
Effects of Liquid Fructose Supplementation and Chronic Unpredictable Stress on Uterine Contractile Activity in Nonpregnant Rats
 , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of Liquid Fructose Supplementation and/or Stress on Estrous Cycles Length and Regularity
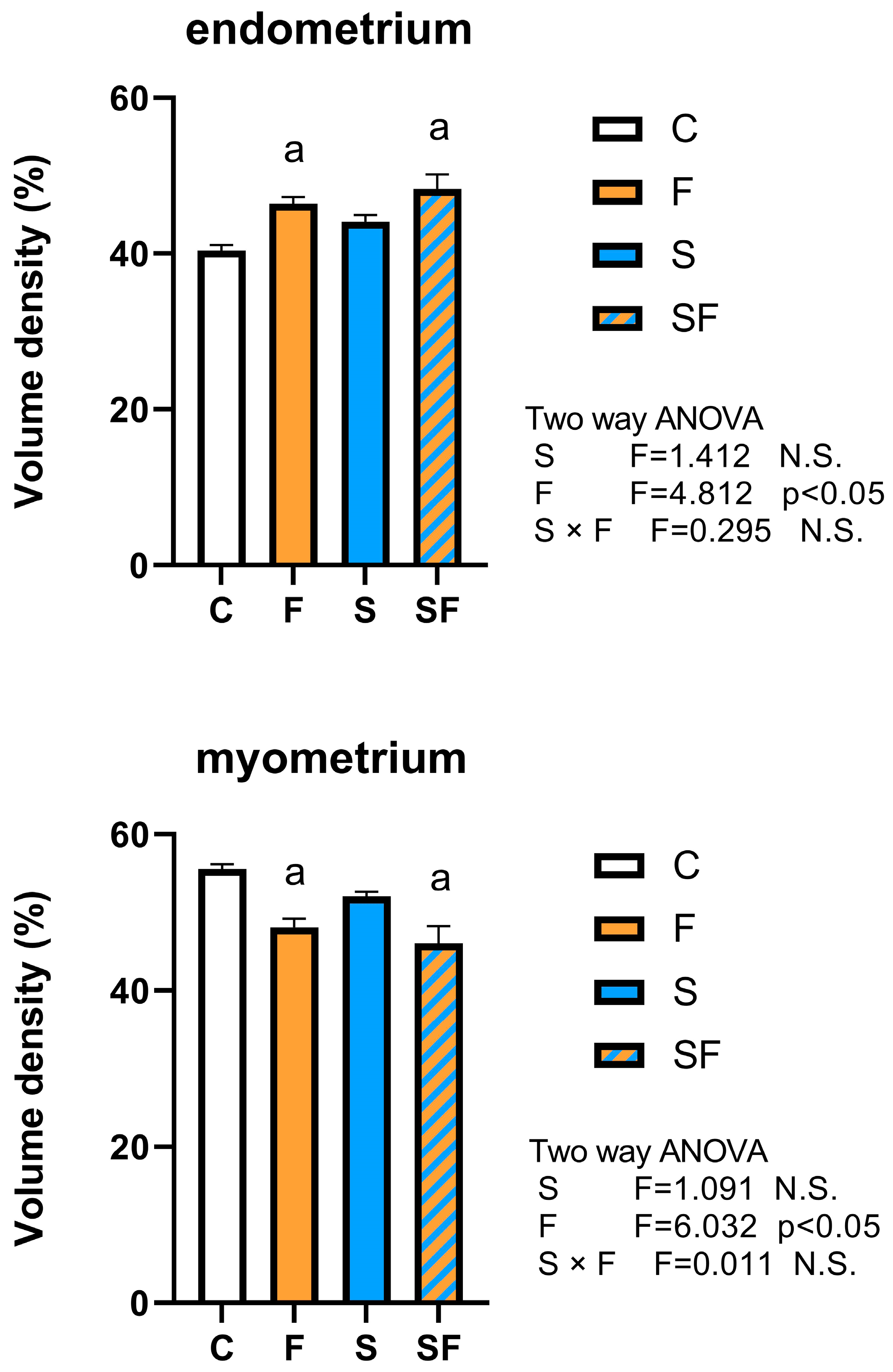
2.2. Histological and Stereological Analysis
2.3. Effects of Liquid Fructose Supplementation and/or Stress on Antioxidant Enzymes Activity
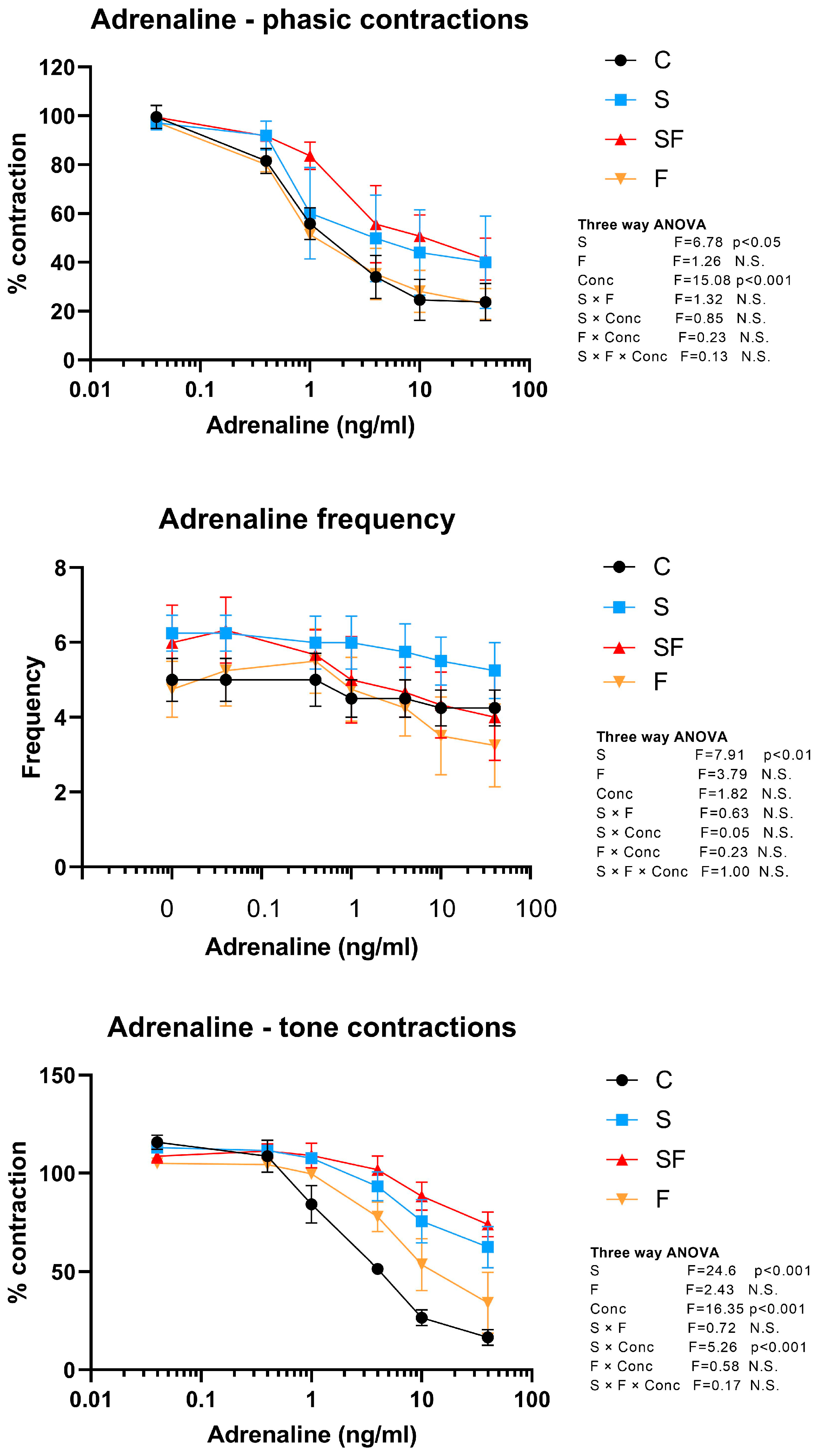
2.4. Effects of Liquid Fructose Supplementation and/or Stress on Uterine Contractility
2.4.1. Effects of Adrenaline on Phasic and Tone Uterine Contractions
2.4.2. Effects of Oxytocin on Phasic Uterine Contractions
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Tissue Preparation
4.3. Isolated Organ Bath Studies
4.4. Antioxidant Enzymes Activity
4.5. Histological and Stereological Analysis
4.6. Data Presentation and Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dallman, M.F.; Pecoraro, N.; Akana, S.F.; La Fleur, S.E.; Gomez, F.; Houshyar, H.; Bell, M.E.; Bhatnagar, S.; Laugero, K.D.; Manalo, S. Chronic stress and obesity: A new view of “comfort food”. Proc. Natl. Acad. Sci. USA 2003, 100, 11696–11701. [Google Scholar] [CrossRef]
- Tamashiro, K.L.; Sakai, R.R.; Shively, C.A.; Karatsoreos, I.N.; Reagan, L.P. Chronic stress, metabolism, and metabolic syndrome. Stress 2011, 14, 468–474. [Google Scholar] [CrossRef]
- Rosmond, R. Role of stress in the pathogenesis of the metabolic syndrome. Psychoneuroendocrinology 2005, 30, 1–10. [Google Scholar] [CrossRef]
- Taskinen, M.R.; Soderlund, S.; Bogl, L.H.; Hakkarainen, A.; Matikainen, N.; Pietilainen, K.H.; Rasanen, S.; Lundbom, N.; Bjornson, E.; Eliasson, B.; et al. Adverse effects of fructose on cardiometabolic risk factors and hepatic lipid metabolism in subjects with abdominal obesity. J. Intern. Med. 2017, 282, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Popkin, B.M.; Bray, G.A.; Despres, J.P.; Hu, F.B. Sugar-sweetened beverages, obesity, type 2 diabetes mellitus, and cardiovascular disease risk. Circulation 2010, 121, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Fontana, R.; Della Torre, S. The Deep Correlation between Energy Metabolism and Reproduction: A View on the Effects of Nutrition for Women Fertility. Nutrients 2016, 8, 87. [Google Scholar] [CrossRef]
- Sharma, R.; Biedenharn, K.R.; Fedor, J.M.; Agarwal, A. Lifestyle factors and reproductive health: Taking control of your fertility. Reprod. Biol. Endocrinol. 2013, 11, 66. [Google Scholar] [CrossRef]
- Broughton, D.E.; Moley, K.H. Obesity and female infertility: Potential mediators of obesity’s impact. Fertil. Steril. 2017, 107, 840–847. [Google Scholar] [CrossRef]
