
miRNAs in Follicular and Oviductal Fluids Support Global DNA Demethylation in Early-Stage Embryos


Abstract
1. Introduction
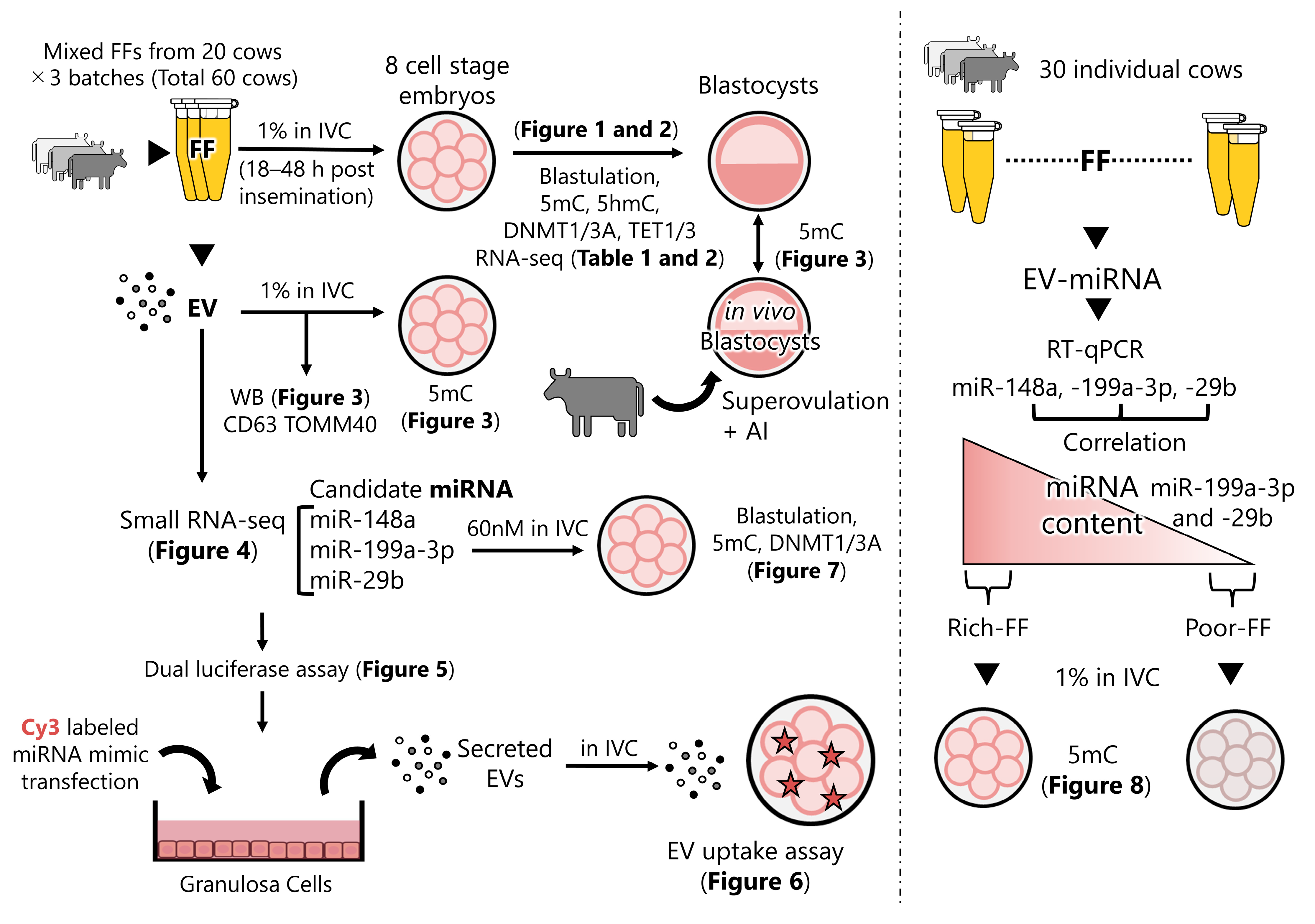
2. Results
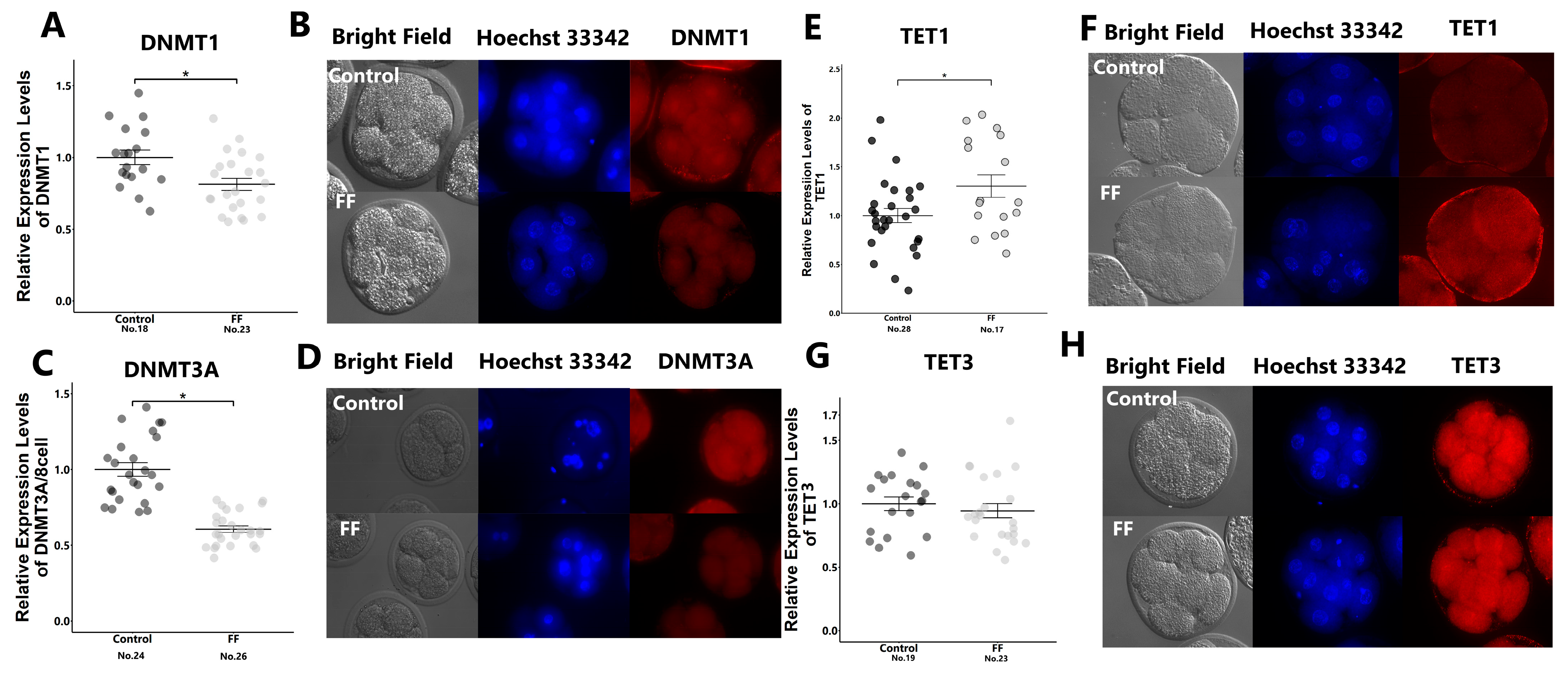
2.1. EVs in FF Induced the Demethylation of In Vitro-Produced Embryos
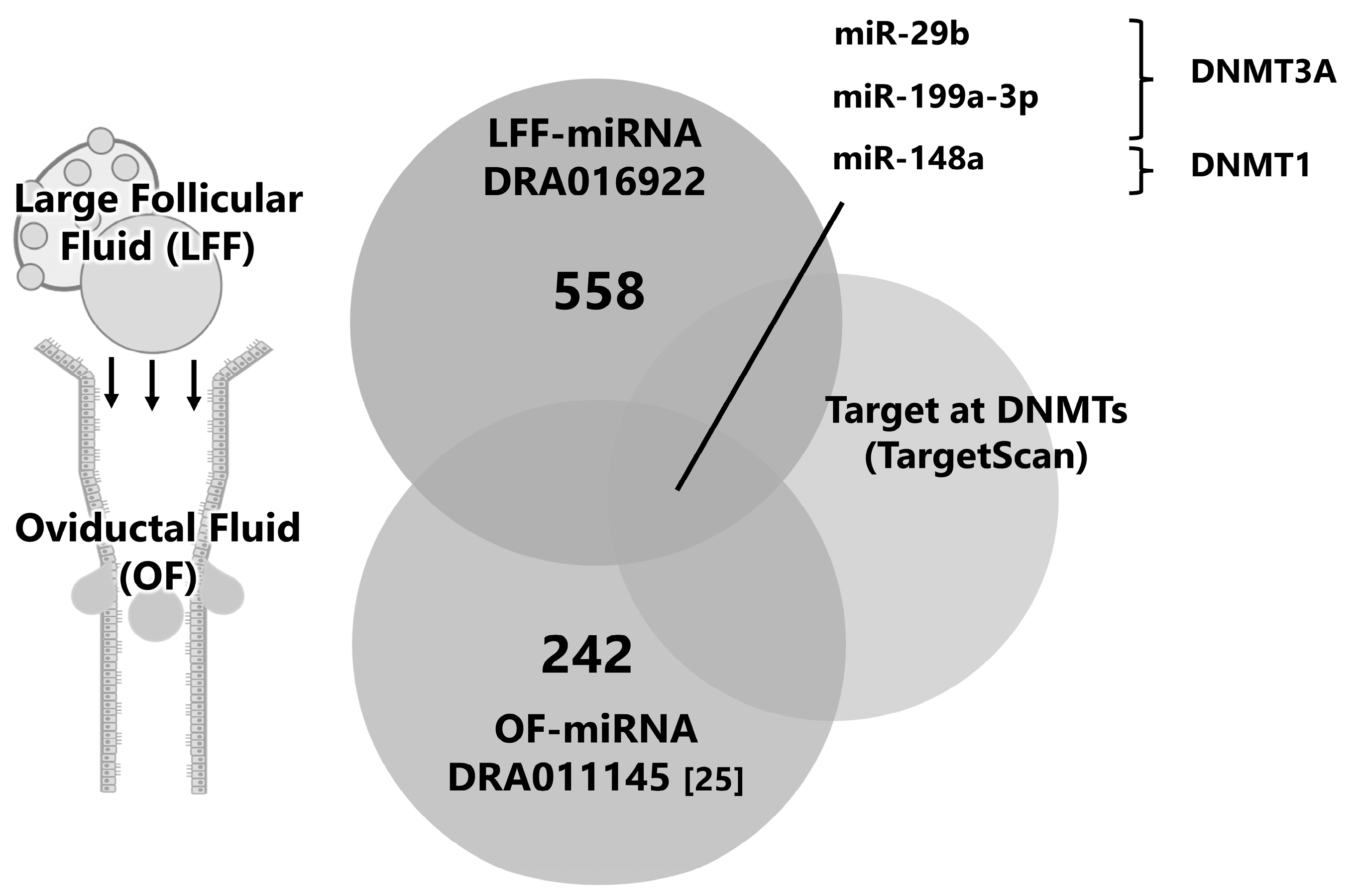
2.2. Identification of Molecules Affecting DNA Methylation
2.3. miRNA Mimic Transfection and Assay of EV Uptake by Embryos
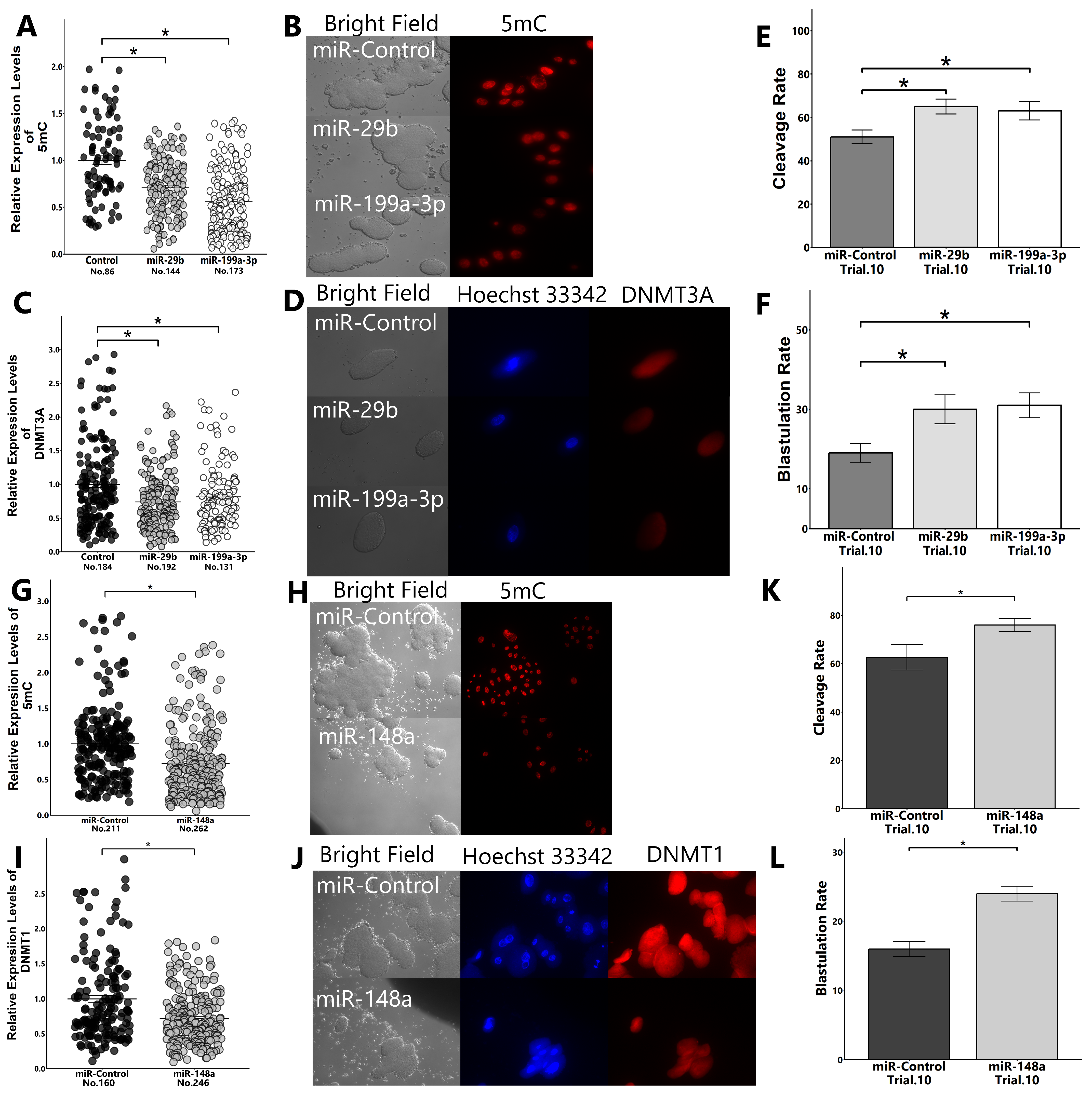
2.4. Transfection of Zygotes with miR-29b, miR-199a-3p, and miR-148a
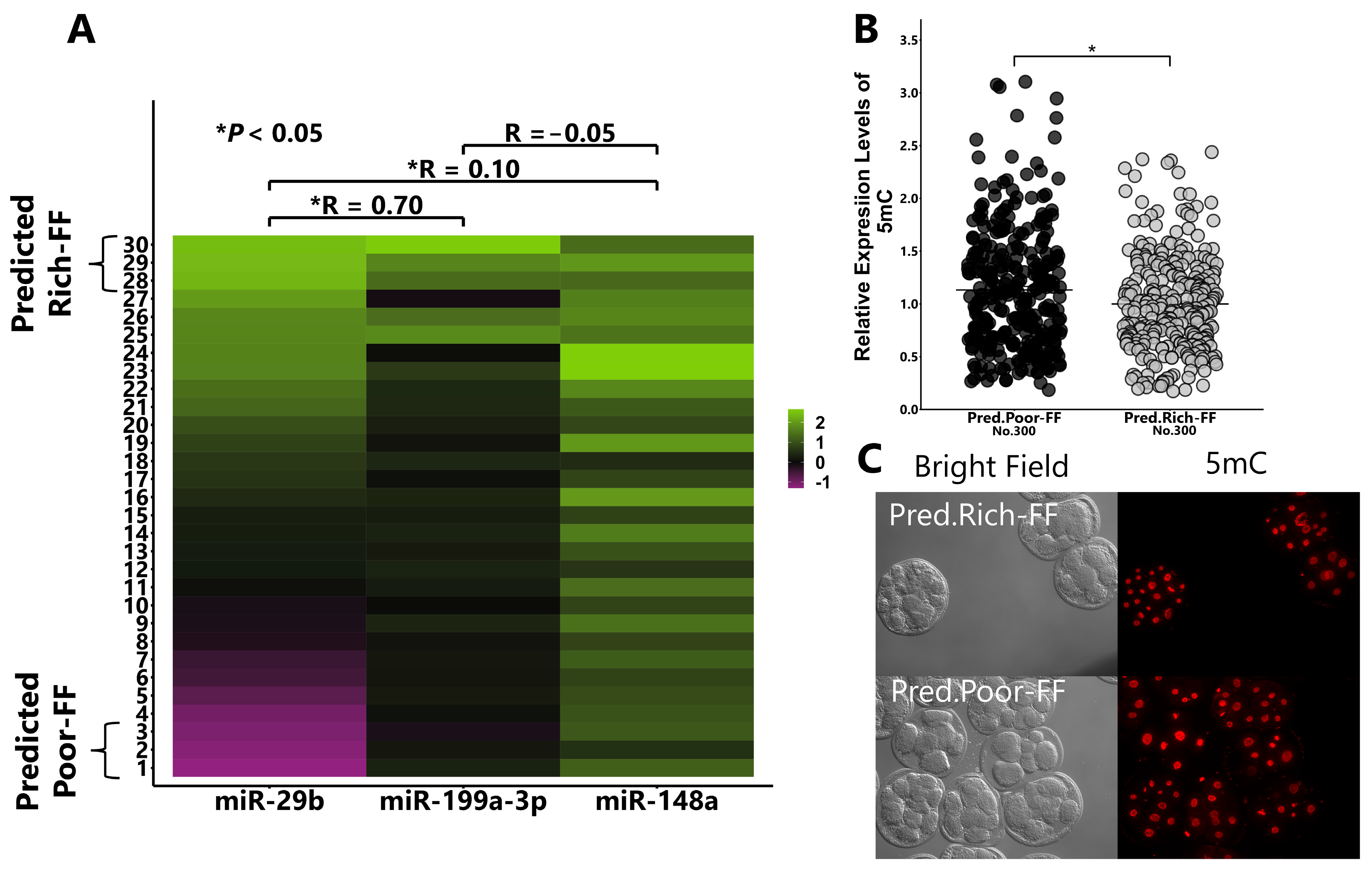
2.5. Validation of the Hypothesis That FF Containing High Levels of miR-29b, 199-3p, and 148a Reduce DNA Methylation Levels
2.6. RNA-seq of FF-Treated 8-Cell-Stage Embryos and Blastocysts
3. Discussion
4. Materials and Methods
4.1. Chemicals and Medium
4.2. Oocyte Collection, IVM, IVF, and IVC
4.3. Preparation of FF
4.4. Effect of Supplementation of IVC Medium with FF on Embryonic Development
4.5. Immunostaining
4.6. Isolation of EVs from FF and the Extraction of miRNAs from EVs
4.7. Collection and Preparation of Live Granulosa Cells
4.8. Western Blotting
4.9. Superovulation and Flushing of Embryos
4.10. Small RNA Sequences of miRNAs
4.11. Preparation of the pmirGLO Vector and miRNA Mimics
4.12. Dual Luciferase Reporter Assay
4.13. Preparation of Cy3-Labeled miRNA Mimic
4.14. EV-Uptake Assay
4.15. Effect of Supplementation of IVC Medium with miRNA Mimics on Embryonic Development
4.16. Selection of FF Based on the miRNA Content Using RT-qPCR
4.17. RNA-seq and Analysis of the Data
4.18. Publicly Available Database of miRNAs in Bovine FF and OF
4.19. Experimental Design
4.20. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnwell, C.V.; Farin, P.W.; Whisnant, C.S.; Alexander, J.E.; Farin, C.E. Maternal serum progesterone concentration and early conceptus development of bovine embryos produced in vivo or in vitro. Domest. Anim. Endocrinol. 