
Unraveling the Role of RNase L Knockout in Alleviating Immune Response Activation in Mice Bone Marrow after Irradiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
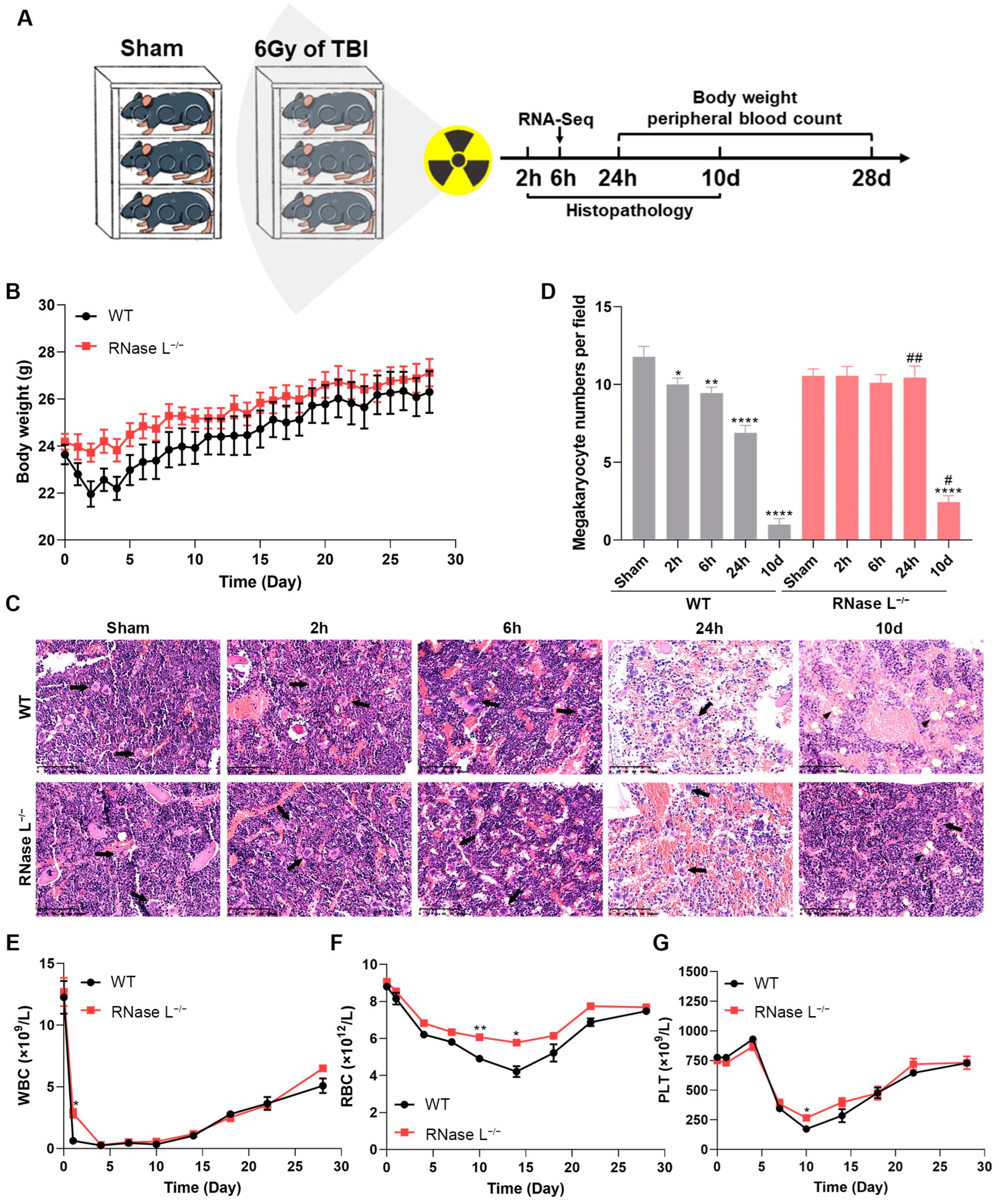
2.1. Irradiation-Induced Body Injury Is Partially Relieved in RNase L−/− Mice
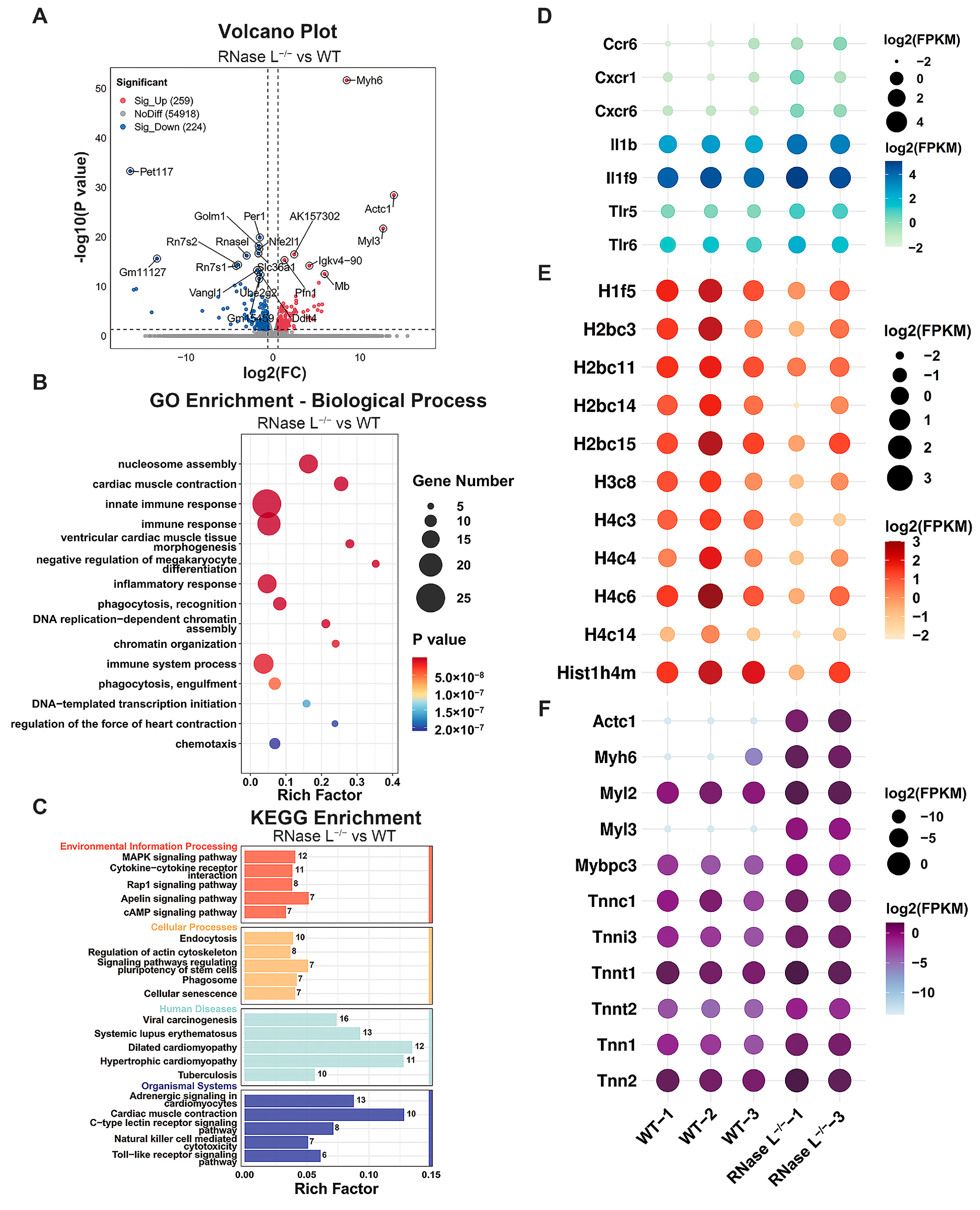
2.2. Effect of RNase L Knockout on Gene Expression in Bone Marrow Cells
2.3. The Expression of Rnase L and Oas Family Genes Is Upregulated after Irradiation
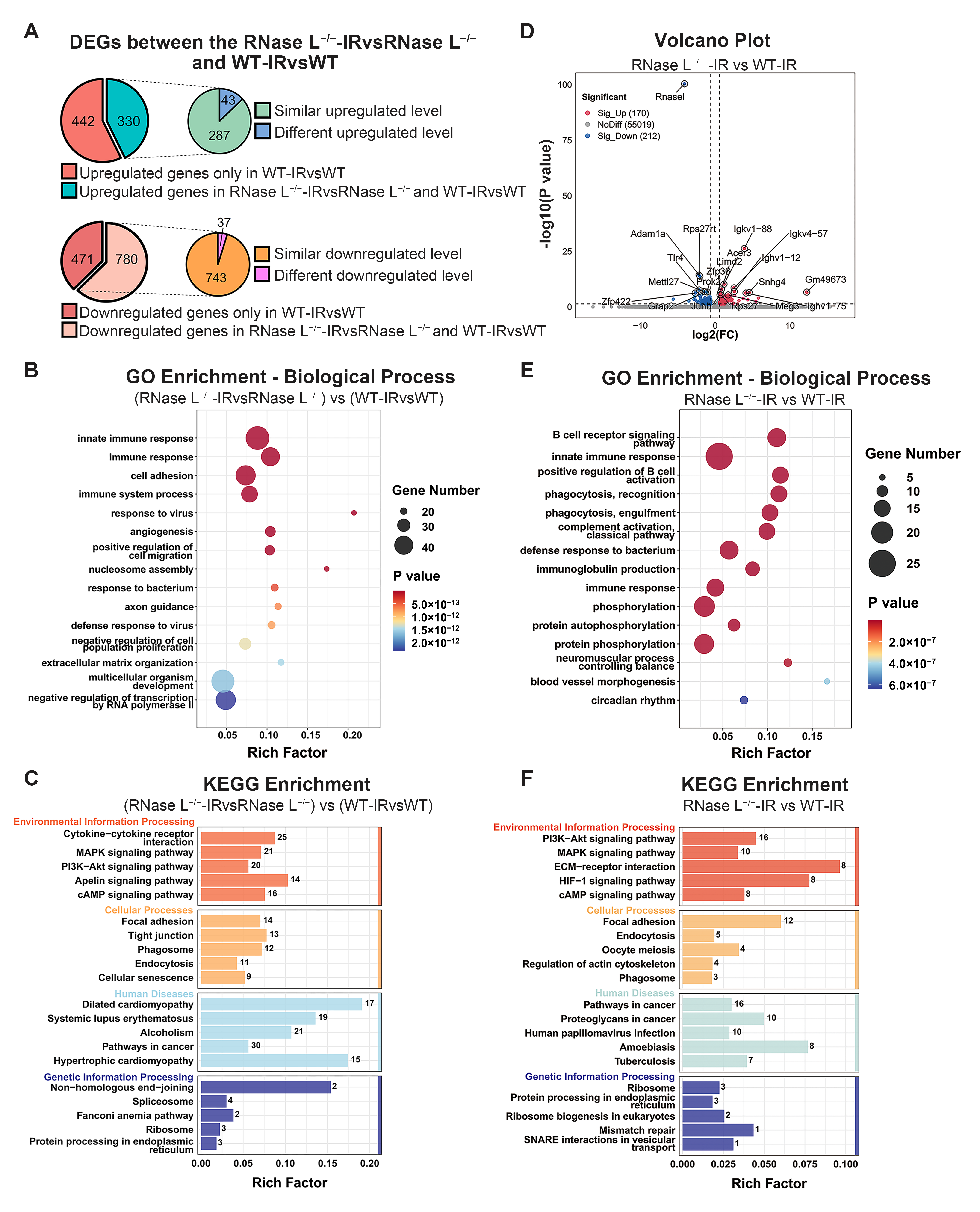
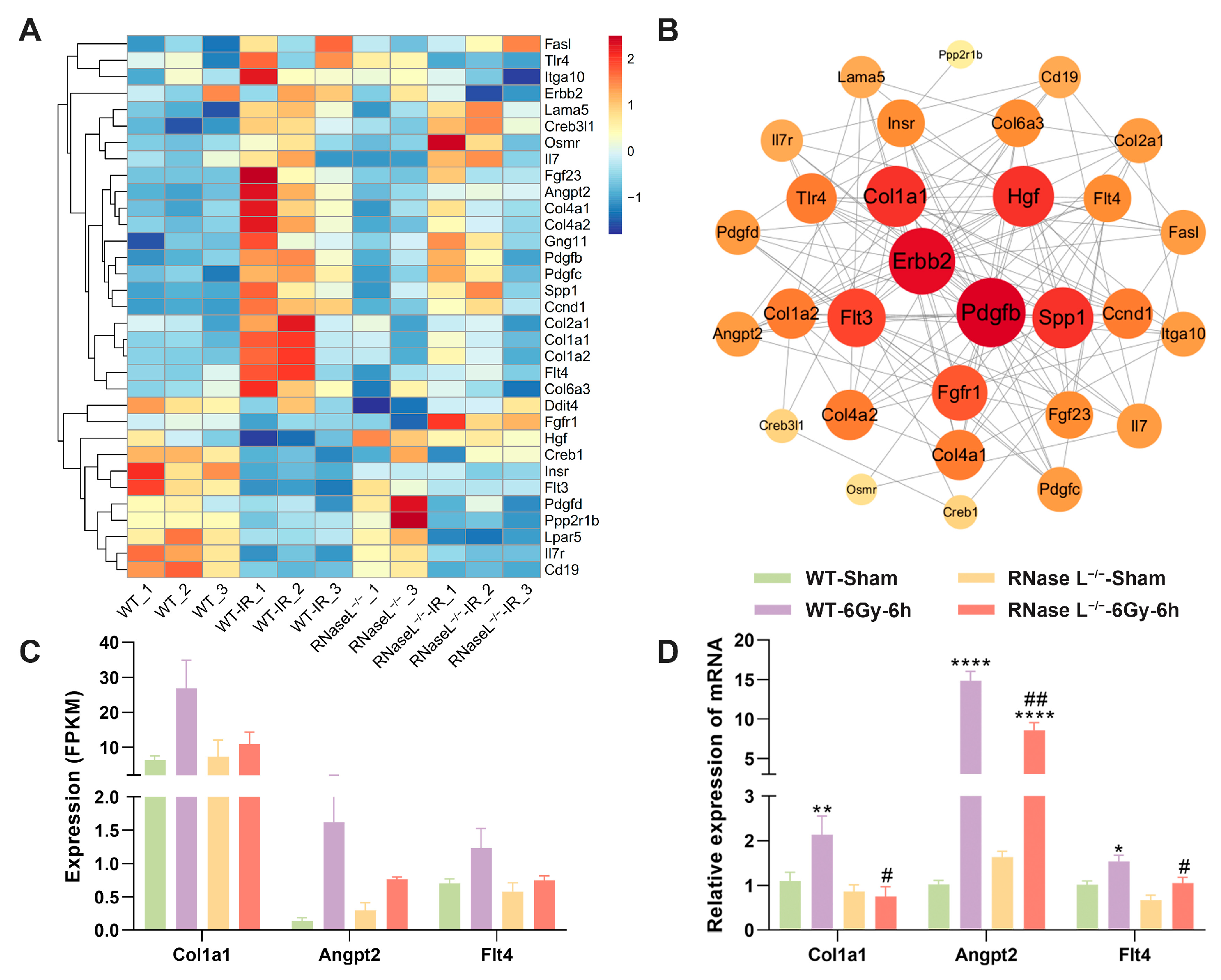
2.4. Effect of RNase L Knockout on Gene Expression in Bone Marrow Cells after Irradiation
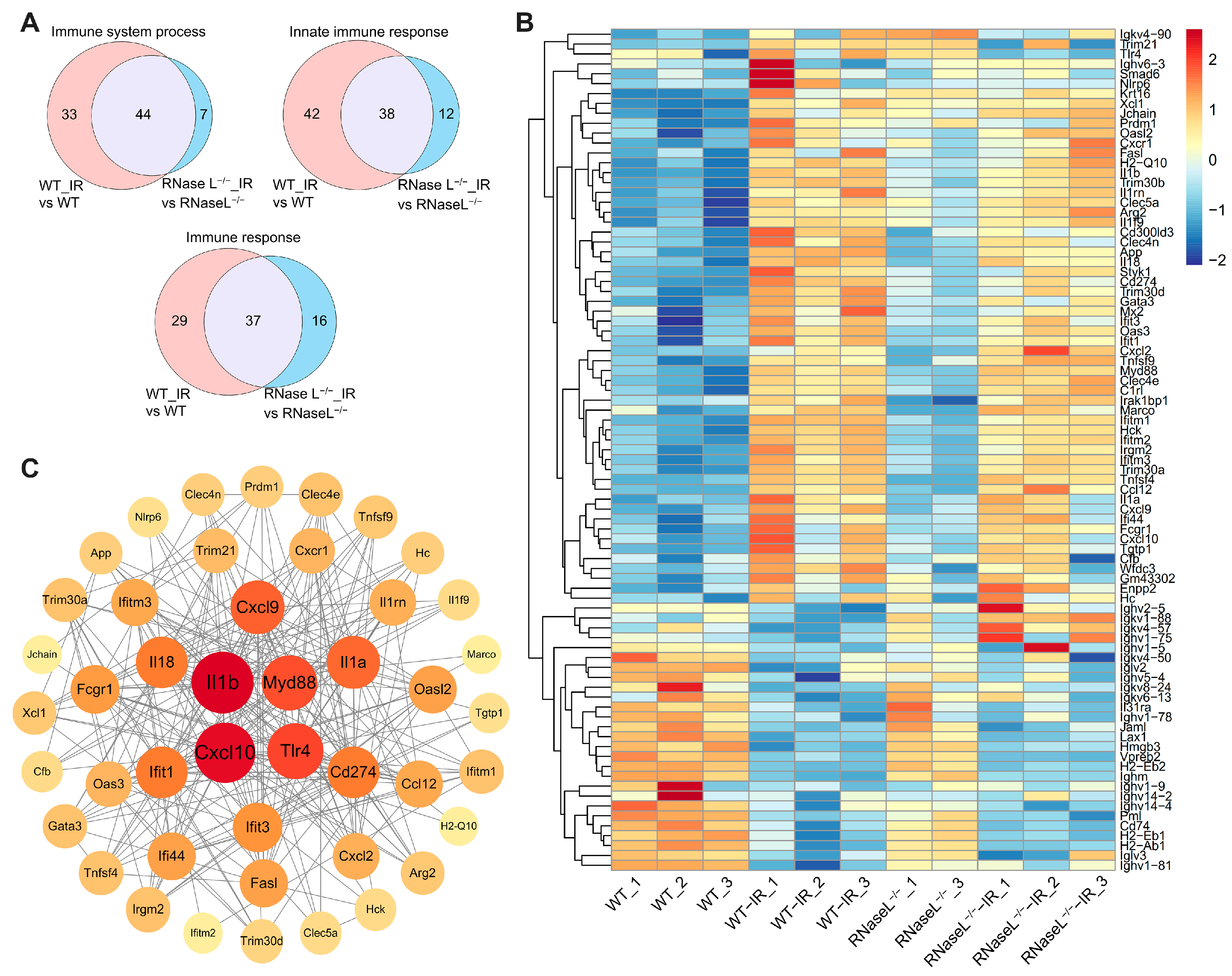
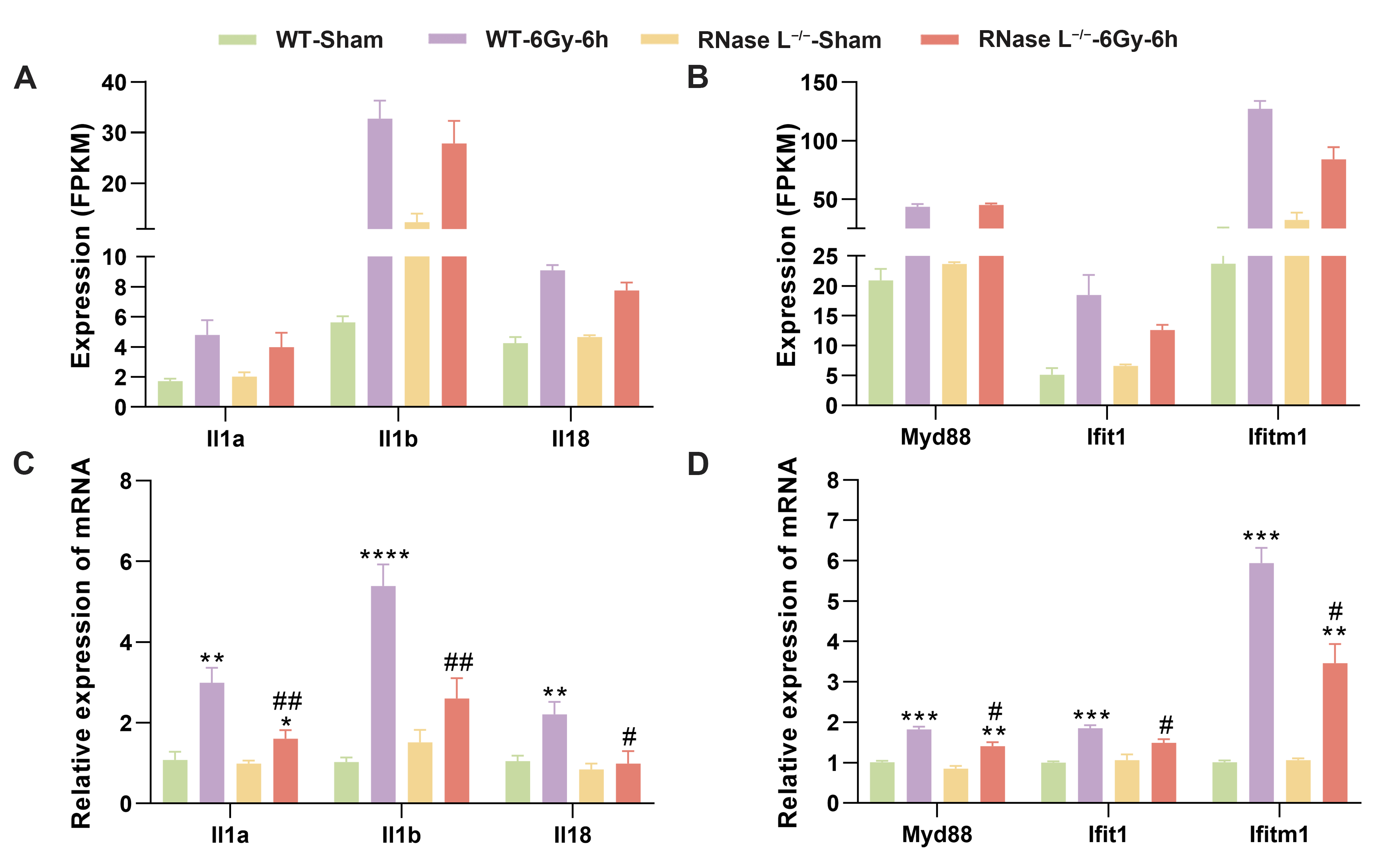
2.5. RNase L Deficiency Inhibits the Upregulation of Immune Response-Related Genes after Irradiation
