Half LIM-Domain Protein 2 (FHL2) Induces Neuropeptide Y (NPY) in Macrophages in Visceral Adipose Tissue and Promotes Diet-Induced Obesity" International Journal of Molecular Sciences 24, no. 19: 14943. https://doi.org/10.3390/ijms241914943
APA StyleSommer, J., Ehnis, H., Seitz, T., Schneider, J., Wild, A. B., Moceri, S., Buechler, C., Bozec, A., Weber, G. F., Merkel, S., Beckervordersandforth, R., Steinkasserer, A., Schüle, R., Trebicka, J., Hartmann, A., Bosserhoff, A., von Hörsten, S., Dietrich, P., & Hellerbrand, C. (2023). Four-and-a-Half LIM-Domain Protein 2 (FHL2) Induces Neuropeptide Y (NPY) in Macrophages in Visceral Adipose Tissue and Promotes Diet-Induced Obesity. International Journal of Molecular Sciences, 24(19), 14943. https://doi.org/10.3390/ijms241914943