2015, 52, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Schieve, L.A.; Meikle, S.F.; Ferre, C.; Peterson, H.B.; Jeng, G.; Wilcox, L.S. Low and very low birth weight in infants conceived with use of assisted reproductive technology. N. Engl. J. Med. 2002, 346, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Reefhuis, J.; Honein, M.A.; Schieve, L.A.; Correa, A.; Hobbs, C.A.; Rasmussen, S.A.; National Birth Defects Prevention Study. Assisted reproductive technology and major structural birth defects in the United States. Hum. Reprod. 2009, 24, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Urrego, R.; Rodriguez-Osorio, N.; Niemann, H. Epigenetic disorders and altered gene expression after use of Assisted Reproductive Technologies in domestic cattle. Epigenetics 2014, 9, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Uysal, F.; Ozturk, S.; Akkoyunlu, G. DNMT1, DNMT3A and DNMT3B proteins are differently expressed in mouse oocytes and early embryos. J. Mol. Histol. 2017, 48, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.S.; Østrup, O.; Østrup, E.; Vejlsted, M.; Niemann, H.; Lucas-Hahn, A.; Petersen, B.; Li, J.; Callesen, H.; Hyttel, P. DNA methylation in porcine preimplantation embryos developed in vivo and produced by in vitro fertilization, parthenogenetic activation and somatic cell nuclear transfer. Epigenetics 2011, 6, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Giritharan, G.; Delle Piane, L.; Donjacour, A.; Esteban, F.J.; Horcajadas, J.A.; Maltepe, E.; Rinaudo, P. In vitro culture of mouse embryos reduces differential gene expression between inner cell mass and trophectoderm. Reprod. Sci. 2012, 19, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Aizawa, T.; Munakata, Y.; Iwata, H. Comparison of gene expression and mitochondria number between bovine blastocysts obtained in vitro and in vivo. J. Reprod. Dev. 2020, 66, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Lyons, R.A.; Saridogan, E.; Djahanbakhch, O. The effect of ovarian follicular fluid and peritoneal fluid on Fallopian tube ciliary beat frequency. Hum. Reprod. 2006, 21, 52–56. [Google Scholar] [CrossRef]
