Abstract
Lipids are organic nonpolar molecules with essential biological and economic importance. While the genetic pathways and regulatory networks of lipid biosynthesis and metabolism have been extensively studied and thoroughly reviewed in oil crops such as soybeans, less attention has been paid to the biological roles of lipids in rice, a staple food for the global population and a model species for plant molecular biology research, leaving a considerable knowledge gap in the biological roles of lipids. In this review, we endeavor to furnish a current overview of the advancements in understanding the genetic foundations and physiological functions of lipids, including triacylglycerol, fatty acids, and very-long-chain fatty acids. We aim to summarize the key genes in lipid biosynthesis, metabolism, and transcriptional regulation underpinning rice’s developmental and growth processes, biotic stress responses, abiotic stress responses, fertility, seed longevity, and recent efforts in rice oil genetic improvement.
1. Introduction
Lipids are organic nonpolar molecules containing hydrogen, carbon, and oxygen atoms, which provide the framework for the structure and function of living cells. As a major type of lipid, triglycerides are stored as fat in adipose cells, serving as the high-density energy-storage depot for organisms and providing thermal insulation. Some lipids, like steroid hormones, act as chemical messengers between cells, tissues, and organs [1]. In terms of cellular function, lipids are integral components of cell membranes, contributing to their structure, flexibility, and permeability. They are also involved in cell signaling and the regulation of membrane-bound enzymes [2]. Plant-derived or vegetable oils are of significant importance to human society due to their versatile and wide-ranging applications. These oils are crucial dietary fats for human consumption and are extensively utilized in culinary practices and food production. They also serve as essential raw materials for the production of biofuels, offering sustainable alternatives to traditional fossil fuels. In the cosmetic and personal care industry, plant-derived oils are prized for their emollient and moisturizing properties, making them valuable ingredients in skincare products, soaps, and hair care formulations. Furthermore, these oils play a vital role in industrial processes, contributing to manufacturing paints, lubricants, and various commercial products [3]. Their diverse applications highlight the pivotal role of plant-derived oils in both consumer products and industrial processes.
Given the biological and economic importance of plant-derived lipids, the genetic pathways and regulatory networks of lipid biosynthesis and metabolism have been extensively studied and thoroughly reviewed, particularly in oil crops such as soybeans, rapeseed, and sunflowers [4,5,6]. Rice is a staple food for more than half of the world’s population, particularly in Asia. The lipids in rice grains comprise triglycerides (TAGs), phospholipids, fatty acids, and other bioactive lipid components, with a total amount ranging from 2 to 3%, much lower than oil crops [7]. The lipids are concentrated in rice embryos and brans, essentially determining milling, appearance, and cooking quality [7]. In addition, lipids are the essential building blocks of cellular membranes and act as signal molecules to modulate various biological processes [2]. Despite the full recognition of the significant roles of lipids in rice, less attention has been paid to lipids when compared to proteins and nucleic acids in the rice research community, leaving a considerable knowledge gap in the biological roles of lipids [8]. Fortunately, recent molecular genetic research has uncovered a number of rice genes involved in lipid biosynthesis, metabolism, and signaling pathways underpinning various biological processes. This review endeavors to furnish a current overview of the advancements in understanding the genetic foundation and physiological functions of lipids in the context of rice’s developmental and growth processes, biotic stress responses, abiotic stress responses, and fertility, as well as seed longevity (Figure 1; Table 1).
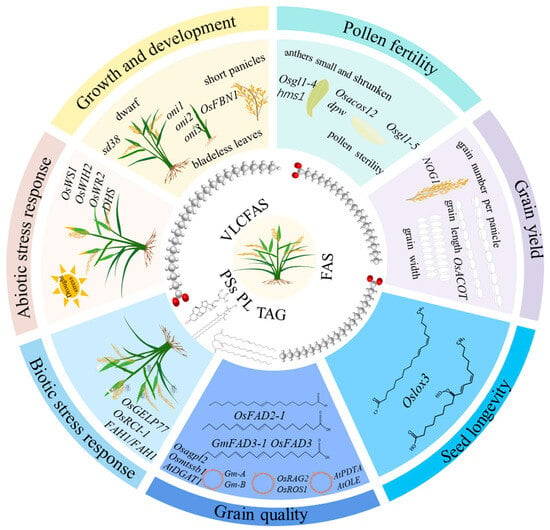
Figure 1.
Significant roles of lipids in rice. FAS: fatty acids; PSs: phytosterols; PL: Phospholipid; TAG: triacylglycerol; VLCFAS: very-long-chain fatty acids.

Table 1.
A summary of the lipid-related genes and their functions in rice.
2. Roles of Lipids in Rice Growth and Development
In very-long-chain fatty acids (VLCFAs) biosynthesis, β-ketoacyl CoA synthase (E.C.2.3.1.119, KCS) is the key enzyme. Knock-out of WSL1 encoding a typical KCS protein resulted in pleiotropic phenotypes, including reduced growth, leaf fusion, sparse wax crystals, and low fertility. The observed phenotype might be attributed to the significant reduction of VLCFA precursors of C20–C24 and total cuticular wax load on wsl1 leaf blades and sheaths [9].
From a series of rice onion mutants showing small cabbage-like shoots or bladeless leaves, three lipid-related genes involved in the shoot apical meristem were fine-mapped [43]. As a fatty acid elongase (β-ketoacyl CoA synthase), ONI1 biochemically synthesizes VLCFAs in the outermost epidermal cell layer [10]. Similarly, ONI2 catalyzes the first step of elongation reactions of a carbon chain of VLCFAs. Disruption of ONI2 led to tiny shoots in which leaves were fused and ceased growing after germination [11]. ONI3 was shown to be the ortholog of Arabidopsis HOTHEAD, containing glucose-methanol-choline (GMC) oxidoreductase and NAD(P)-binding Rossmann-like domains. It is functionally involved in the biosynthesis of long-chain fatty acids. Compared to oni1 and oni2, oni3/mini1 showed more severe growth defects with fused neighboring organs and finally became lethal after germination [12,13].
Fibrillins (FBNs) are a conserved plastid-lipid-associated protein (PAP) family modulating lipid metabolism. In rice, Fibrillin 1 (OsFBN1), a chloroplast-localized protein, was found to specifically bind C18 and C20 fatty acids in vitro. Over-expression lines of OsFBN1 exhibited more tillers, short panicles, poor grain-filling percentage, and JA levels than wild-type and RNAi- silencing lines under heat stress. More interestingly, more plastoglobules, which are defined as a hub of lipid metabolism in the chloroplast [14], were observed in the over-expression lines, suggesting the essential role of rice OsFBN1 in plastoglobule formation and plant growth [44].
Zhang et al. (2022) identified a rice semi-dwarf mutant, semi-dwarf 38 (sd38), with significantly reduced cell length. They revealed that SD38 encodes a fatty acid elongase responsible for the synthesis of C24:0 VLCFAs (VLCFAs). Exogenous application of VLCFA (C24:0) or ethephon could partially reverse the dwarf phenotype of sd38 [15].
3. Lipids Control Rice Pollen Fertility
The outer layer of the anther, called the anther cuticle, and the pollen wall’s outer layer, known as pollen exine, are essential lipid layers for the development of the male reproductive organ [45]. The anther cuticle comprises cutin and cuticle wax, while the exine is composed of sporopollenin, a highly resistant biopolymer derived from fatty acids [46]. It is well established that defective lipid synthesis in the cuticle or exine leads to nonviable pollen and sterile plants. To date, numerous genes related to lipid metabolism that cause male sterility have been identified, and their functions have been elucidated in rice and other plant species [45].