- Sanchez-Garrido, M.A.; Tena-Sempere, M. Metabolic dysfunction in polycystic ovary syndrome: Pathogenic role of androgen excess and potential therapeutic strategies. Mol. Metab. 2020, 35, 100937. [Google Scholar] [CrossRef] [PubMed]
- Chavarro, J.E.; Rich-Edwards, J.W.; Rosner, B.A.; Willett, W.C. A prospective study of dietary carbohydrate quantity and quality in relation to risk of ovulatory infertility. Eur. J. Clin. Nutr. 2009, 63, 78–86. [Google Scholar] [CrossRef]
- Ullah, R.; Su, Y.; Shen, Y.; Li, C.; Xu, X.; Zhang, J.; Huang, K.; Rauf, N.; He, Y.; Cheng, J.; et al. Postnatal feeding with high-fat diet induces obesity and precocious puberty in C57BL/6J mouse pups: A novel model of obesity and puberty. Front. Med. 2017, 11, 266–276. [Google Scholar] [CrossRef]
- Ullah, R.; Raza, A.; Rauf, N.; Shen, Y.; Zhou, Y.D.; Fu, J. Postnatal Feeding With a Fat Rich Diet Induces Precocious Puberty Independent of Body Weight, Body Fat, and Leptin Levels in Female Mice. Front. Endocrinol. 2019, 10, 758. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.A.; Kim, H.R.; Kim, Y.B.; Kim, H.S.; Lee, S.H. Effect of High Fructose Corn Syrup (HFCS) Intake on the Female Reproductive Organs and Lipid Accumulation in Adult Rats. Dev. Reprod. 2017, 21, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Valsamakis, G.; Chrousos, G.; Mastorakos, G. Stress, female reproduction and pregnancy. Psychoneuroendocrinology 2019, 100, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Vigil, P.; Melendez, J.; Soto, H.; Petkovic, G.; Bernal, Y.A.; Molina, S. Chronic Stress and Ovulatory Dysfunction: Implications in Times of COVID-19. Front. Glob. Women’s Health 2022, 3, 866104. [Google Scholar] [CrossRef]
- Dorfman, M.; Arancibia, S.; Fiedler, J.L.; Lara, H.E. Chronic intermittent cold stress activates ovarian sympathetic nerves and modifies ovarian follicular development in the rat. Biol. Reprod. 2003, 68, 2038–2043. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Wang, J.Z.; Li, J.J.; Wei, D.L.; Sui, H.S.; Zhang, Z.H.; Zhou, P.; Tan, J.H. Maternal restraint stress diminishes the developmental potential of oocytes. Biol. Reprod. 2011, 84, 672–681. [Google Scholar] [CrossRef]
- Zhao, X.; Ma, R.; Zhang, X.; Cheng, R.; Jiang, N.; Guo, M.; Rong, B.; Liu, Y.; Chen, M.; Feng, W.; et al. Reduced growth capacity of preimplantation mouse embryos in chronic unpredictable stress model. Mol. Reprod. Dev. 2021, 88, 80–95. [Google Scholar] [CrossRef]
- Wray, S.; Prendergast, C. The Myometrium: From Excitation to Contractions and Labour. Adv. Exp. Med. Biol. 2019, 1124, 233–263. [Google Scholar] [CrossRef]
