
Sodium Acetate Enhances Neutrophil Extracellular Trap Formation via Histone Acetylation Pathway in Neutrophil-like HL-60 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Sodium Acetate Treatment Increased NOX-Independent NETosis in nHL-60 Cells
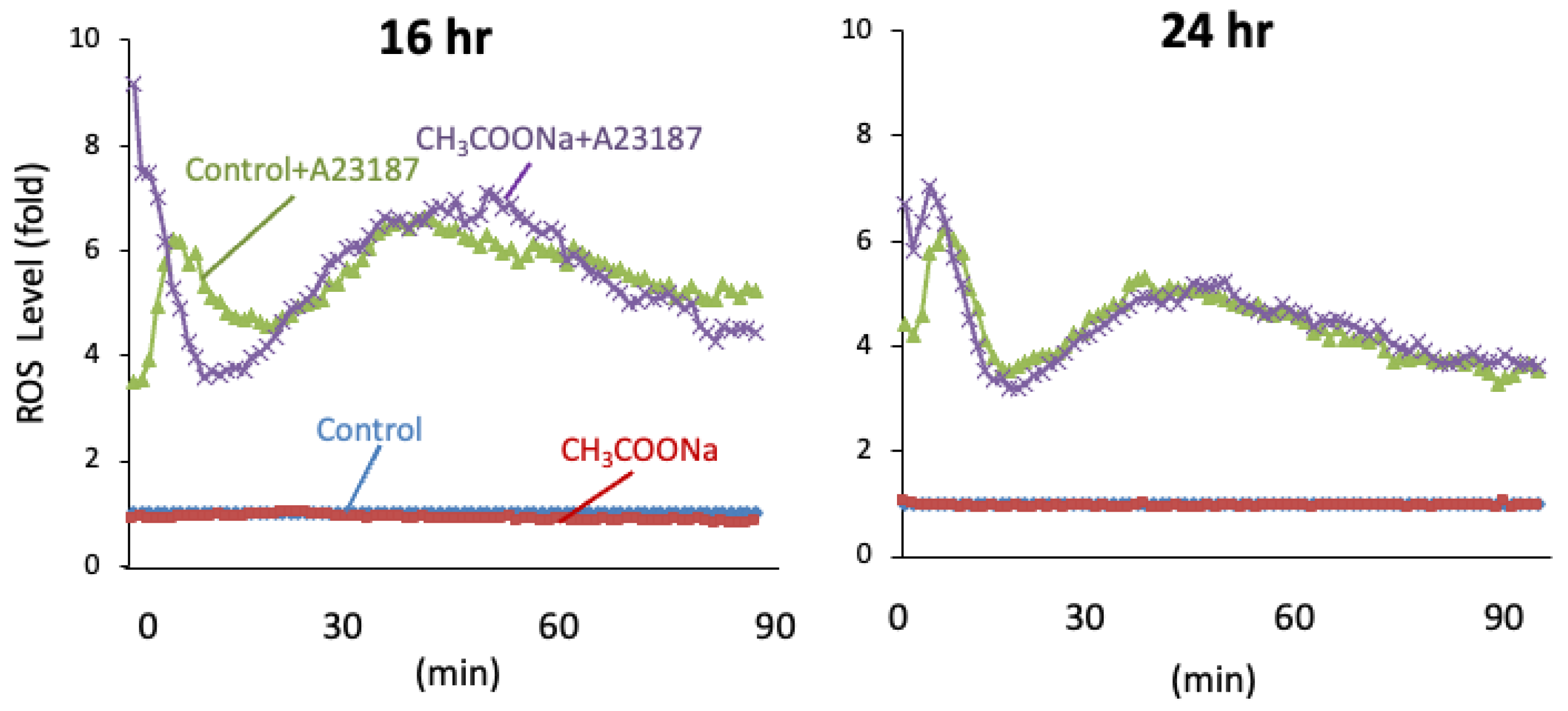
2.2. Sodium Acetate Treatment Did Not Increase ROS Production in nHL-60 Cells
2.3. Effect of ACSS2 (Acetyl-CoA Synthetase 2) Inhibitor on Extracellular DNA Release
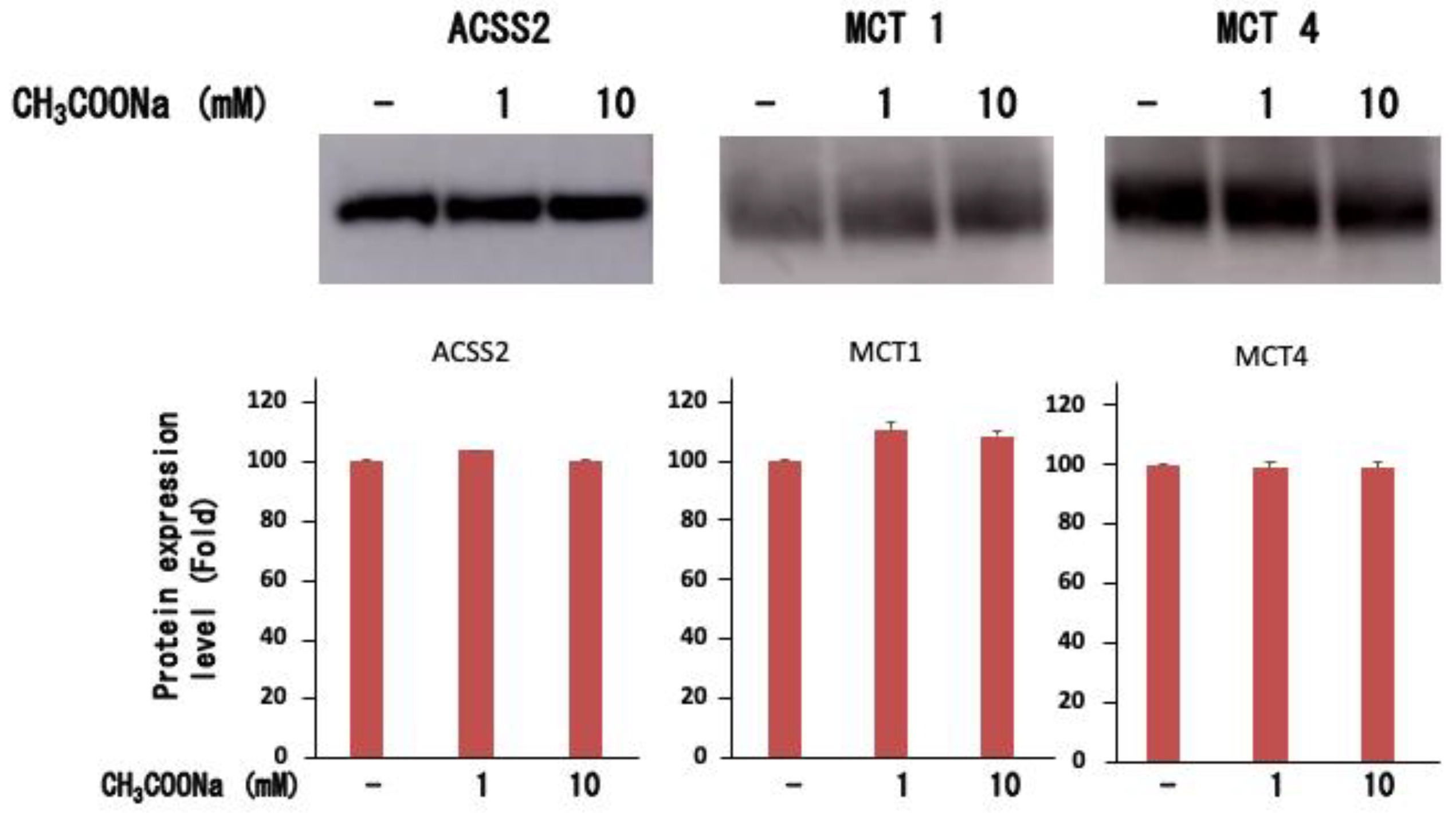
2.4. Expression of ACSS2, MCT-1, and MCT-4
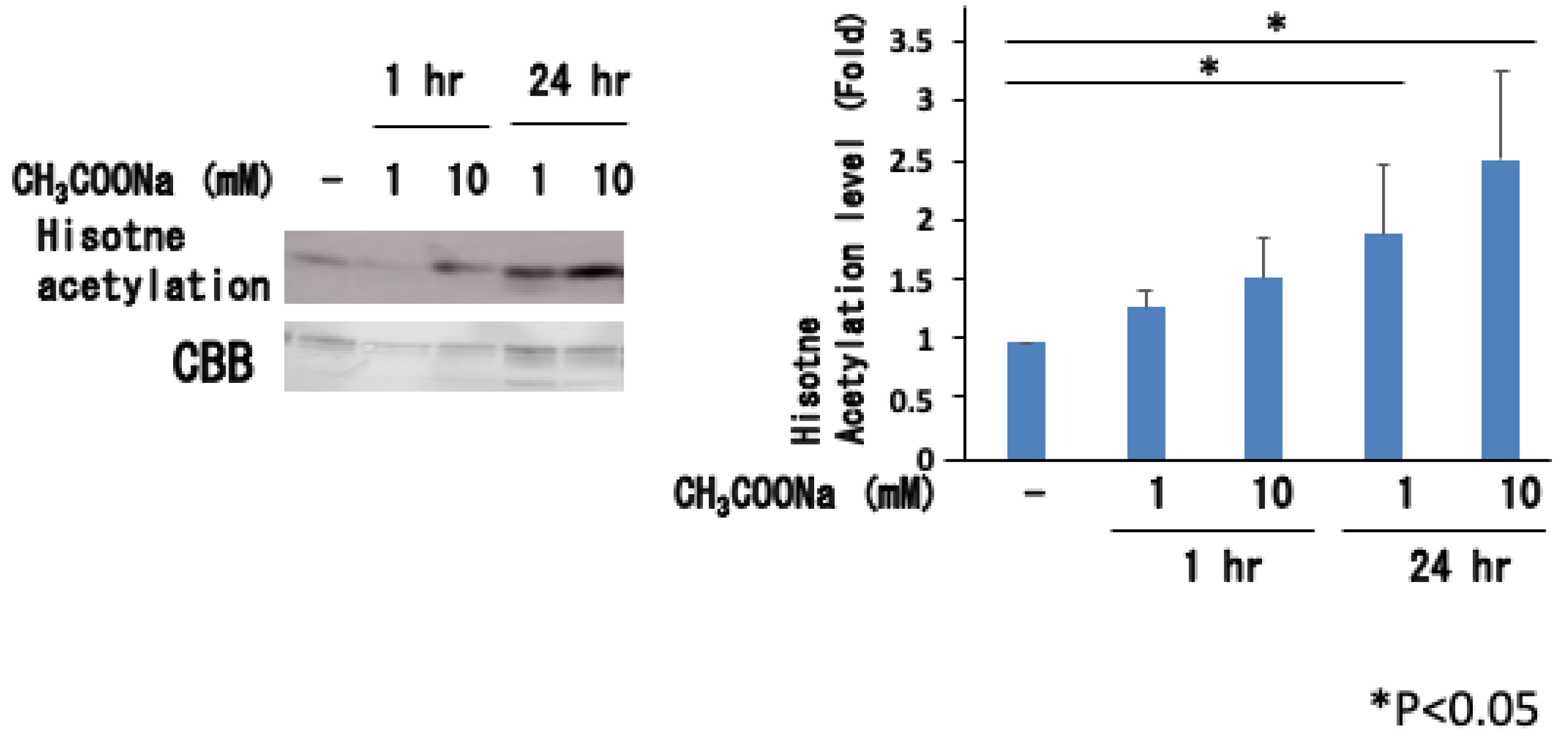
2.5. Sodium Acetate Treatment Increased Histone Acetylation
2.6. Histone Acetylation Decreased after Stimulation of A23187
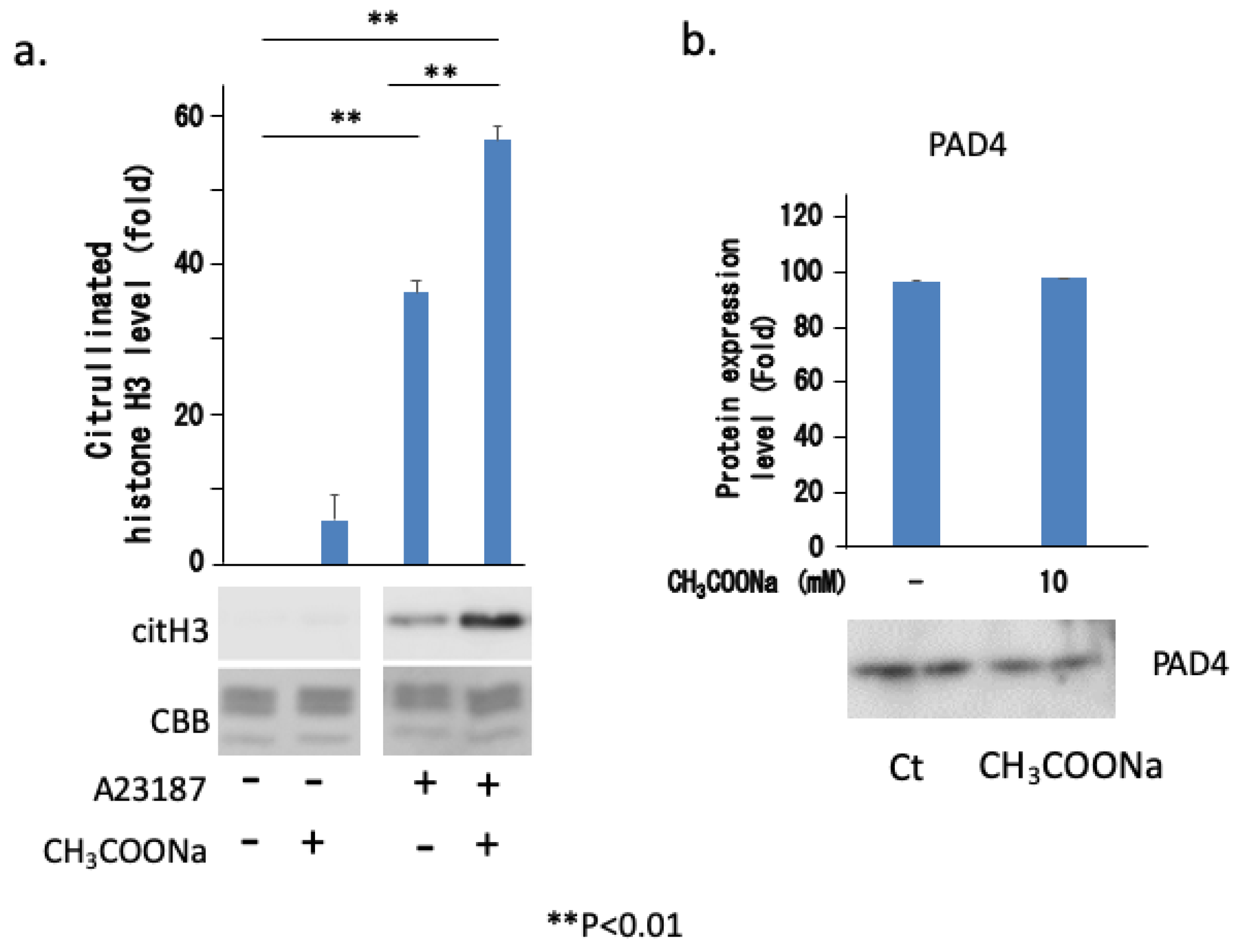
2.7. Sodium Acetate Treatment Increased NETosis by Increasing Histone Citrullination
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Sodium Acetate
4.3. SYTOX Green NETosis Assay
4.4. NET Visualization
4.5. ROS Measurement
4.6. Quantification of Extracellular DNA
4.7. Total Cell Protein Extraction
4.8. Histone Extraction
4.9. Western Blotting
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, N.; Radic, M. Citrullination of autoantigens implicates NETosis in the induction of autoimmunity. Ann. Rheum. Dis. 2014, 73, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Knackstedt, S.L.; Georgiadou, A.; Apel, F.; Abu-Abed, U.; Moxon, C.A.; Cunnington, A.J.; Raupach, B.; Cunningham, D.; Langhorne, J.; Kruger, R.; et al. Neutrophil extracellular traps drive inflammatory pathogenesis in malaria. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Apel, F.; Zychlinsky, A.; Kenny, E.F. The role of neutrophil extracellular traps in rheumatic diseases. Nat. Rev. Rheumatol. 