
The Protective Role of KANK1 in Podocyte Injury

 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
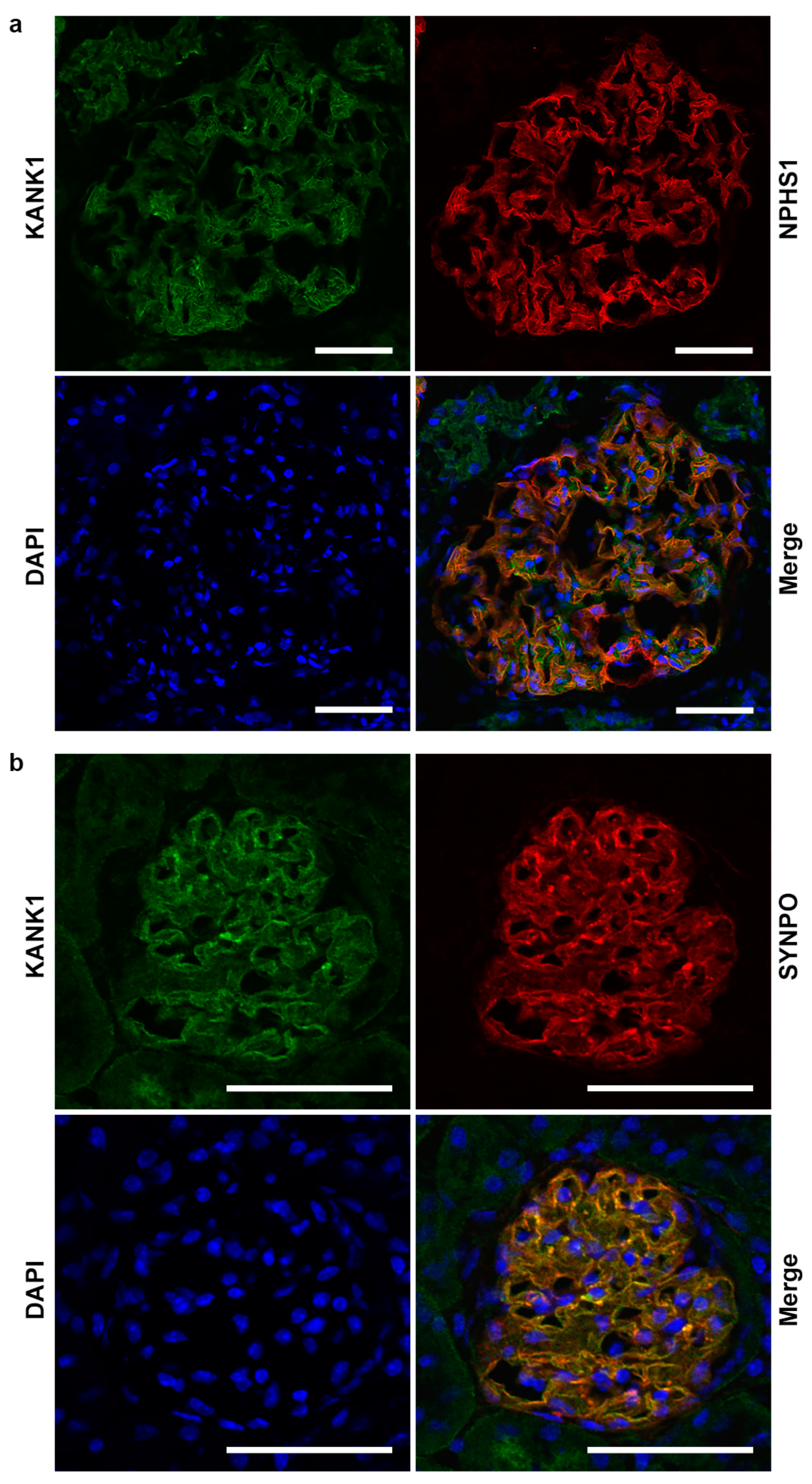
2.1. KANK1 Protein Is Expressed in Both Human and Mouse Podocytes
2.2. Kank1 Was Successfully Knocked Out in Mouse Podocytes
2.3. The Inactivation of Kank1 in Podocytes at Two or Six Months Did Not Result in Albuminuria
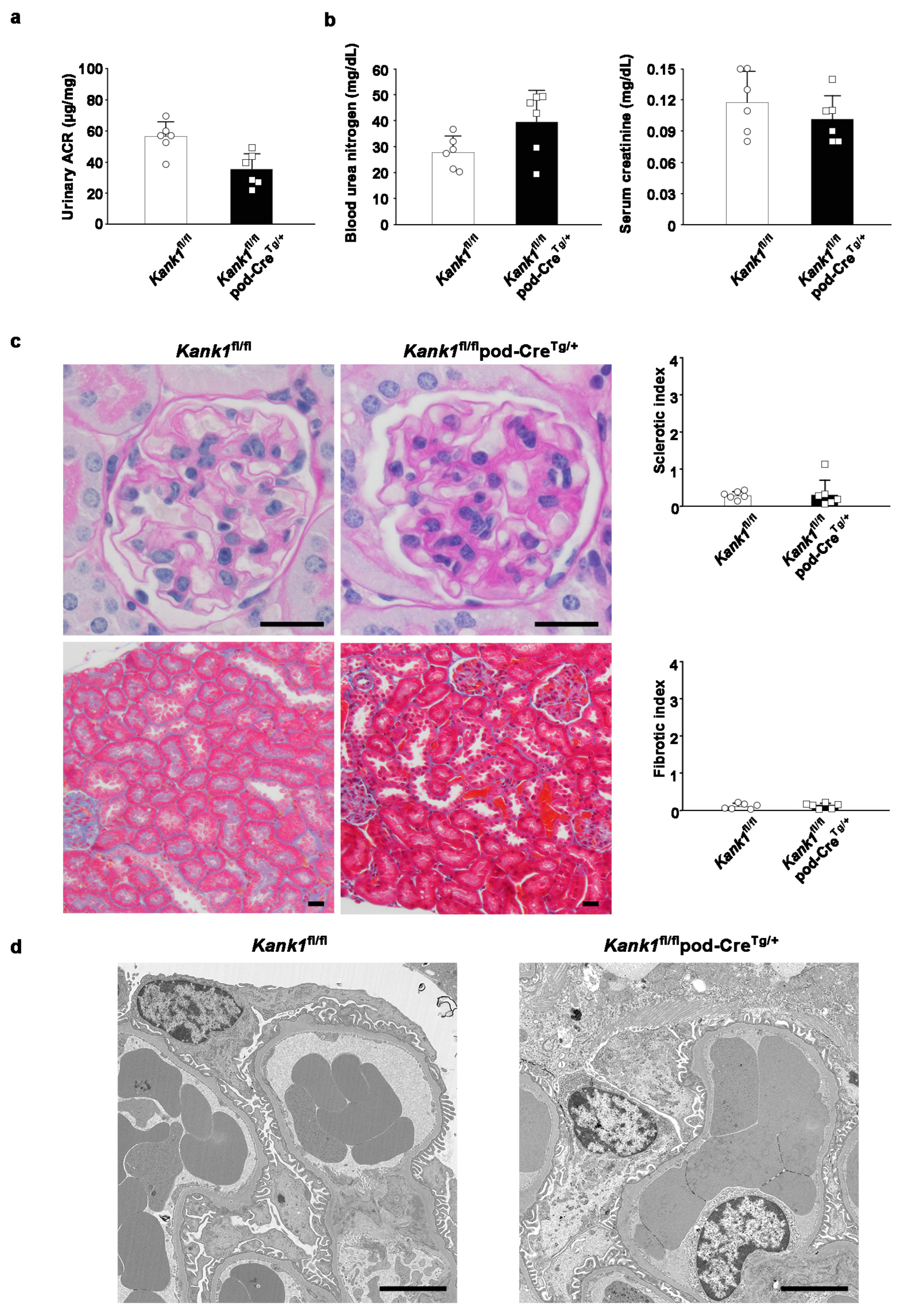
2.4. The Inactivation of Kank1 in Podocytes at One Year Did Not Result in Abnormalities in Urine, Blood, or Histological Tests
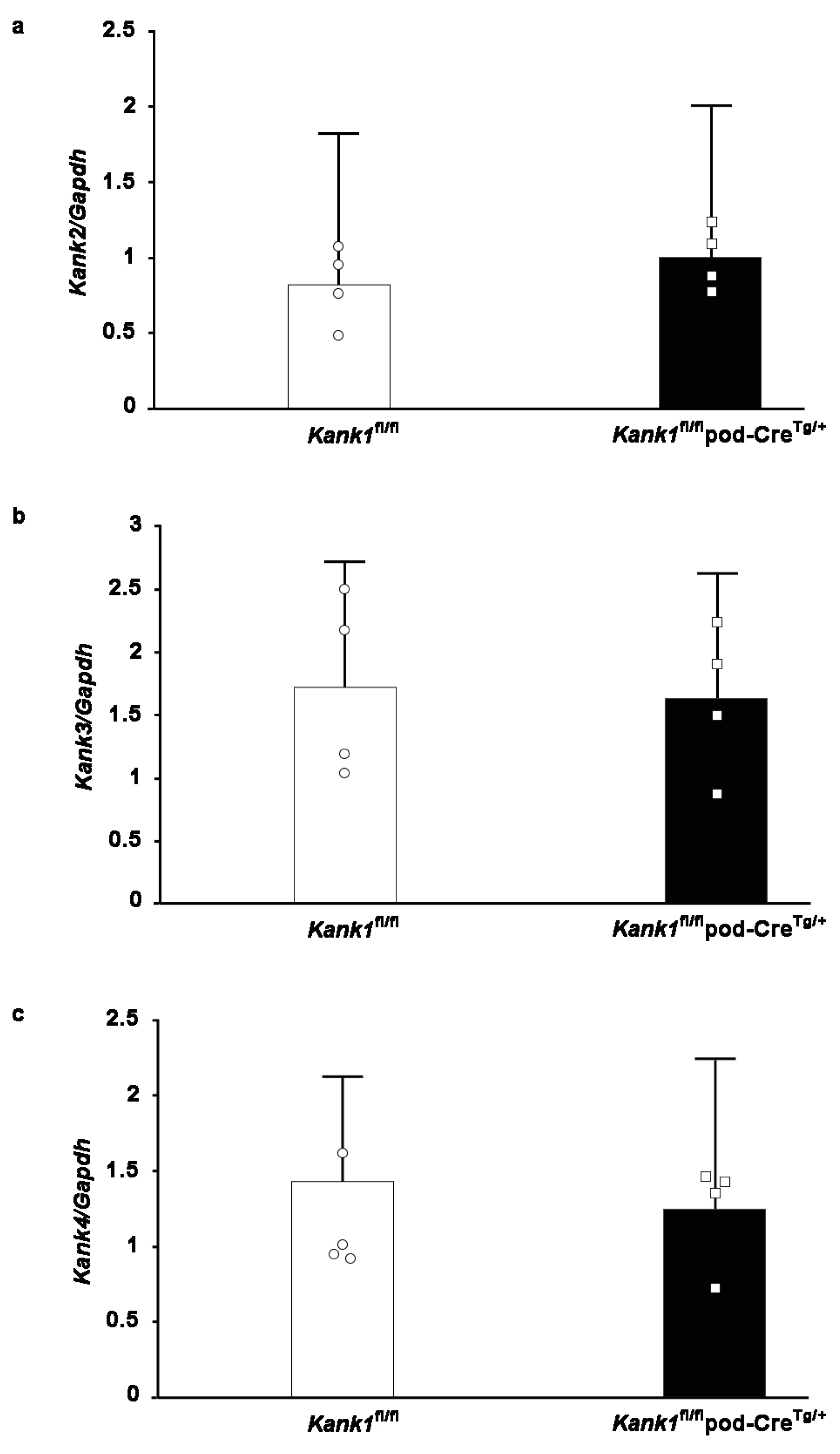
2.5. Compensation by other Members of the Kank Family Was Not Evident in Podocyte-Specific Kank1 Knockout Mice
2.6. Albuminuria and Glomerular Sclerosis Evident in Podocyte-Specific Kank1 Knockout Mice under Pathological Conditions
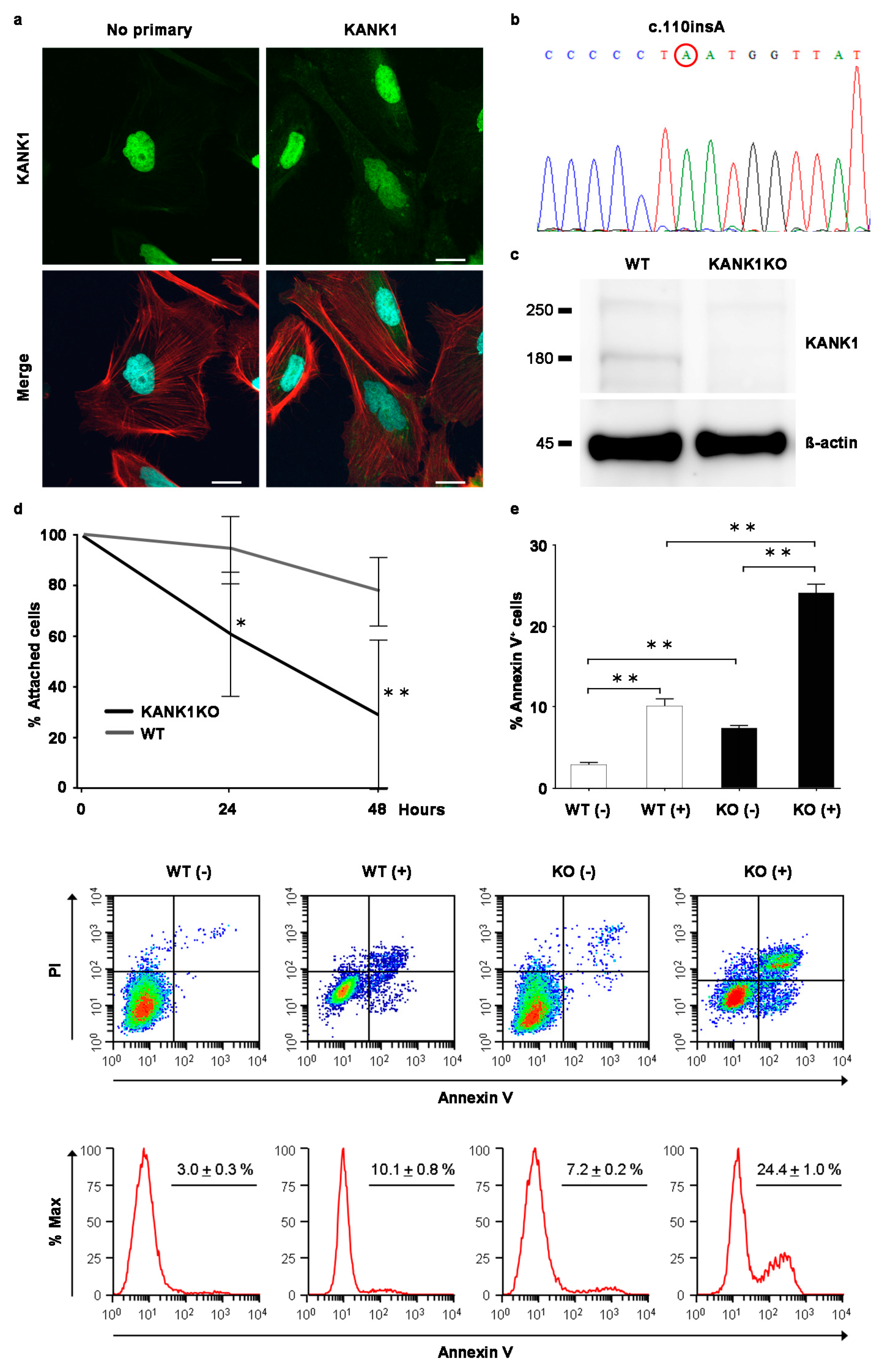
2.7. Generation of the Human Immortalized KANK1 Knockout (KANK1KO) Podocyte Cell Line
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Isolation of Mouse Glomeruli
4.3. RNA Extraction and Reverse Transcriptase (RT)-PCR
4.4. Immunofluorescence for KANK1 in Humans or WT Mice
4.5. Western Blotting
4.6. Immunofluorescence Studies in Mice
4.7. Urine and Blood Analyses
4.8. Light Microscopic Analyses
4.9. Transmission Electron Microscopic Analyses
4.10. Immunofluorescence in Human Podocytes
4.11. Generation of a KANK1KO Human Immortalized Podocyte Cell Line
4.12. Detachment Assays
4.13. Apoptosis Assays