- Hasan, M.M.; Viil, J.; Lättekivi, F.; Ord, J.; Reshi, Q.U.A.; Jääger, K.; Velthut-Meikas, A.; Andronowska, A.; Jaakma, Ü.; Salumets, A.; et al. Bovine Follicular Fluid and Extracellular Vesicles Derived from Follicular Fluid Alter the Bovine Oviductal Epithelial Cells Transcriptome. Int. J. Mol. Sci. 2020, 21, 5365. [Google Scholar] [CrossRef]
- Hamdi, M.; Lopera-Vasquez, R.; Maillo, V.; Sanchez-Calabuig, M.J.; Núnez, C.; Gutierrez-Adan, A.; Rizos, D. Bovine oviductal and uterine fluid support in vitro embryo development. Reprod. Fertil. Dev. 2018, 30, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Barrera, A.D.; García, E.V.; Hamdi, M.; Sánchez-Calabuig, M.J.; López-Cardona, Á.P.; Balvís, N.F.; Rizos, D.; Gutiérrez-Adán, A. Embryo culture in presence of oviductal fluid induces DNA methylation changes in bovine blastocysts. Reproduction 2017, 154, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Qu, P.; Luo, S.; Du, Y.; Zhang, Y.; Song, X.; Yuan, X.; Lin, Z.; Li, Y.; Liu, E. Extracellular vesicles and melatonin benefit embryonic develop by regulating reactive oxygen species and 5-methylcytosine. J. Pineal Res. 2020, 68, e12635. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.L.V.; Cañón-Beltrán, K.; Cajas, Y.N.; Hamdi, M.; Yaryes, A.; Millán de la Blanca, M.G.; Beltrán-Breña, P.; Mazzarella, R.; da Silveira, J.C.; Gutiérrez-Adán, A.; et al. Extracellular vesicles from oviductal and uterine fluids supplementation in sequential in vitro culture improves bovine embryo quality. J. Anim. Sci. Biotechnol. 2022, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Mobarak, H.; Heidarpour, M.; Lolicato, F.; Nouri, M.; Rahbarghazi, R.; Mahdipour, M. Physiological impact of extracellular vesicles on female reproductive system; highlights to possible restorative effects on female age-related fertility. BioFactors 2019, 45, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Inoue, Y.; Shinozawa, A.; Tanaka, K.; Shirasuna, K.; Iwata, H. miR-17-5p in bovine oviductal fluid affects embryo development. Mol. Cell. Endocrinol. 2022, 551, 111651. [Google Scholar] [CrossRef]