2018, 14, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Krumbholz, M.; Schonermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Grone, H.J.; Brinkmann, V.; Jenne, D.E. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H.; Sonoda, A.; Yamamoto, M.; Kawashima, Y.; Takishita, Y.; Morita, A.; Tsutsumi, T.; Tsuchiya, M.; Sato, E.F. 17-β-estradiol enhances neutrophil extracellular trap formation by interaction with estrogen membrane receptor. Arch. Biochem. Biophys. 2019, 663, 64–70. [Google Scholar] [CrossRef]
- Portier, I.; Campbell, R.A.; Denorme, F. Mechanisms of immunothrombosis in COVID-19. Curr. Opin. Hematol. 2021, 28, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Anders, H.J.; Bilyy, R.; Bowlin, G.L.; Daniel, C.; De Lorenzo, R.; Egeblad, M.; Henneck, T.; Hidalgo, A.; Hoffmann, M.; et al. Patients with COVID-19: In the dark-NETs of neutrophils. Cell Death Differ. 2021, 28, 3125–3139. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.E.; McGuone, D.; Xu, M.L.; Jane-Wit, D.; Mitchell, R.N.; Libby, P.; Pober, J.S. Coronavirus Disease 2019 (COVID-19) Coronary Vascular Thrombosis: Correlation with Neutrophil but Not Endothelial Activation. Am. J. Pathol. 2022, 192, 112–120. [Google Scholar] [CrossRef]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X.; Huang, L.; Hoffmann, D.; Lu, M.; Qiu, Y. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11, 2063. [Google Scholar]
