
IL-10 and TGF-β, but Not IL-17A or IFN-γ, Potentiate the IL-15-Induced Proliferation of Human T Cells: Association with a Decrease in the Expression of β2m-Free HLA Class I Molecules Induced by IL-15

 and
and 
Abstract
1. Introduction
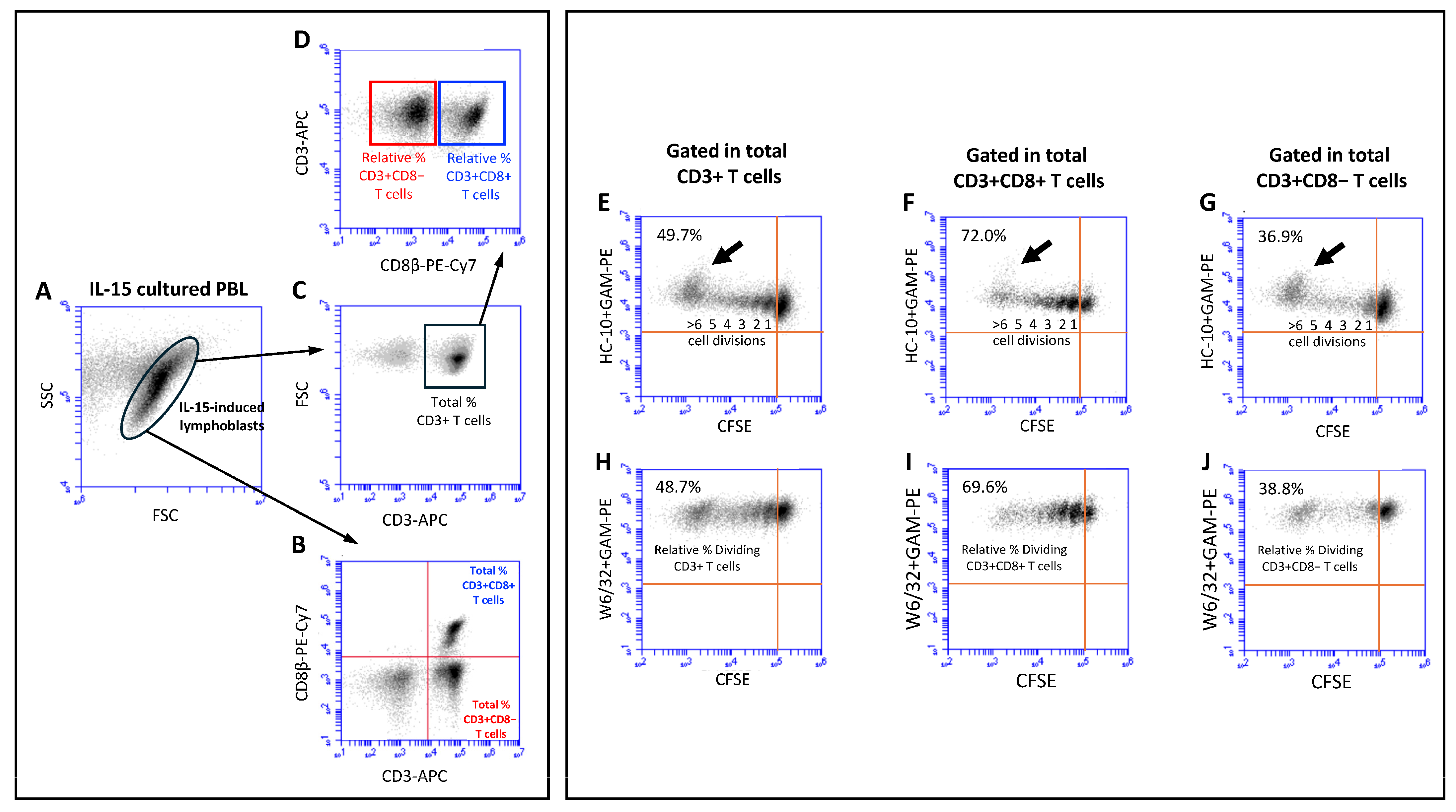
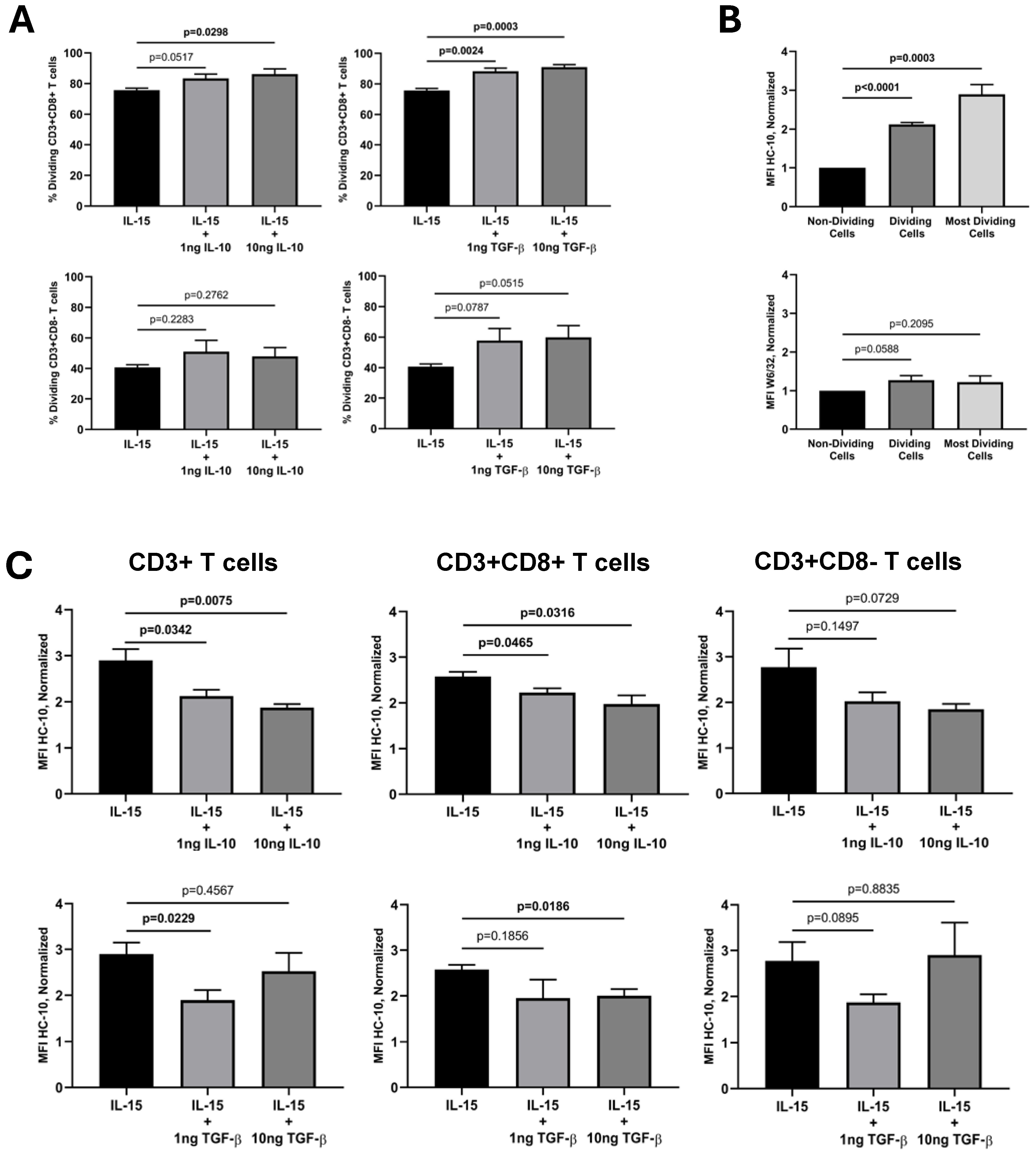
2. Results
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Cells Isolation, CFSE Labeling and Culture Conditions
4.3. Flow Cytometry Studies
4.4. Quantification of Cell Proliferation
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akdis, C.A.; Joss, A.; Akdis, M.; Faith, A.; Blaser, K. A molecular basis for T cell suppression by IL-10: CD28-associated IL-10 receptor inhibits CD28 tyrosine phosphorylation and phosphatidylinositol 3-kinase binding. FASEB J. 2000, 14, 1666–1668. [Google Scholar] [CrossRef] [PubMed]
- de Waal Malefyt, R.; Haanen, J.; Spits, H.; Roncarolo, M.G.; te Velde, A.; Figdor, C.; Johnson, K.; Kastelein, R.; Yssel, H.; de Vries, J.E. Interleukin 10 (IL-10) and viral IL-10 strongly reduce antigen-specific human T cell proliferation by diminishing the antigen-presenting capacity of monocytes via downregulation of class II major histocompatibility complex expression. J. Exp. Med. 1991, 174, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Mollo, S.B.; Harrington, L.E.; Zajac, A.J. IL-10 Regulates Memory T Cell Development and the Balance between Th1 and Follicular Th Cell Responses during an Acute Viral Infection. J. Immunol. 2016, 197, 1308–1321. [Google Scholar] [CrossRef]
- Allavena, P.; Piemonti, L.; Longoni, D.; Bernasconi, S.; Stoppacciaro, A.; Ruco, L.; Mantovani, A. IL-10 prevents the differentiation of monocytes to dendritic cells but promotes their maturation to macrophages. Eur. J. Immunol. 1998, 28, 359–369. [Google Scholar] [CrossRef]
- Coomes, S.M.; Kannan, Y.; Pelly, V.S.; Entwistle, L.J.; Guidi, R.; Perez-Lloret, J.; Nikolov, N.; Müller, W.; Wilson, M.S. CD4+ Th2 cells are directly regulated by IL-10 during allergic airway inflammation. Mucosal. Immunol. 2017, 10, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.; Gagliani, N.; Esplugues, E.; O’Connor, W., Jr.; Huber, F.J.; Chaudhry, A.; Kamanaka, M.; Kobayashi, Y.; Booth, C.J.; Rudensky, A.Y.; et al. Th17 cells express interleukin-10 receptor and are controlled by Foxp3(-) and Foxp3+ regulatory CD4+ T cells in an interleukin-10-dependent manner. Immunity 2011, 34, 554–565. [Google Scholar] [CrossRef]