2.6. RNase L Regulates Irradiation-Induced Immunoglobulins Expression
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Irradiation Exposure and Bone Marrow Extraction
4.3. Bone Marrow Histology and Complete Blood Count
4.4. RNA Extraction and RNA Sequencing (RNA-Seq)
4.5. Bioinformatic Analysis
4.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Validation Analysis
4.7. Serum Immunoglobulins Level Analysis by Cytometric Bead Array (CBA)
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shao, L.; Luo, Y.; Zhou, D. Hematopoietic stem cell injury induced by ionizing radiation. Antioxid. Redox Signal. 2014, 20, 1447–1462. [Google Scholar] [CrossRef]
- Zong, L.; Gao, Z.; Xie, W.; Tong, J.; Cao, Y. Role of NF-κB activation in mouse bone marrow stromal cells exposed to 900-MHz radiofrequency fields (RF). J. Toxicol. Environ. Health A 2019, 82, 46–51. [Google Scholar] [CrossRef]
- Hladik, D.; Tapio, S. Effects of ionizing radiation on the mammalian brain. Mutat. Res. Rev. Mutat. Res. 2016, 770, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, S.; Rana, J.N. Microwave radiation and the brain: Mechanisms, current status, and future prospects. Int. J. Mol. Sci. 2022, 23, 9288. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jenrow, K.A.; Brown, S.L. Mechanisms of radiation-induced normal tissue toxicity and implications for future clinical trials. Radiat. Oncol. J. 2014, 32, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Olabisi, A.O. Radiation: A poly-traumatic hit leading to multi-organ injury. Cell Biosci. 2019, 9, 25. [Google Scholar] [CrossRef]
- Rios, C.I.; Cassatt, D.R.; Hollingsworth, B.A.; Satyamitra, M.M.; Tadesse, Y.S.; Taliaferro, L.P.; Winters, T.A.; DiCarlo, A.L. Commonalities