- Douda, D.N.; Khan, M.A.; Grasemann, H.; Palaniyar, N. SK3 channel and mitochondrial ROS mediate NADPH oxidase-independent NETosis induced by calcium influx. Proc. Natl. Acad. Sci. USA 2015, 112, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Takishita, Y.; Yasuda, H.; Shimizu, M.; Matsuo, A.; Morita, A.; Tsutsumi, T.; Tsuchiya, M.; Sato, E.F. Formation of neutrophil extracellular traps in mitochondrial DNA-deficient cells. J. Clin. Biochem. Nutr. 2020, 66, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Farahvash, A.; Douda, D.N.; Licht, J.C.; Grasemann, H.; Sweezey, N.; Palaniyar, N. JNK Activation Turns on LPS-and Gram-Negative Bacteria-Induced NADPH Oxidase-Dependent Suicidal NETosis. Sci. Rep. 2017, 7, 3409. [Google Scholar] [CrossRef]
- Khan, M.A.; Palaniyar, N. Transcriptional firing helps to drive NETosis. Sci. Rep. 2017, 7, 41749. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef]
- Kirchner, T.; Moller, S.; Klinger, M.; Solbach, W.; Laskay, T.; Behnen, M. The impact of various reactive oxygen species on the formation of neutrophil extracellular traps. Mediat. Inflamm. 2012, 2012, 849136. [Google Scholar] [CrossRef] [PubMed]
- Al-Khafaji, A.B.; Tohme, S.; Yazdani, H.O.; Miller, D.; Huang, H.; Tsung, A. Superoxide induces Neutrophil Extracellular Trap Formation in a TLR-4 and NOX-dependent mechanism. Mol. Med. 2016, 22, 621–631. [Google Scholar] [CrossRef]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef]
- Boffa, L.C.; Vidali, G.; Mann, R.S.; Allfrey, V.G. Suppression of histone deacetylation in vivo and in vitro by sodium butyrate. J. Biol. Chem. 1978, 253, 3364–3366. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Nakao, S.; Moriya, Y.; Furuyama, S.; Niederman, R.; Sugiya, H. Propionic acid stimulates superoxide generation in human neutrophils. Cell Biol. Int. 1998, 22, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Schlatterer, K.; Beck, C.; Schoppmeier, U.; Peschel, A.; Kretschmer, D. Acetate sensing by GPR43 alarms neutrophils and protects from severe sepsis. Commun. Biol. 2021, 4, 928. [Google Scholar] [CrossRef] [PubMed]