- Murai, M.; Turovskaya, O.; Kim, G.; Madan, R.; Karp, C.L.; Cheroutre, H.; Kronenberg, M. Interleukin 10 acts on regulatory T cells to maintain expression of the transcription factor Foxp3 and suppressive function in mice with colitis. Nat. Immunol. 2009, 10, 1178–1184. [Google Scholar] [CrossRef]
- Cohen, S.B.A.; Katsikis, P.D.; Feldmann, M.; Londei, M. IL-10 enhances expression of the IL-2 receptor α chain on T cells. Immunology 1994, 83, 329–332. [Google Scholar] [PubMed]
- Groux, H.; Bigler, M.; de Vries, J.E.; Roncarolo, M.-G. Inhibitory and Stimulatory Effects of IL-10 on Human CD8 + T Cells. J. Immunol. 1998, 160, 3188–3193. [Google Scholar] [CrossRef] [PubMed]
- Rowbottom, A.W.; Lepper, M.A.; Garland, R.J.; Cox, C.V.; Corley, E.G. Interleukin-10-induced CD8 cell proliferation. Immunology 1999, 98, 80–89. [Google Scholar] [CrossRef]
- Gorelik, L.; Constant, S.; Flavell, R.A. Mechanism of transforming growth factor beta-induced inhibition of T helper type 1 differentiation. J. Exp. Med. 2002, 195, 1499–1505. [Google Scholar] [CrossRef]
- Chen, C.H.; Seguin-Devaux, C.; Burke, N.A.; Oriss, T.B.; Watkins, S.C.; Clipstone, N.; Ray, A. Transforming growth factor beta blocks Tec kinase phosphorylation, Ca2+ influx, and NFATc translocation causing inhibition of T cell differentiation. J. Exp. Med. 2003, 197, 1689–1699. [Google Scholar] [CrossRef]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, S.M. Conversion of peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 2003, 198, 1875–1886. [Google Scholar] [CrossRef]
- Geissmann, F.; Revy, P.; Regnault, A.; Lepelletier, Y.; Dy, M.; Brousse, N.; Amigorena, S.; Hermine, O.; Durandy, A. TGF-beta 1 prevents the noncognate maturation of human dendritic Langerhans cells. J. Immunol. 1999, 162, 4567–4575. [Google Scholar] [CrossRef] [PubMed]
- Denniston, A.K.; Kottoor, S.H.; Khan, I.; Oswal, K.; Williams, G.P.; Abbott, J.; Wallace, G.R.; Salmon, M.; Rauz, S.; Murray, P.I.; et al. Endogenous cortisol and TGF-beta in human aqueous humor contribute to ocular immune privilege by regulating dendritic cell function. J. Immunol. 2011, 186, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Lopes, J.E.; Chong, M.M.; Ivanov, I.I.; Min, R.; Victora, G.D.; Shen, Y.; Du, J.; Rubtsov, Y.P.; Rudensky, A.Y.; et al. TGF-beta-induced Foxp3 inhibits Th17 cell differentiation by antagonizing RORgammat function. Nature 2008, 453, 236–240. [Google Scholar] [CrossRef]
