Abstract
The genus Henricia is known to have intraspecific morphological variations, making species identification difficult. Therefore, molecular phylogeny analysis based on genetic characteristics is valuable for species identification. We present complete mitochondrial genomic sequences of Henricia longispina aleutica, H. reniossa, and H. sanguinolenta for the first time in this study. This study will make a significant contribution to our understanding of Henricia species and its relationships within the class Asteroidea. Lengths of mitochondrial genomes of the three species are 16,217, 16,223, and 16,194 bp, respectively, with a circular form. These genomes contained 13 protein-coding genes, two ribosomal RNA genes, 22 transfer RNA genes, and a D-loop. The gene order and direction aligned with other asteroid species. Phylogenetic relationship analysis showed that our Henricia species were in a monophyletic clade with other Henricia species and in a large clade with species (Echinaster brasiliensis) from the same family. These findings provide valuable insight into understanding the phylogenetic relationships of species in the genus Henricia.
1. Introduction
The mitochondrial genome is a double-stranded circular molecule typically ranging from 14,000 bp to 17,000 bp in length. It contains a gene complement encoding 13 protein-coding genes (PCGs), 22 transfer RNA genes (tRNA), and two ribosomal RNA genes (rRNA) [1]. Mitogenome sequencing is widely employed to elucidate phylogenetic and evolutionary relationships among species [2,3]. The mitogenome provides valuable insight into rearrangement patterns and phylogenetic relationships [4]. The class Asteroidea de Blainville, 1830 (sea stars), belonging to the phylum Echinodermata Klein, 1778, represents the second most diverse class in this phylum, encompassing 1800 species categorized into 38 families [5]. These intriguing creatures inhabit oceans worldwide, with tropical Atlantic and Indo-Pacific regions showcasing the highest diversity across various depths [6]. Asteroids have garnered significant attention due to their unique chemical structures, particularly their steroid metabolites, which offer rich opportunities for exploration. Starfish are also renowned for their wide range of biological effects, including antiviral, cytotoxic, antifungal, antibacterial, anti-inflammatory, anticancer, analgesic, and neurogenic actions [7,8].
Genus Henricia Gray, 1840, is a large genus that belongs to the family Echinasteridae Verrill, 1870. Henricia is known for its wide-ranging geographical distribution and intraspecific morphological variations. These variations are due to differences in environmental conditions that can result in phenotypic plasticity, making it difficult to identify species within the genus [9,10,11]. Henricia species are distributed in cold water. They exhibit great diversity in high latitudinal regions and deep-sea settings in the North Pacific Ocean, the Atlantic Ocean, and the East Sea of Korea [12]. According to recent studies by Mah, Henricia is currently composed of 97 species [13], including 21 species reported in South Korea [14].
Many species within the genus Henrica share similar characteristics, making it difficult to identify them based on their appearance alone [9,12]. An alternative approach is to use molecular data such as DNA barcoding to simplify species diagnosis. This method has become more common when species diagnosis is challenging due to limited or variable diagnostic morphological characteristics [15,16,17,18]. In this study, we used a mitogenomic approach that could provide more accurate data than single gene analysis. To construct our phylogenetic analysis of Henricia species, we utilized complete mitochondrial genomes of three Henricia species and 39 other species within the class Asteroidea. By examining other species within the class Asteroidea, we aimed to gain a comprehensive understanding of inter-specific phylogenetic relationships.
2. Results
2.1. Mitogenome Features and AT/GC-Skew
We investigated the mitogenomes of three Henricia species, H. longispina aleutica, H. reniossa, and H. sanguinolenta. Our findings showed that the lengths of their mitogenomes were 16,217, 16,194, and 16,223 bp, respectively. Each of these mitogenomes included 13 PCGs, 22 tRNAs, and two rRNAs. The base composition analysis of H. longispina aleutica revealed the following percentages: A (36.2%), T (26.2%), C (24.1%), and G (13.5%). The complete mitochondrial genome of H. longispina aleutica exhibited a preference for GC content (37.5%). Additionally, all 13 PCGs, as well as tRNA and rRNA genes, demonstrate anti-G bias. Similarly, base composition analysis of H. reniossa showed the following percentages: A (35.4%), T (25.7%), C (25.0%), and G (13.9%). The complete mitochondrial genome of H. reniossa displays a preference for GC content (38.8%). All 13 PCGs, as well as the tRNA and rRNA genes, also demonstrated an anti-G bias. Lastly, the base composition analysis of H. sanguinolenta showed the following percentages: A (35.6%), T (25.8%), C (24.6%), and G (13.9%). The complete mitochondrial genome of H. sanguinolenta also exhibited a preference for GC content (38.5%). All 13 PCGs, tRNA, and rRNA genes also demonstrate an anti-G bias (Table 1).

Table 1.
Nucleotide compositions of complete mitochondrial genomes of the three Henricia species in this study.