4.14. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chesney, R.W. The idiopathic nephrotic syndrome. Curr. Opin. Pediatr. 1999, 11, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Mekahli, D.; Liutkus, A.; Ranchin, B.; Yu, A.; Bessenay, L.; Girardin, E.; Van Damme-Lombaerts, R.; Palcoux, J.B.; Cachat, F.; Lavocat, M.P.; et al. Long-term outcome of idiopathic steroid-resistant nephrotic syndrome: A multicenter study. Pediatr. Nephrol. 2009, 24, 1525–1532. [Google Scholar] [CrossRef]
- Rosenberg, A.Z.; Kopp, J.B. Focal Segmental Glomerulosclerosis. Clin. J. Am. Soc. Nephrol. 2017, 12, 502–517. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, C.E.; Lovric, S.; Ashraf, S.; Pabst, W.L.; Gee, H.Y.; Kohl, S.; Engelmann, S.; Vega-Warner, V.; Fang, H.; Halbritter, J.; et al. A single-gene cause in 29.5% of cases of steroid-resistant nephrotic syndrome. J. Am. Soc. Nephrol. 2015, 26, 1279–1289. [Google Scholar] [CrossRef] [PubMed]
- Lovric, S.; Ashraf, S.; Tan, W.; Hildebrandt, F. Genetic testing in steroid-resistant nephrotic syndrome: When and how? Nephrol. Dial. Transplant. 2016, 31, 1802–1813. [Google Scholar] [CrossRef]
- Gee, H.Y.; Zhang, F.; Ashraf, S.; Kohl, S.; Sadowski, C.E.; Vega-Warner, V.; Zhou, W.; Lovric, S.; Fang, H.; Nettleton, M.; et al. KANK deficiency leads to podocyte dysfunction and nephrotic syndrome. J. Clin. Investig. 2015, 125, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Roy, B.C.; Hatano, N.; Aoyagi, T.; Gohji, K.; Kiyama, R. A novel ankyrin repeat-containing gene (Kank) located at 9p24 is a growth suppressor of renal cell carcinoma. J. Biol. Chem. 2002, 277, 36585–36591. [Google Scholar] [CrossRef] [PubMed]