- Gagliani, N.; Amezcua Vesely, M.C.; Iseppon, A.; Brockmann, L.; Xu, H.; Palm, N.W.; de Zoete, M.R.; Licona-Limón, P.; Paiva, R.S.; Ching, T.; et al. Th17 cells transdifferentiate into regulatory T cells during resolution of inflammation. Nature 2015, 523, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.A.; Massagué, J. TGF-beta directly targets cytotoxic T cell functions during tumor evasion of immune surveillance. Cancer Cell. 2005, 8, 369–380. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Schoenborn, J.R.; Wilson, C.B. Regulation of Interferon-γ During Innate and Adaptive Immune Responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar] [CrossRef]
- Siegel, J.P. Effects of interferon-gamma on the activation of human T lymphocytes. Cell Immunol. 1988, 111, 461–472. [Google Scholar] [CrossRef]
- Berry, S.P.D.; Dossou, C.; Kashif, A.; Sharifinejad, N.; Azizi, G.; Hamedifar, H.; Sabzvari, A.; Zian, Z. The role of IL-17 and anti-IL-17 agents in the immunopathogenesis and management of autoimmune and inflammatory diseases. Int. Immunopharmacol. 2022, 102, 108402. [Google Scholar] [CrossRef] [PubMed]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.L.; Hooibrink, B.; Arosa, F.A.; van Lier, R.A. IL-15 induces antigen-independent expansion and differentiation of human naive CD8+ T cells in vitro. Blood 2003, 102, 2541–2546. [Google Scholar] [CrossRef]
- Correia, M.P.; Cardoso, E.M.; Pereira, C.F.; Neves, R.; Uhrberg, M.; Arosa, F.A. Hepatocytes and IL-15: A favorable microenvironment for T cell survival and CD8+ T cell differentiation. J. Immunol. 2009, 182, 6149–6159. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.P.; Costa, A.V.; Uhrberg, M.; Cardoso, E.M.; Arosa, F.A. IL-15 induces CD8+ T cells to acquire functional NK receptors capable of modulating cytotoxicity and cytokine secretion. Immunobiology 2011, 216, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.G.; Powis, S.J.; Arosa, F.A. Misfolding of major histocompatibility complex class I molecules in activated T cells allows cis-interactions with receptors and signaling molecules and is associated with tyrosine phosphorylation. J. Biol. Chem. 2004, 279, 53062–53070. [Google Scholar] [CrossRef]
