
Proteomic Analysis of Domestic Cat Blastocysts and Their Secretome Produced in an In Vitro Culture System without the Presence of the Zona Pellucida

 and
and
Abstract
1. Introduction
2. Results
2.1. Experiment 1: In Vitro Development and Proteomic Analysis of Domestic Cat Blastocysts Cultured with (ZI) and without (ZF) the Zona Pellucida
2.1.1. In Vitro Development of Domestic Cat Embryos from the ZF and ZI Groups
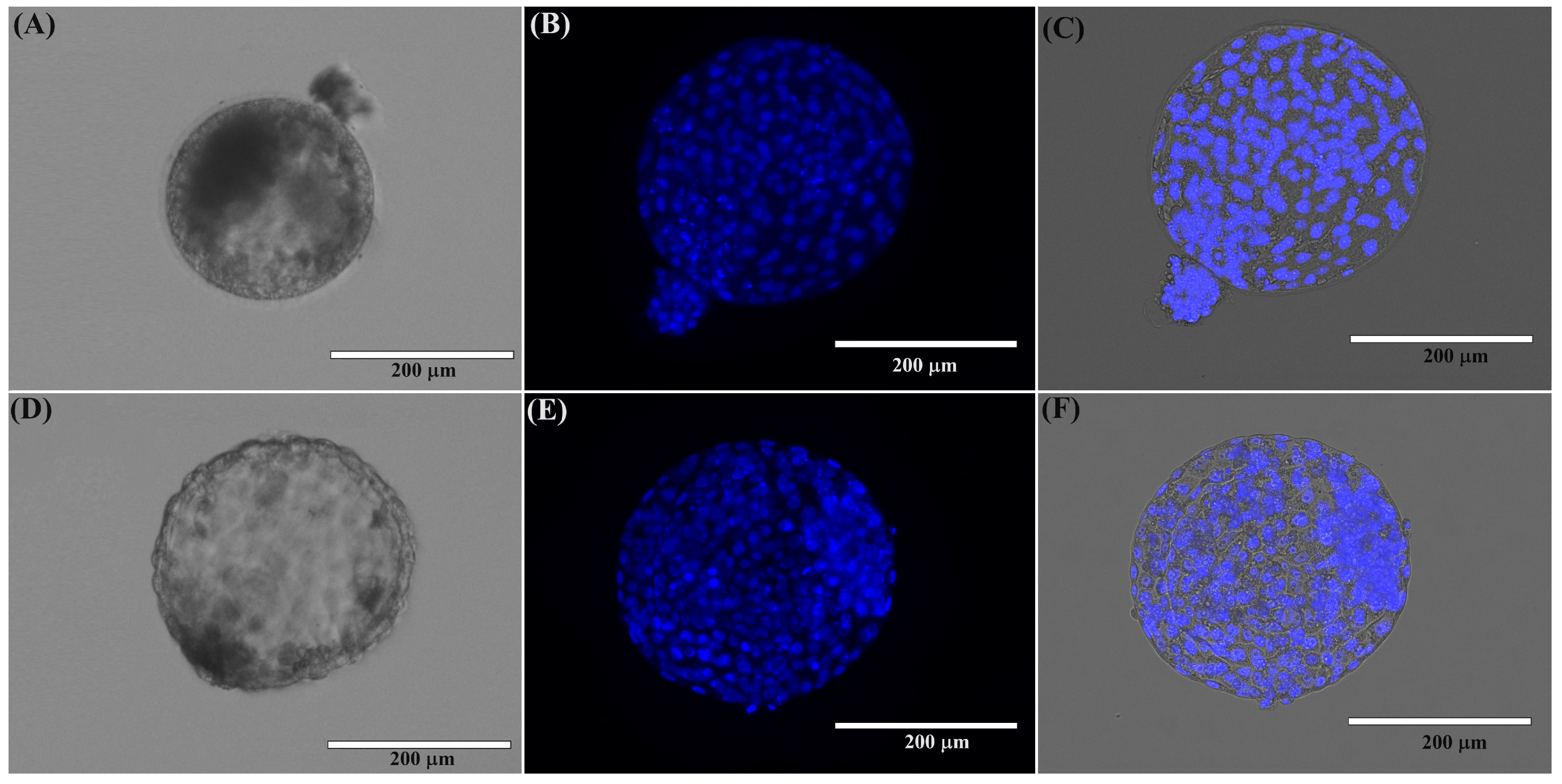
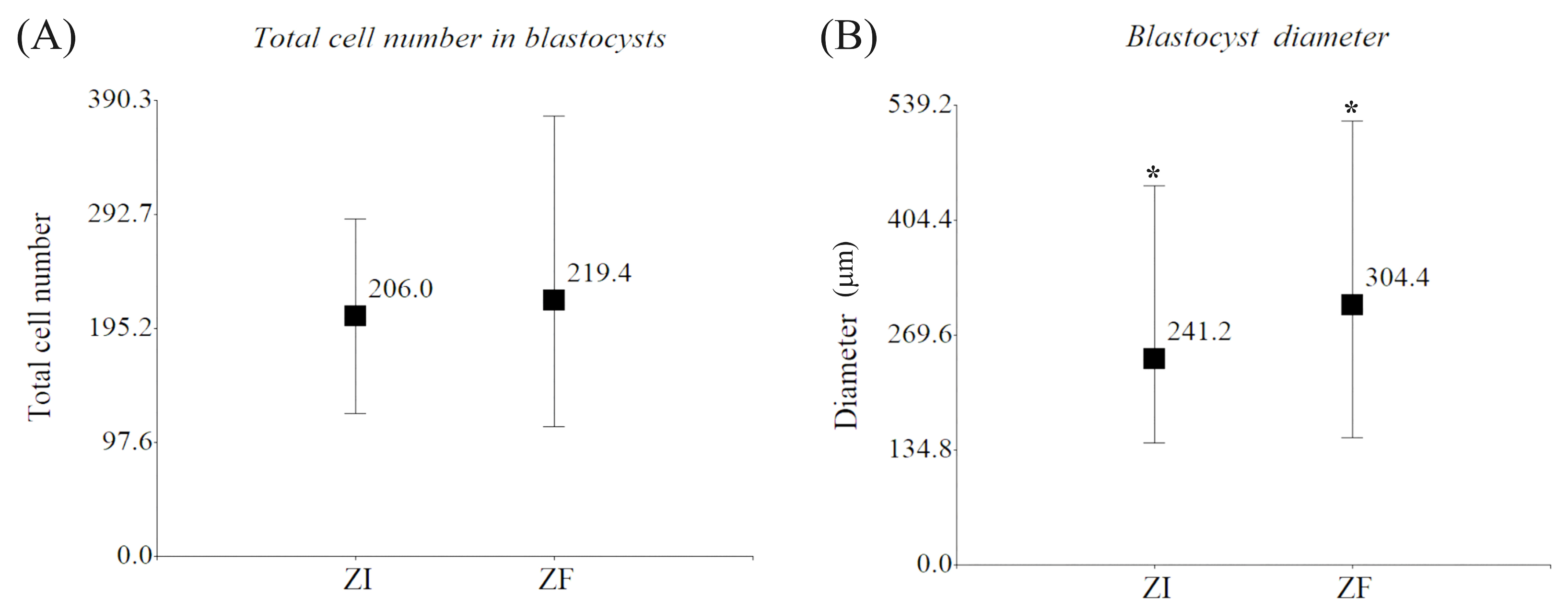
2.1.2. Morphological Evaluation of Domestic Cat Blastocysts
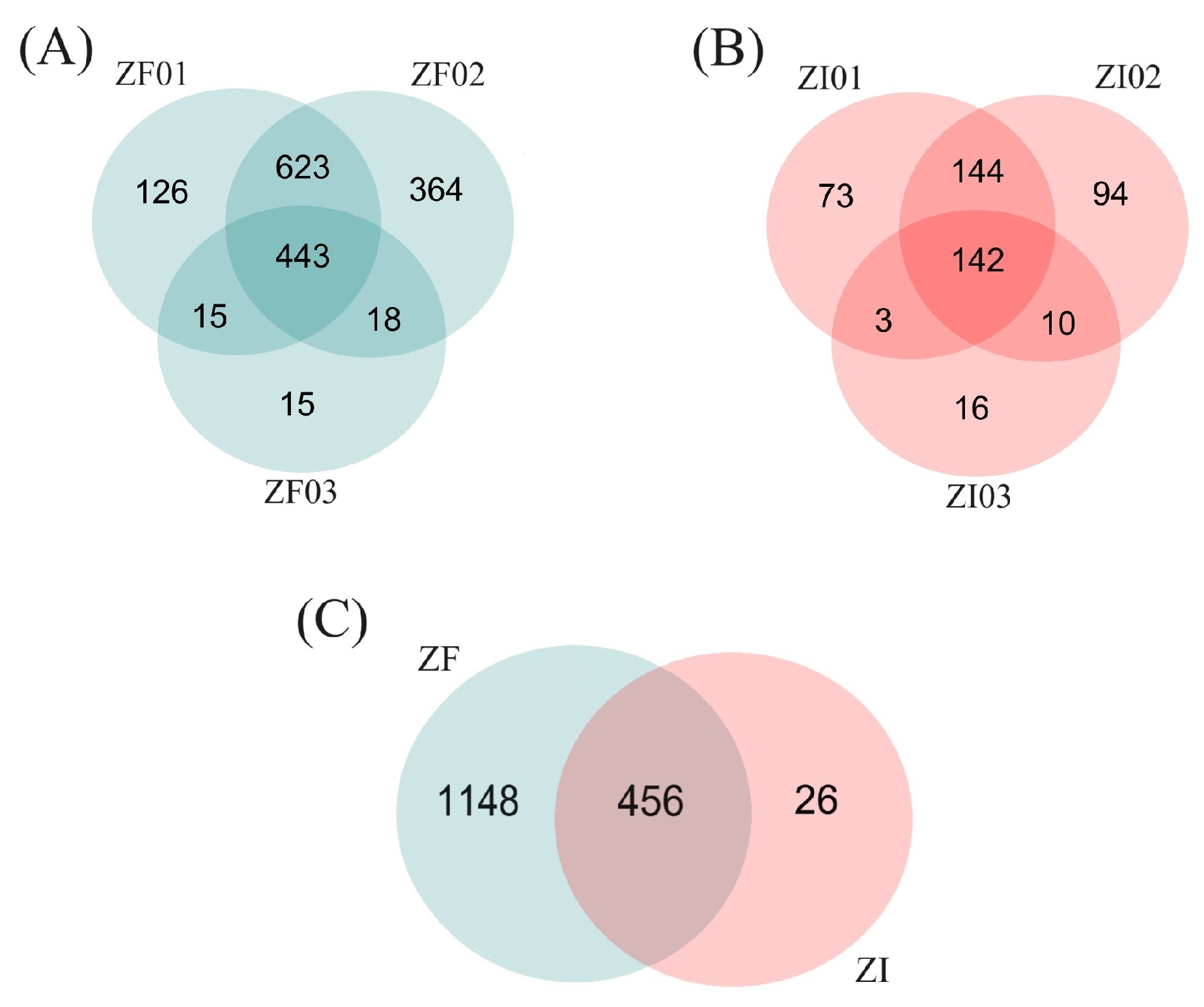
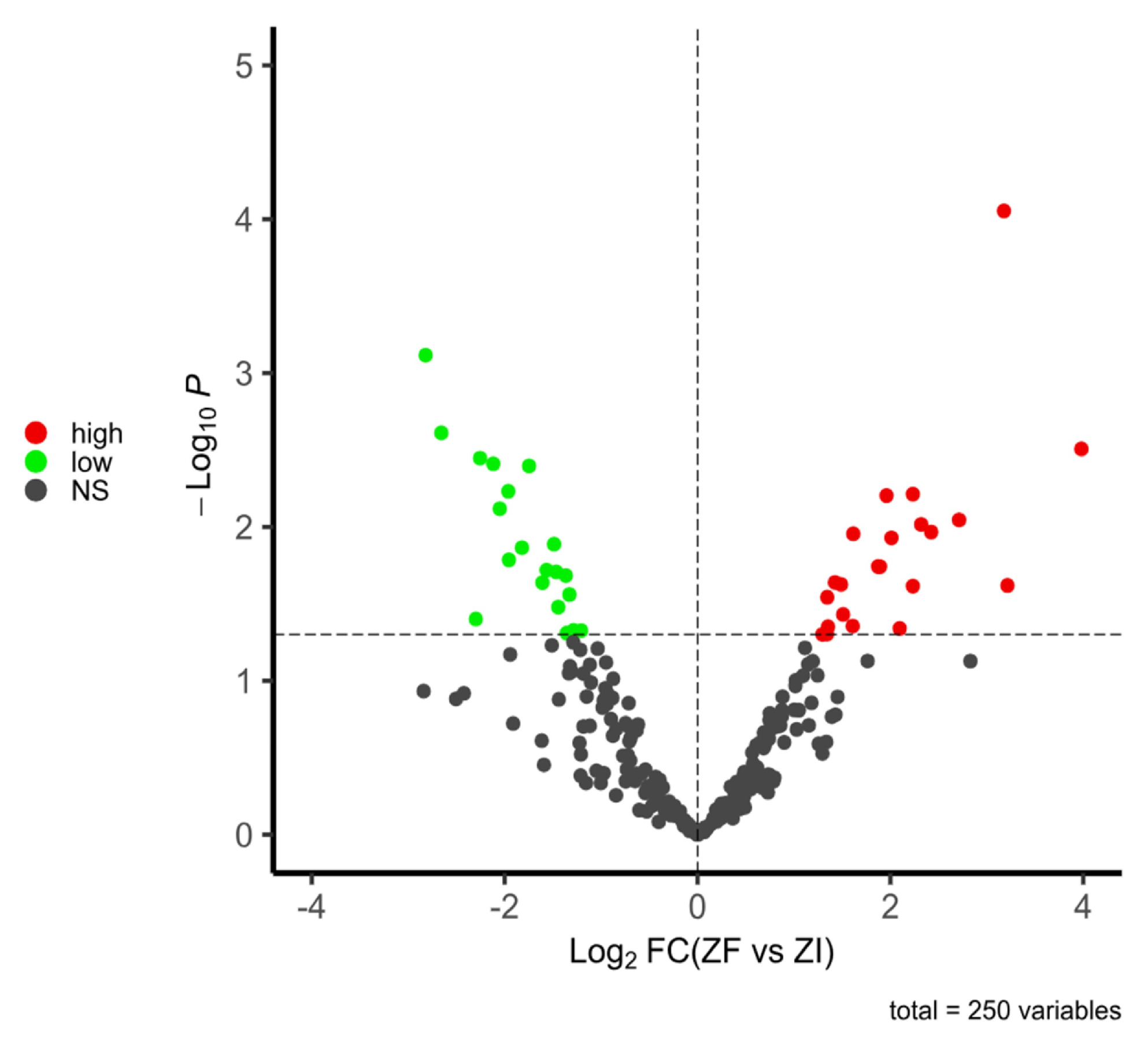
2.1.3. Proteomic Analysis of Domestic Cat ZF and ZI Blastocysts
Protein Identification
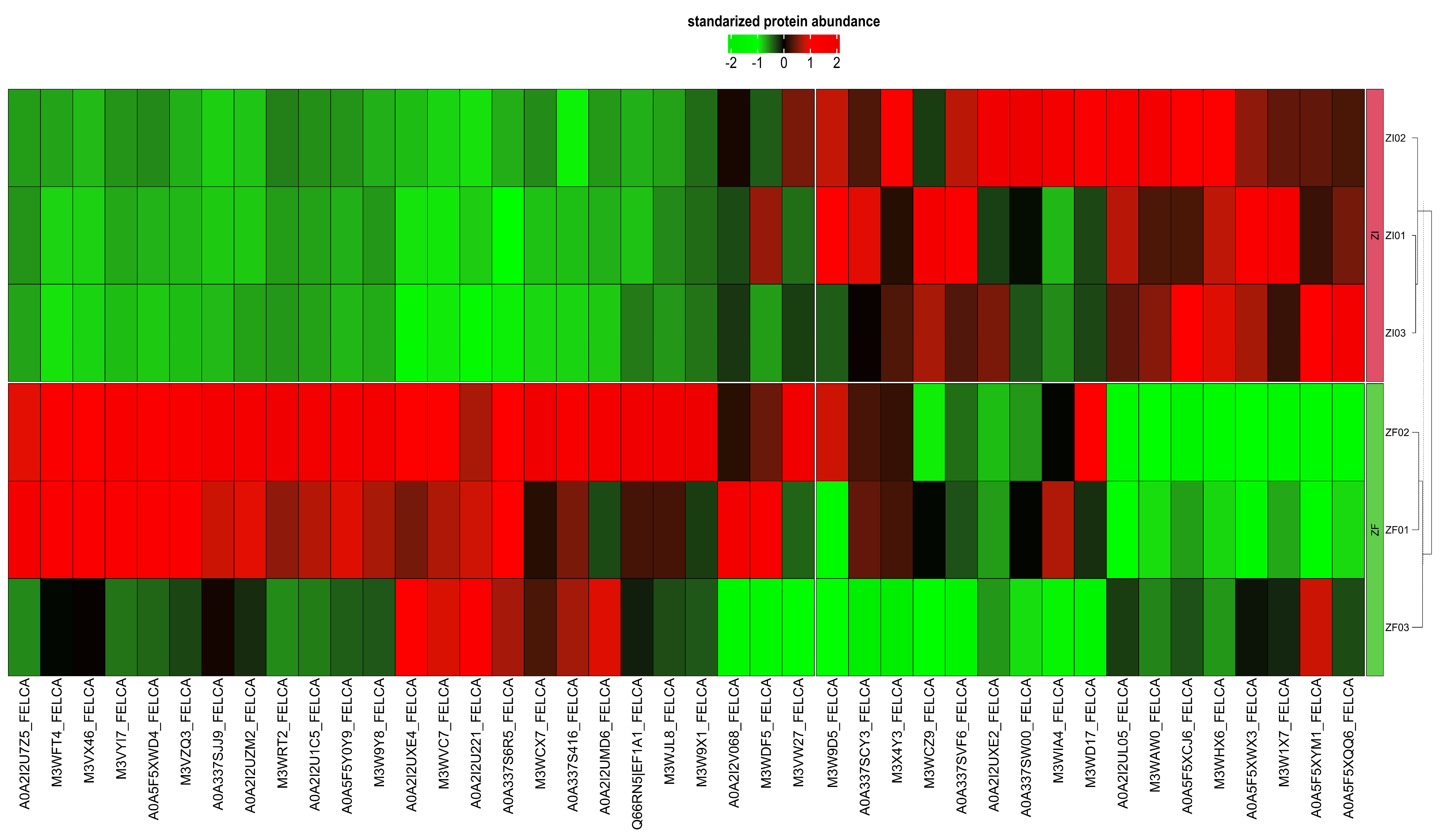
Differential Expression Analysis
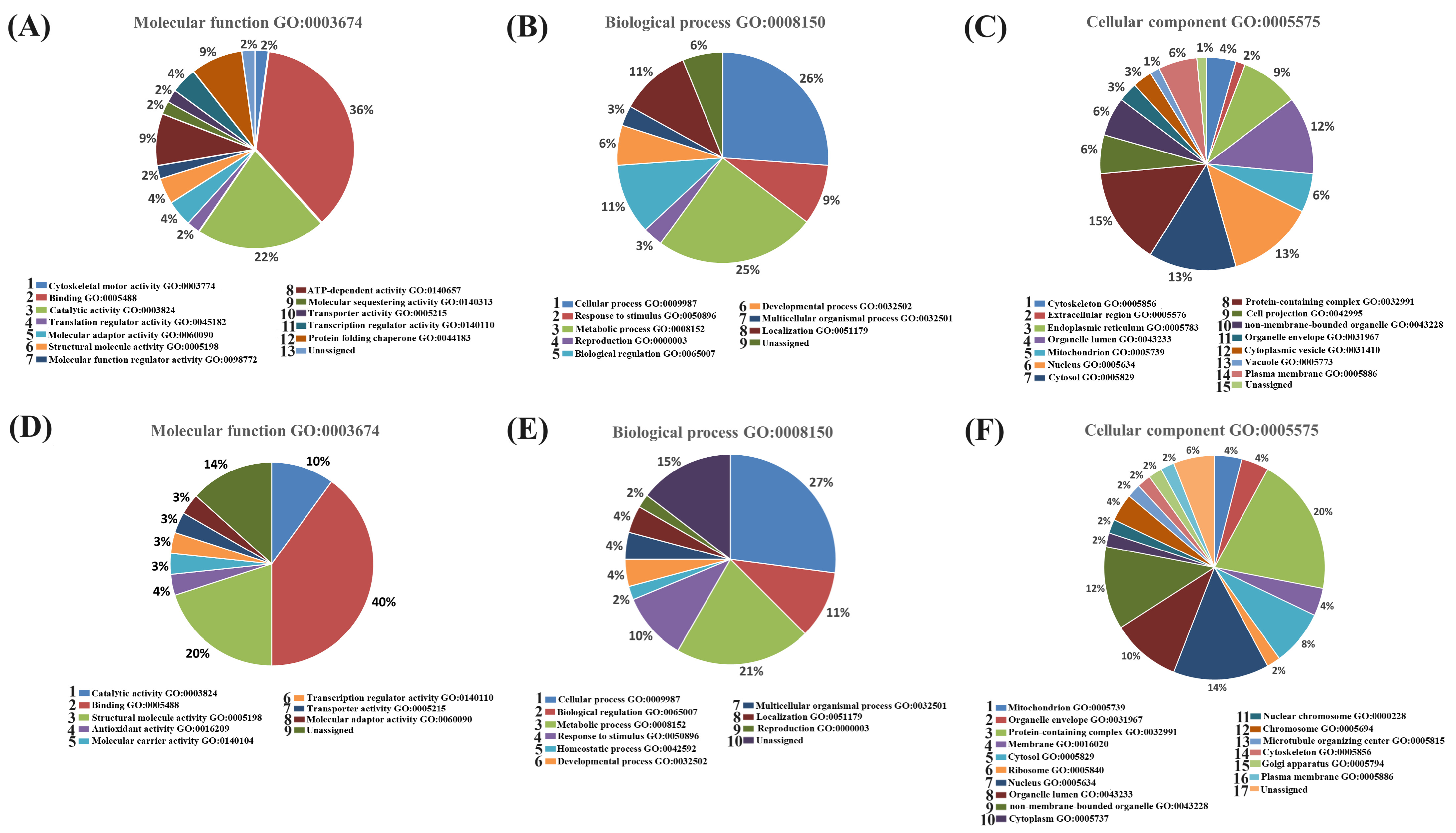
2.1.4. Functional Classification of Differentially Expressed Proteins from the ZF and ZI Blastocysts
2.2. Experiment 2: Proteomic Analysis of the Secretome of Domestic Cat Blastocysts Cultured with and without the Zona Pellucida
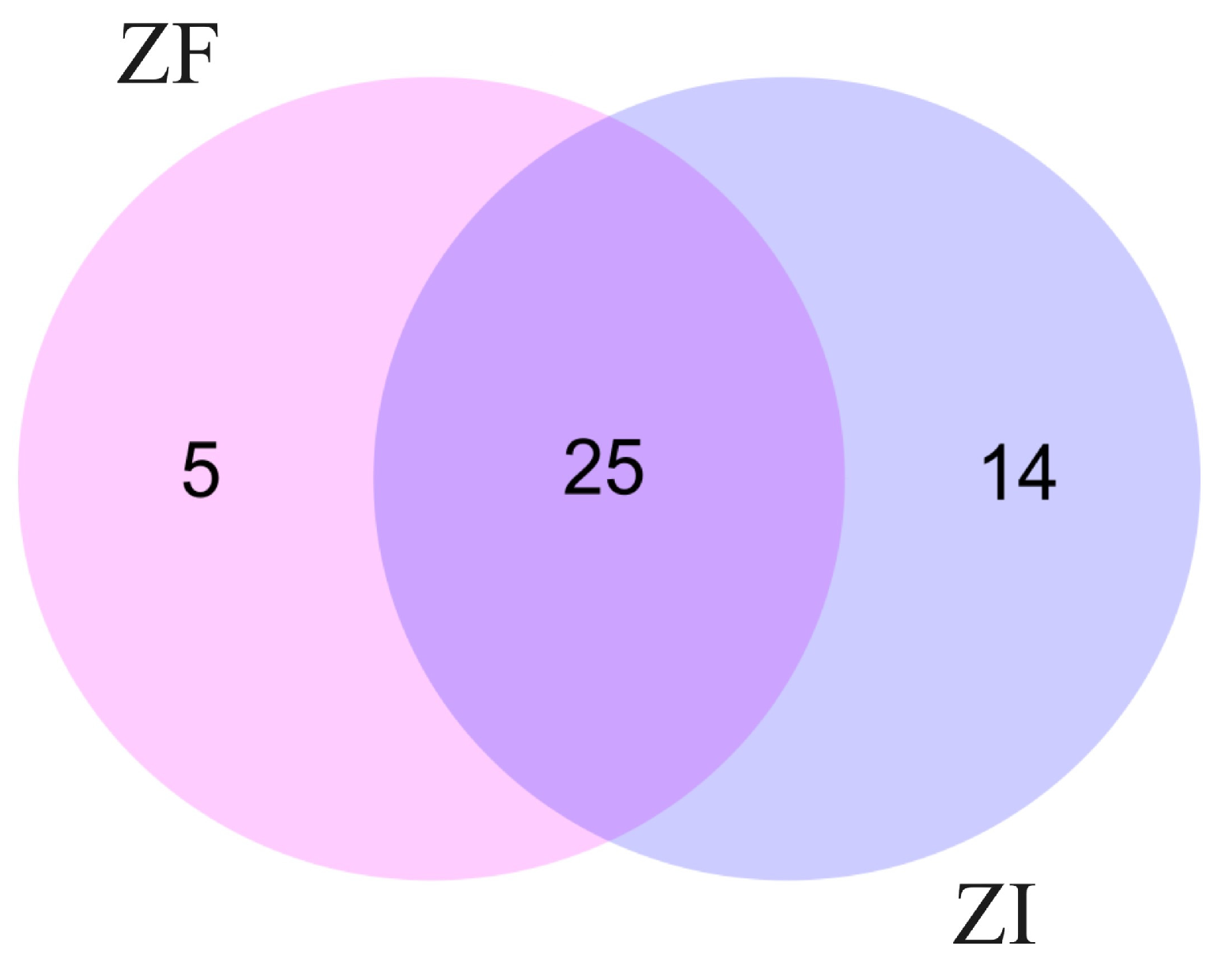
2.2.1. Protein Identification in Conditioned Culture Media of ZF and ZI Blastocysts
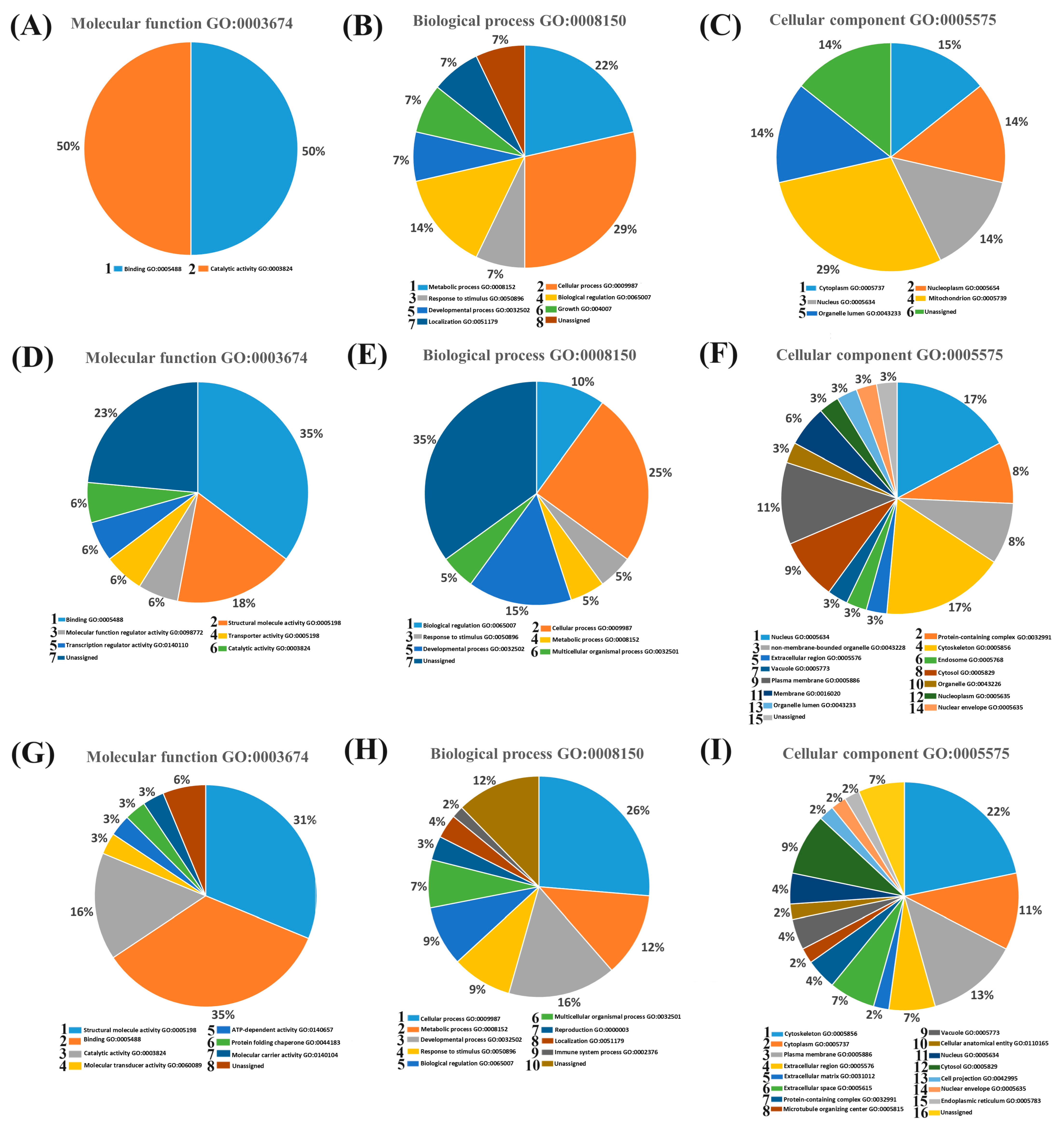
2.2.2. Functional Classification of Proteins Identified in the Conditioned Culture Media
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design
4.3. Ovariohysterectomy and Cumulus–Oocyte Complex (COC) Collection
4.4. In Vitro Maturation
4.5. Sperm Collection and In Vitro Fertilization
4.6. In Vitro Embryo Culture
4.7. Sample Collection
4.8. Morphological Evaluation of Blastocysts
4.8.1. Diameter Measurement
4.8.2. Total Cell Count
4.9. Proteomic Analysis
4.9.1. Protein Extraction
4.9.2. Protein Digestion for Mass Spectrometry
4.9.3. Liquid Chromatography–Tandem Mass Spectrometry (LC–MS/MS)
4.9.4. Protein Identification
4.9.5. Protein Functional Classification
4.10. Statistical Analysis
4.10.1. In Vitro Embryo Development and Morphological Analysis
4.10.2. Protein Quantification LFQ (Label-Free Quantification)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thongphakdee, A.; Sukparangsi, W.; Comizzoli, P.; Chatdarong, K. Reproductive biology and biotechnologies in wild felids. Theriogenology 2020, 150, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.E. Thirty years of assisted reproductive technology in the domestic cat: A selected summary. Rev. Bras. Reprod. Anim. 2019, 43, 129–136. [Google Scholar]
- Songsasen, N.; Comizzoli, P.; Nagashima, J.; Fujihara, M.; Wildt, D.E. The Domestic Dog and Cat as Models for Understanding the Regulation of Ovarian Follicle Development In Vitro. Reprod. Domest. Anim. 2012, 47, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.E. Forty years of assisted reproduction research in non-domestic, wild and endangered mammals. Rev. Bras. Reprod. Anim. 2019, 43, 160–167. [Google Scholar]
- Pope, C.E. Aspects of in vivo oocyte production, blastocyst development, and embryo transfer in the cat. Theriogenology 2014, 81, 126–137. [Google Scholar] [CrossRef]
- Pope, C.E.; Gómez, M.C.; Dresser, B.L. In vitro production and transfer of cat embryos in the 21st century. Theriogenology 2006, 66, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Palomares, F.; Brito, D. What we (don’t) know about the effects of habitat loss and fragmentation on felids. Oryx 2015, 49, 96–106. [Google Scholar] [CrossRef]
- Pukazhenthi, B.S.; Neubauer, K.; Jewgenow, K.; Howard, J.; Wildt, D.E. The impact and potential etiology of teratospermia in the domestic cat and its wild relatives. Theriogenology 2006, 66, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Pukazhenthi, B.S.; Wildt, D.E. Which reproductive technologies are most relevant to studying, managing and conserving wildlife? Reprod. Fertil. Dev. 2003, 16, 33–46. [Google Scholar] [CrossRef]
- Shahverdi, M.; Akbarinejad, V.; Dalman, A.; Hajinasrollah, M.; Vojgani, M.; Vash, N.T.; Nasr-Esfahani, M.H.; Eftekhari-Yazdi, P. Effect of Mechanical Micro-Vibrations on The Efficiency of Leopard Inter-Species Somatic Cell Nuclear Transfer. Cell J. (Yakhteh) 2022, 24, 612–619. [Google Scholar]