- Iniguez-Gutierrez, L.; Godinez-Mendez, L.A.; Fafutis-Morris, M.; Padilla-Arellano, J.R.; Corona-Rivera, A.; Bueno-Topete, M.R.; Rojas-Rejon, O.A.; Delgado-Rizo, V. Physiological concentrations of short-chain fatty acids induce the formation of neutrophil extracellular traps in vitro. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420958949. [Google Scholar] [CrossRef] [PubMed]
- Ohbuchi, A.; Kono, M.; Takenokuchi, M.; Imoto, S.; Saigo, K. Acetate moderately attenuates the generation of neutrophil extracellular traps. Blood Res. 2018, 53, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xu, H.; Du, X.; Lit, L.; Walker, W.; Lu, A.; Ran, R.; Gregg, J.P.; Reilly, M.; Pancioli, A.; et al. Gene expression in blood changes rapidly in neutrophils and monocytes after ischemic stroke in humans: A microarray study. J. Cereb. Blood Flow. Metab. 2006, 26, 1089–1102. [Google Scholar] [CrossRef]
- Mendoza, M.; Egervari, G.; Sidoli, S.; Donahue, G.; Alexander, D.C.; Sen, P.; Garcia, B.A.; Berger, S.L. Enzymatic transfer of acetate on histones from lysine reservoir sites to lysine activating sites. Sci. Adv. 2022, 8, eabj5688. [Google Scholar] [CrossRef]
- Hamam, H.J.; Khan, M.A.; Palaniyar, N. Histone Acetylation Promotes Neutrophil Extracellular Trap Formation. Biomolecules 2019, 9, 32. [Google Scholar] [CrossRef]
- Hamam, H.J.; Palaniyar, N. Histone Deacetylase Inhibitors Dose-Dependently Switch Neutrophil Death from NETosis to Apoptosis. Biomolecules 2019, 9, 184. [Google Scholar] [CrossRef]