- Malik, M.; Roh, M.; England, S.K. Uterine contractions in rodent models and humans. Acta Physiol. 2021, 231, e13607. [Google Scholar] [CrossRef]
- Warren, A.Y.; Matharoo-Ball, B.; Shaw, R.W.; Khan, R.N. Hydrogen peroxide and superoxide anion modulate pregnant human myometrial contractility. Reproduction 2005, 130, 539–544. [Google Scholar] [CrossRef]
- Appiah, I.; Milovanovic, S.; Radojicic, R.; Nikolic-Kokic, A.; Orescanin-Dusic, Z.; Slavic, M.; Trbojevic, S.; Skrbic, R.; Spasic, M.B.; Blagojevic, D. Hydrogen peroxide affects contractile activity and anti-oxidant enzymes in rat uterus. Br. J. Pharmacol. 2009, 158, 1932–1941. [Google Scholar] [CrossRef] [PubMed]
- Orescanin-Dusic, Z.; Tatalovic, N.; Vidonja-Uzelac, T.; Nestorov, J.; Nikolic-Kokic, A.; Mijuskovic, A.; Spasic, M.; Paskulin, R.; Bresjanac, M.; Blagojevic, D. The Effects of Ibogaine on Uterine Smooth Muscle Contractions: Relation to the Activity of Antioxidant Enzymes. Oxid. Med. Cell Longev. 2018, 2018, 5969486. [Google Scholar] [CrossRef]
- Carvajal, J.A.; Oporto, J.I. The Myometrium in Pregnant Women with Obesity. Curr. Vasc. Pharmacol. 2021, 19, 193–200. [Google Scholar] [CrossRef]
- Wray, S. Insights into the uterus. Exp. Physiol. 2007, 92, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Wray, S.; Arrowsmith, S. Uterine Excitability and Ion Channels and Their Changes with Gestation and Hormonal Environment. Annu. Rev. Physiol. 2021, 83, 331–357. [Google Scholar] [CrossRef] [PubMed]
- Hjollund, N.H.; Jensen, T.K.; Bonde, J.P.; Henriksen, T.B.; Andersson, A.M.; Kolstad, H.A.; Ernst, E.; Giwercman, A.; Skakkebaek, N.E.; Olsen, J. Distress and reduced fertility: A follow-up study of first-pregnancy planners. Fertil. Steril. 1999, 72, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.Y.; Chen, H.H.; Zhang, N.; Ding, M.X.; Qiu, Y.E.; Pan, X.M.; Fang, Y.S.; Lin, Y.P.; Zheng, Q.; Wang, W.Q. Effects of chronic unpredictable mild stress on ovarian reserve in female rats: Feasibility analysis of a rat model of premature ovarian failure. Mol. Med. Rep. 2018, 18, 532–540. [Google Scholar] [CrossRef]
- Light, H.R.; Tsanzi, E.; Gigliotti, J.; Morgan, K.; Tou, J.C. The type of caloric sweetener added to water influences weight gain, fat mass, and reproduction in growing Sprague-Dawley female rats. Exp. Biol. Med. 2009, 234, 651–661. [Google Scholar] [CrossRef]
- Kovacevic, S.; Nestorov, J.; Matic, G.; Elakovic, I. Fructose and stress induce opposite effects on lipid metabolism in the visceral adipose tissue of adult female rats through glucocorticoid action. Eur. J. Nutr. 2017, 56, 2115–2128. [Google Scholar] [CrossRef]