- Praxedes, É.A.; Borges, A.A.; Santos, M.V.; Pereira, A.F. Use of somatic cell banks in the conservation of wild felids. Zoo Biol. 2018, 37, 258–263. [Google Scholar] [CrossRef]
- Gómez, M.C.; Pope, C.E.; Ricks, D.M.; Lyons, J.; Dumas, C.; Dresser, B.L. Cloning endangered felids using heterospecific donor oocytes and interspecies embryo transfer. Reprod. Fertil. Dev. 2008, 21, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.C.; Pope, C.E.; Kutner, R.H.; Ricks, D.M.; Lyons, L.A.; Ruhe, M.; Dumas, C.; Lyons, J.; López, M.; Dresser, B.L.; et al. Nuclear transfer of sand cat cells into enucleated domestic cat oocytes is affected by cryopreservation of donor cells. Cloning Stem Cells 2008, 10, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.C.; Earle Pope, C.; Giraldo, A.; Lyons, L.A.; Harris, R.F.; King, A.L.; Cole, A.; Godke, R.A.; Dresser, B.L. Birth of African Wildcat cloned kittens born from domestic cats. Cloning Stem Cells 2004, 6, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.C.; Jenkins, J.A.; Giraldo, A.; Harris, R.F.; King, A.; Dresser, B.L.; Pope, C.E. Nuclear transfer of synchronized African wild cat somatic cells into enucleated domestic cat oocytes. Biol. Reprod. 2003, 69, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Lagutina, I.; Fulka, H.; Lazzari, G.; Galli, C. Interspecies somatic cell nuclear transfer: Advancements and problems. Cell. Reprogram. 2013, 15, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Gómez, M.C.; Pope, C.E.; Biancardi, M.N.; Dumas, C.; Galiguis, J.; Morris, A.C.; Wang, G.; Dresser, B.L. Trichostatin A modified histone covalent pattern and enhanced expression of pluripotent genes in interspecies black-footed cat cloned embryos but did not improve in vitro and in vivo viability. Cell. Reprogram. 2011, 13, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Imsoonthornruksa, S.; Lorthongpanich, C.; Sangmalee, A.; Srirattana, K.; Laowtammathron, C.; Tunwattana, W.; Somsa, W.; Ketudat-Cairns, M.; Parnpai, R. Abnormalities in the transcription of reprogramming genes related to global epigenetic events of cloned endangered felid embryos. Reprod. Fertil. Dev. 2010, 22, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Alvarez, L.; Sharbati, J.; Sharbati, S.; Cox, J.F.; Einspanier, R.; Castro, F.O. Differential gene expression in bovine elongated (Day 17) embryos produced by somatic cell nucleus transfer and in vitro fertilization. Theriogenology 2010, 74, 45–59. [Google Scholar] [CrossRef]
- Vajta, G.; Bartels, P.; Joubert, J.; de la Rey, M.; Treadwell, R.; Callesen, H. Production of a healthy calf by somatic cell nuclear transfer without micromanipulators and carbon dioxide incubators using the Handmade Cloning (HMC) and the Submarine Incubation System (SIS). Theriogenology 2004, 62, 1465–1472. [Google Scholar] [CrossRef]
- Boiani, M.; Eckardt, S.; Leu, N.A.; Schöler, H.R.; McLaughlin, K.J. Pluripotency deficit in clones overcome by clone–clone aggregation: Epigenetic complementation? EMBO J. 2003, 22, 5304–5312. [Google Scholar] [CrossRef] [PubMed]
- Veraguas, D.; Aguilera, C.; Echeverry, D.; Saez-Ruiz, D.; Castro, F.O.; Rodriguez-Alvarez, L. Embryo aggregation allows the production of kodkod (Leopardus guigna) blastocysts after interspecific SCNT. Theriogenology 2020, 158, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Moro, L.N.; Hiriart, M.I.; Buemo, C.P.; Jarazo, J.; Sestelo, A.; Veraguas, D.; Rodriguez-Alvarez, L.; Salamone, D.F. Cheetah interspecific SCNT followed by embryo aggregation improves in vitro development but not pluripotent gene expression. Reproduction 2015, 150, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Moro, L.N.; Jarazo, J.; Buemo, C.; Hiriart, M.I.; Sestelo, A.; Salamone, D.F. Tiger, Bengal and Domestic Cat Embryos Produced by Homospecific and Interspecific Zona-Free Nuclear Transfer. Reprod. Domest. Anim. 2015, 50, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Gambini, A.; Maserati, M. A journey through horse cloning. Reprod. Fertil. Dev. 2018, 30, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Vajta, G. Handmade cloning: The future way of nuclear transfer? Trends Biotechnol. 2007, 25, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Cortez, J.V.; Vajta, G.; Valderrama, N.M.; Portocarrero, G.S.; Quintana, J.M. High pregnancy and calving rates with a limited number of transferred handmade cloned bovine embryos. Cell. Reprogram. 2018, 20, 4–8. [Google Scholar] [CrossRef]