- Libersky, E.A.; Boatman, D.E. Progesterone concentrations in serum, follicular fluid, and oviductal fluid of the golden hamster during the periovulatory period. Biol. Reprod. 1995, 53, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Avilés, M.; Gutiérrez-Adán, A.; Coy, P. Oviductal secretions: Will they be key factors for the future ARTs? Mol. Hum. Reprod. 2010, 16, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.S.; Canha-Gouveia, A.; París-Oller, E.; Coy, P. Supplementation of bovine follicular fluid during in vitro maturation increases oocyte cumulus expansion, blastocyst developmental kinetics, and blastocyst cell number. Theriogenology 2019, 126, 222–229. [Google Scholar] [CrossRef]
- Azari-Dolatabad, N.; Raes, A.; Pavani, K.C.; Asaadi, A.; Angel-Velez, D.; Van Damme, P.; Leroy, J.L.M.R.; Van Soom, A.; Pascottini, O.B. Follicular fluid during individual oocyte maturation enhances cumulus expansion and improves embryo development and quality in a dose-specific manner. Theriogenology 2021, 166, 38–45. [Google Scholar] [CrossRef]
- Hung, W.T.; Hong, X.; Christenson, L.K.; McGinnis, L.K. Extracellular Vesicles from Bovine Follicular Fluid Support Cumulus Expansion. Biol. Reprod. 2015, 93, 117. [Google Scholar] [CrossRef] [PubMed]
- Hung, W.T.; Navakanitworakul, R.; Khan, T.; Zhang, P.; Davis, J.S.; McGinnis, L.K.; Christenson, L.K. Stage-specific follicular extracellular vesicle uptake and regulation of bovine granulosa cell proliferation. Biol. Reprod. 2017, 97, 644–655. [Google Scholar] [CrossRef] [PubMed]
- da Silveira, J.C.; Andrade, G.M.; Del Collado, M.; Sampaio, R.V.; Sangalli, J.R.; Silva, L.A.; Pinaffi, F.V.L.; Jardim, I.B.; Cesar, M.C.; Nogueira, M.F.G.; et al. Supplementation with small-extracellular vesicles from ovarian follicular fluid during in vitro production modulates bovine embryo development. PLoS ONE 2017, 12, e0179451. [Google Scholar] [CrossRef] [PubMed]
- Navakanitworakul, R.; Hung, W.T.; Gunewardena, S.; Davis, J.S.; Chotigeat, W.; Christenson, L.K. Characterization and Small RNA Content of Extracellular Vesicles in Follicular Fluid of Developing Bovine Antral Follicles. Sci. Rep. 2016, 6, 25486. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Inoue, Y.; Hara, S.; Itou, J.; Shirasuna, K.; Iwata, H. microRNAs associated with the quality of follicular fluids affect oocyte and early embryonic development. Reprod. Med. Biol. 2024, 23, e12559. [Google Scholar] [CrossRef] [PubMed]