- Conejeros, I.; Lopez-Osorio, S.; Zhou, E.; Velasquez, Z.D.; Del Rio, M.C.; Burgos, R.A.; Alarcon, P.; Chaparro-Gutierrez, J.J.; Hermosilla, C.; Taubert, A. Glycolysis, monocarboxylate transport, and purinergic signaling are key events in Eimeria bovis-induced NETosis. Front. Immunol. 2022, 13, 842482. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kabe, Y.; Kanai, A.; Sugiura, Y.; Hida, S.; Taniguchi, S.; Takahashi, T.; Matsui, H.; Yasukawa, Z.; Itou, H.; et al. Short-chain fatty acids bind to apoptosis-associated speck-like protein to activate inflammasome complex to prevent Salmonella infection. PLoS Biol. 2020, 18, e3000813. [Google Scholar] [CrossRef] [PubMed]
- Munzer, P.; Negro, R.; Fukui, S.; di Meglio, L.; Aymonnier, K.; Chu, L.; Cherpokova, D.; Gutch, S.; Sorvillo, N.; Shi, L.; et al. NLRP3 Inflammasome Assembly in Neutrophils Is Supported by PAD4 and Promotes NETosis Under Sterile Conditions. Front. Immunol. 2021, 12, 683803. [Google Scholar] [CrossRef] [PubMed]
- Lien, T.S.; Sun, D.S.; Hung, S.C.; Wu, W.S.; Chang, H.H. Dengue Virus Envelope Protein Domain III Induces Nlrp3 Inflammasome-Dependent NETosis-Mediated Inflammation in Mice. Front. Immunol. 2021, 12, 618577. [Google Scholar] [CrossRef] [PubMed]
- Westerterp, M.; Fotakis, P.; Ouimet, M.; Bochem, A.E.; Zhang, H.; Molusky, M.M.; Wang, W.; Abramowicz, S.; la Bastide-van Gemert, S.; Wang, N.; et al. Cholesterol Efflux Pathways Suppress Inflammasome Activation, NETosis, and Atherogenesis. Circulation 2018, 138, 898–912. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.P.; Denu, J.M. Short-chain fatty acids activate acetyltransferase p300. eLife 2021, 10, e72171. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, J.F.; Hany, L.; Barat, C.; Ouellet, M.; Tremblay, M.J. Epigenetic Metabolite Acetate Inhibits Class I/II Histone Deacetylases, Promotes Histone Acetylation, and Increases HIV-1 Integration in CD4+ T Cells. J. Virol. 2017, 91, e01943-16. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, D.; Yao, H.; Doret, P.; Hao, G.; Shen, Q.; Qiu, H.; Zhang, X.; Wang, Y.; Chen, G.; et al. Coordination of PAD4 and HDAC2 in the regulation of p53-target gene expression. Oncogene 2010, 29, 3153–3162. [Google Scholar] [CrossRef]
- Saiki, M.; Watase, M.; Matsubayashi, H.; Hidaka, Y. Recognition of the N-terminal histone H2A and H3 peptides by peptidylarginine deiminase IV. Protein Pept. Lett. 2009, 16, 1012–1016. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef]
- Nuutinen, H.; Lindros, K.; Hekali, P.; Salaspuro, M. Elevated blood acetate as indicator of fast ethanol elimination in chronic alcoholics. Alcohol 1985, 2, 623–626. [Google Scholar] [CrossRef]
- Tang, C.; Offermanns, S. FFA2 and FFA3 in Metabolic Regulation. In Free Fatty Acid Receptors; Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2017; Volume 236, pp. 205–220. [Google Scholar]
- Collins, S.J.; Ruscetti, F.W.; Gallagher, R.E.; Gallo, R.C. Terminal differentiation of human promyelocytic leukemia cells induced by dimethyl sulfoxide and other polar compounds. Proc. Natl. Acad. Sci. USA 1978, 75, 2458–2462. [Google Scholar] [CrossRef] [PubMed]
- Bunce, C.M.; Fisher, A.G.; Toksoz, D.; Brown, G. Isolation and characterisation of dimethylsulphoxide resistant variants from the human promyeloid cell line HL60. Exp. Hematol. 1983, 11, 828–833. [Google Scholar] [PubMed]