Wax-deficient anther1 (Wda1), designated as OsGL1-5, was cloned from a T-DNA insertional mutant showing pollen sterility and significant defects in the biosynthesis of VLCFAs in both layers. It was observed that epicuticular wax crystals were absent in the outer layer of the anther, and pollen exine formation was compromised in the mutant anthers [16]. Besides Wda1, rice acyl-CoA synthetase5 (OsACOS5) is also implicated in pollen exine development and fertility [17,18].
In plant cuticles, fatty alcohols synthesized by fatty acyl-CoA reductase are important fatty constituents [46]. DPW (Defective Pollen Wall) has been identified as a novel fatty acid reductase mediating the production of 1-hexadecanol. DPW and its Arabidopsis ortholog MS2 are functionally conserved, as their corresponding mutants display defected anther and degenerated pollen grains with an irregular exine [19,47]. In dpw anthers, cutin monomers were dramatically reduced with an altered composition of cuticular wax, soluble fatty acids, and alcohols. The work showcased the link between primary fatty alcohol synthesis, anther cuticle, and pollen sporopollenin biosynthesis in monocots and dicots [19].
GDSL esterases and lipases, featured by the conserved motif Gly-Asp-Ser-Leu, are a subfamily of hydrolytic/lipolytic enzymes [48]. Zhao et al. (2020) map-based cloned an endoplasmic reticulum-localized GDSL lipase gene RMS2 from an irradiation-induced mutant population. rms2 exhibited complete male sterility, while the vegetative growth was normal. Biochemically, RMS2 possesses lipid hydrolase activity, while the dysfunction of RMS2 led to significant changes in the content of 16 lipid components and numerous other metabolites. Interestingly, RMS2 might be a key node in the rice male fertility regulatory network, since master male fertility regulators Undeveloped Tapetum1 (UDT1) and Persistent Tapetal Cell1 (PTC1) could activate its transcription [49].
A recent study revealed that lipid metabolism might be subject to epigenetic regulation [20]. PEM1 (pollen expressed MBD-like 1) encoding a methyl-CpG-binding domain protein is involved in the pollen exine development. pem1 anthers became small and shrunken with 30% lower viable pollen grains when compared to the WT. Unlike many of the other male sterile mutants showing retarded tapetum degradation, pem1 mutants had regular PCD progress of the tapetum. However, they exhibited abnormal Ubisch body formation, delayed exine occurrence, defective exines, and increased anther cuticles [20]. Additionally, a novel alpha integrin-like protein DPW3 (Defective Pollen Wall3) has been associated with pollen wall formation and fertility, although the detailed molecular mechanism requires further exploration [50].
Humidity-sensitive genic male sterility (HGMS) lines have great potential in hybrid rice breeding. However, HGMS-related genes have yet to be identified. Chen et al. (2020) isolated a rice HGMS mutant that exhibited male sterility at low humidity but was fully fertile at high humidity [21]. The causal gene HUMIDITY-SENSITIVE GENIC MALE STERILITY 1 (HMS1) encodes a putative β-ketoacyl-CoA synthase. Through interacting with its co-factor HMS1-INTERACTING PROTEIN (HMS1I), HMS1 promotes the conversion of C24:0 and C26:0 fatty acids into C26:0 and C28:0 fatty acids on the pollen wall, which helps to protect pollen from dehydration under low-humidity conditions. In a yeast system, HMS1 and HMS1I were found to catalyze the biosynthesis of VLCFAs longer than C24. Notably, the authors developed an HGMS line using the HMS1 gene with convertible pollen fertility under different humidity conditions, suggesting that HMS1 can be potentially used for hybrid breeding in indica and japonica rice.
Unlike other reported OsGLs involved in drought resistance, OsGL1-4/CER1 was identified as a critical regulator in male reproductive development. Ni et al. (2018) revealed that OsGL1-4/CER1 functions in VLC alkanes biosynthesis, underpinning anther development and plastids differentiation. OsGL1-4/CER1 transcription was robustly detected in the developed tapetum and bicellular pollen cells. Meanwhile, disruption of OsGL1-4/CER1 significantly reduced the content of VLC alkanes (C25 and C27), finally leading to sterile pollen grains containing fewer amyloplasts [22]. A few years later, two independent groups both found that the pollen sterility of cer1 is attributed to the excessively fast dehydration at anthesis and defective adhesion and hydration under normal conditions, which could be recovered by artificial high humidity [51,52]. Ni et al. (2021) further specified that the lipid composition of tryphine is the primary reason for such humidity-sensitive genic male sterility, since the fertility could be recovered by co-pollination with mixed OsCER1Cas mutant and maize pollens [52].
4. Lipids Involves in the Regulation of Grain Yield
Enoyl-CoA hydratase (ECH) catalyzes the second step in fatty acid metabolism’s physiologically important beta-oxidation pathway [53]. A Chinese research group found that NUMBER OF GRAINS 1 (NOG1) encoding an enoyl-CoA hydratase/isomerase regulates grain number per panicle without penalty on the panicle number and grain weight. A 12-bp indel in the promoter affected the transcription of NOG1, which finally contributed to the variations in total fatty acids and linolenic acid (LA, C18:3) among different varieties. In addition to the fatty acids, NOG1 is involved in the biosynthesis of JA, since C18:3 is the synthetic precursor of JA. Introgression of NOG1 in modern cultivars achieved up to a 25.8% increase in grain yield, showing the massive potential of lipid-related genes in crop genetic improvement [23].
Zhao et al. (2019) identified a seed-predominantly expressed microRNA, miR1432. The suppression of miR1432 facilitated grain filling rate and could increase grain yield up to 17.14% in field trials. In their search for the downstream targets of miR1432, the authors discovered that rice Acyl-CoA thioesterase (OsACOT) is a genuine target gene negatively regulated by miR1432-directed cleavage. OsACOT serves as a crucial enzyme in the fatty acid desaturation and elongation pathway, particularly in the conversion of the fatty acid 16:0 into 18:2. Intriguingly, over-expression of miR1432-resistant form of OsACOT mimicked miR1432 suppression lines and resulted in an increased yield of approximately 50% [24].
5. Grain Quality Is Affected by Lipid Contents and Components
Although rice grains possess a relatively low lipid content, the lipids in rice bran and grains exhibit a balanced composition of saturated (e.g., C16:0) and unsaturated (e.g., C18:1 and C18:2) fatty acids, which presents rice as a valuable and healthful oil source for human consumption [54]. Over the past few decades, substantial efforts have been dedicated to investigating the genetic underpinnings of rice lipid metabolism, with a particular emphasis on fatty acids. Employing a classic QTL mapping strategy, three QTLs, qRFC-1, qRFC-2, and qRFC-5, that accounted for 44% of the fat content variation were mapped in a doubled haploid population [55]. Later, two other groups did similar work, identifying nearly 20 QTLs contributing to grain lipid content. Nevertheless, no candidate genes underlying the QTLs were uncovered [56,57]. Ying et al. (2012) mapped 29 fatty acid content-associated QTLs, including eight QTLs that were repeatedly detected across multiple years and 11 QTLs harboring orthologs of key Arabidopsis lipid metabolism enzymes. Interestingly, a strong QTL for oleic (18:1) and linoleic (18:2) acids was associated with a rice ortholog of a gene encoding acyl-CoA: diacylglycerol acyltransferase (DGAT) and another for palmitic acid (16:0) mapped similarly to the acyl-ACP thioesterase (FatB) gene ortholog [58]. In 2021, a genome-wide association study was conducted in a diverse panel of 533 cultivated rice accessions and identified 46 loci controlling oil composition and concentration. Genes involved in lipid metabolism, namely PAL6, LIN6, MYR2, and ARA6, are significant contributors to the natural variance in oil composition within rice subpopulations. Notably, qPAL6 exhibits a substantial effect, explaining 49.86% and 57.44% of the phenotypic variance in the C16:0 composition. Similarly, qLIN6 accounts for 28.56% and 55.31% of the phenotypic variance in C18:2 conversion from C18:1. Furthermore, qMYR2 encodes a myristoyl-ACP thioesterase responsible for the hydrolysis of specific acyl-ACPs, leading to the production of saturated fatty acids. Disruption of qMYR2 results in lowered C14:0 and C16:0 but increased C18:2. Additionally, qARA6, annotated as a 3-ketoacyl-CoA synthase, modulates the biosynthesis of very-long-chain fatty acids (C20–C22). Functional impairment of qARA6 significantly reduces the C20:0, C20:1, C20:2, and C22:0 components [59].