- Asaadi, A.; Dolatabad, N.A.; Atashi, H.; Raes, A.; Van Damme, P.; Hoelker, M.; Hendrix, A.; Pascottini, O.B.; Van Soom, A.; Kafi, M.; et al. Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality. Int. J. Mol. Sci. 2021, 22, 578. [Google Scholar] [CrossRef] [PubMed]
- Arand, J.; Reijo Pera, R.A.; Wossidlo, M. Reprogramming of DNA methylation is linked to successful human preimplantation development. Histochem. Cell Biol. 2021, 156, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.T. DNA methylation remodeling in vitro and in vivo. Curr. Opin. Genet. Dev. 2015, 34, 82–87. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, J.; Zhang, S.; Zhai, Y.; Jiang, Y.; An, X.; Ma, X.; Zhang, X.; Li, Z.; Tang, B. Tet3 is required for normal in vitro fertilization preimplantation embryos development of bovine. Mol. Reprod. Dev. 2019, 86, 298–307. [Google Scholar] [CrossRef]
- Duan, J.; Zhu, L.; Dong, H.; Zheng, X.; Jiang, Z.; Chen, J.; Tian, X.C. Analysis of mRNA abundance for histone variants, histone- and DNA-modifiers in bovine in vivo and in vitro oocytes and embryos. Sci. Rep. 2019, 9, 1217. [Google Scholar] [CrossRef]
- Deng, M.; Zhang, G.; Cai, Y.; Liu, Z.; Zhang, Y.; Meng, F.; Wang, F.; Wan, Y. DNA methylation dynamics during zygotic genome activation in goat. Theriogenology 2020, 156, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Sattari, M.; Kohvakka, A.; Moradi, E.; Rauhala, H.; Urhonen, H.; Isaacs, W.B.; Nykter, M.; Murtola, T.J.; Tammela, T.L.J.; Latonen, L.; et al. Identification of long noncoding RNAs with aberrant expression in prostate cancer metastases. Endocr.-Relat. Cancer 2023, 30, e220247. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.J. Mitochondria in early development: Linking the microenvironment, metabolism and the epigenome. Reproduction 2019, 157, R159–R179. [Google Scholar] [CrossRef]