2.2. PCG Characteristics
Mitochondrial genomes of the three Henricia species were analyzed. PCGs in H. longispina aleutica and H. sanguinolenta spanned 11,473 bp, while H. reniossa spanned 11,476 bp. NADH5 was the longest at 1878 bp, while ATP8 was the shortest at 165 bp in H. longispina aleutica and H. sanguinolenta. NADH5 was the longest PCG, consisting of 1881 bp, while ATP8 was the shortest at 165 bp in H. reniossa (Figure 1, Figure 2 and Figure 3, Table 2). These PCGs collectively encoded 3824 amino acids for H. longispina aleutica and H. sanguinolenta and 3825 amino acids for H. reniossa.
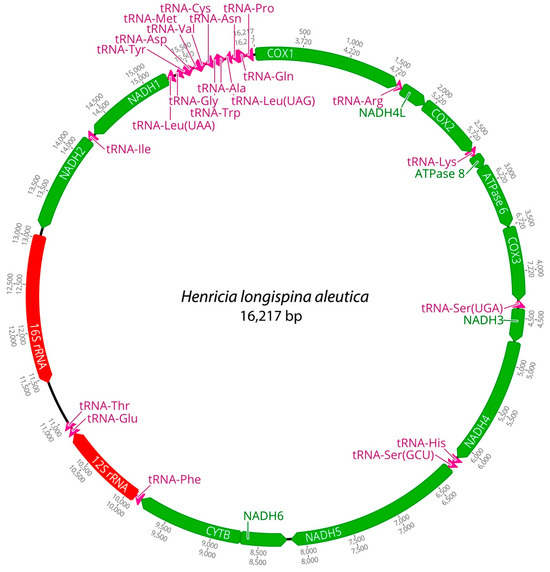
Figure 1.
Complete mitochondrial genome of Henricia longispina aleutica in this study.
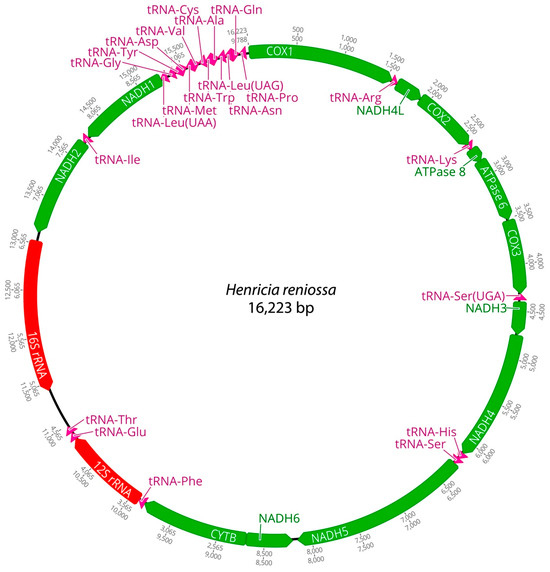
Figure 2.
Complete mitochondrial genome of Henricia reniossa in this study.
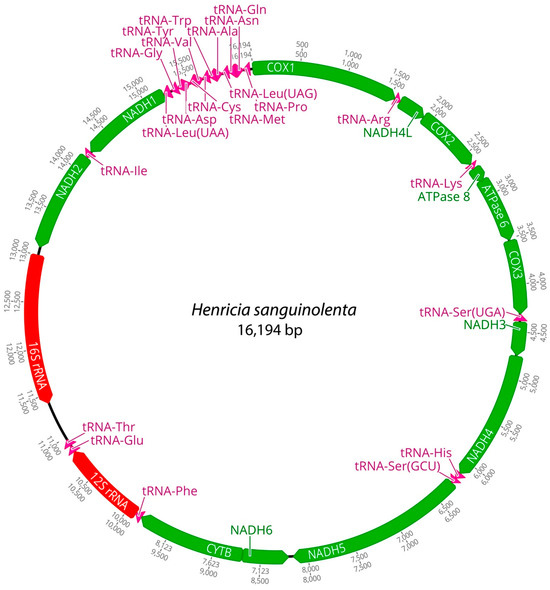
Figure 3.
Complete mitochondrial DNA of Henricia sanguinolenta in this study.
Table 2.
Mitochondrial genes and associated features of the three Henricia species in this study (intergenic space (IGS) is described as intergenic (+) or overlapping nucleotides).
The majority of the PCG genes of H. longispina aleutica, H. reniossa, and H. sanguinolenta began with the start codon ATG (Table 2). However, NAD4L and NADH3 of H. longispina aleutica and H. sanguinolenta started with ATC and ATT, respectively. In H. reniossa, start codons for NAD4L, NADH3, NADH2, and NADH1 were ATC, ATT, GTG, and GTG, respectively. The most common stop codon in the three Henricia species was TAA. However, COX1 and NADH4 utilized TGA, and CYTB employed TAG in H. longispina aleutica and H. sanguinolenta. COX1 and CYTB utilized TGA as a stop codon in H. reniossa (Table 2). The positive strand of H. longispina aleutica, H. reniossa, and H. sanguinolenta mitogenome contained 10 PCGs, while the negative strand contained NADH1, NADH2, and NADH6.