- Kunz, G.; Noe, M.; Herbertz, M.; Leyendecker, G. Uterine peristalsis during the follicular phase of the menstrual cycle: Effects of oestrogen, antioestrogen and oxytocin. Hum. Reprod. Update 1998, 4, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Wray, S.; Noble, K. Sex hormones and excitation-contraction coupling in the uterus: The effects of oestrous and hormones. J. Neuroendocrinol. 2008, 20, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Kunz, G.; Herbertz, M.; Noe, M.; Leyendecker, G. Sonographic evidence for the involvement of the utero-ovarian counter-current system in the ovarian control of directed uterine sperm transport. Hum. Reprod. Update 1998, 4, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Kunz, G.; Leyendecker, G. Uterine peristaltic activity during the menstrual cycle: Characterization, regulation, function and dysfunction. Reprod. Biomed. Online 2002, 4 (Suppl. S3), 5–9. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Kendrick, A.; Quenby, S.; Wray, S. Contractility and calcium signaling of human myometrium are profoundly affected by cholesterol manipulation: Implications for labor? Reprod. Sci. 2007, 14, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, S.; Elakovic, I.; Vojnovic Milutinovic, D.; Nikolic-Kokic, A.; Blagojevic, D.; Matic, G.; Tappy, L.; Djordjevic, A.; Brkljacic, J. Fructose-Rich Diet Attenuates Stress-Induced Metabolic Disturbances in the Liver of Adult Female Rats. J. Nutr. 2021, 151, 3661–3670. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, I.H.; Harris, J.S. The effect of Adrenalin on the pregnant human uterus. Am. J. Obstet. Gynecol. 1950, 59, 775–784. [Google Scholar] [CrossRef]
- Young, R.C. Myocytes, myometrium, and uterine contractions. Ann. N. Y. Acad. Sci. 2007, 1101, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lopez, A.L.; Jaime, H.B.; Escobar Villanueva, M.D.C.; Padilla, M.B.; Palacios, G.V.; Aguilar, F.J.A. Chronic unpredictable mild stress generates oxidative stress and systemic inflammation in rats. Physiol. Behav. 2016, 161, 15–23. [Google Scholar] [CrossRef]
- Sahin, E.; Gumuslu, S. Alterations in brain antioxidant status, protein oxidation and lipid peroxidation in response to different stress models. Behav. Brain Res. 2004, 155, 241–248. [Google Scholar] [CrossRef]
- Sahin, E.; Gumuslu, S. Immobilization stress in rat tissues: Alterations in protein oxidation, lipid peroxidation and antioxidant defense system. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 144, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Gumuslu, S. Stress-dependent induction of protein oxidation, lipid peroxidation and anti-oxidants in peripheral tissues of rats: Comparison of three stress models (immobilization, cold and immobilization-cold). Clin. Exp. Pharmacol. Physiol. 2007, 34, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, J.; Djordjevic, A.; Adzic, M.; Niciforovic, A.; Radojcic, M.B. Chronic stress differentially affects antioxidant enzymes and modifies the acute stress response in liver of Wistar rats. Physiol. Res. 2010, 59, 729–736. [Google Scholar] [CrossRef]