The economic significance of rice oil has led to the implementation of various strategies aimed at enhancing the lipid content and healthy fatty acid components through genetic means. One key strategy involves boosting lipid synthesis efficiency by utilizing lipid rate-limiting enzymes or regulator genes from rice or other plant species. WRINKLED1 (WRI1) is a master regulator of seed oil biosynthesis in Arabidopsis. Over-expression of AtWRI1 in rice increased fatty acid content by 30–40% in vegetative organs but decreased fatty acid content in the endosperm, whereas the endosperm-specific expression of CoWRI1 from coconut showed minor effects on rice oil content [25,60]. Over-expression of RAG2, a member of 14-to-16-kDa α-amylase/trypsin inhibitors in rice, gave rise to a 10–30% increase in lipid content and higher storage protein levels [26].
However, given that the overall carbon flux to starch is dominant over lipid biosynthesis, it is not surprising that minor effects could be observed when only lipid biosynthesis is enhanced [31]. As suggested by a report on Chlamydomonas reinhardtii, in which lipid content increased by 3.5-fold when an essential starch synthesis gene was knocked out [61], blocking the starch biosynthesis might be an efficient strategy to divert the carbon flux to lipid biosynthesis. Indeed, Wei et al. (2017) knocked out an ADP-glucose pyrophosphorylase (AGP) large subunit gene to have the oil content in rice grains over 10% [27].
The protein oleosin is notably abundant in the oil bodies of plant seeds, and it serves a crucial function in the accumulation of lipids. By introducing two soybean oleosin genes that encode 24 kDa proteins in rice, under the control of an embryo-specific rice promoter REG-2, the transgenic rice showed more oil bodies in smaller size and a significant increase in seed lipid content. Specifically, the lipid content increased by 36.93% and 46.06% compared to the control, while the fatty acid profiles of triacylglycerols remained unchanged [28]. In addition, modifying OsROS1 and OsMitssb1 significantly increased the thickness of rice aleurone layer cells, which could finally lead to an over-48% increase in grain oil content, representing an alternative strategy in rice lipid metabolic engineering [29,62].
In pursuit of achieving a balanced carbon flux between carbohydrates and lipids in rice, Izadi-Darbandi et al. (2020) undertook the metabolic engineering of rice grains by integrating Arabidopsis genes such as AtWRI1, AtOle, diacylglycerol acyltransferase (AtDGAT), and phospholipid:diacylglycerol acyltransferase (AtPDAT) to facilitate the final step of TAG assembly. This intervention substantially increased TAG, oleic acid, palmitic acid, and total oil contents by 26%, 28%, 27%, and 70%, respectively [30]. In 2023, Liu et al. published a significant case of multigene engineering aimed at augmenting lipid content in rice grains. The goal was achieved by the concurrent knock-out of two rice genes, AGPL2 and Mtssb1, and the endosperm-specific expression of DGAT1 from Arabidopsis. These modifications effectively redirected carbon flux from starch to lipid, resulting in a substantial increase in oil biosynthesis, elevating the lipid content from approximately 2% to around 12%. Meanwhile, the fatty acid compositions were significantly altered with more beneficial components such as GLA. A lipid content of 12% is very close to oil crops like soybean, which represented the highest lipid level ever reported in rice or other starchy-type grains [31]. This work highlighted a practical approach for genetically improving oil contents in rice and other crops with starchy grains.
Rice oil comprises approximately 18% palmitic acid, 36% oleic acid, and 37% linoleic acid. Linoleic acid is particularly susceptible to non-enzymatic oxidation due to its oxidative instability [63]. The high oleic acid content in rice has consistently been a desirable characteristic due to its enhanced stability and beneficial nutritional profile. It is well known that the microsomal omega-6 fatty acid desaturase (FAD2) catalyzes the conversion of oleic acid to linoleic acid in seed oil. In Arabidopsis and other cops, knock-out or knock-down of FAD2 successfully produced high oleic and low linoleic oil seeds [64,65]. In rice, the suppression of FAD2-1, one of the transcripts of FAD2 predominantly expressed in seeds, through RNA interference (RNAi), has been observed to lead to an increase in oleic acid and a reduction in linoleic and palmitic acids in the grains. This finding suggests that OsFAD2-1 represents a viable target for enhancing the genetic profile of fatty acids in rice [32,33]. In an attempt to enhance the α-linolenic acid (ALA) content and to reduce the ratio of linoleic acid (LA) to ALA, Liu et al. (2012) systematically introduced six ω-3 (Δ-15) fatty acid desaturase (FAD) genes from rice and soybean into the rice. The ALA contents in the seeds of GmFAD3-1 and OsFAD3 over-expression lines exhibited a consistent increase of 23.8- and 27.9-fold across multiple generations. Comparative analysis revealed that the endosperm-specific promoters GluC and REG outperformed the constitutive maize Ubi-1 promoter in enhancing ALA levels in rice embryos and bran [34,66].
6. Lipids Regulate Seed Longevity in Rice
The longevity and viability of seeds are closely associated with the presence of lipid peroxidation products, specifically malondialdehyde and acetaldehyde. These products can lead to cell damage and intoxication by reacting with macromolecules. Linolenic acid (LNA) and LA are the most significant polyunsaturated fatty acids. The peroxidation of LNA and LA is typically linked to the breakdown of cell structure, the formation of cytotoxic products, and the release of volatile decomposition products, all of which contribute to the deterioration of rice seed longevity. Lipoxygenases, a conserved family of fatty acid dioxygenases, facilitate the addition of oxygen to polyunsaturated fatty acids such as LNA and LA. Among the three embryo-derived LOX isoenzymes, LOX3 is at a dominant level and has been linked to fatty acid peroxidation during seed storage and stored grain quality [67]. Xu et al. (2015) found that LOX3 is capable of producing 9-hydroperoxyoctadecadienoic acid (9-HPOD) using LA as substrate. Meanwhile, suppressing LOX3 expression in rice endosperm successfully extended rice seed longevity under either artificial aging or natural aging conditions, whereas the major agronomic traits remain unchanged [35]. In addition, the knock-down of LOX3 could also effectively protect the functional component β-carotene from deterioration in the carotenoid-enriched golden rice [68].