2.3. Relative Synonymous Codon Usage
Codons with relative synonymous codon usage (RSCU) values above 1.00 indicated a positive codon usage bias (CUB). They were considered abundant codons. Those with RSCU values below 1.00 indicated a negative CUB. They were considered less abundant codons [19]. Among the 13 PCGs of H. longispina aleutica, H. reniossa, and H. sanguinolenta, the most frequently used codons encoded serine (3.2%) for H. longispina aleutica, serine (2.56%) for H. reniossa, and serine (3.2%) for H. sanguinolenta had the most frequently encoded amino acids. Alanine, arginine, glutamate, glycine, and glutamine were the lowest frequencies among the three Henricia species (Table 3).
Table 3.
Results from the relative synonymous codon usage (RSCU) analysis for PCGs of the mitochondrial genomes of the three Henricia species in this study.
2.4. Characteristics of rRNA and tRNA Genes
The analysis of H. longispina aleutica, H. reniossa, and H. sanguinolenta revealed that their mitochondrial genomes encoded two rRNA genes: 12S and 16S rRNA. The 12S rRNA gene was located on the positive strand, while the 16S rRNA gene was situated on the negative strand of all three Henricia species (Figure 1, Figure 2 and Figure 3, Table 2). The 12S rRNA gene had a length of 931, 922, or 928 bp. In contrast, the 16S rRNA gene spanned 1585, 1594, and 1607 bp, respectively. GC contents of H. longispina aleutica, H. reniossa, and H. sanguinolenta rRNAs were 37.4%, 37.7%, and 38.0%, respectively (Table 2). The 12S rRNA gene was positioned between two tRNA genes, phenylalanine, and glutamic acid, while the 16S rRNA gene resided between NADH2 and the D-loop region of H. longispina aleutica, H. reniossa, and H. sanguinolenta (Figure 1, Figure 2 and Figure 3, Table 2).
A total of 22 tRNA genes were identified within the three Henricia mitogenomes, 11 of which were encoded on the positive strand. Lengths of these tRNA genes ranged from 67 bp to 72 bp, with tRNA-Arg being the shortest and tRNA-Cys being the longest in H. longispina aleutica. In H. reniossa, tRNA genes ranged from 67 bp to 73 bp, with tRNA-Arg being the shortest and tRNA-Cys being the longest. Similarly, in H. sanguinolenta, tRNA-Lys was the shortest at 57 bp, while tRNA-Ile was the longest at 73 bp (Table 2). H. longispina aleutica, H. reniossa, and H. sanguinolenta tRNAs had GC contents of 35.1%, 36.2%, and 35.5%, respectively.
2.5. Phylogenetic Relationships and Gene Arrangement
Within class Asteroidea, a clade consisting of 42 species from six orders and 23 families was observed, confirming the systematic classification (Figure 4). The phylogenetic tree displayed a distinct separation of 42 asteroid mitogenomes from the outgroup. The three Henricia species in this study (H. longispina aleutica, H. reniossa, and H. sanguinolenta) belonged to the clade of family Echinasteridae, forming a monophyletic group alongside other Henrica species (H. leviuscula and H. pachyderma) and Echinaster brasiliensis from the same family. Within this clade, H. reniossa was a sister taxon of H. leviuscula, with a 100% bootstrap support value. Similarly, H. longispina aleutica was a sister taxon of H. sanguinolenta, with a 100% bootstrap support value as well (Figure 4).
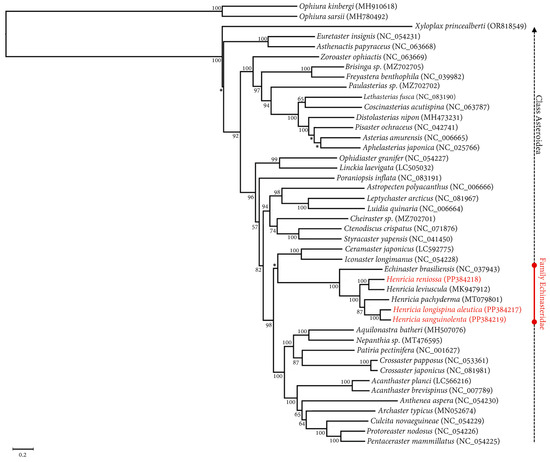
Figure 4.
The maximum likelihood tree of the 3 Henricia species of this study, 42 asteroid species, and two ophiuroids as an outgroup based on nucleotide sequences of 13 PCGs. The asterisk (*) denotes a bootstrap support value under 50%.
3. Materials and Methods
3.1. Sample Collection
Henricia specimens were collected from several locations in the East Sea, Korea. They were collected by trimix and scuba diving (Table 4). All specimens were preserved in an ethyl alcohol solution (>95%) immediately after collection. These preserved specimens were stored at the Marine Echinoderms Resources Bank of Korea (MERBK) at Sahmyook University and assigned voucher numbers (Table 4).
Table 4.
Information collected and voucher number of Henricia species in this study.