- Djordjevic, J.; Djordjevic, A.; Adzic, M.; Radojcic, M.B. Chronic social isolation compromises the activity of both glutathione peroxidase and catalase in hippocampus of male wistar rats. Cell Mol. Neurobiol. 2010, 30, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Che, Y.; Zhou, Z.; Shu, Y.; Zhai, C.; Zhu, Y.; Gong, S.; Cui, Y.; Wang, J.F. Chronic unpredictable stress impairs endogenous antioxidant defense in rat brain. Neurosci. Lett. 2015, 584, 208–213. [Google Scholar] [CrossRef]
- Kovacevic, S.; Nestorov, J.; Matic, G.; Elakovic, I. Fructose-enriched diet induces inflammation and reduces antioxidative defense in visceral adipose tissue of young female rats. Eur. J. Nutr. 2017, 56, 151–160. [Google Scholar] [CrossRef]
- Lopes, A.; Vilela, T.C.; Taschetto, L.; Vuolo, F.; Petronilho, F.; Dal-Pizzol, F.; Streck, E.L.; Ferreira, G.C.; Schuck, P.F. Evaluation of the effects of fructose on oxidative stress and inflammatory parameters in rat brain. Mol. Neurobiol. 2014, 50, 1124–1130. [Google Scholar] [CrossRef]
- Seraphim, D.C.C.; Punaro, G.R.; Fernandes, T.O.; Ginoza, M.; Lopes, G.S.; Higa, E.M.S. Assessment of fructose overload in the metabolic profile and oxidative/nitrosative stress in the kidney of senescent female rats. Exp. Gerontol. 2017, 99, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, J.; Dudzinska, W.; Skotnicka, E.; Sielatycka, K.; Daniel, I. The Impact of a Diet Containing Sucrose and Systematically Repeated Starvation on the Oxidative Status of the Uterus and Ovary of Rats. Nutrients 2019, 11, 1544. [Google Scholar] [CrossRef]
- Spasojevic, I.; Bajic, A.; Jovanovic, K.; Spasic, M.; Andjus, P. Protective role of fructose in the metabolism of astroglial C6 cells exposed to hydrogen peroxide. Carbohydr. Res. 2009, 344, 1676–1681. [Google Scholar] [CrossRef]
- Spasojevic, I.; Mojovic, M.; Blagojevic, D.; Spasic, S.D.; Jones, D.R.; Nikolic-Kokic, A.; Spasic, M.B. Relevance of the capacity of phosphorylated fructose to scavenge the hydroxyl radical. Carbohydr. Res. 2009, 344, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Semchyshyn, H.M. Fructation in vivo: Detrimental and protective effects of fructose. Biomed. Res. Int. 2013, 2013, 343914. [Google Scholar] [CrossRef] [PubMed]