- Nagaraj, R.; Sharpley, M.S.; Chi, F.; Braas, D.; Zhou, Y.; Kim, R.; Clark, A.T.; Banerjee, U. Nuclear Localization of Mitochondrial TCA Cycle Enzymes as a Critical Step in Mammalian Zygotic Genome Activation. Cell 2017, 168, 210–223.e11. [Google Scholar] [CrossRef] [PubMed]
- Verruma, C.G.; Santos, R.S.; Marchesi, J.A.P.; Sales, S.L.A.; Vila, R.A.; Rios, Á.F.L.; Furtado, C.L.M.; Ramos, E.S. Dynamic methylation pattern of H19DMR and KvDMR1 in bovine oocytes and preimplantation embryos. J. Assist. Reprod. Genet. 2024, 41, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Wang, H.; Wu, Z.; Zhang, J.; Liu, Z.; He, H. The expression patterns of DNA methylation reprogramming related genes are associated with the developmental competence of cloned embryos after zygotic genome activation in pigs. Gene Expr. Patterns GEP 2015, 18, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Cao, Y.; Zhai, Y.; Ma, X.; An, X.; Zhang, S.; Li, Z. MicroRNA-29b regulates DNA methylation by targeting Dnmt3a/3b and Tet1/2/3 in porcine early embryo development. Dev. Growth Differ. 2018, 60, 197–204. [Google Scholar] [CrossRef]
- Wang, S.; Chen, L.; Zhu, Y.; Jiang, W. Characterization of microRNAs in spent culture medium associated with human embryo quality and development. Ann. Transl. Med. 2021, 9, 1648. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Jiang, R.; Jiang, Y.; Su, Y.; Wang, S. A validated model for individualized prediction of pregnancy outcome in woman after fresh cycle of Day 5 single blastocyst transfer. Sci. Rep. 2023, 13, 10016. [Google Scholar] [CrossRef]
- Tan, K.; Wang, X.; Zhang, Z.; Miao, K.; Yu, Y.; An, L.; Tian, J. Downregulation of miR-199a-5p Disrupts the Developmental Potential of In Vitro-Fertilized Mouse Blastocysts. Biol. Reprod. 2016, 95, 54. [Google Scholar] [CrossRef]
- Liu, B.; Xu, Q.; Wang, Q.; Feng, S.; Lai, F.; Wang, P.; Zheng, F.; Xiang, Y.; Wu, J.; Nie, J.; et al. The landscape of RNA Pol II binding reveals a stepwise transition during ZGA. Nature 2020, 587, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.T.; Muralimanoharan, S.; Gogate, A.A.; Mendelson, C.R. Human Trophoblast Differentiation Is Associated With Profound Gene Regulatory and Epigenetic Changes. Endocrinology 2019, 160, 2189–2203. [Google Scholar] [CrossRef] [PubMed]