3.2. Species Identification Based on the Morphological Characteristics
The genus Henricia Gray, 1840, in Korea, was primarily classified into two groups: the imbricated group and the reticulated group [14]. Among the specimens examined in this study, H. sanguinolenta (O.F. Müller, 1776) and H. longispina aleutica Fisher, 1911, were categorized under the reticulated group, while H. reniossa asiatica Djakonov, 1958, was assigned to the imbricated group. Subsequently, detailed morphological analyses were conducted on the studied specimens, comparing them with closely related species. The slender-armed H. longispina aleutica was compared to H. pacifica Hayashi, 1940, to elucidate morphological similarities and differences. Additionally, variations in the characteristics of H. reniossa asiatica were carefully examined in comparison to H. reniossa reniossa Hayashi, 1940. For further details on the examination of the studied specimens, refer to [14,20,21].
3.3. DNA Extraction and Mitochondrial DNA Amplification
Genomic DNAs were extracted from gonadal tissues using a DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. Mitochondrial DNA amplification and analysis followed the method described by Lee and Shin [19]. Mitochondrial DNA amplification was done using the REPLI-g Mitochondrial DNA Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. The quality and concentration of amplified DNAs were assessed using a Nanodrop One-C (Thermo Fisher Scientific, Waltham, MA, USA). Next-generation sequencing (NGS) analysis was performed using genome analysis units at the National Instrumentation Center for Environmental Management at Seoul National University in Korea. A genomic library was constructed from the genomic DNA using a Kapa Hyper Prep Kit (Kapa Biosystems, Woburn, MA, USA), using paired-end reading, which was followed by NGS on the Illumina Hi-Seq 2500 platform (San Diego, CA, USA). The contigs of the mitogenome were assembled using the de novo assembly method on Geneious Prime 2023.1.1 (Biomatters Ltd., Auckland, New Zealand).
3.4. Mitogenome Annotation and Sequence Analysis
Complete mitogenomes of H. longispina aleutica, H. reniossa, and H. sanguinolenta were annotated by referencing three mitogenomes of closely related taxa from the family Echinasteridae available in the National Center for Biotechnology Information (NCBI) database (Table 5). Contigs of the mitogenome were assembled using the de novo assembly method with Geneious v.11.1.5 (Biomatters Ltd., Auckland, New Zealand). Subsequently, the complete mitogenome was annotated using the same software. tRNA genes were identified using the ARWEN website (http://130.235.244.92/ARWEN/ (accessed on 1 February 2024)) [20] and tRNAscan-SE online [21] using the “Invertebrate Mito” search mode. Geneious v.11.1.5 was also used to calculate the DNA base composition and codon preference of H. longispina aleutica, H. reniossa, and H. sanguinolenta mitogenomes. DNA base preference was determined using the following formulas: GC-skew = (G − C)/(G + C) and AT-skew = (A − T)/(A + T).
Table 5.
Mitochondrial genomes of the phylum Echinodermata in this study, including three newly reported mitogenomes of Henricia longispina aleutica, H. reniossa, and H. sanguinolenta.
3.5. Phylogenetic Analysis
Phylogenetic analysis of the 13 PCGs in this study utilized the maximum likelihood (ML) method through RAxML 8.2 [43]. The dataset used for analysis consisted of 41 complete mitogenomes obtained from NCBI. This dataset included 42 species of class Asteroidea, including H. longispina aleutica, H. reniossa, and H. sanguinolenta, and two species of class Ophiuroidea, which served as the outgroup (Table 2). Alignment of mitogenome sequences was done using MAFFT [44]. The best-fit substitution model for the nucleotide dataset comprising 13 PCGs was determined using jModelTest 2.1.1 [45,46]. The selected substitution model for this dataset was GTR + I + G. For ML analysis, bootstrap resampling with 1000 iterations was conducted using the rapid option.
4. Discussion
Phylogenetic analysis is a valuable tool for identifying biological characteristics and studying species relationships and evolutionary history [47]. In previous investigations, morphological analyses were performed to identify species and determine evolutionary relationships. Subsequently, morphological analysis has been found to be insufficient for identifying species because morphological properties change faster than molecular analysis in response to changing geographical factors, environment, and climate. In addition, the damage may happen to the specimen’s body [48]. Therefore, mitochondrial DNA analysis, widely used in phylogenetic studies [49], was performed in this study to identify species. Due to its faster evolutionary rate than nuclear genetic markers, mtDNA is highly polymorphic. Thus, it can be used effectively as a DNA marker for breed identification [50].
Henricia species are spread over a wide range of geographic locations, meaning that different populations may encounter different environmental conditions. This can result in variations in physical characteristics, making it difficult to identify species within the genus. A combination of morphological and molecular phylogeny studies can be used for species identification to overcome this challenge.