7. Lipids Related to Abiotic Stress Response in Rice
Cuticular wax on the leaf aerial surfaces is the outermost diffusion barrier against the uncontrolled loss or uptake of water and gases [69]. To most plants, cuticular waxes are complex mixtures of primarily VLCFAs, hydrocarbons, alcohols, flavonoids, and so on [46], though the proportions of the major constituents may vary from each other. Because the defective accumulation or altered composition of wax could be visually detected, screening of cuticular wax mutants like eceriferum (cer) in Arabidopsis and Glossy1 in maize has been carried out for decades [46,70]. In rice, Islam et al. (2009) systematically identified 11 homologs of maize GL1, designated as OsGL1-1 to OsGL1-11. According to phylogenetic analysis, most of the OsGL1s could be induced by abiotic stresses and might be involved in cuticular or epicuticular wax biosynthesis [71]. At least, osgl1-1, osgl1-2, osgl1-3, and osgl1-6 have been found to participate in drought stress response in rice. Compared to the WT, the osgl1-2 exhibited less wax crystallization, a thinner cuticular layer, and reduced total cuticular wax amount, leading to higher sensitivity to drought stress at the reproductive stage. In contrast, the over-expression lines showed the opposite phenotypes [71]. OsGL1-1 was annotated as a sterol desaturase or short-chain dehydrogenase/reductase. OsGL1-3 protein has a conserved fatty acid hydroxylase domain (FAH domain) and a WAX2 C-terminal domain, while OsGL1-6 gene is a member of the fatty aldehyde decarbonylase gene family. The osgl1-1, osgl1-3 and osgl1-6 mutants showed very similar phenotypes to osgl1-2, with decreased cuticular wax deposition, thinner cuticular membranes, and hypersensitivity to drought stress [72,73,74].
It was found that osa-miR1848 regulates OsWS1, a membrane-bound O-acyl transferase gene member, to determine wax biosynthesis [36]. OsWS1 and osa-miR1848 exhibited a negative time-and-spatial co-expression pattern, especially under water-deficit treatment, suggesting antagonistic roles of the two factors. Compared to the control, OsWS1 over-expression lines displayed 35% higher VLCFA contents, denser wax papillae around the stoma, and a higher survival rate upon water-deficit treatment. Conversely, OsWS1-RNAi and osa-miR1848 over-expression plants exhibited opposing changes.
OsWIH2 is another reported fatty acid synthesis enzyme that positively regulates rice drought resistance by alleviating water loss and reactive oxygen species (ROS) accumulation and altering wax content [37]. OsWIH2 works as a complex with HOTHEAD (HTH), which is an α-alcohol dehydrogenase catalyzing the biosynthesis of long-chain α-,ω-dicarboxylic fatty acids (LCFAs) in the cutin and wax biosynthesis pathway [75]. Meanwhile, drought-inducible transcription factor OsbHLH130 could activate the transcription of OsWIH2, thus forming a bHLH130-OsWIH2-HTH regulatory module in cuticular wax biosynthesis and drought response.
Besides the wax or cuticle synthesis enzymes, several genes involved in the regulation of wax biosynthesis and drought tolerance have been identified. These include the transcription factor CFL1, OsWR2, and DWA1 [39,76,77,78]. Wang et al. (2018) reported that DROUGHT HYPERSENSITIVE (DHS) negatively regulates wax biosynthesis by promoting the turnover of an HD-ZIP IV family member ROC4, thereby influencing drought tolerance [38]. Over-expression of DHS significantly reduced the cuticular wax contents and conferred drought hypersensitivity. As a RING-type E3 ligase, DHS functions in mediating the ubiquitination of ROC4 in vivo, subsequently leading to its degradation via the ubiquitin/26S proteasome-mediated pathway. Notably, the DHS-ROC4 module has been identified as directly targeting Os-BDG, a putative regulator of wax biosynthesis. Thus, this study elucidates a precisely regulated molecular mechanism that links wax biosynthesis with drought stress response in rice, offering valuable insights for developing drought-tolerant rice cultivars.
Xiang et al. (2022) identified Salt Tolerance and Heading Date 1 (STH1) from the African rice species Oryza glaberrima. STH1 encodes an α/β hydrolase family member primarily localized in the major sites of peroxisomal β-oxidation, including the cytoplasm and mitochondria. It catalyzes the hydrolytic degradation of unsaturated fatty acids to maintain fatty acid metabolic homeostasis and membrane fluidity, thus controlling rice salt, heat, and osmotic tolerance. Correspondingly, rice seedlings showed a more significant growth acceleration in terms of shoot length when grown on a mannitol-containing medium supplemented with the fatty acid 11-eicosenoic acid, implying that metabolic flux redirection by blocking fatty acids from degradation can enhance plant tolerance to various abiotic stresses. STH1HP46 is found to be a rare allele in modern Asian cultivars, which shows great promise in boosting grain yield under salt stress [79].
Very few favorable thermo-tolerance genes have been identified, mostly working through reactive oxygen species scavenging, cytotoxic protein elimination, and unfolded protein renaturation [80,81]. Kan et al. (2022) revealed a novel “TT2-SCT1-OsWR2” thermal tolerance mechanism linking G protein, Ca2+ sensing, and wax metabolism [82]. Known as a Gγ subunit protein, TT2 was cloned as a major QTL contributing to heat tolerance. Heat-treated plants carrying a weak allele of TT2 had enhanced thermo-tolerance, exhibiting less-severe reductions in wax content compared with plants with the normal TT2 allele. The authors have validated that the heat signal increases cytosolic Ca2+, which subsequently undergoes conversion into an active form facilitated by CaM. Following this, CaM binds to SCT1 to repress its activity, thereby decreasing the transcription of OsWR2, the pivotal regulator of wax biosynthesis [39]. Consequently, this perturbation results in reduced wax content and induces thermo-sensitivity. Field trial experiment results showed that TT2 governs rice thermo-tolerance in both vegetative and reproductive stages. Under heat stress, NIL-TT2HPS32 plants significantly outperformed NIL-TT2HJX plants with 54.7% higher grain yield, showing promising potential in breeding thermo-tolerant rice varieties.
8. Rice Biotic Stress Response Is Mediated by Lipids
Microdomains, also called membrane rafts, are small, heterogeneous, liquid-ordered domains composed mainly of sphingolipids and sterols [83]. Since plant defense-related proteins primarily congregate in plasma membrane microdomains, innate immunity is associated with microdomains’ functions. By knocking down two fatty acid 2-hydroxylases catalyzing the 2-hydroxylation of sphingolipid fatty acids, Nagano et al. (2016) obtained fah1 fah2 genetic lines harboring fewer microdomains and higher susceptibility to rice blast fungus infection. Os-RbohB and Os-RbohH are microdomains-located NADPH oxidases involved in the pathogen defense process. They revealed that microdomains are required for the dynamics of the GTPase Rac1 and respiratory burst oxidase homologs (Rbohs) in response to chitin elicitors, regulating chitin-induced immunity through ROS signaling mediated by the Rac1-RbohB pathway [40].
Many plant phosphatidylinositol phosphates, particularly PtdIns(4,5)P2, have been demonstrated to be disease-susceptibility factors [84]. From a rice lesion mimic mutant with broad-spectrum disease resistance, Sha et al. (2023) cloned a cytidine diphosphate diacylglycerol synthase gene RBL1 involved in phospholipid biosynthesis. rbl1 had drastically reduced the amount of phosphatidylinositol and its derivative PtdIns(4,5)P2, which is enriched in the biotrophic interfacial complex and extra-invasive hyphal membrane structure associate with effector secretion and fungal infection. Intriguingly, the authors generated a valuable allele named RBL1Δ12, which confers broad-spectrum disease resistance to 10 M. oryzae field strains, five Xoo strains, and two rice false smut Ustilaginoidea virens strains in model rice variety backgrounds. At the same time, no yield penalty was observed in multiple field trials [85]. The work demonstrated the feasibility of balancing rice growth and immunity via genetically editing lipid-related genes.