- Gambini, A.; Jarazo, J.; Olivera, R.; Salamone, D.F. Equine cloning: In vitro and in vivo development of aggregated embryos. Biol. Reprod. 2012, 87, 15. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Kragh, P.M.; Zhang, Y.; Li, J.; Schmidt, M.; Bøgh, I.B.; Zhang, X.; Purup, S.; Jørgensen, A.L.; Pedersen, A.M.; et al. Piglets born from handmade cloning, an innovative cloning method without micromanipulation. Theriogenology 2007, 68, 1104–1110. [Google Scholar] [CrossRef]
- Kanda, M.; Oikawa, H.; Nakao, H.; Tsutsui, T. Early embryonic development in vitro and embryo transfer in the cat. J. Veter-Med. Sci. 1995, 57, 641–646. [Google Scholar] [CrossRef]
- Malin, K.; Witkowska-Piłaszewicz, O.; Papis, K. The many problems of somatic cell nuclear transfer in reproductive cloning of mammals. Theriogenology 2022, 189, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Veraguas-Davila, D.; Cordero, M.F.; Saez, S.; Saez-Ruiz, D.; Gonzalez, A.; Saravia, F.; Castro, F.O.; Rodriguez-Alvarez, L. Domestic cat embryos generated without zona pellucida are capable of developing in vitro but exhibit abnormal gene expression and a decreased implantation rate. Theriogenology 2021, 174, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Veraguas-Dávila, D.; Saéz-Ruíz, D.; Álvarez, M.C.; Saravia, F.; Castro, F.O.; Rodríguez-Alvarez, L. Analysis of trophectoderm markers in domestic cat blastocysts cultured without zona pellucida. Zygote 2022, 30, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Huang, T.; Wu, T.; Bai, H.; Kawahara, M.; Takahashi, M. Zona pellucida removal by acid Tyrode’s solution affects pre-and post-implantation development and gene expression in mouse embryos. Biol. Reprod. 2022, 107, 1228–1241. [Google Scholar] [CrossRef] [PubMed]
- Veraguas-Dávila, D.; Caamaño, D.; Saéz-Ruiz, D.; Vásquez, Y.; Saravia, F.; Castro, F.O.; Rodríguez-Alvarez, L. Zona pellucida removal modifies the expression and release of specific microRNAs in domestic cat blastocysts. Zygote 2023, 31, 544–556. [Google Scholar] [CrossRef]
- Gygi, S.; Rochon, Y.; Franza, R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Katz-Jaffe, M.G.; Gardner, D.K.; Schoolcraft, W.B. Proteomic analysis of individual human embryos to identify novel biomarkers of development and viability. Fertil. Steril. 2006, 85, 101–107. [Google Scholar] [CrossRef]
- Katz-Jaffe, M.G.; McReynolds, S.; Gardner, D.K.; Schoolcraft, W.B. The role of proteomics in defining the human embryonic secretome. Mol. Hum. Reprod. 2009, 15, 271–277. [Google Scholar] [CrossRef]
- Hathout, Y. Approaches to the study of the cell secretome. Expert Rev. Proteom. 2007, 4, 239–248. [Google Scholar] [CrossRef]
- Hu, S.; Loo, J.A.; Wong, D.T. Human body fluid proteome analysis. Proteomics 2006, 6, 6326–6353. [Google Scholar] [CrossRef]
- Kulasingam, V.; Diamandis, E.P. Proteomics analysis of conditioned media from three breast cancer cell lines: A mine for biomarkers and therapeutic targets. Mol. Cell. Proteom. 2007, 6, 1997–2011. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Corrales, F.J.; Caamaño, J.N.; Díez, C.; Trigal, B.; Mora, M.I.; Martín, D.; Carrocera, S.; Gómez, E. Proteome of the early embryo–maternal dialogue in the cattle uterus. J. Proteome Res. 2012, 11, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Katz-Jaffe, M.G.; Linck, D.W.; Schoolcraft, W.B.; Gardner, D.K. A proteomic analysis of mammalian preimplantation embryonic development. Reproduction 2005, 130, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, J.; Wang, Y.; Zhao, W.; Wang, W.; Cui, J.; Yang, J.; Yue, Y.; Zhang, S.; Chu, M.; et al. A proteomic atlas of ligand–receptor interactions at the ovine maternal–fetal interface reveals the role of histone lactylation in uterine remodeling. J. Biol. Chem. 2022, 298, 101456. [Google Scholar] [CrossRef] [PubMed]