- Kakinuma, N.; Zhu, Y.; Wang, Y.; Roy, B.C.; Kiyama, R. Kank proteins: Structure, functions and diseases. Cell. Mol. Life Sci. 2009, 66, 2651–2659. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Kakinuma, N.; Wang, Y.; Kiyama, R. Kank proteins: A new family of ankyrin-repeat domain-containing proteins. Biochim. Biophys. Acta 2008, 1780, 128–133. [Google Scholar] [CrossRef]
- Roy, B.C.; Kakinuma, N.; Kiyama, R. Kank attenuates actin remodeling by preventing interaction between IRSp53 and Rac1. J. Cell Biol. 2009, 184, 253–267. [Google Scholar]
- Li, C.C.; Kuo, J.C.; Waterman, C.M.; Kiyama, R.; Moss, J.; Vaughan, M. Effects of brefeldin A-inhibited guanine nucleotide-exchange (BIG) 1 and KANK1 proteins on cell polarity and directed migration during wound healing. Proc. Natl. Acad. Sci. USA 2011, 108, 19228–19233. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, B.P.; Gough, R.E.; Ammon, Y.C.; van de Willige, D.; Post, H.; Jacquemet, G.; Altelaar, A.M.; Heck, A.J.; Goult, B.T.; Akhmanova, A. Talin-KANK1 interaction controls the recruitment of cortical microtubule stabilizing complexes to focal adhesions. elife 2016, 5, e18124. [Google Scholar] [CrossRef]
- Chen, N.P.; Sun, Z.; Fässler, R. The Kank family proteins in adhesion dynamics. Curr. Opin. Cell Biol. 2018, 54, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Goult, B.T.; Ballestrem, C.; Zacharchenko, T. The structural basis of the talin-KANK1 interaction that coordinates the actin and microtubule cytoskeletons at focal adhesions. Open Biol. 2023, 13, 230058. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Onishi, Y.; Kakinuma, N.; Roy, B.C.; Aoyagi, T.; Kiyama, R. Alternative splicing of the human Kank gene produces two types of Kank protein. Biochem. Biophys. Res. Commun. 