- Ventura, E.E.; Davis, J.N.; Goran, M.I. Sugar content of popular sweetened beverages based on objective laboratory analysis: Focus on fructose content. Obesity 2011, 19, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Joels, M.; Karst, H.; Alfarez, D.; Heine, V.M.; Qin, Y.; van Riel, E.; Verkuyl, M.; Lucassen, P.J.; Krugers, H.J. Effects of chronic stress on structure and cell function in rat hippocampus and hypothalamus. Stress 2004, 7, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Marcondes, F.K.; Bianchi, F.J.; Tanno, A.P. Determination of the estrous cycle phases of rats: Some helpful considerations. Braz. J. Biol. 2002, 62, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Mijuskovic, A.; Kokic, A.N.; Dusic, Z.O.; Slavic, M.; Spasic, M.B.; Blagojevic, D. Chloride channels mediate sodium sulphide-induced relaxation in rat uteri. Br. J. Pharmacol. 2015, 172, 3671–3686. [Google Scholar] [CrossRef] [PubMed]
- Ristic, N.; Nestorovic, N.; Manojlovic-Stojanoski, M.; Filipovic, B.; Sosic-Jurjevic, B.; Milosevic, V.; Sekulic, M. Maternal dexamethasone treatment reduces ovarian follicle number in neonatal rat offspring. J. Microsc. 2008, 232, 549–557. [Google Scholar] [CrossRef]
- Gundersen, H.J.; Jensen, E.B. The efficiency of systematic sampling in stereology and its prediction. J. Microsc. 1987, 147, 229–263. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Fructose | Stress | Stress + Fructose | |
|---|---|---|---|---|
| Duration of the estrous cycle (days) | 5 ± 1.7 | 5 ± 2.2 | 5 ± 0.7 | * n.d. (prolonged diestrus) |
| Females with irregular cycles | 0/6 | 1/6 | 1/6 | 5/6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oreščanin Dušić, Z.; Kovačević, S.; Ristić, N.; Vojnović Milutinović, D.; Vidonja Uzelac, T.; Blagojević, D.; Djordjevic, A.; Brkljačić, J. Effects of Liquid Fructose Supplementation and Chronic Unpredictable Stress on Uterine Contractile Activity in Nonpregnant Rats. Int. J. Mol. Sci. 2024, 25, 6770. https://doi.org/10.3390/ijms25126770
Oreščanin Dušić Z, Kovačević S, Ristić N, Vojnović Milutinović D, Vidonja Uzelac T, Blagojević D, Djordjevic A, Brkljačić J. Effects of Liquid Fructose Supplementation and Chronic Unpredictable Stress on Uterine Contractile Activity in Nonpregnant Rats. International Journal of Molecular Sciences. 2024; 25(12):6770. https://doi.org/10.3390/ijms25126770
Chicago/Turabian StyleOreščanin Dušić, Zorana, Sanja Kovačević, Nataša Ristić, Danijela Vojnović Milutinović, Teodora Vidonja Uzelac, Duško Blagojević, Ana Djordjevic, and Jelena Brkljačić. 2024. "Effects of Liquid Fructose Supplementation and Chronic Unpredictable Stress on Uterine Contractile Activity in Nonpregnant Rats" International Journal of Molecular Sciences 25, no. 12: 6770. https://doi.org/10.3390/ijms25126770