Chichvarkhin [51] discovered seven different species of Henricia in Vostok Bay and two species from an adjacent area. These species were identified using morphological characteristics and DNA barcoding, which utilized partial mitochondrial gene (16S rRNA) sequences. Phylogenetic relationships of 17 species (H. alexeyi, H. compacta, H. densispina, H. djakonovi, H. granulifera, H. hayashii, H. lineata, H. nipponica, H. obesa, H. oculate, H. ohshimai, H. olga, H. pacifica, H. sanguinolenta, H. sp, H. tumida, and H. uluudax) and 39 specimens were determined using the neighbor-joining method (NJ). Results indicated that H. sanguinolenta formed a monophyletic clade and a large clade with H. hayashii, H. obesa, and H. compacta, with a 63% bootstrap support value. Additionally, H. djakonovi demonstrated a close relationship with H. tumida, with a 67% bootstrap support value. However, Ubagan [52] studied the molecular phylogeny and morphological characteristics of the species Henricia djakonovi using ten species of the genus Henricia (H. epiphysialis, H. leviuscula, H. nipponica, H. oculata, H. perforate, H. regularis, H. reniossa, H. reticulata, H. sanguinolenta, and H. djakonovi). Phylogenetic relationships were identified using ML based on mitochondrial DNA gene cytochrome c oxidase subunit I (COI). Results showed that H. djakonovi formed a monophyletic clade with a 78% bootstrap support value, while H. sanguinolenta showed a sister relationship with H. regularis, with low bootstrap support value. H. reniossa showed a sister relationship with H. nipponica, with a low bootstrap support value. H. leviuscula had a monophyly clade with a 100% bootstrap support value.
Scientists often use a partial gene of the mitochondrial genome, such as 16S rRNA and COI, to establish evolutionary relationships between species. However, results from these methods can differ and relationships between species can be unstable. Despite this, mitochondrial DNA is crucial for understanding evolutionary biology and tracing relationships between populations [1]. In this study, we successfully obtained the first complete mitochondrial genomes for H. longispina aleutica, H. reniossa, and H. sanguinolenta with GenBank accession numbers of PP384217, PP384218, and PP384219, respectively.
Lee and Shin [31] examined the complete mitochondrial genome of H. leviuscula. They reported phylogenetic relationships using ML based on 13 PCGs from 11 species of class Asteroidea (Acanthaster brevispinus, A. planci, Aphelasterias japonica, Asterias amurensis, Echinaster (othilia) brasiliensis, Aquilonastra bather, Distolasterias nipon, H. leviuscula, Luidia quniaria, Patiria pectinifera, and Astropecten polyacanthus). Results indicated that H. leviuscula had a close relationship with E. brasiliensis, with a 100% bootstrap support value. Similarly, the phylogenetic relationship of H. pachyderma has been analyzed using the ML approach based on 13 PCGs of 32 complete mitochondrial genomes of echinoderms. Of them, 16 contained asteroids. Results showed that H. pachyderma was more closely related to H. leviuscula than E. brasiliensis, with a strong bootstrap support value of 100%. It formed a large clade with E. brasiliensis with a 92% bootstrap support value [32].
As part of our investigation, we analyzed the phylogenetic relationships of 42 species belonging to the class Asteroidea, which included six species from the family Echinasteridae (E. brasiliensis, H. leviuscula, H. longispina aleutica, H. pachyderma, H. reniossa, and H. sanguinolenta). We performed ML analysis based on 13 PCGs. Results showed that H. longispina aleutica was closely related to H. sanguinolenta with a 100% bootstrap support value. Similarly, H. reniossa was closely related to H. leviuscula with a 100% bootstrap support value. The family Echinasteridae was identified as a monophyletic clade separate from the remaining families within the class Asteroid (Figure 4). Furthermore, the molecular study showed that Henricia species also formed a monophyletic clade within the family Echinasteridae. While morphologically similar, the seastar genus Henricia Gray, 1840, was more complex than other asteroid groups due to its highly variable characteristics, ability to interbreed, and production of intermediates [53,54]. The main diagnostic characteristics of Henricia can be determined by examining its abactinal and actinal morphological features. This includes observing the shape and number of abactinal and actinal spines, the structure of abactinal and actinal skeletons, and the number of ambulacral spines present. The abactinal skeleton of some Henricia species is typically arranged in a closely interlocking pattern, whereas others have a more open, reticulated structure [55]. Therefore, morphological features can vary within Henricia species. H. longispina aleutica Fisher, 1911 has a denuded abactinal spine. Its abactinal skeleton consists of lobed and elongated shapes that come together to form a rhomboid coarse meshwork of different sizes [20]. H. reniossa Hayashi, 1940, has a closely meshed dorsal skeleton with five or six long and stout inner spines. Its abactinal plates are arranged in several longitudinal series exclusively at the proximal abactinolateral section of the arms. H. sanguinolenta Fisher, 1911, has a weakly reticulated structure with narrow meshes in its abactinal skeleton. Its abactinal plate is small, with one to five abactinal papulae [20].
Findings from previous studies and mitogenomes uncovered in this research augment existing genomic resources for use in further evolutionary research on the Asteroidea class and beyond. These findings will be instrumental in conservation genetics.
5. Conclusions
In this study, we successfully determined complete mitogenomes and investigated the phylogenetic relationships of three species in the genus Henricia: H. longispina aleutica, H. reniossa, and H. sanguinolenta. Mitogenomes of these species comprised 13 protein-coding genes (PCGs), two rRNA genes, 22 tRNA genes, and a non-coding region. The total lengths of these complete mitogenomes were 16,217, 16,223, and 16,194 bp, respectively, consistent with reported lengths of other published echinoderm species. This study provides valuable insight into the phylogenetic and evolutionary relationships of genus Henricia and other echinoderm species. Additionally, the complete mitochondrial genomes of this study could be used to identify H. longispina aleutica, H. reniossa, and H. sanguinolenta more easily and provide further support for morphological identification under limited information by offering insight gained from mtDNA analysis.