In addition to its functions in seed longevity, Lipoxygenase 3 (LOX3) may negatively influence rice blast disease resistance. lox3 mutants grow normally with high levels of methyl-linolenate and reactive oxygen species (ROS). After M. oryzae infection, lox3 plants exhibited serious blast symptoms and reduced defense response, accompanied by increased ROS-mediated cell death. Moreover, exogenous JA repressed hyphal expansion and ROS-mediated cell death, finally alleviating blast symptoms in the mutant [86].
A recent publication described the functional characterization of OsGELP77, a major QTL contributing to broad-spectrum resistance in rice. OsGELP77 is annotated as an ER-localized, GDSL-type lipase with typical lipase activity. Based on its pathogen- and JA-inducible expression pattern, OsGELP77 was found to positively regulate resistance to various pathogens, possibly through modulating glycerolipid metabolism, glycerophospholipid metabolism, and sphingolipid metabolism as well as JA homeostasis. In addition, OsWRKY45 may work upstream of OsGELP77 to trigger rice immunity. Elevated expression of OsGELP77 or pyramiding of a natural elite haplotype OsGELP77Hap3 significantly increased resistance to various pathogens to achieve higher yield [41].
Oxylipins are a big family of lipid compounds well documented for their function in insect resistance in rice. LOX11 (Lipoxygenase-11)/OsRCI-1 is a chloroplast-localized protein gene in rice showing induced expression by brown planthopper (BPH) infestation. Over-expression of OsRCI-1 elevated the levels of JA, jasmonate-isoleucine, and trypsin protease inhibitors, which imposed decreased colonization, fecundity, and mass of the BPH insects when compared with the WT. Moreover, the decreased attractiveness to BPH and enhanced attractiveness to the parasitoid of oeRCI plants correlated with higher levels of BPH-induced 2-heptanone, 2-heptanol, and 2-heptanone. These results indicate that OsRCI-1 is involved in herbivore-induced JA bursts and plays a role in plant defense [42].
9. Perspectives
This comprehensive review delineates recent progress in elucidating the genetic underpinnings and physiological functions of lipids in rice, a model species for monocots and cereals. Despite the aforementioned advancements, our knowledge of the biological roles of lipids in rice remains relatively rudimentary and fragmented compared to that in Arabidopsis and other oil crops. The bulk of published rice cases hinge on robust correlations between phenotype and variations in lipid content or components. However, the molecular mechanisms underlying the relationship between lipid variation and phenotypic diversity remains largely unexplored. One example that has been thoroughly documented is the role of STH1 in catalyzing the hydrolytic degradation of unsaturated fatty acids, which is essential for maintaining membrane fluidity [79]. Meanwhile, a large number of rice lipid-related genes have been characterized via a forward genetics approach utilizing randomly collected mutants and map-based cloning, which resulted in a fragmented comprehension of the rice lipid regulation network. Thus, there is a need to allocate additional resources to systematically explore the gene families associated with lipid biosynthesis, metabolism, and signaling. Finally, as has been shown in a few cases in rice [87], the advancement of new mass spectrometric techniques in lipidomic research has the potential to significantly enhance the identification of novel lipid components within the pathways and networks of cellular lipids in rice biological systems.
Author Contributions
Conceptualization, K.Z., Z.L. (Zhengliang Luo), J.Z., S.W. and X.Z. investigation, W.H., Z.L. (Zemin Liu) and X.M.; writing—original draft preparation, K.Z., J.Z., S.W. and X.Z.; writing—review and editing, K.Z., S.W. and X.Z.; supervision, X.Z., S.T. and J.W.; funding acquisition, K.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the agricultural science and technology innovation fund project of Hunan Province (China) (2023CX09), Central Public-interest Scientific Institution Basal Research Fund (No. Y2023PT17), National Key Research and Development Program of China (2023YFD1201201) and Changsha Natural Science Foundation (kq2208128).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
References
- Gurr, M.I.; Harwoord, J.L.; Frayn, K.N.; Murphy, D.J.; Michell, R.H. Lipids: Biochemistry, Biotechnology and Health, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2016. [Google Scholar]
- Suh, M.C.; Kim, H.U.; Nakamura, Y. Plant lipids: Trends and beyond. J. Exp. Bot. 2022, 73, 2715–2720. [Google Scholar] [CrossRef]
- Tao, B. Industrial Applications for Plant Oils and Lipids. In Bioprocessing for Value-Added Products from Renewable Resources; Yang, S.-T., Ed.; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Harwood, J.L.; Guschina, I.A. Regulation of lipid synthesis in oil crops. FEBS Lett. 2013, 587, 2079–2081. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Lu, H.; Zhang, Y.; Cheng, J.; Huang, B.; Lu, X.; Sheteiwy, M.S.; Kuang, S.; Shao, H. Oil crop genetic modification for producing added value lipids. Crit. Rev. Biotechnol. 2020, 40, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Sagun, J.V.; Yadav, U.P.; Alonso, A.P. Progress in understanding and improving oil content and quality in seeds. Front. Plant Sci. 2023, 14, 1116894. [Google Scholar] [CrossRef]
- Zhou, Z.K.; Robards, K.; Helliwell, S.; Blanchard, C. Composition and functional properties of rice. Int. J. Food Sci. Technol. 2002, 37, 849–868. [Google Scholar] [CrossRef]
- Li, P.; Chen, Y.H.; Lu, J.; Zhang, C.Q.; Liu, Q.Q.; Li, Q.F. Genes and Their Molecular Functions Determining Seed Structure, Components, and Quality of Rice. Rice 2022, 15, 18. [Google Scholar] [CrossRef]
- Yu, D.M.; Ranathunge, K.; Huang, H.; Pei, Z.Y.; Franke, R.; Schreiber, L.; He, C. Wax Crystal-Sparse Leaf1 encodes a beta-ketoacyl CoA synthase involved in biosynthesis of cuticular waxes on rice leaf. Planta 2008, 228, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Kimura, F.; Hirakata, K.; Tsuda, K.; Takasugi, T.; Eiguchi, M.; Nakagawa, K.; Kurata, N. Fatty acid elongase is required for shoot development in rice. Plant J. 2011, 66, 680–688. [Google Scholar] [CrossRef]