2005, 330, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Lindenmeyer, M.T.; Eichinger, F.; Sen, K.; Anders, H.J.; Edenhofer, I.; Mattinzoli, D.; Kretzler, M.; Rastaldi, M.P.; Cohen, C.D. Systematic analysis of a novel human renal glomerulus-enriched gene expression dataset. PLoS ONE 2010, 5, 11545. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Artomov, M.; Brähler, S.; Stander, M.C.; Shamsan, G.; Sampson, M.G.; White, J.M.; Kretzler, M.; Miner, J.H.; Jain, S.; et al. A role for genetic susceptibility in sporadic focal segmental glomerulosclerosis. J. Clin. Investig. 2016, 126, 1067–1078. [Google Scholar] [CrossRef]
- Kramer-Zucker, A.G.; Wiessner, S.; Jensen, A.M.; Drummond, I.A. Organization of the pronephric filtration apparatus in zebrafish requires Nephrin, Podocin and the FERM domain protein Mosaic eyes. Dev. Biol. 2005, 285, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Ebarasi, L.; He, L.; Hultenby, K.; Takemoto, M.; Betsholtz, C.; Tryggvason, K.; Majumdar, A. A reverse genetic screen in the zebrafish identifies crb2b as a regulator of the glomerular filtration barrier. Dev. Biol. 2009, 334, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Almosailleakh, M.; Bentivegna, S.; Narcisi, S.; Benquet, S.J.; Gillberg, L.; Montaño-Almendras, C.P.; Savickas, S.; Schoof, E.M.; Wegener, A.; Luche, H.; et al. Loss of the KN Motif and AnKyrin Repeat Domain 1 (KANK1) Leads to Lymphoid Compartment Dysregulation in Murine Model. Genes 2023, 14, 1947. [Google Scholar] [CrossRef]