- Cañón-Beltrán, K.; Cajas, Y.N.; Peréz-Cerezales, S.; Leal, C.L.V.; Agirregoitia, E.; Gutierrez-Adán, A.; González, E.M.; Rizos, D. Nobiletin enhances the development and quality of bovine embryos in vitro during two key periods of embryonic genome activation. Sci. Rep. 2021, 11, 11796. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.M.; Hoelker, M.; Noferesti, S.S.; Salilew-Wondim, D.; Tholen, E.; Looft, C.; Rings, F.; Uddin, M.J.; Spencer, T.E.; Schellander, K.; et al. Exosomal and Non-Exosomal Transport of Extra-Cellular microRNAs in Follicular Fluid: Implications for Bovine Oocyte Developmental Competence. PLoS ONE 2013, 8, e78505. [Google Scholar] [CrossRef]
- Ikeda, S.; Sugimoto, M.; Kume, S. Lipofection of siRNA into bovine 8-16-cell stage embryos using zona removal and the well-of-the-well culture system. J. Reprod. Dev. 2018, 64, 199–202. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway | Count | p-Value |
|---|---|---|
| Metabolic pathways | 76 | 1.0 × 104 |
| PI3K-Akt signaling pathway | 20 | 3.0 × 102 |
| Carbon metabolism | 10 | 8.0 × 103 |
| Lysosome | 10 | 3.0 × 102 |
| Biosynthesis of amino acids | 9 | 2.0 × 103 |
| Glucagon signaling pathway | 9 | 1.0 × 102 |
| Serotonergic synapse | 9 | 3.0 × 102 |
| Arachidonic acid metabolism | 8 | 2.0 × 102 |
| Starch and sucrose metabolism | 5 | 2.0 × 102 |
| African trypanosomiasis | 5 | 5.0 × 102 |
| GO: Biological process | Count | p-value |
| positive regulation of gene expression | 31 | 1.0 × 103 |
| Cell adhesion | 26 | 1.0 × 103 |
| Lipid metabolic process | 21 | 8.0 × 105 |
| Negative regulation of gene expression | 21 | 4.0 × 103 |
| Defense response to virus | 19 | 2.0 × 102 |
| Positive regulation of cell migration | 18 | 5.0 × 103 |
| Proteolysis | 18 | 1.0 × 102 |
| Homophilic cell adhesion via plasma membrane adhesion molecules | 15 | 3.0 × 103 |
| Axon guidance | 14 | 3.0 × 102 |
| Phosphorylation | 13 | 2.0 × 102 |
| KEGG Pathway | Count | p-Value |
|---|---|---|
| Pathways in cancer | 19 | 2.0 × 102 |