- Tsuda, K.; Akiba, T.; Kimura, F.; Ishibashi, M.; Moriya, C.; Nakagawa, K.; Kurata, N.; Ito, Y. ONION2 Fatty Acid Elongase is Required for Shoot Development in Rice. Plant Cell Physiol. 2013, 54, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.X.; Hu, J.; Xu, J.; Yu, H.P.; Shi, Z.Y.; Xiong, G.S.; Zhu, L.; Zeng, D.; Zhang, G.; Gao, Z.; et al. Identification and characterization of Mini1, a gene regulating rice shoot development. J. Integr. Plant Biol. 2015, 57, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Akiba, T.; Hibara, K.I.; Kimura, F.; Tsuda, K.; Shibata, K.; Ishibashi, M.; Moriya, C.; Nakagawa, K.; Kurata, N.; Itoh, J.I.; et al. Organ fusion and defective shoot development in oni3 mutants of rice. Plant Cell Physiol. 2014, 55, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Shanmugabalaji, V.; Zita, W.; Collombat, J.; Kessler, F. Plastoglobules: A hub of lipid metabolism in the chloroplast. Adv. Bot. Res. 2022, 101, 91–119. [Google Scholar]
- Zhang, X.B.; Wang, Y.; Wang, X.W.; Zhu, Z.; Zhang, X.F.; Jia, L.Q.; Li, Y.Y.; Tian, W.J.; Chen, H.Y.; Zhu, X.Y.; et al. A very-long-chain fatty acid synthesis gene, SD38, influences plant height by activating ethylene biosynthesis in rice. Plant J. 2022, 112, 1084–1097. [Google Scholar] [CrossRef]
- Jung, K.H.; Han, M.J.; Lee, D.Y.; Lee, Y.S.; Schreiber, L.; Franke, R.; Faust, A.; Yephremov, A.; Saedler, H.; Kim, Y.W.; et al. Wax-deficient anther1 is involved in cuticle and wax production in rice anther walls and is required for pollen development. Plant Cell 2006, 18, 3015–3032. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.J.; Liang, W.Q.; Chen, M.J.; Zhang, D.B.; Zhao, X.X.; Shi, J.X. Rice fatty acyl-CoA synthetase OsACOS12 is required for tapetum programmed cell death and male fertility. Planta 2017, 246, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Li, D.D.; Guo, Z.L.; Shi, Q.S.; Xiong, S.X.; Zhang, C.; Zhu, J.; Yang, Z.N. OsACOS12, an orthologue of Arabidopsis acyl-CoA synthetase5, plays an important role in pollen exine formation and anther development in rice. BMC Plant Biol. 2016, 16, 256. [Google Scholar] [CrossRef]
- Shi, J.; Tan, H.X.; Yu, X.H.; Liu, Y.Y.; Liang, W.Q.; Ranathunge, K.; Franke, R.B.; Schreiber, L.; Wang, Y.; Kai, G.; et al. Defective Pollen Wall Is Required for Anther and Microspore Development in Rice and Encodes a Fatty Acyl Carrier Protein Reductase. Plant Cell 2011, 23, 2225–2246. [Google Scholar] [CrossRef]
- Song, Y.Y.; Tang, Y.Y.; Liu, L.T.; Xu, Y.Y.; Wang, T. The methyl-CpG-binding domain family member PEM1 is essential for Ubisch body formation and pollen exine development in rice. Plant J. 2022, 111, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Q.; Zhang, Z.G.; Ni, E.D.; Lin, J.W.; Peng, G.Q.; Huang, J.L.; Zhu, L.Y.; Deng, L.; Yang, F.F.; Luo, Q.; et al. HMS1 interacts with HMS1I to regulate very-long-chain fatty acid biosynthesis and the humidity-sensitive genic male sterility in rice (Oryza sativa). New Phytol. 2020, 225, 2077–2093. [Google Scholar] [CrossRef] [PubMed]
- Ni, E.D.; Zhou, L.Y.; Li, J.; Jiang, D.G.; Wang, Z.H.; Zheng, S.Y.; Qi, H.; Zhou, Y.; Wang, C.M.; Xiao, S.; et al. OsCER1 Plays a Pivotal Role in Very-Long-Chain Alkane Biosynthesis and Affects Plastid Development and Programmed Cell Death of Tapetum in Rice (Oryza sativa L.). Front. Plant Sci. 2018, 9, 1217. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Wu, S.; Zhu, Z.F.; Liu, F.X.; Fu, Y.C.; Cai, H.W.; Sun, X.Y.; Gu, P.; Xie, D.X.; Tan, L.B.; et al. NOG1 increases grain production in rice. Nat. Commun. 2017, 8, 1497. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-F.; Peng, T.; Sun, H.-Z.; Teotia, S.; Wen, H.-L.; Du, Y.-X.; Zhang, J.; Li, J.Z.; Tnag, G.L.; Xue, H.W.; et al. miR1432-OsACOT (Acyl-CoA thioesterase) module determines grain yield via enhancing grain filling rate in rice. Plant Biotechnol. J. 2019, 17, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Tian, R.C.; Gao, Z.Q.; Yang, H.B. Characterization of AtWRI1 in fatty acids and starch synthesis in rice. Biosci. Biotechnol. Biochem. 2019, 83, 1807–1814. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, X.; Zhou, D.; Ouyang, Y.; Yao, J. Overexpression of the 16-kDa α-amylase/trypsin inhibitor RAG2 improves grain yield and quality of rice. Plant Biotechnol. J. 2017, 15, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Jiao, G.; Lin, H.; Sheng, Z.; Shao, G.; Xie, L.; Tang, S.; Xu, Q.; Hu, P. GRAIN INCOMPLETE FILLING 2 regulates grain filling and starch synthesis during rice caryopsis development. J. Integr. Plant Biol. 2017, 59, 134–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.X.; Liu, H.L.; Qu, L.Q. Embryo-specific expression of soybean oleosin altered oil body morphogenesis and increased lipid content in transgenic rice seeds. Theor. Appl. Genet. 2013, 126, 2289–2297. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, X.; Yao, X.; Yu, R.; Larkin, P.J.; Liu, C.-M. Mutations in the DNA demethylase OsROS1 result in a thickened aleurone and improved nutritional value in rice grains. Proc. Natl. Acad. Sci. USA 2018, 115, 11327–11332. [Google Scholar] [CrossRef]
- Izadi-Darbandi, A.; Younessi-Hamzekhanlu, M.; Sticklen, M. Metabolically engineered rice biomass and grain using genes associated with lipid pathway show high level of oil content. Mol. Biol. Rep. 2020, 47, 7917–7927. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.; Ying, J.; Shu, Y.; Liu, W.; Li, G.; Chen, L.; Luo, J.; Wang, S.; Wang, Y.; et al. Multi-gene engineering boosts oil content in rice grains. Plant Commun. 2024, 5, 100736. [Google Scholar] [CrossRef]
- Zaplin, E.S.; Liu, Q.; Li, Z.; Butardo, V.M.; Blanchard, C.L.; Rahman, S. Production of high oleic rice grains by suppressing the expression of the OsFAD2-1 gene. Funct. Plant Biol. 2013, 40, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, G.J.; Liu, Q.; Shreshtha, P.; Li, Z.Y.; Rahman, S. RNAi-mediated down-regulation of the expression of OsFAD2-1: Effect on lipid accumulation and expression of lipid biosynthetic genes in the rice grain. BMC Plant Biol. 2016, 16, 189. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.J.; Liu, H.L.; Dong, X.B.; Tian, L.H.; Xiao, L.; Xu, Y.N.; Qu, L.Q. Increasing α-linolenic acid content in rice bran by embryo-specific expression of ω3/Δ15-desaturase gene. Mol. Breed. 2014, 33, 987–996. [Google Scholar] [CrossRef]