- Banliat, C.; Mahé, C.; Lavigne, R.; Com, E.; Pineau, C.; Labas, V.; Guyonnet, B.; Mermillod, P.; Saint-Dizier, M. The proteomic analysis of bovine embryos developed in vivo or in vitro reveals the contribution of the maternal environment to early embryo. BMC Genom. 2022, 23, 839. [Google Scholar] [CrossRef] [PubMed]
- Bragança, G.M.; Alcantara-Neto, A.S.; Batista, R.I.; Brandao, F.Z.; Freitas, V.J.; Mermillod, P.; Souza-Fabjan, J.M. Oviduct fluid during IVF moderately modulates polyspermy in in vitro-produced goat embryos during the non-breeding season. Theriogenology 2021, 168, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Almiñana, C.; Corbin, E.; Tsikis, G.; Alcântara-Neto, A.S.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct extracellular vesicles protein content and their role during oviduct–embryo cross-talk. Reproduction 2017, 154, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Grabacka, M.M.; Gawin, M.; Pierzchalska, M. Phytochemical modulators of mitochondria: The search for chemopreventive agents and supportive therapeutics. Pharmaceuticals 2014, 7, 913–942. [Google Scholar] [CrossRef]
- Hao, J.; Hu, H.; Jiang, Z.; Yu, X.; Li, C.; Chen, L.; Xia, Y.; Liu, D.; Wang, D. microRNA-670 modulates Igf2bp1 expression to regulate RNA methylation in parthenogenetic mouse embryonic development. Sci. Rep. 2020, 10, 4782. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, W.K.; Colleoni, S.; Galli, C.; Paris, D.B.; Colenbrander, B.; Roelen, B.A.; Stout, T.A. Maternal age and in vitro culture affect mitochondrial number and function in equine oocytes and embryos. Reprod. Fertil. Dev. 2015, 27, 957–968. [Google Scholar] [CrossRef]
- Trifonova, E.A.; Gavrilenko, M.M.; Babovskaya, A.A.; Zarubin, A.A.; Svarovskaya, M.G.; Izhoykina, E.V.; Stepanov, I.A.; Serebrova, V.N.; Kutsenko, I.G.; Stepanov, V.A. Alternative Splicing Landscape of Placental Decidual Cells during Physiological Pregnancy. Russ. J. Genet. 2022, 58, 1257–1265. [Google Scholar] [CrossRef]
- Leoni, G.G.; Berlinguer, F.; Succu, S.; Bebbere, D.; Mossa, F.; Madeddu, M.; Ledda, S.; Bogliolo, L.; Naitana, S. A new selection criterion to assess good quality ovine blastocysts after vitrification and to predict their transfer into recipients. Mol. Reprod. Dev. 2008, 75, 373–382. [Google Scholar] [CrossRef]
- Dun, M.D.; Smith, N.D.; Baker, M.A.; Lin, M.; Aitken, R.J.; Nixon, B. The chaperonin containing TCP1 complex (CCT/TRiC) is involved in mediating sperm-oocyte interaction. J. Biol. Chem. 2011, 286, 36875–36887. [Google Scholar] [CrossRef]
- Del Alamo, M.R.; Katila, T.; Palviainen, M.; Reilas, T. Effects of intrauterine devices on proteins in the uterine lavage fluid of mares. Theriogenology 2021, 165, 1–9. [Google Scholar] [CrossRef]
- Kepkova, K.V.; Vodicka, P.; Toralova, T.; Lopatarova, M.; Cech, S.; Dolezel, R.; Havlicek, V.; Besenfelder, U.; Kuzmany, A.; Sirard, M.-A.; et al. Transcriptomic analysis of in vivo and in vitro produced bovine embryos revealed a developmental change in cullin 1 expression during maternal-to-embryonic transition. Theriogenology 2011, 75, 1582–1595. [Google Scholar] [CrossRef]
- Yi, Y.; Zeng, Y.; Sam, T.W.; Hamashima, K.; Tan, R.J.; Warrier, T.; Phua, J.X.; Taneja, R.; Liou, Y.-C.; Li, H.; et al. Ribosomal proteins regulate 2-cell-stage transcriptome in mouse embryonic stem cells. Stem Cell Rep. 2023, 18, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Bastos, H.B.; Martinez, M.N.; Camozzato, G.C.; Estradé, M.J.; Barros, E.; Vital, C.E.; Vidigal, P.M.; Meikle, A.; Jobim, M.I.; Gregory, R.M.; et al. Proteomic profile of histotroph during early embryo development in mares. Theriogenology 2019, 125, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.M.; Lloyd, R.E.; Fazeli, A.; Sostaric, E.; Georgiou, A.S.; Satake, N.; Watson, P.F.; Holt, W.V. Effects of HSPA8, an evolutionarily conserved oviductal protein, on boar and bull spermatozoa. Reproduction 2009, 137, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Berryman, M.; Franck, Z.; Bretscher, A. Ezrin is concentrated in the apical microvilli of a wide variety of epithelial cells whereas moesin is found primarily in endothelial cells. J. Cell Sci. 1993, 105, 1025–1043. [Google Scholar] [CrossRef]