- Arosa, F.A.; Santos, S.G.; Powis, S.J. Open conformers: The hidden face of MHC-I molecules. Trends Immunol. 2007, 28, 115–123. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Filippone, E.J.; Amato-Menker, C.J.; Arosa, F.A.; Das, B.; Ou, Y.; Norin, A.J. Antibodies to cryptic epitopes on HLA class I and class II heavy chains bound to single antigen beads: Clinically relevant? Transpl. Immunol. 2021, 69, 101482. [Google Scholar] [CrossRef]
- Hudson, L.E.; Allen, R. Leukocyte Ig-Like Receptors—A Model for MHC Class I Disease Associations. Front. Immunol. 2016, 7, 281. [Google Scholar] [CrossRef]
- Smith, C.; Santi, M.; Rajan, B.; Rushing, E.J.; Choi, M.R.; Rood, B.R.; Cornelison, R.; MacDonald, T.J.; Vukmanovic, S. A novel role of HLA class I in the pathology of medulloblastoma. J. Transl. Med. 2009, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, E.M.; Esgalhado, A.J.; Patrão, L.; Santos, M.; Neves, V.P.; Martinez, J.; Patto, M.A.V.; Silva, H.; Arosa, F.A. Distinctive CD8+ T cell and MHC class I signatures in polycythemia vera patients. Ann. Hematol. 2018, 97, 1563–1575. [Google Scholar] [CrossRef]
- Arosa, F.A.; Esgalhado, A.J.; Reste-Ferreira, D.; Cardoso, E.M. Open MHC Class I Conformers: A Look through the Looking Glass. Int. J. Mol. Sci. 2021, 22, 9738. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, T.S.; Chakrabarti, A.; Edidin, M. Interaction of class I human leukocyte antigen (HLA-I) molecules with insulin receptors and its effect on the insulin-signaling cascade. Mol. Biol. Cell. 1997, 8, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.G.; Antoniou, A.N.; Sampaio, P.; Powis, S.J.; Arosa, F.A. Lack of tyrosine 320 impairs spontaneous endocytosis and enhances release of HLA-B27 molecules. J. Immunol. 2006, 176, 2942–2949. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Bieri, G.; Gontier, G.; Müller, S.; Smith, L.K.; Snethlage, C.E.; White CW 3rd Maybury-Lewis, S.Y.; Villeda, S.A. MHC class I H2-Kb negatively regulates neural progenitor cell proliferation by inhibiting FGFR signaling. PLoS Biol. 2021, 19, e3001311. [Google Scholar] [CrossRef]
- Esgalhado, A.J.; Reste-Ferreira, D.; Weinhold, S.; Uhrberg, M.; Cardoso, E.M.; Arosa, F.A. In vitro IL-15-activated human naïve CD8+ T cells down-modulate the CD8β chain and become CD8αα T cells. Front. Immunol. 2024, 15, 1252439. [Google Scholar] [CrossRef]
- Alspach, E.; Lussier, E.D.M.; Schreiber, R.D. Interferon γ and its important roles in promoting and inhibiting spontaneous and therapeutic cancer immunity. Cold Spring Harb. Perspect. Biol. 2019, 11, a028480. [Google Scholar] [CrossRef]