Author Contributions
Conceptualization, M.A. and T.L.; methodology, M.A. and T.L.; software, M.A. and T.L.; validation, M.A.; formal analysis, M.A. and T.L.; investigation, M.A. and T.L.; resources, T.L.; data curation, M.A.; writing—original draft preparation, M.A. and T.L.; writing—review and editing, M.A., H.J. and T.L.; visualization, M.A.; supervision, H.J. and T.L.; project administration, T.L.; funding acquisition, T.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the management of Marine Fishery Bio-resources Center (2024), funded by the National Marine Biodiversity Institute of Korea (MABIK). It was also supported by a grant (No. 2021R1I1A2058017) of the Basic Science Research Program through the National Research Foundation (NRF) funded by the Ministry of Education, Republic of Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The genome sequence data that support the findings of this study are openly available in GenBank of NCBI at https://www.ncbi.nlm.nih.gov (accessed on 20 February 2024), under accession nos. PP384217, PP384218, and PP384219.
Acknowledgments
We thank Michael Dadole Ubagan (Marine Biological Resource Institute of Sahmyook University) for advising on morphological identification in this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Rubinoff, D.; Cameron, S.; Will, K. A genomic perspective on the shortcomings of mitochondrial DNA for “barcoding” identification. J. Hered. 2006, 97, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Dool, S.E.; Puechmaille, S.J.; Foley, N.M.; Allegrini, B.; Bastian, A.; Mutumi, G.L.; Maluleke, T.G.; Odendaal, L.J.; Teeling, E.C.; Jacobs, D.S. Nuclear introns outperform mitochondrial DNA in inter-specific phylogenetic reconstruction: Lessons from horseshoe bats (Rhinolophidae: Chiroptera). Mol. Phylogenet. Evol. 2016, 97, 196–212. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhou, C.; Tran, N.T.; Sun, Z.; Wu, J.; Ge, H.; Lu, Z.; Zhong, C.; Zhu, Z.; Yang, Q. Comparison of the complete mitochondrial genome of Phyllophorus liuwutiensis (Echinodermata: Holothuroidea: Phyllophoridae) to that of other sea cucumbers. FEBS Open Bio 2020, 10, 1587–1600. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Vantomme, L.; Jossart, Q.; Gérard, K.; Danis, B.; Moreau, C. Preliminary Assessment of Sea Star (Echinodermata, Asteroidea) Diversity in the Coastal Magellanic Region (South Chile) and Their Geographical Distribution. Diversity 2023, 15, 1129. [Google Scholar] [CrossRef]
- Balakirev, E.S. Trans-Species Polymorphism in Mitochondrial Genome of Camarodont Sea Urchins. Genes 2019, 10, 592. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.V.; Minale, L.; Riccio, R.; Zollo, F. Steroidal oligoglycosides and polyhydroxysteroids from Echinoderms. Fortschr. Chem. Org. Naturst. 1993, 62, 75–308. [Google Scholar] [CrossRef] [PubMed]
- Ivanchina, N.V.; Kicha, A.A.; Stonik, V.A. Steroid glycosides from marine organisms. Steroids 2011, 76, 425–454. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.M.; Downey, M.E. Starfishes of the Atlantic. 1992. Available online: https://api.semanticscholar.org/CorpusID:82206354 (accessed on 1 February 2024).
- Fontanella, F.; Hopkins, T.; Féral, J.; David, B. Preliminary phylogeny of Echinaster (Othilia) from the Gulf of Mexico based on morphological characters (Echinodermata: Asteroidea). Echinoderm Res. 2001, 337, 91–95. [Google Scholar]
- Knott, J.R.; Phillips, F.; Reheis, M.C.; Sada, D.; Jayko, A.; Axen, G. Geologic and hydrologic concerns about pupfish divergence during the last glacial maximum. Proc. R. Soc. B 2018, 285, 20171648. [Google Scholar] [CrossRef] [PubMed Central]
- Mah, C.L.; Blake, D.B. Global diversity and phylogeny of the Asteroidea (Echinodermata). PLoS ONE 2012, 7, e35644. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mah, C.L. New Genera, Species, and observations on the biology of Antarctic Valvatida (Asteroidea). Zootaxa 2023, 5310, 1–88. [Google Scholar] [CrossRef] [PubMed]
- Ubagan, M.D.; Lee, J.; Shin, S.; Lee, T. Two newly recorded echinoderms from the mesophotic zone in Korea. J. Species Res. 2023, 12, 180–188. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ward, R.D.; Holmes, B.H.; O’Hara, T.D. DNA barcoding discriminates echinoderm species. Mol. Ecol. Resour. 2008, 8, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Knott, K.E.; Ringvold, H.; Blicher, M.E. Morphological and molecular analysis of Henricia Gray, 1840 (Asteroidea: Echinodermata) from the Northern Atlantic Ocean. Zool. J. Linn. Soc. 2018, 182, 791–807. [Google Scholar] [CrossRef]