- Pan, W.; Sun, K.; Tang, K.; Xiao, Q.; Ma, C.; Yu, C.; Wei, Z. Structural insights into ankyrin repeat-mediated recognition of the kinesin motor protein KIF21A by KANK1, a scaffold protein in focal adhesion. J. Biol. Chem. 2018, 293, 1944–1956. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Sasamura, H.; Nakamura, M.; Azegami, T.; Oguchi, H.; Sakamaki, Y.; Itoh, H. KLF4-dependent epigenetic remodeling modulates podocyte phenotypes and attenuates proteinuria. J. Clin. Investig. 2014, 124, 2523–2537. [Google Scholar] [CrossRef]
- Bakker, W.W.; van Dael, C.M.; Pierik, L.J.; van Wijk, J.A.; Nauta, J.; Borghuis, T.; Kapojos, J.J. Altered activity of plasma hemopexin in patients with minimal change disease in relapse. Pediatr. Nephrol. 2005, 20, 1410–1415. [Google Scholar] [CrossRef] [PubMed]
- Bakker, W.W.; Borghuis, T.; Harmsen, M.C.; van den Berg, A.; Kema, I.P.; Niezen, K.E.; Kapojos, J.J. Protease activity of plasma hemopexin. Kidney Int. 2005, 68, 603–610. [Google Scholar] [CrossRef]
- Candelier, J.J.; Lorenzo, H.K. Idiopathic nephrotic syndrome and serum permeability factors: A molecular jigsaw puzzle. Cell Tissue Res. 2020, 379, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.W.; Harris, D.C. Adriamycin nephropathy: A model of focal segmental glomerulosclerosis. Nephrology 2011, 16, 30–38. [Google Scholar] [CrossRef]
- Takemoto, M.; Asker, N.; Gerhardt, H.; Lundkvist, A.; Johansson, B.R.; Saito, Y.; Betsholtz, C. A new method for large scale isolation of kidney glomeruli from mice. Am. J. Pathol. 2002, 161, 799–805. [Google Scholar] [CrossRef]