- Hwang, I.S.; Shim, J.; Oh, K.B.; Lee, H.; Park, M.R. cd26 Knockdown Negatively Affects Porcine Parthenogenetic Preimplantation Embryo Development. Animals 2022, 12, 1662. [Google Scholar] [CrossRef]
- Cui, X.S.; Li, X.Y.; Jeong, Y.J.; Jun, J.H.; Kim, N.H. Gene expression of cox5a, 5b, or 6b1 and their roles in preimplantation mouse embryos. Biol. Reprod. 2006, 74, 601–610. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Athavale, D.M.; Barré, A.; Kranyak, A.C.; Lal, A.; Blalock, J.L.; Zimmerman, S.; Chang, T.A.; Robinson, R.D.; Wininger, J.D.; Roudebush, W.E.; et al. Pro-apoptotic gene expression in blastocoel fluid from euploid day-5 embryos is associated with negative pregnancy outcomes. Fertil. Steril. 2019, 112, e261. [Google Scholar] [CrossRef]
- Rincón, J.A.; Pradieé, J.; Remião, M.H.; Collares, T.V.; Mion, B.; Gasperin, B.G.; Rovani, M.T.; Corrêa, M.N.; Pegoraro, L.M.C.; Schneider, A. Effect of high-density lipoprotein on oocyte maturation and bovine embryo development in vitro. Reprod. Domest. Anim. 2019, 54, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Loyer, P.; Trembley, J.H.; Katona, R.; Kidd, V.J.; Lahti, J.M. Role of CDK/cyclin complexes in transcription and RNA splicing. Cell. Signal. 2005, 17, 1033–1051. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, J.; Yu, W.; Liu, G.; Chen, Q. Expression of stage-specific genes during zygotic gene activation in preimplantation mouse embryos. Zool. Sci. 2003, 20, 1389–1393. [Google Scholar] [CrossRef] [PubMed]
- Soto-Moreno, E.J.; Balboula, A.; Spinka, C.; Rivera, R.M. Serum supplementation during bovine embryo culture affects their development and proliferation through macroautophagy and endoplasmic reticulum stress regulation. PLoS ONE 2021, 16, e0260123. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Vassena, R.; Latham, K.E. Effects of in vitro oocyte maturation and embryo culture on the expression of glucose transporters, glucose metabolism and insulin signaling genes in rhesus monkey oocytes and preimplantation embryos. Mol. Hum. Reprod. 2007, 13, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Mohebi, M.; Berjis, K.; Ghahremani, A.; Modarressi, M.H.; Ghafouri-Fard, S. A novel variant in TLE6 is associated with embryonic developmental arrest (EDA) in familial female infertility. Sci. Rep. 2022, 12, 17664. [Google Scholar] [CrossRef]
- Liu, J.; Tan, Z.; He, J.; Jin, T.; Han, Y.; Hu, L.; Huang, S. Two novel mutations in PADI6 and TLE6 genes cause female infertility due to arrest in embryonic development. J. Assist. Reprod. Genet. 2021, 38, 1551–1559. [Google Scholar] [CrossRef]
- Alazami, A.M.; Awad, S.M.; Coskun, S.; Al-Hassan, S.; Hijazi, H.; Abdulwahab, F.M.; Poizat, C.; Alkuraya, F.S. TLE6 mutation causes the earliest known human embryonic lethality. Genome Biol. 2015, 16, 240. [Google Scholar] [CrossRef]
- Barrientos, G.; Freitag, N.; Tirado-Gonzalez, I.; Unverdorben, L.; Jeschke, U.; Thijssen, V.L.; Blois, S.M. Involvement of galectin-1 in reproduction: Past, present and future. Hum. Reprod. Update 2014, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Noli, L.; Khorsandi, S.E.; Pyle, A.; Giritharan, G.; Fogarty, N.; Capalbo, A.; Devito, L.; Jovanovic, V.M.; Khurana, P.; Rosa, H.; et al. Effects of thyroid hormone on mitochondria and metabolism of human preimplantation embryos. Stem Cells 2020, 38, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.; Jensen, A.M.; Harboe, T.; Møgster, M.; Jensen, R.M.; Mjaavatten, O.; Birkeland, E.; Spriet, E.; Sandven, L.; Furmanek, T.; et al. Molecular mechanisms involved in Atlantic halibut (Hippoglossus hippoglossus) egg quality: Impairments at transcription and protein folding levels induce inefficient protein and energy homeostasis during early development. bioRxiv 2022. [Google Scholar] [CrossRef]
- Uniprot. 2024. Available online: https://www.uniprot.org/uniprotkb/A0A2I2V068/entry (accessed on 15 February 2024).