- Wakita, D.; Fujita, T.; Kajihara, H. Molecular systematics and morphological analyses of the subgenus Setihenricia (Echinodermata: Asteroidea: Henricia) from Japan. Species Divers. 2019, 24, 119–135. [Google Scholar] [CrossRef][Green Version]
- Gun, L.; Yumiao, R.; Haixian, P.; Liang, Z. Comprehensive analysis and comparison on the codon usage pattern of whole Mycobacterium tuberculosis coding genome from different area. BioMed Res. Int. 2018, 2018, 3574976. [Google Scholar] [CrossRef] [PubMed]
- Ubagan, M.D.; Shin, S. Two newly recorded sea stars of genus Henricia (Asteroidea: Spinulosida: Echinasteridae) from the East Sea, Korea. Korean J. Environ. Biol. 2023, 41, 364–369. [Google Scholar] [CrossRef]
- Ubagan, M.D.; Shin, S. A newly recorded sea star of genus Henricia (Asteroidea: Spinulosida: Echinasteridae) from the East Sea, Korea. J. Species Res. 2019, 8, 109–112. [Google Scholar] [CrossRef]
- Sun, S.; Xiao, N.; Sha, Z. Mitogenomics provides new insights into the phylogenetic relationships and evolutionary history of deep-sea sea stars (Asteroidea). Sci. Rep. 2022, 12, 4656. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Liu, J.; Zhang, H. The first complete mitochondrial genome of the Mariana Trench Freyastera benthophila (Asteroidea: Brisingida: Brisingidae) allows insights into the deep-sea adaptive evolution of Brisingida. Ecol. Evol. 2018, 8, 10673–10686. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, M.; Komatsu, M.; Araki, T.; Asakawa, S.; Yokobori, S.-I.; Watanabe, K.; Wada, H. The phylogenetic status of Paxillosida (Asteroidea) based on complete mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2005, 36, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Tan, M.; Meng, G.; Yang, S.; Su, X.; Liu, S.; Song, W.; Li, Y.; Wu, Q.; Zhang, A. Multiplex sequencing of pooled mitochondrial genomes—A crucial step toward biodiversity analysis using mito-metagenomics. Nucleic Acids Res. 2014, 42, e166. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Zeng, X.; Ni, G. The complete mitochondrial genome of the starfish Coscinasterias acutispina Stimpson, 1862 (Echinodermata, Forcipulatida) from the East China Sea. Mitochondrial DNA Part B 2022, 7, 1843–1844. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Shin, S. Complete mitochondrial genome analysis of Distolasterias nipon (Echinodermata, Asteroidea, Forcipulatida). Mitochondrial DNA Part B 2018, 3, 1290–1291. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Liu, J.; Zhang, H. The complete mitochondrial genome of Styracaster yapensis (Paxillosida: Porcellanasteridae): Characterization and phylogenetic position. Mitochondrial DNA Part B 2019, 4, 81–82. [Google Scholar] [CrossRef]
- Payne, C.Y.; Tilic, E.; Boschen-Rose, R.E.; Gannon, A.; Stiller, J.; Hiley, A.S.; Grupe, B.M.; Mah, C.L.; Rouse, G.W. Xyloplax princealberti (Asteroidea, Echinodermata): A New Species That Is Not Always Associated with Wood Falls. Diversity 2023, 15, 1212. [Google Scholar] [CrossRef]
- Seixas, V.C.; Ventura, C.R.R.; Paiva, P.C. The complete mitochondrial genome of the sea star Echinaster (Othilia) brasiliensis (Asteroidea: Echinasteridae). Conserv. Genet. Resour. 2019, 11, 151–155. [Google Scholar] [CrossRef]
- Lee, T.; Shin, S. Complete mitochondrial genome of the taxonomically notorious sea star, Henricia leviuscula (Asteroidea, Spinulosida, Echinasteridae), from South Korea. Mitochondrial DNA Part B 2019, 4, 2656–2657. [Google Scholar] [CrossRef]
- Lee, T.; Shin, S. Complete mitochondrial genome of Henricia pachyderma (Asteroidea, Spinulosida, Echinasteridae) and phylogenetic analysis. Mitochondrial DNA Part B 2020, 5, 1483–1484. [Google Scholar] [CrossRef]
- Yasuda, N.; Hamaguchi, M.; Sasaki, M.; Nagai, S.; Saba, M.; Nadaoka, K. Complete mitochondrial genome sequences for Crown-of-thorns starfish Acanthaster planci and Acanthaster brevispinus. BMC Genom. 2006, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, H.; Kajitani, R.; Nakamura, Y.; Takahashi, K.; Okuno, M.; Kobayashi, F.; Shinoda, T.; Toyoda, A.; Suzuki, Y.; Thongtham, N. Elucidation of the speciation history of three sister species of crown-of-thorns starfish (Acanthaster spp.) based on genomic analysis. DNA Res. 2021, 28, dsab012. [Google Scholar] [CrossRef] [PubMed]