- Czaja, A.J. Immune Inhibitory Properties and Therapeutic Prospects of Transforming Growth Factor-Beta and Interleukin 10 in Autoimmune Hepatitis. Dig. Dis. Sci. 2022, 67, 1163–1186. [Google Scholar] [CrossRef]
- Zannikou, M.; Fish, E.N.; Platanias, L.C. Signaling by Type I Interferons in Immune Cells: Disease Consequences. Cancers 2024, 16, 1600. [Google Scholar] [CrossRef]
- Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar] [CrossRef] [PubMed]
- Itatani, Y.; Kawada, K.; Sakai, Y. Transforming Growth Factor-β Signaling Pathway in Colorectal Cancer and Its Tumor Microenvironment. Int. J. Mol. Sci. 2019, 20, 5822. [Google Scholar] [CrossRef] [PubMed]
- Papadakis, K.A.; Krempski, J.; Reiter, J.; Svingen, P.; Xiong, Y.; Sarmento, O.F.; Huseby, A.; Johnson, A.J.; Lomberk, G.A.; Urrutia, R.A.; et al. Krüppel-like factor KLF10 regulates transforming growth factor receptor II expression and TGF-β signaling in CD8+ T lymphocytes. Am. J. Physiol. Cell Physiol. 2015, 308, C362–C371. [Google Scholar] [CrossRef] [PubMed]
- Weitzmann, M.N.; Ofotokun, I. Physiological and pathophysiological bone turnover—role of the immune system. Nat. Rev. Endocrinol. 2016, 12, 518–532. [Google Scholar] [CrossRef]
- Arosa, F.A.; Esgalhado, A.J.; Padrão, C.A.; Cardoso, E.M. Divide, Conquer, and Sense: CD8+CD28- T Cells in Perspective. Front. Immunol. 2017, 7, 665. [Google Scholar] [CrossRef]
- Sheng, H.; Marrero, I.; Maricic, I.; Fanchiang, S.S.; Zhang, S.; Sant’Angelo, D.B.; Kumar, K. Distinct PLZF+CD8αα+ Unconventional T cells enriched in liver use a cytotoxic mechanism to limit autoimmunity. J. Immunol. 2019, 203, 2150–2162. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Cytokines | Exp. | Absolute W6/32 MFI Values (auf) | Absolute HC-10 MFI Values (auf) | ||||
|---|---|---|---|---|---|---|---|
| NDC | DC | MDC | NDC | DC | MDC | ||
| IL-15 | 1 | 582,286 | 670,306 | 645,073 | 2434 | 3867 | 7855 |
| 2 | 279,225 | 597,143 | 473,002 | 4670 | 9595 | 10,396 | |
| 3 | 308,030 | 328,094 | 348,512 | 6624 | 14,539 | 19,520 | |
| 4 | 369,864 | 394,137 | 383,243 | 12,406 | 25,252 | 40,912 | |
| Mean ± SEM | 384,851 ± 68,474 | 497,420 ± 81,226 | 462,458 ± 66,281 | 6534 ± 2136 | 13,313 ± 4538 | 19,671 ± 7510 | |
| CV | 35.6 | 32.7 | 28.7 | 65.4 | 68.2 | 76.4 | |
| +1 ng IL-10 | 1 | 690,499 | 696,209 | 592,428 | 2563 | 4973 | 4730 |
| 2 | 313,580 | 577,480 | 534,129 | 5380 | 10,577 | 10,729 | |