- Hwang, K.C.; Cui, X.S.; Park, S.P.; Shin, M.R.; Park, S.Y.; Kim, E.Y.; Kim, N.H. Identification of differentially regulated genes in bovine blastocysts using an annealing control primer system. Mol. Reprod. Dev. 2004, 69, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Paudel, D.; Ouyang, Y.; Huang, Q.; Zhou, W.; Wang, J.; Poorekhorsandi, M.E.; Dhakal, B.; Tong, X. Expression of TRA2B in endometrial carcinoma and its regulatory roles in endometrial carcinoma cells. Oncol. Lett. 2019, 18, 2455–2463. [Google Scholar] [CrossRef] [PubMed]
- Dhaenens, L.; Lierman, S.; De Clerck, L.; Govaert, E.; Deforce, D.; Tilleman, K.; De Sutter, P. Endometrial stromal cell proteome mapping in repeated implantation failure and recurrent pregnancy loss cases and fertile women. Reprod. Biomed. Online 2019, 38, 442–454. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, X.; Tang, B.; Li, C.; Kou, Z.; Li, L.; Liu, W.; Wu, Y.; Kou, X.; Li, J.; et al. Protein expression landscape of mouse embryos during pre-implantation development. Cell Rep. 2017, 21, 3957–3969. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sandhu, S.; Patel, N.; Triggs-Raine, B.; Ding, H. EMG1 is essential for mouse pre-implantation embryo development. BMC Dev. Biol. 2010, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.Y.; Wood, J.L.; Minter-Dykhouse, K.; Ye, L.; Saunders, T.L.; Yu, X.; Chen, J. Polo-like kinase 1 is essential for early embryonic development and tumor suppression. Mol. Cell. Biol. 2008, 28, 6870–6876. [Google Scholar] [CrossRef]
- Wang, W.; Miyamoto, Y.; Chen, B.; Shi, J.; Diao, F.; Zheng, W.; Li, Q.; Yu, L.; Li, L.; Xu, Y.; et al. Karyopherin α deficiency contributes to human preimplantation embryo arrest. J. Clin. Investig. 2023, 133, e159951. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, Z.; Zhang, D.; Zhao, B.; Liu, L.; Xie, Z.; Yao, Y.; Zheng, P. KHDC3L mutation causes recurrent pregnancy loss by inducing genomic instability of human early embryonic cells. PLoS Biol. 2019, 17, e3000468. [Google Scholar] [CrossRef] [PubMed]
- Salilew-Wondim, D.; Tesfaye, D.; Rings, F.; Held-Hoelker, E.; Miskel, D.; Sirard, M.A.; Tholen, E.; Schellander, K.; Hoelker, M. The global gene expression outline of the bovine blastocyst: Reflector of environmental conditions and predictor of developmental capacity. BMC Genom. 2021, 22, 408. [Google Scholar] [CrossRef]
- Kakourou, G.; Jaroudi, S.; Tulay, P.; Heath, C.; Serhal, P.; Harper, J.C.; SenGupta, S.B. Investigation of gene expression profiles before and after embryonic genome activation and assessment of functional pathways at the human metaphase II oocyte and blastocyst stage. Fertil. Steril. 2013, 99, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Z.; Lin, E.; He, P.; Ru, G. Oxidative damage-induced hyperactive ribosome biogenesis participates in tumorigenesis of offspring by cross-interacting with the Wnt and TGF-β1 pathways in IVF embryos. Exp. Mol. Med. 2021, 53, 1792–1806. [Google Scholar] [CrossRef]
- Ni, D.; Huang, X.; Wang, Z.; Deng, L.; Zeng, L.; Zhang, Y.; Lu, D.; Zou, X. Expression characterization and transcription regulation analysis of porcine Yip1 domain family member 3 gene. Asian-Australas J. Anim. Sci. 2020, 33, 398. [Google Scholar] [CrossRef]
- Jia, J.; Han, Z.; Wang, X.; Zheng, X.; Wang, S.; Cui, Y. H2B gene family: A prognostic biomarker and correlates with immune infiltration in glioma. Front. Oncol. 2022, 12, 966817. [Google Scholar] [CrossRef]
- Salilew-Wondim, D.; Saeed-Zidane, M.; Hoelker, M.; Gebremedhn, S.; Poirier, M.; Pandey, H.O.; Tholen, E.; Neuhoff, C.; Held, E.; Besenfelder, U.; et al. Genome-wide DNA methylation patterns of bovine blastocysts derived from in vivo embryos subjected to in vitro culture before, during or after embryonic genome activation. BMC Genom. 2018, 19, 424. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Yuan, Z.; Ye, A.; Wu, T.; Jia, J.; Guo, J.; Zhang, J.; Li, T.; Cheng, X. Low GNG12 expression predicts adverse outcomes: A potential therapeutic target for osteosarcoma. Front. Immunol. 2021, 12, 758845. [Google Scholar] [CrossRef]
- Larson, K.C.; Draper, M.P.; Lipko, M.; Dabrowski, M. Gng12 is a novel negative regulator of LPS-induced inflammation in the microglial cell line BV-2. Inflamm. Res. 2010, 59, 15–22. [Google Scholar] [CrossRef]
- Zhu, J.; Zheng, Y.; Liu, Y.; Chen, M.; Liu, Y.; Li, J. Association between HMGA1 and immunosuppression in hepatocellular carcinoma: A comprehensive bioinformatics analysis. Medicine 2023, 102, e32707. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, Q.; Xie, L. The AlkB Family of Fe (II)/Alpha-Ketoglutarate-Dependent Dioxyg enases Modulates Embryogenesis through Epigenetic Regulation. Curr. Stem Cell Res. Ther. 2018, 13, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, D.; Denchi, E.L.; Danovi, D.; Jochemsen, A.; Capillo, M.; Gobbi, A.; Helin, K.; Pelicci, P.G.; Marine, J.-C. Mdm4 (Mdmx) regulates p53-induced growth arrest and neuronal cell death during early embryonic mouse development. Mol. Cell. Biol. 2002, 22, 5527–5538. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; Gardner, D.K. Lactate Regulates Pyruvate Uptake and Metabolism in the Preimplantation Mouse Embryo. Biol. Reprod. 2000, 62, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K.; Huet-Hudson, Y.M.; Paria, B.C.; McMaster, M.T.; De, S.K.; Dey, S.K. Metallothionein gene expression and metal regulation during preimplantation mouse embryo development (MT mRNA during early development). Dev. Biol. 1991, 145, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kramer, A.C.; McLendon, B.A.; Cain, J.W.; Burghardt, R.C.; Wu, G.; Bazer, F.W.; Johnson, G.A. Elongating porcine conceptuses can utilize glutaminolysis as an anaplerotic pathway to maintain the TCA cycle. Biol. Reprod. 2022, 107, 823–833. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Hou, W.; Yang, X.; Liu, X.; Zhang, Y.; Gao, M.; Zong, M.; Dong, Z.; Liu, Z.; et al. Metabolic control of histone acetylation for precise and timely regulation of minor ZGA in early mammalian embryos. Cell Discov. 2022, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Ooga, M.; Suzuki, M.G.; Aoki, F. Involvement of histone H2B monoubiquitination in the regulation of mouse preimplantation development. J. Reprod. Dev. 2015, 61, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, H.; Wang, X.; Cui, X.; Jin, L. Keratin 86 is up-regulated in the uterus during implantation, induced by oestradiol. BMC Dev. Biol. 2020, 20, 3. [Google Scholar] [CrossRef]
- Wang, B.; Shao, Y. Annexin A2 acts as an adherent molecule under the regulation of steroids during embryo implantation. Mol. Hum. Reprod. 2020, 26, 825–836. [Google Scholar] [CrossRef]
- Kofron, M.; Spagnuolo, A.; Klymkowsky, M.; Wylie, C.; Heasman, J. The roles of maternal α-catenin and plakoglobin in the early Xenopus embryo. Development 1997, 124, 1553–1560. [Google Scholar] [CrossRef]
- Milde, S.; Viebahn, C.; Kirchner, C. Proteolipid protein 2 mRNA is expressed in the rabbit embryo during gastrulation. Mech. Dev. 2001, 106, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, S.; Qian, X.; Kuai, Y.; Xu, Y. The inclusion principles of human embryos in the wow-based time-lapse system: A retrospective cohort study. Front. Endocrinol. 2021, 12, 549216. [Google Scholar] [CrossRef] [PubMed]
- Curry, E.; Easley, J.; Wojtusik, J.; Roth, T. Identification of mink (Neovison vison) fecal proteins during embryonic diapause and placental pregnancy for non-invasive pregnancy diagnosis in wildlife. In Proceedings of the III International Symposium on Embryonic Diapause, Ascona, Switzerland, 2–6 June 2020; pp. 2–6. [Google Scholar]
- Dracopoli, N.; Brett, F. Serum aminopeptidases in pregnant vervet monkeys. Biochem. Genet. 1982, 20, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Keil, R.; Rietscher, K.; Hatzfeld, M. Antagonistic regulation of intercellular cohesion by plakophilins 1 and 3. J. Investig. Dermatol. 2016, 136, 2022–20299. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Green, K.J. Regulation of desmosome assembly and adhesion. Semin. Cell Dev. Biol. 2004, 15, 665–677. [Google Scholar] [CrossRef]
- Fleming, T.P.; Garrod, D.R.; Elsmore, A.J. Desmosome biogenesis in the mouse preimplantation embryo. Development 1991, 112, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Jacob, J.T.; Coulombe, P.A.; Kwan, R.; Omary, M.B. Types I and II keratin intermediate filaments. Cold Spring Harb. Perspect. Biol. 2018, 10, a018275. [Google Scholar] [CrossRef] [PubMed]