- Xu, H.B.; Wei, Y.D.; Zhu, Y.S.; Lian, L.; Xie, H.G.; Cai, Q.H.; Chen, Q.S.; Lin, Z.P.; Wang, Z.H.; Xie, H.; et al. Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity. Plant Biotechnol. J. 2015, 13, 526–539. [Google Scholar] [CrossRef]
- Xia, K.F.; Ou, X.J.; Gao, C.Z.; Tang, H.D.; Jia, Y.X.; Deng, R.F.; Xu, X.L.; Zhang, M.Y. OsWS1 involved in cuticular wax biosynthesis is regulated by osa-miR1848. Plant Cell Environ. 2015, 38, 2662–2673. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.Y.; Gao, S.X.; Li, J.; Song, P.Y.; Zhang, Q.; Guo, J.F.; Wang, X.Y.; Han, X.Y.; Wang, X.J.; Zhu, Y.; et al. The bHLH transcription factor regulated gene OsWIH2 is a positive regulator of drought tolerance in rice. Plant Physiol. Biochem. 2021, 169, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Tian, X.J.; Zhao, Q.Z.; Liu, Z.Q.; Li, X.F.; Ren, Y.K.; Tang, J.Q.; Fang, J.; Xu, Q.J.; Bu, Q.Y. The E3 Ligase DROUGHT HYPERSENSITIVE Negatively Regulates Cuticular Wax Biosynthesis by Promoting the Degradation of Transcription Factor ROC4 in Rice. Plant Cell 2018, 30, 228–244. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.Y.; Jenks, M.A.; Liu, J.; Liu, A.L.; Zhang, X.W.; Xiang, J.H.; Zou, J.; Peng, Y.; Chen, X.B. Overexpression of Transcription Factor OsWR2 Regulates Wax and Cutin Biosynthesis in Rice and Enhances its Tolerance to Water Deficit. Plant Mol. Biol. Rep. 2014, 32, 719–731. [Google Scholar] [CrossRef]
- Nagano, M.; Ishikawa, T.; Fujiwara, M.; Fukao, Y.; Kawano, Y.; Kawai-Yamada, M.; Shimamoto, K. Plasma Membrane Microdomains Are Essential for Rac1-RbohB/H-Mediated Immunity in Rice. Plant Cell 2016, 28, 1966–1983. [Google Scholar] [CrossRef]
- Zhang, M.J.; Chen, D.; Tian, J.J.; Cao, J.B.; Xie, K.B.; He, Y.Q.; Yuan, M. OsGELP77, a QTL for broad-spectrum disease resistance and yield in rice, encodes a GDSL-type lipase. Plant Biotechnol. J. 2024, 22, 1352–1371. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.H.; Wang, L.; Li, C.Z.; Cao, M.J.; Wang, J.N.; Yao, Z.L.; Zhou, S.Y.; Zhou, G.X.; Zhang, D.Y.; Lou, Y.G. The lipoxygenase gene is involved in the biosynthesis of herbivore-induced JAs and regulates plant defense and growth in rice. Plant Cell Environ. 2022, 45, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Ito, Y.; Yamaki, S.; Miyao, A.; Hirochika, H.; Kurata, N. Isolation and mapping of three rice mutants that showed ectopic expression of KNOX genes in leaves. Plant Sci. 2009, 177, 131–135. [Google Scholar] [CrossRef]
- Li, J.J.; Yang, J.; Zhu, B.H.; Xie, G.S. Overexpressing OsFBN1 enhances plastoglobule formation, reduces grain-filling percent and jasmonate levels under heat stress in rice. Plant Sci. 2019, 285, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.Y.; Wu, S.W.; Li, Z.W.; An, X.L.; Tian, Y.H. Lipid Metabolism: Critical Roles in Male Fertility and Other Aspects of Reproductive Development in Plants. Mol. Plant 2020, 13, 955–983. [Google Scholar] [CrossRef] [PubMed]
- Kunst, L.; Samuels, A.L. Biosynthesis and secretion of plant cuticular wax. Prog. Lipid Res. 2003, 42, 51–80. [Google Scholar] [CrossRef] [PubMed]
- Aarts, M.G.M.; Hodge, R.; Kalantidis, K.; Florack, D.; Wilson, Z.A.; Mulligan, B.J.; Scott, R.; Pereira, A. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. Plant J. 1997, 12, 615–623. [Google Scholar] [CrossRef]
- Akoh, C.C.; Lee, G.C.; Liaw, Y.C.; Huang, T.H.; Shaw, J.F. GDSL family of serine esterases/lipases. Prog. Lipid Res. 2004, 43, 534–552. [Google Scholar] [CrossRef]
- Zhao, J.; Long, T.; Wang, Y.F.; Tong, X.H.; Tang, J.; Li, J.L.; Wang, H.M.; Tang, L.Q.; Li, Z.Y.; Shu, Y.Z.; et al. RMS2 Encoding a GDSL Lipase Mediates Lipid Homeostasis in Anthers to Determine Rice Male Fertility. Plant Physiol. 2020, 182, 2047–2064. [Google Scholar] [CrossRef]
- Mondol, P.C.; Xu, D.W.; Duan, L.; Shi, J.X.; Wang, C.H.; Chen, X.F.; Chen, M.J.; Hu, J.P.; Liang, W.Q.; Zhang, D.B. Defective Pollen Wall 3 (DPW3), a novel alpha integrin-like protein, is required for pollen wall formation in rice. New Phytol. 2020, 225, 807–822. [Google Scholar] [CrossRef]
- Yu, B.; Liu, L.T.; Wang, T. Deficiency of very long chain alkanes biosynthesis causes humidity-sensitive male sterility via affecting pollen adhesion and hydration in rice. Plant Cell Environ. 2019, 42, 3340–3354. [Google Scholar] [CrossRef]
- Ni, E.D.; Deng, L.; Chen, H.Q.; Lin, J.W.; Ruan, J.M.; Liu, Z.L.; Zhuang, C.X.; Zhou, H. OsCER1 regulates humidity-sensitive genic male sterility through very-long-chain (VLC) alkane metabolism of tryphine in rice. Funct. Plant Biol. 2021, 48, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, G.; Liu, H.W. Enoyl-CoA hydratase: Reaction, mechanism, and inhibition. Bioorganic Med. Chem. 2003, 11, 9–20. [Google Scholar] [CrossRef]
- Krishna, A.G.G.; Hemakumar, K.H.; Khatoon, S. Study on the composition of rice bran oil and its higher free fatty acids value. J. Am. Oil Chem. Soc. 2006, 83, 117–120. [Google Scholar] [CrossRef]
- Hu, Z.L.; Li, P.; Zhou, M.Q.; Zhang, Z.H.; Wang, L.X.; Zhu, L.H.; Zhu, Y.G. Mapping of quantitative trait loci (QTLs) for rice protein and fat content using doubled haploid lines. Euphytica 2004, 135, 47–54. [Google Scholar] [CrossRef]
- Liu, W.J.; Zeng, J.; Jiang, G.H.; He, Y.Q. QTLs identification of crude fat content in brown rice and its genetic basis analysis using DH and two backcross populations. Euphytica 2009, 169, 197–205. [Google Scholar] [CrossRef]
- Qin, Y.; Kim, S.M.; Zhao, X.; Lee, H.S.; Jia, B.Y.; Kim, K.M.; Eun, M.Y.; Sohn, J.K. QTL detection and MAS selection efficiency for lipid content in brown rice (Oryza sativa L.). Genes Genom. 2010, 32, 506–512. [Google Scholar] [CrossRef]
- Ying, J.Z.; Shan, J.X.; Gao, J.P.; Zhu, M.Z.; Shi, M.; Lin, H.X. Identification of Quantitative Trait Loci for Lipid Metabolism in Rice Seeds. Mol. Plant 2012, 5, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Xia, D.; Li, P.B.; Ao, Y.T.; Xu, X.D.; Wan, S.S.; Li, Y.H.; Wu, B.; Shi, H.; Wang, K.Y.; et al. Genetic architecture and key genes controlling the diversity of oil composition in rice grains. Mol. Plant 2021, 14, 456–469. [Google Scholar] [CrossRef]
- Sun, R.; Ye, R.; Gao, L.; Zhang, L.; Wang, R.; Mao, T.; Zheng, Y.S.; Li, D.D.; Lin, Y.J. Characterization and Ectopic Expression of CoWRI1, an AP2/EREBP Domain-Containing Transcription Factor from Coconut (Cocos nucifera L.) Endosperm, Changes the Seeds Oil Content in Transgenic Arabidopsis thaliana and Rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 63. [Google Scholar] [CrossRef]