- Quek, Z.B.R.; Chang, J.J.M.; Ip, Y.C.A.; Huang, D. Complete mitochondrial genome of the sea star Archaster typicus (Asteroidea: Archasteridae). Mitochondrial DNA Part B 2019, 4, 3130–3132. [Google Scholar] [CrossRef] [PubMed]
- Quek, Z.B.R.; Chang, J.J.M.; Ip, Y.C.A.; Chan, Y.K.S.; Huang, D. Mitogenomes reveal alternative initiation codons and lineage-specific gene order conservation in echinoderms. Mol. Biol. Evol. 2021, 38, 981–985. [Google Scholar] [CrossRef]
- Asakawa, S.; Himeno, H.; Miura, K.-I.; Watanabe, K. Nucleotide sequence and gene organization of the starfish Asterina pectinifera mitochondrial genome. Genetics 1995, 140, 1047–1060. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Hiruta, S.F.; Arai, M.; Shimizu, M.; Mah, C.L.; Fujita, T.; Setiamarga, D.H. The first complete mitochondrial genome of the Northern Pacific deep-sea goniasterid sea star Ceramaster japonicus (Sladen, 1889) determined using NGS-based shotgun sequencing. Mitochondrial DNA Part B 2021, 6, 1406–1408. [Google Scholar] [CrossRef] [PubMed]
- Hiruta, S.F.; Arai, M.; Chavanich, S.; Viyakarn, V.; Fujita, T. Complete mitochondrial genome of a sea star, Linckia laevigata (Echinodermata, Asteroidea, Valvatida, Ophidiasteridae). Mitochondrial DNA Part B 2020, 5, 1342–1343. [Google Scholar] [CrossRef]
- Alboasud, M.; Jeong, H.; Lee, T. The complete mitochondrial genome of Poraniopsis inflata (Asteroidea: Valvatida: Poraniidae) from Dokdo Island, Korea. Mitochondrial DNA Part B 2024, 9, 290–294. [Google Scholar] [CrossRef]
- Guo, J.; Fan, S.; Xu, Q. Study on genetic diversity of Ophiura sarsii population in the Arctic. Prog. Mar. Sci. 2021, 39. [Google Scholar] [CrossRef]
- Li, Y.-X.; Wang, X.-T.; Dong, Y.; Fan, S.-L.; Xu, Q.-Z. The complete mitochondrial genome of Ophiura kinbergi (Ophiuroidea, Ophiurina): Genome structure and phylogenetics. Mitochondrial DNA Part B 2020, 5, 1309–1310. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and high-performance computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Rubinoff, D.; Holland, B.S. Between two extremes: Mitochondrial DNA is neither the panacea nor the nemesis of phylogenetic and taxonomic inference. Syst. Biol. 2005, 54, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Waikagul, J.; Thaekham, U. Approaches to Research on the Systematics of Fish-Borne Trematodes; Academic Press: Waltham, MA, USA, 2014; Available online: https://books.google.co.kr/books?hl=en&lr=&id=Nsu7AgAAQBAJ&oi=fnd&pg=PP1&dq=28.%09Waikagul,+J.%3B+Thaekham,+U.+Approaches+to+Research+on+the+Systematics+of+Fish-Borne+Trematodes%3B+Academic+Press:+Waltham+Cam-bridge,+MA,+USA++,+2014.&ots=M9pQKTwzm6&sig=HswsYJ_TFPysOjH_z-s1YBYH_1U&redir_esc=y#v=onepage&q&f=false (accessed on 1 February 2014).
- Harrison, R.G. Animal mitochondrial DNA as a genetic marker in population and evolutionary biology. Trends Ecol. Evol. 1989, 4, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, S.; Komuro, T.; Izawa, H.; Tsutsumi, H. Analysis of human mitochondrial DNA polymorphisms in the Japanese population. Biochem. Genet. 2013, 51, 33–70. [Google Scholar] [CrossRef] [PubMed]
- Chichvarkhin, A.; Chichvarkhina, O.; Wakita, D. Sea stars of the genus Henricia Gray, 1840 (Echinodermata, Asteroidea) from Vostok Bay, Sea of Japan. PeerJ 2019, 7, e6585. [Google Scholar] [CrossRef] [PubMed Central]
- Ubagan, M.D.; Alboasud, M.S.A.; Lee, T. Morphological Description and Molecular Analysis of Newly Recorded Asteroid, Henricia djakonovi Chichvarkhin, 2017 (Asteroidea: Spinulosida: Echinasteridae), from Dokdo Island, Korea. Taxonomy 2023, 3, 46–54. [Google Scholar] [CrossRef]
- Chichvarkhin, A. Henricia djakonovi sp. nov.(Echinodermata, Echinasteridae): A new sea star species from the Sea of Japan. PeerJ 2017, 5, e2863. [Google Scholar] [CrossRef] [PubMed] [PubMed Central][Green Version]
- Madsen, F.J. The Henricia Sanguinolenta Complex (Echinodermata, Asteroidea) of the Norwegian Sea and Adjacent Waters: A Re-Evaluation, with Notes on Related Species; Zoological Museum, University of Copenhagen: Copenhagen, Denmark, 1987. [Google Scholar]
- Ubagan, M.D.; Lee, T.; Kim, P.; Shin, S. A new species of the genus Henricia (Asteroidea, Spinulosida, Echinasteridae) from South Korea. ZooKeys 2020, 997, 1. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).