- Yasuda, H.; Ohashi, A.; Nishida, S.; Kamiya, T.; Suwa, T.; Hara, H.; Takeda, J.; Itoh, Y.; Adachi, T. Exendin-4 induces extracellular-superoxide dismutase through histone H3 acetylation in human retinal endothelial cells. J. Clin. Biochem. Nutr. 2016, 59, 174–181. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasuda, H.; Takishita, Y.; Morita, A.; Tsutsumi, T.; Nakagawa, N.; Sato, E.F. Sodium Acetate Enhances Neutrophil Extracellular Trap Formation via Histone Acetylation Pathway in Neutrophil-like HL-60 Cells. Int. J. Mol. Sci. 2024, 25, 8757. https://doi.org/10.3390/ijms25168757
Yasuda H, Takishita Y, Morita A, Tsutsumi T, Nakagawa N, Sato EF. Sodium Acetate Enhances Neutrophil Extracellular Trap Formation via Histone Acetylation Pathway in Neutrophil-like HL-60 Cells. International Journal of Molecular Sciences. 2024; 25(16):8757. https://doi.org/10.3390/ijms25168757
Chicago/Turabian StyleYasuda, Hiroyuki, Yutaka Takishita, Akihiro Morita, Tomonari Tsutsumi, Naoya Nakagawa, and Eisuke F. Sato. 2024. "Sodium Acetate Enhances Neutrophil Extracellular Trap Formation via Histone Acetylation Pathway in Neutrophil-like HL-60 Cells" International Journal of Molecular Sciences 25, no. 16: 8757. https://doi.org/10.3390/ijms25168757
APA StyleYasuda, H., Takishita, Y., Morita, A., Tsutsumi, T., Nakagawa, N., & Sato, E. F. (2024). Sodium Acetate Enhances Neutrophil Extracellular Trap Formation via Histone Acetylation Pathway in Neutrophil-like HL-60 Cells. International Journal of Molecular Sciences, 25(16), 8757. https://doi.org/10.3390/ijms25168757