- Starkova, T.Y.; Artamonova, T.O.; Ermakova, V.V.; Chikhirzhina, E.V.; Khodorkovskii, M.A.; Tomilin, A.N. The profile of post-translational modifications of histone H1 in chromatin of mouse embryonic stem cells. Acta Naturae 2019, 11, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Patterson, M.; Mikkola, H.K.; Lowry, W.E.; Kurdistani, S.K. Dynamic distribution of linker histone H1.5 in cellular differentiation. PLoS Genet. 2012, 8, e1002879. [Google Scholar] [CrossRef]
- Constantinescu, D.; Gray, H.L.; Sammak, P.J.; Schatten, G.P.; Csoka, A.B. Lamin A/C expression is a marker of mouse and human embryonic stem cell differentiation. Stem Cells 2006, 24, 177–185. [Google Scholar] [CrossRef]
- Röber, R.A.; Weber, K.; Osborn, M. Differential timing of nuclear lamin A/C expression in the various organs of the mouse embryo and the young animal: A developmental study. Development 1989, 105, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Belaadi, N.; Pernet, L.; Aureille, J.; Chadeuf, G.; Rio, M.; Vaillant, N.; Vitiello, E.; Lafanechère, L.; Loirand, G.; Guilluy, C. SUN2 regulates mitotic duration in response to extracellular matrix rigidity. Proc. Natl. Acad. Sci. USA 2022, 119, e2116167119. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Yan, Q.; Zhang, M.; Zhu, Y.; Liu, J.; Jiang, Y.; Zhen, X.; Xu, M.; Yue, Q.; Zhou, J.; et al. FHL1 mediates HOXA10 deacetylation via SIRT2 to enhance blastocyst-epithelial adhesion. Cell Death Discov. 2022, 8, 461. [Google Scholar] [CrossRef] [PubMed]
- Moreira, P.N.; Fernández-Gonzalez, R.; Ramirez, M.A.; Pérez-Crespo, M.; Rizos, D.; Pintado, B.; Gutiérrez-Adán, A. Differential effects of culture and nuclear transfer on relative transcript levels of genes with key roles during preimplantation. Zygote 2006, 14, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Zhu, R.; Dai, F.; Yang, D.; Liu, S.; Zheng, Y.; Wu, M.; Deng, Z.; Wang, Z.; Zhang, Y.; Tan, W.; et al. The mechanism of insulin-like growth factor II mRNA-binging protein 3 induce decidualization and maternal-fetal interface cross talk by TGF-β1 in recurrent spontaneous abortion. Front. Cell Dev. Biol. 2022, 10, 862180. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N 6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Xu, H.; Chen, B.; Wang, W.; Wang, L.; Sun, X.; Sang, Q. Expanding the genetic and phenotypic spectrum of female infertility caused by TLE6 mutations. J. Assist. Reprod. Genet. 2020, 37, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Bebbere, D.; Masala, L.; Albertini, D.F.; Ledda, S. The subcortical maternal complex: Multiple functions for one biological structure? J. Assist. Reprod. Genet. 2016, 33, 1431–1438. [Google Scholar] [CrossRef]
- Huang, X.; Sun, Q.; Chen, D.; Yang, W.; Zhang, J.; Liu, R.; Zhang, P.; Huang, L.; Zhang, M.; Fu, Q. Nlrp5 and Tle6 expression patterns in buffalo oocytes and pre-implantation embryos. Reprod. Domest. Anim. 2022, 57, 481–488. [Google Scholar] [CrossRef]
- Xie, H.; Zhang, B.; Xia, T.; Cui, J.; Pan, F.; Li, Y.; Fang, F.; Zhang, Y.; Ling, Y.; Liu, Y. Ezrin Thr567 phosphorylation participates in mouse oocyte maturation, fertilization, and early embryonic development. Fertil. Early Embryonic Dev. 2024. [Google Scholar] [CrossRef]
- Dard, N.; Louvet, S.; Santa-Maria, A.; Aghion, J.; Martin, M.; Mangeat, P.; Maro, B. In vivo functional analysis of ezrin during mouse blastocyst formation. Dev. Biol. 2001, 233, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Louvet, S.; Aghion, J.; Santa-Maria, A.; Mangeat, P.; Maro, B. Ezrin becomes restricted to outer cells following asymmetrical division in the preimplantation mouse embryo. Dev. Biol. 1996, 177, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Liu, X.; Wang, Z.; Qiao, F.; Wang, J.; Zhang, J.; Zhang, Y.; Qing, S.; Wang, Y. Effect of ezrin regulation by sperm-borne miR-183 on the formation of microvilli and early development of bovine embryos. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Bustin, M.; Reeves, R. High-mobility-group chromosomal proteins: Architectural components that facilitate chromatin function. Prog. Nucleic Acid. Res. Mol. Biol. 1996, 54, 35–100b. [Google Scholar] [PubMed]
- Matsubara, K.; Matsubara, Y.; Uchikura, Y.; Takagi, K.; Yano, A.; Sugiyama, T. HMGA1 is a potential driver of preeclampsia pathogenesis by interference with extravillous trophoblasts invasion. Biomolecules 2021, 11, 822. [Google Scholar] [CrossRef]
- Fusco, A.; Fedele, M. Roles of HMGA proteins in cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Kolundžić, N.; Bojić-Trbojević, Ž.; Kovačević, T.; Stefanoska, I.; Kadoya, T.; Vićovac, L. Galectin-1 is part of human trophoblast invasion machinery-a functional study in vitro. PLoS ONE 2011, 6, e28514. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, U.; Walzel, H.; Mylonas, I.; Papadopoulos, P.; Shabani, N.; Kuhn, C.; Schulze, S.; Friese, K.; Karsten, U.; Anz, D. The human endometrium expresses the glycoprotein mucin-1 and shows positive correlation for Thomsen-Friedenreich epitope expression and galectin-1 binding. J. Histochem. Cytochem. 2009, 57, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Petronczki, M.; Lénárt, P.; Peters, J.M. Polo on the rise—From mitotic entry to cytokinesis with Plk1. Dev. Cell 2008, 14, 646–659. [Google Scholar] [CrossRef]
- Barr, F.A.; Silljé, H.H.; Nigg, E.A. Polo-like kinases and the orchestration of cell division. Nat. Rev. Mol. Cell Biol. 2004, 5, 429–441. [Google Scholar] [CrossRef]
- Kumagai, A.; Dunphy, W.G. Purification and molecular cloning of Plx1, a Cdc25-regulatory kinase from Xenopus egg extracts. Science 1996, 273, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Eschrich, D.; Buchhaupt, M.; Kötter, P.; Entian, K.D. Nep1p (Emg1p), a novel protein conserved in eukaryotes and archaea, is involved in ribosome biogenesis. Curr. Genet. 2002, 40, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.L.; Moura, Y.B.; Viana, J.V.; de Oliveira, L.R.; Praxedes, É.A.; Vieira, J.B.; Sales, S.L.A.; Silva, H.V.R.; Luciano, M.C.D.S.; Pessoa, C.; et al. Full confluency, serum starvation, and roscovitine for inducing arrest in the G/G phase of the cell cycle in puma skin-derived fibroblast lines. Anim. Reprod. 2023, 20, e20230017. [Google Scholar] [CrossRef] [PubMed]
- Młodawska, W.; Mrowiec, P.; Bochenek, M.; Wnęk, K.; Kochan, J.; Nowak, A.; Niżański, W.; Prochowska, S.; Pałys, M. Effect of serum starvation and contact inhibition on dermal fibroblast cell cycle synchronization in two species of wild felids and domestic cat. Ann. Anim. Sci. 2022, 22, 1245–1255. [Google Scholar] [CrossRef]
- Veraguas, D.; Gallegos, P.F.; Castro, F.O.; Rodriguez-Alvarez, L. Cell cycle synchronization and analysis of apoptosis-related gene in skin fibroblasts from domestic cat (Felis silvestris catus) and kodkod (Leopardus guigna). Reprod. Domest. Anim. 2017, 52, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, I.; Watt, F.M. The role of keratins in modulating carcinogenesis via communication with cells of the immune system. Cell Stress 2019, 3, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.; Thompson, B.; Fisk, J.N.; Nebert, D.W.; Bruford, E.A.; Vasiliou, V.; Bunick, C.G. Update of the keratin gene family: Evolution, tissue-specific expression patterns, and relevance to clinical disorders. Hum. Genom. 2022, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Woodward, B.J.; Lenton, E.A.; Neil, S.M. Requirement of preimplantation human embryos for extracellular calmodulin for development. Hum. Reprod. 1993, 8, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Nasr-Esfahani, M.H.; Johnson, M.H. How does transferrin overcome the in vitro block to development of the mouse preimplantation embryo? Reproduction 1992, 96, 41–48. [Google Scholar] [CrossRef][Green Version]
- Sawada, H.; Yamazaki, K.; Hoshi, M. Trypsin-like hatching protease from mouse embryos: Evidence for the presence in culture medium and its enzymatic properties. J. Exp. Zool. 1990, 254, 83–87. [Google Scholar] [CrossRef]
- Perona, R.M.; Wassarman, P.M. Mouse blastocysts hatch in vitro by using a trypsin-like proteinase associated with cells of mural trophectoderm. Dev. Biol. 1986, 114, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Engel, W.; Franke, W.; Petzoldt, U. Isozymes as genetic markers in early mammalian development. In Isozymes V3: Developmental Biology; Academic Press: Cambridge, MA, USA, 2012; p. 67. [Google Scholar]
- Markova, M.D.; Nikolova, V.P.; Chakarova, I.V.; Zhivkova, R.S.; Dimitrov, R.K.; Delimitreva, S.M. Intermediate filament distribution patterns in maturing mouse oocytes and cumulus cells. Biocell 2015, 39, 1–7. [Google Scholar]
- South, A.P.; Wan, H.; Stone, M.G.; Dopping-Hepenstal, P.J.; Purkis, P.E.; Marshall, J.F.; Leigh, I.M.; Eady, R.A.; Hart, I.R.; McGrath, J.A. Lack of plakophilin 1 increases keratinocyte migration and reduces desmosome stability. J. Cell Sci. 2003, 116, 3303–3314. [Google Scholar] [CrossRef] [PubMed]