- Lahdenperä, J.; Kilpeläinen, P.; Liu, X.L.; Pikkarainen, T.; Reponen, P.; Ruotsalainen, V.; Tryggvason, K. Clustering-induced tyrosine phosphorylation of nephrin by Src family kinases. Kidney Int. 2003, 64, 404–413. [Google Scholar] [CrossRef]
- Murata, T.; Katayama, K.; Oohashi, T.; Jahnukainen, T.; Yonezawa, T.; Sado, Y.; Ishikawa, E.; Nomura, S.; Tryggvason, K.; Ito, M. COL4A6 is dispensable for autosomal recessive Alport syndrome. Sci. Rep. 2016, 6, 29450. [Google Scholar] [CrossRef]
- Katayama, K.; Kawano, M.; Naito, I.; Ishikawa, H.; Sado, Y.; Asakawa, N.; Murata, T.; Oosugi, K.; Kiyohara, M.; Ishikawa, E.; et al. Irradiation prolongs survival of Alport mice. J. Am. Soc. Nephrol. 2008, 19, 1692–1700. [Google Scholar] [CrossRef]
- Lin, M.H.; Miller, J.B.; Kikkawa, Y.; Suleiman, H.Y.; Tryggvason, K.; Hodges, B.L.; Miner, J.H. Laminin-521 Protein Therapy for Glomerular Basement Membrane and Podocyte Abnormalities in a Model of Pierson Syndrome. J. Am. Soc. Nephrol. 2018, 29, 1426–1436. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, A.; Katayama, K.; Ito, Y.; Joh, K.; Toda, M.; Yasuma, T.; D’Alessandro-Gabazza, C.N.; Kawachi, H.; Yan, K.; Ito, M.; et al. Podocyte-specific Crb2 knockout mice develop focal segmental glomerulosclerosis. Sci. Rep. 2021, 11, 20556. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J.; Coward, R.J.; Inward, C.D.; Farren, T.; Xing, C.Y.; Ni, L.; Mathieson, P.W.; Mundel, P. A conditionally immortalized human podocyte cell line demonstrating nephrin and podocin expression. J. Am. Soc. Nephrol. 