| 3 | 299,515 | 410,883 | 438,325 | 6835 | 12,862 | 15,825 | |
| 4 | 531,147 | 611,837 | 628,632 | 12,310 | 18,768 | 29,602 | |
| Mean ± SEM | 458,685 ± 93,710 | 574,102 ± 59,851 | 548,379 ± 41,529 | 6772 ± 2048 | 11,795 ± 2855 | 15,222 ± 5303 | |
| CV | 40.9 | 20.9 | 15.21 | 60.5 | 48.4 | 69.7 | |
| +10 ng IL-10 | 1 | 561,224 | 696,209 | 609,248 | 2324 | 3548 | 4292 |
| 2 | 280,539 | 479,845 | 542,653 | 5251 | 9570 | 9474 | |
| 3 | 320,450 | 353,883 | 498,150 | 7532 | 10,370 | 16,433 | |
| 4 | 488,084 | 563,177 | 537,863 | 14,953 | 20,244 | 27,516 | |
| Mean ± SEM | 412,574 ± 66,907 | 523,279 ± 71,925 | 546,979 ± 23,028 | 7515 ± 2699 | 10,933 ± 3457 | 14,429 ± 5022 | |
| CV | 32.4 | 27.5 | 8.4 | 71.8 | 63.2 | 69.6 | |
| +1 ng TGF-β | 1 | 506,734 | 350,418 | 615,149 | 4153 | 4094 | 6223 |
| 2 | 285,272 | 481,154 | 525,100 | 7249 | 9929 | 12,392 | |
| 3 | 273,987 | 479,845 | 383,225 | 7657 | 9590 | 19,352 | |
| 4 | 305,044 | 432,657 | 445,050 | 11,863 | 16,577 | 22,266 | |
| Mean ± SEM | 342,759 ± 55,034 | 436,019 ± 30,682 | 492,131 ± 50,247 * | 7731 ± 1584 | 10,048 ± 2554 | 15,058 ± 3600 * | |
| CV | 32.1 | 14.1 | 20.4 | 41.0 | 50.8 | 47.8 | |
| +10 ng TGF-β | 1 | 449,394 | 497,858 | 596,349 | 2870 | 3165 | 6614 |
| 2 | 240,816 | 473,261 | 491,321 | 5692 | 9772 | 12,335 | |
| 3 | 260,754 | 364,314 | 436,399 | 6025 | 10,912 | 22,536 | |
| 4 | 275,224 | 490,359 | 490,673 | 9598 | 15,586 | 18,287 | |
| Mean ± SEM | 306,547 ± 48,135 | 456,448 ± 31,140 | 503,686 ± 33,462 * | 6046 ± 1379 | 9859 ± 2561 | 14,943 ± 3476 * | |
| CV | 31.4 | 13.6 | 13.3 | 45.6 | 52.0 | 46.5 | |
| +1 ng IL-17A | 1 | 450,968 | 915,144 | 494,647 | 5152 | 7059 | 8190 |
| 2 | 346,997 | 570,424 | 643,442 | 4946 | 7998 | 12,446 | |
| 3 | 336,243 | 433,139 | 425,421 | 6515 | 15,365 | 18,809 | |
| 4 | 441,588 | 416,545 | 486,243 | 16399 | 24,189 | 36,747 | |
| Mean ± SEM | 393,949 ± 30,352 | 583,813 ± 115,701 | 512,438 ± 46,311 | 8253 ± 2738 | 13,653 ± 3973 | 19,048 ± 6290 | |
| CV | 15.4 | 39.6 | 18.1 | 66.3 | 58.2 | 66.0 | |
| +10 ng IL-17A | 1 | 830,854 | 934,290 | 863,447 | 3646 | 4992 | 8420 |
| 2 | 344,445 | 590,201 | 630,329 | 4392 | 8418 | 11,873 | |
| 3 | 394,027 | 494,165 | 477,729 | 7869 | 15,631 | 13,492 | |
| 4 | 478,755 | 574,801 | 532,176 | 15043 | 25,087 | 40,330 | |
| Mean ± SEM | 512,020 ± 109,835 | 648,364 ± 97,607 | 625,920 ± 85,238 | 7738 ± 2603 | 13,532 ± 4444 | 18,529 ± 7344 | |
| CV | 42.9 | 30.1 | 27.2 | 67.3 | 65.7 | 79.3 | |