- Li, Y.T.; Han, D.X.; Hu, G.R.; Sommerfeld, M.; Hu, Q.A. Inhibition of starch synthesis results in overproduction of lipids in Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2010, 107, 258–268. [Google Scholar] [CrossRef]
- Li, D.Q.; Wu, X.B.; Wang, H.F.; Feng, X.; Yan, S.J.; Wu, S.Y.; Liu, J.X.; Yao, X.F.; Bai, A.N.; Zhao, H.; et al. Defective mitochondrial function by mutation in THICK ALEURONE 1 encoding a mitochondrion-targeted single-stranded DNA-binding protein leads to increased aleurone cell layers and improved nutrition in rice. Mol. Plant 2022, 15, 1638–1639. [Google Scholar] [CrossRef]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Ageing of stored rice: Changes in chemical and physical attributes. J. Cereal. Sci. 2002, 35, 65–78. [Google Scholar] [CrossRef]
- Stoutjesdijk, P.A.; Singh, S.P.; Liu, Q.; Hurlstone, C.J.; Waterhouse, P.A.; Green, A.G. hpRNA-mediated targeting of the Arabidopsis FAD2 gene gives highly efficient and stable silencing. Plant Physiol. 2002, 129, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Singh, S.P.; Green, A.G. High-stearic and high-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing. Plant Physiol. 2002, 129, 1732–1743. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Yin, Z.J.; Xiao, L.; Xu, Y.N.; Qu, L.Q. Identification and evaluation of ω-3 fatty acid desaturase genes for hyperfortifying α-linolenic acid in transgenic rice seed. J. Exp. Bot. 2012, 63, 3279–3287. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Ise, K.; Li, C.Y.; Honda, I.; Iwai, Y.; Matsukura, U. Volatile components in stored rice [Oryza sativa (L.)] of varieties with and without lipoxygenase-3 in seeds. J. Agric. Food Chem. 1999, 47, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Gayen, D.; Ali, N.; Sarkar, S.N.; Datta, S.K.; Datta, K. Down-regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta 2015, 242, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Kerstiens, G. Water transport in plant cuticles: An update. J. Exp. Bot. 2006, 57, 2493–2499. [Google Scholar] [CrossRef]
- Jenks, M.A.; Tuttle, H.A.; Eigenbrode, S.D.; Feldmann, K.A. Leaf Epicuticular Waxes of the Eceriferum Mutants in Arabidopsis. Plant Physiol. 1995, 108, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Du, H.; Ning, J.; Ye, H.Y.; Xiong, L.Z. Characterization of Glossy1-homologous genes in rice involved in leaf wax accumulation and drought resistance. Plant Mol. Biol. 2009, 70, 443–456. [Google Scholar] [CrossRef]
- Qin, B.X.; Tang, D.; Huang, J.; Li, M.; Wu, X.R.; Lu, L.L.; Wang, K.J.; Yu, H.X.; Chen, J.M.; Gu, M.H.; et al. Rice OsGL1-1 is involved in leaf cuticular wax and cuticle membrane. Mol. Plant 2011, 4, 985–995. [Google Scholar] [CrossRef]
- Zhou, X.Y.; Li, L.Z.; Xiang, J.H.; Gao, G.F.; Xu, F.X.; Liu, A.L.; Zhang, X.W.; Peng, Y.; Chen, X.B.; Wan, X.Y. OsGL1-3 is Involved in Cuticular Wax Biosynthesis and Tolerance to Water Deficit in Rice. PLoS ONE 2015, 10, e116676. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.Y.; Ni, E.D.; Yang, J.W.; Zhou, H.; Liang, H.; Li, J.; Jiang, D.G.; Wang, Z.H.; Liu, Z.L.; Zhuang, C.X. Rice OsGL1-6 is involved in leaf cuticular wax accumulation and drought resistance. PLoS ONE 2013, 8, e65139. [Google Scholar] [CrossRef] [PubMed]
- Lolle, S.J.; Hsu, W.; Pruitt, R.E. Genetic analysis of organ fusion in. Genetics 1998, 149, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.H.; Li, S.B.; He, S.; Wassmann, F.; Yu, C.H.; Qin, G.J.; Schreiber, L.; Qu, L.J.; Gu, H.Y. CFL1, a WW Domain Protein, Regulates Cuticle Development by Modulating the Function of HDG1, a Class IV Homeodomain Transcription Factor, in Rice and Arabidopsi. Plant Cell 2011, 23, 3392–3411. [Google Scholar] [CrossRef] [PubMed]
- Mawlong, I.; Ali, K.; Kurup, D.; Yadav, S.; Tyagi, A. Isolation and characterization of an AP2/ERF-type drought stress inducible transcription factor encoding gene from rice. J. Plant Biochem. Biotechnol. 2014, 23, 42–51. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Xiong, L.Z. Putative megaenzyme DWA1 plays essential roles in drought resistance by regulating stress-induced wax deposition in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 17790–17795. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.H.; Yu, J.J.; Liao, B.; Shan, J.X.; Ye, W.W.; Dong, N.Q.; Guo, T.; Kan, Y.; Zhang, H.; Yang, Y.B.; et al. An a/f3 hydrolase family member negatively regulates salt tolerance but promotes flowering through three distinct functions in rice. Mol. Plant 2022, 15, 1908–1930. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Li, X.M.; Lin, H.X.; Chong, K. Crop Improvement Through Temperature Resilience. Annu. Rev. Plant Biol. 2019, 70, 753–780. [Google Scholar] [CrossRef] [PubMed]
- Kan, Y.; Mu, X.R.; Zhang, H.; Gao, J.; Shan, J.X.; Ye, W.W.; Lin, H.X. TT2 controls rice thermotolerance through SCT1-dependent alteration of wax biosynthesis. Nat. Plants 2022, 8, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Mongrand, S.; Stanislas, T.; Bayer, E.M.F.; Lherminier, J.; Simon-Plas, F. Membrane rafts in plant cells. Trends Plant Sci. 2010, 15, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Zhou, Z.Q.; Li, Q.; Zhai, C.; Liu, L.J.; Quilichini, T.D.; Gao, P.; Kessler, S.A.; Jaillais, Y.; Datla, R.; et al. Specific Recruitment of Phosphoinositide Species to the Plant-Pathogen Interfacial Membrane Underlies Arabidopsis Susceptibility to Fungal Infection. Plant Cell 2020, 32, 1665–1688. [Google Scholar] [CrossRef] [PubMed]
- Sha, G.; Sun, P.; Kong, X.J.; Han, X.Y.; Sun, Q.P.; Fouillen, L.; Zhao, J.; Li, Y.; Yang, L.; Wang, Y.; et al. Genome editing of a rice CDP-DAG synthase confers multipathogen resistance. Nature 2023, 618, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Su, S.Y.; Tang, P.; Zuo, R.B.; Chen, H.F.; Zhao, T.Q.; Yang, S.M.; Yang, J. Exogenous Jasmonic Acid Alleviates Blast Resistance Reduction Caused by LOX3 Knockout in Rice. Biomolecules 2023, 13, 1197. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Z.; Ouyang, B.; Wang, F.; Lan, D.; Wang, Y. Lipidomics analysis of rice bran during storage unveils mechanisms behind dynamic changes in functional lipid molecular species. Food Chem. 2024, 447, 138946. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).