2002, 13, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Reiser, J.; Oh, J.; Shirato, I.; Asanuma, K.; Hug, A.; Mundel, T.M.; Honey, K.; Ishidoh, K.; Kominami, E.; Kreidberg, J.A.; et al. Podocyte migration during nephrotic syndrome requires a coordinated interplay between cathepsin L and alpha3 integrin. J. Biol. Chem. 2004, 279, 34827–34832. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oda, K.; Katayama, K.; Zang, L.; Toda, M.; Tanoue, A.; Saiki, R.; Yasuma, T.; D’Alessandro-Gabazza, C.N.; Shimada, Y.; Mori, M.; et al. The Protective Role of KANK1 in Podocyte Injury. Int. J. Mol. Sci. 2024, 25, 5808. https://doi.org/10.3390/ijms25115808
Oda K, Katayama K, Zang L, Toda M, Tanoue A, Saiki R, Yasuma T, D’Alessandro-Gabazza CN, Shimada Y, Mori M, et al. The Protective Role of KANK1 in Podocyte Injury. International Journal of Molecular Sciences. 2024; 25(11):5808. https://doi.org/10.3390/ijms25115808
Chicago/Turabian StyleOda, Keiko, Kan Katayama, Liqing Zang, Masaaki Toda, Akiko Tanoue, Ryosuke Saiki, Taro Yasuma, Corina N. D’Alessandro-Gabazza, Yasuhito Shimada, Mutsuki Mori, and et al. 2024. "The Protective Role of KANK1 in Podocyte Injury" International Journal of Molecular Sciences 25, no. 11: 5808. https://doi.org/10.3390/ijms25115808
APA StyleOda, K., Katayama, K., Zang, L., Toda, M., Tanoue, A., Saiki, R., Yasuma, T., D’Alessandro-Gabazza, C. N., Shimada, Y., Mori, M., Suzuki, Y., Murata, T., Hirai, T., Tryggvason, K., Gabazza, E. C., & Dohi, K. (2024). The Protective Role of KANK1 in Podocyte Injury. International Journal of Molecular Sciences, 25(11), 5808. https://doi.org/10.3390/ijms25115808