| +1 ng IFN-g | 1 | 631,849 | 636,441 | 596,433 | 3291 | 5212 | 5805 |
| 2 | 371,853 | 597,143 | 641,127 | 6061 | 8127 | 7446 | |
| 3 | 384,973 | 527,285 | 498,277 | 5927 | 15,056 | 19,497 | |
| 4 | 409,159 | 461,234 | 451,065 | 12,310 | 19,702 | 26,071 | |
| Mean ± SEM | 449,459 ± 61,286 | 555,526 ± 38,695 | 546,726 ± 43,666 | 6897 ± 1914 | 12,024 ± 3288 | 14,705 ± 4865 | |
| CV | 27.3 | 13.9 | 16.0 | 55.5 | 54.7 | 66.2 | |
| +10 ng IFN-g | 1 | 757,165 | 763,603 | 748,453 | 3381 | 5834 | 8353 |
| 2 | 390,485 | 640,182 | 606,920 | 5970 | 8136 | 7781 | |
| 3 | 412,008 | 563,646 | 539,970 | 5932 | 13,927 | 18,633 | |
| 4 | 394,570 | 462,345 | 414,055 | 13,533 | 19,158 | 23,188 | |
| Mean ± SEM | 488,557 ± 89,658 | 607,444 ± 63,528 | 577,350 ± 69,650 | 7204 ± 2195 | 11,764 ± 2996 | 14,489 ± 3824 | |
| CV | 36.7 | 20.9 | 24.1 | 60.9 | 50.9 | 52.8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, L.H.; Peixoto, H.A.; Cardoso, E.M.; Esgalhado, A.J.; Arosa, F.A. IL-10 and TGF-β, but Not IL-17A or IFN-γ, Potentiate the IL-15-Induced Proliferation of Human T Cells: Association with a Decrease in the Expression of β2m-Free HLA Class I Molecules Induced by IL-15. Int. J. Mol. Sci. 2024, 25, 9376. https://doi.org/10.3390/ijms25179376
Duarte LH, Peixoto HA, Cardoso EM, Esgalhado AJ, Arosa FA. IL-10 and TGF-β, but Not IL-17A or IFN-γ, Potentiate the IL-15-Induced Proliferation of Human T Cells: Association with a Decrease in the Expression of β2m-Free HLA Class I Molecules Induced by IL-15. International Journal of Molecular Sciences. 2024; 25(17):9376. https://doi.org/10.3390/ijms25179376
Chicago/Turabian StyleDuarte, Leila H., Hugo A. Peixoto, Elsa M. Cardoso, André J. Esgalhado, and Fernando A. Arosa. 2024. "IL-10 and TGF-β, but Not IL-17A or IFN-γ, Potentiate the IL-15-Induced Proliferation of Human T Cells: Association with a Decrease in the Expression of β2m-Free HLA Class I Molecules Induced by IL-15" International Journal of Molecular Sciences 25, no. 17: 9376. https://doi.org/10.3390/ijms25179376
APA StyleDuarte, L. H., Peixoto, H. A., Cardoso, E. M., Esgalhado, A. J., & Arosa, F. A. (2024). IL-10 and TGF-β, but Not IL-17A or IFN-γ, Potentiate the IL-15-Induced Proliferation of Human T Cells: Association with a Decrease in the Expression of β2m-Free HLA Class I Molecules Induced by IL-15. International Journal of Molecular Sciences, 25(17), 9376. https://doi.org/10.3390/ijms25179376