between COVID-19 and radiation injury. Radiat. Res. 2021, 195, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Geng, F.; Chen, J.; Tang, S.; Azzam, E.I.; Zhang, J.; Zhang, S. Additional evidence for commonalities between COVID-19 and radiation injury: Novel insight into COVID-19 candidate drugs. Radiat. Res. 2022, 198, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, X.; Lei, X.; Xiao, X.; Jiao, T.; Ma, R.; Dong, X.; Jiang, Q.; Wang, W.; Shi, Y.; et al. Sensing of cytoplasmic chromatin by cGAS activates innate immune response in SARS-CoV-2 infection. Signal Transduct. Target. Ther. 2021, 6, 382. [Google Scholar] [CrossRef] [PubMed]
- Hayman, T.J.; Baro, M.; MacNeil, T.; Phoomak, C.; Aung, T.N.; Cui, W.; Leach, K.; Iyer, R.; Challa, S.; Sandoval-Schaefer, T.; et al. STING enhances cell death through regulation of reactive oxygen species and DNA damage. Nat. Commun. 2021, 12, 2327. [Google Scholar] [CrossRef]
- Sharma, A.; Kontodimas, K.; Bosmann, M. The MAVS immune recognition pathway in viral infection and sepsis. Antioxid. Redox Signal. 2021, 35, 1376–1392. [Google Scholar] [CrossRef]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef]
- Khodarev, N.N. Intracellular RNA sensing in mammalian cells: Role in stress response and cancer therapies. Int. Rev. Cell Mol. Biol. 2019, 344, 31–89. [Google Scholar] [CrossRef]
- Tigano, M.; Vargas, D.C.; Tremblay-Belzile, S.; Fu, Y.; Sfeir, A. Nuclear sensing of breaks in mitochondrial DNA enhances immune surveillance. Nature 2021, 591, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Hassel, B.A.; Silverman, R.H. Expression cloning of 2-5A-dependent RNAase: A uniquely regulated mediator of interferon action. Cell 1993, 72, 753–765. [Google Scholar] [CrossRef]