| PI3K-Akt signaling pathway | 14 | 4.0 × 102 |
| Oxytocin signaling pathway | 10 | 3.0 × 103 |
| Cholinergic synapse | 9 | 2.0 × 103 |
| Transcriptional misregulation in cancer | 9 | 4.0 × 102 |
| Phospholipase D signaling pathway | 8 | 3.0 × 102 |
| Efferocytosis | 8 | 4.0 × 102 |
| GO: Biological process | Count | p-value |
| Regulation of transcription from RNA polymerase II promoter | 42 | 8.0 × 103 |
| Negative regulation of transcription from RNA polymerase II promoter | 25 | 2.0 × 103 |
| Positive regulation of transcription from RNA polymerase II promoter | 25 | 1.0 × 102 |
| Cell differentiation | 19 | 5.0 × 103 |
| Activation of GTPase activity | 7 | 1.0 × 102 |
| SMAD protein signal transduction | 6 | 9.0 × 103 |
| Lipid transport | 6 | 1.0 × 102 |
| Positive regulation of endothelial cell migration | 5 | 1.0 × 102 |
| MAPK cascade | 5 | 3.0 × 102 |
| Cellular response to cAMP | 4 | 3.0 × 102 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, S.; Inoue, Y.; Hamazaki, M.; Hara, S.; Noguchi, T.; Shirasuna, K.; Iwata, H. miRNAs in Follicular and Oviductal Fluids Support Global DNA Demethylation in Early-Stage Embryos. Int. J. Mol. Sci. 2024, 25, 5872. https://doi.org/10.3390/ijms25115872
Aoki S, Inoue Y, Hamazaki M, Hara S, Noguchi T, Shirasuna K, Iwata H. miRNAs in Follicular and Oviductal Fluids Support Global DNA Demethylation in Early-Stage Embryos. International Journal of Molecular Sciences. 2024; 25(11):5872. https://doi.org/10.3390/ijms25115872
Chicago/Turabian StyleAoki, Sogo, Yuki Inoue, Mao Hamazaki, Shunsuke Hara, Tatsuo Noguchi, Koumei Shirasuna, and Hisataka Iwata. 2024. "miRNAs in Follicular and Oviductal Fluids Support Global DNA Demethylation in Early-Stage Embryos" International Journal of Molecular Sciences 25, no. 11: 5872. https://doi.org/10.3390/ijms25115872
APA StyleAoki, S., Inoue, Y., Hamazaki, M., Hara, S., Noguchi, T., Shirasuna, K., & Iwata, H. (2024). miRNAs in Follicular and Oviductal Fluids Support Global DNA Demethylation in Early-Stage Embryos. International Journal of Molecular Sciences, 25(11), 5872. https://doi.org/10.3390/ijms25115872