- Gallicano, G.I.; Kouklis, P.; Bauer, C.; Yin, M.; Vasioukhin, V.; Degenstein, L.; Fuchs, E. Desmoplakin is required early in development for assembly of desmosomes and cytoskeletal linkage. J. Cell Biol. 1998, 143, 2009–2022. [Google Scholar] [CrossRef] [PubMed]
- Veraguas, D.; Gallegos, P.F.; Velasquez, A.E.; Castro, F.O.; Rodriguez-Alvarez, L. FSH stimulation of anestrous cats improves oocyte quality and development of parthenogenetic embryos. Theriogenology 2017, 87, 25–35. [Google Scholar] [CrossRef]
- Veraguas, D.; Cuevas, S.R.; Gallegos, P.F.; Saez-Ruiz, D.; Castro, F.O.; Rodriguez-Alvarez, L. eCG stimulation in domestic cats increases the expression of gonadotrophin-induced genes improving oocyte competence during the non-breeding season. Reprod. Domest. Anim. 2018, 53, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Veraguas, D.; Saez, S.; Aguilera, C.; Echeverry, D.; Gallegos, P.F.; Saez-Ruiz, D.; Castro, F.O.; Rodriguez-Alvarez, L. In vitro and in vivo development of domestic cat embryos generated by in vitro fertilization after eCG priming and oocyte in vitro maturation. Theriogenology 2020, 146, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, A.E.; Castro, F.O.; Veraguas, D.; Cox, J.F.; Lara, E.; Briones, M.; Rodriguez-Alvarez, L. Splitting of IVP bovine blastocyst affects morphology and gene expression of resulting demi-embryos during in vitro culture and in vivo elongation. Zygote 2016, 24, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Vajta, G.; Korösi, T.; Du, Y.; Nakata, K.; Ieda, S.; Kuwayama, M.; Nagy, Z.P. The Well-of-the-Well system: An efficient approach to improve embryo development. Reprod. Biomed. Online 2008, 17, 73–81. [Google Scholar] [CrossRef]
- Kong, A.T.; Leprevost, F.V.; Avtonomov, D.M.; Mellacheruvu, D.; Nesvizhskii, A.I. MSFragger: Ultrafast and comprehensive peptide identification in mass spectrometry–based proteomics. Nat. Methods 2017, 14, 513–520. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | n* | Total Oocytes | Cleavages (%Mean ± SD) | Morulae (%Mean ± SD) | Blastocysts (%Mean ± SD) | Hatching Blastocysts (%Mean ± SD) |
|---|---|---|---|---|---|---|
| ZI | 10 | 351 | 140 (39.9 ± 11.6) | 71 (50.7 ± 14.3) | 44 (31.4 ± 8.5) | 8 (5.7 ± 10.6) |
| ZF | 10 | 470 | 201 (42.8 ± 15.8) | 95 (47.3 ± 27.9) | 62 (30.8 ± 17.8) |
| Protein ID | Log2FC | Name | Function | References |
|---|---|---|---|---|
| A0A2I2U1C5_FELCA | 3.98060042 | MYH9 | Fertilization, early development | [46,47] |
| M3W9X1_FELCA | 3.21413703 | PDIA4 | Morula to blastocyst transition | [45] |
| M3VZQ3_FELCA | 3.17734479 | TUFM | Mitochondrial translation | [48] |
| M3VYI7_FELCA | 2.71239201 | IGF2BP1 | m6A modification | [49] |
| A0A337S6R5_FELCA | 2.42297848 | ATP5PF | ATP production | [50] |
| A0A2I2UXE4_FELCA | 2.31927503 | HNRNPH1 | Post-transcriptional regulation | [51] |
| A0A5F5XWD4_FELCA | 2.23313382 | HSP90B1 | Stress and damage protection | [52] |
| A0A2I2U221_FELCA | 2.23313259 | CCT8 | Oocyte sperm interaction, cell growth | [53,54] |
| M3WRT2_FELCA | 2.0962291 | RPL8 | Ribosomal biogenesis | [55] |
| M3W9Y8_FELCA | 2.01100493 | RPL18 | Ribosomal biogenesis | [56] |
| A0A2I2UZM2_FELCA | 1.95964111 | HSPA8 | Cell differentiation, fertilization | [46,57,58] |
| A0A5F5Y0Y9_FELCA | 1.89221244 | EZR | Microvilli formation | [59] |
| M3WCX7_FELCA | 1.87232765 | EEF1B2 | Ribosomal translation factor | [60] |
| M3VX46_FELCA | 1.61502009 | COX5B | Mitochondrial function | [61] |
| A0A2I2U7Z5_FELCA | 1.60935135 | BCAP31 | Apoptosis, embryo implantation | [62] |
| EF1A1_FELCA | 1.51020066 | EEF1A1 | Ribosomal translation factor | [60] |
| A0A2I2UMD6_FELCA | 1.48842654 | HDLBP | Anti-inflammatory, antioxidant | [63] |
| A0A337SJJ9_FELCA | 1.42487437 | RNPS1 | Pre-mRNA splicing | [64] |
| M3WVC7_FELCA | 1.35282905 | YWHAZ | Cellular communication system | [65] |
| M3WFT4_FELCA | 1.34473579 | HSPA5 | Stress marker | [66] |
| A0A337S416_FELCA | 1.3425531 | ALDOA | Glycolysis in oocytes and embryos | [67] |
| M3WJL8_FELCA | 1.29586264 | TLE6 | Infertility, developmental arrest | [68,69,70] |
| Protein ID | Log2FC | Name | Functions | References |
|---|---|---|---|---|
| M3W9D5_FELCA | −1.20664542 | LGALS1 | Blastocyst attachment | [71] |
| M3WIA4_FELCA | −1.28571479 | COA3 | Mitochondrial respiratory chain | [72] |
| M3X4Y3_FELCA | −1.32858383 | UQCRFS1 | Mitochondrial structural | [73] |
| A0A2I2V068_FELCA | −1.35619626 | PSME3IP1 | Transmembrane protein | [74] |
| M3WD17_FELCA | −1.36580332 | RPL12 | Protein synthesis, hatching | [75] |
| M3WDF5_FELCA | −1.44449596 | TRA2B | mRNA splicing, endometrium | [76] |
| M3VW27_FELCA | −1.46532 | RPS25 | Repeated implantation failure | [77] |
| M3WAW0_FELCA | −1.48787899 | RRP9 | Morula to blastocyst transition | [78] |
| A0A2I2UL05_FELCA | −1.5655253 | PRXL2A | Antioxidant, embryo–maternal crosstalk | [44] |
| M3W1X7_FELCA | −1.60814943 | EMG1 | Preimplantation development | [79] |
| M3WHX6_FELCA | −1.74681032 | PLK1 | Mitosis, embryo development | [80] |
| A0A5F5XWX3_FELCA | −1.82092779 | KPNA2 | Preimplantation embryo arrest | [81] |
| A0A5F5XQQ6_FELCA | −1.9562502 | KHDC3L | Epiblast stability and viability | [82] |
| A0A337SCY3_FELCA | −1.9626275 | RPL36A | Protein synthesis in oocytes and embryos | [83,84] |
| A0A337SW00_FELCA | −2.0509994 | H2BC3 | DNA repair in embryos | [85] |
| A0A5F5XCJ6_FELCA | −2.11846902 | YIPF3 | Golgi transport, ER function | [86] |
| M3WCZ9_FELCA | −2.25459988 | H2BC18 | Immunity | [87] |
| A0A5F5XYM1_FELCA | −2.29930597 | COL1A2 | Embryo extracellular matrix | [88] |
| A0A2I2UXE2_FELCA | −2.65692623 | GNG12 | Inflammation, immunity | [89,90] |
| A0A337SVF6_FELCA | −2.81974135 | HMGA1 | Immunosuppression | [91] |
| Protein ID | Group | Name | Function | References |
|---|---|---|---|---|
| A0A337SNF5_FELCA | ZF | FTO | Epigenetic modulations | [92] |
| A0A337S839_FELCA | ZF | MDM4 | p53 regulation, cell proliferation | [93] |
| A0A337S8L1_FELCA | ZF | LDHA | Embryo metabolism | [94] |
| A0A337S103_FELCA | ZF | MT1X | Metal ion homeostasis, stress response | [95] |
| M3W8Q7_FELCA | ZF | GLUD1 | Glutaminolysis metabolism | [96,97] |
| A0A2I2USF3_FELCA | ZI | H2BC17 | Regulation of embryo development | [98] |
| A0A5F5XCD9_FELCA | ZI | KRT86 | Embryo implantation | [99] |
| A0A337SCD2_FELCA | ZI | ANXA2 | Cell adhesion, embryo implantation | [100] |
| A0A2I2UBN8_FELCA | ZI | JUP | Cell adhesion, embryo morphology | [101] |
| M3W114_FELCA | ZI | PLP2 | Gastrulation | [102] |
| A0A2I2UGZ3_FELCA | ZI | KRT32 | Cell adhesion, differentiation, and migration | [103] |
| A0A5F5XSP5_FELCA | ZI | LAP3 | Pregnancy biomarker | [104,105] |
| M3W2I5_FELCA | ZI | PKP1 | Desmosome stabilization and maturation | [106] |
| M3WCK3_FELCA | ZI | DSP | Desmosome formation, trophoblast stabilization | [107,108] |
| M3VUF5_FELCA | ZI | KRT82 | Intermediate filament formation | [109] |
| M3W584_FELCA | ZI | H1-5 | Differentiation of pluripotent cells | [110,111] |
| A0A5K1VP01_FELCA | ZI | LMNA | Cell plasticity and differentiation | [112,113] |
| A0A337SS59_FELCA | ZI | SUN2 | Mitosis, mitotic spindle formation | [114] |
| A0A2I2U7V8_FELCA | ZI | FHL1 | Blastocyst–epithelial adhesion | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veraguas-Dávila, D.; Zapata-Rojas, C.; Aguilera, C.; Saéz-Ruiz, D.; Saravia, F.; Castro, F.O.; Rodriguez-Alvarez, L. Proteomic Analysis of Domestic Cat Blastocysts and Their Secretome Produced in an In Vitro Culture System without the Presence of the Zona Pellucida. Int. J. Mol. Sci. 2024, 25, 4343. https://doi.org/10.3390/ijms25084343
Veraguas-Dávila D, Zapata-Rojas C, Aguilera C, Saéz-Ruiz D, Saravia F, Castro FO, Rodriguez-Alvarez L. Proteomic Analysis of Domestic Cat Blastocysts and Their Secretome Produced in an In Vitro Culture System without the Presence of the Zona Pellucida. International Journal of Molecular Sciences. 2024; 25(8):4343. https://doi.org/10.3390/ijms25084343
Chicago/Turabian StyleVeraguas-Dávila, Daniel, Camila Zapata-Rojas, Constanza Aguilera, Darling Saéz-Ruiz, Fernando Saravia, Fidel Ovidio Castro, and Lleretny Rodriguez-Alvarez. 2024. "Proteomic Analysis of Domestic Cat Blastocysts and Their Secretome Produced in an In Vitro Culture System without the Presence of the Zona Pellucida" International Journal of Molecular Sciences 25, no. 8: 4343. https://doi.org/10.3390/ijms25084343
APA StyleVeraguas-Dávila, D., Zapata-Rojas, C., Aguilera, C., Saéz-Ruiz, D., Saravia, F., Castro, F. O., & Rodriguez-Alvarez, L. (2024). Proteomic Analysis of Domestic Cat Blastocysts and Their Secretome Produced in an In Vitro Culture System without the Presence of the Zona Pellucida. International Journal of Molecular Sciences, 25(8), 4343. https://doi.org/10.3390/ijms25084343