- Hovanessian, A.G. On the discovery of interferon-inducible, double-stranded RNA activated enzymes: The 2′–5′ oligoadenylate synthetases and the protein kinase PKR. Cytokine Growth Factor Rev. 2007, 18, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Jha, B.K.; Silverman, R.H. New insights into the role of RNase L in innate immunity. J. Interferon Cytokine Res. 2011, 31, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Malathi, K.; Dong, B.; Gale, M., Jr.; Silverman, R.H. Small self-RNA generated by RNase L amplifies antiviral innate immunity. Nature 2007, 448, 816–819. [Google Scholar] [CrossRef]
- Malathi, K.; Saito, T.; Crochet, N.; Barton, D.J.; Gale, M., Jr.; Silverman, R.H. RNase L releases a small RNA from HCV RNA that refolds into a potent PAMP. RNA 2010, 16, 2108–2119. [Google Scholar] [CrossRef]
- Malathi, K.; Paranjape, J.M.; Bulanova, E.; Shim, M.; Guenther-Johnson, J.M.; Faber, P.W.; Eling, T.E.; Williams, B.R.; Silverman, R.H. A transcriptional signaling pathway in the IFN system mediated by 2′–5′-oligoadenylate activation of RNase L. Proc. Natl. Acad. Sci. USA 2005, 102, 14533–14538. [Google Scholar] [CrossRef]
- Chen, I.Y.; Ichinohe, T. Response of host inflammasomes to viral infection. Trends Microbiol. 2015, 23, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Crouse, J.; Kalinke, U.; Oxenius, A. Regulation of antiviral T cell responses by type I interferons. Nat. Rev. Immunol. 2015, 15, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.P.; Marshall, H.D.; Laidlaw, B.J.; Staron, M.M.; Kaech, S.M.; Craft, J. Transcription factor STAT3 and type I interferons are corepressive insulators for differentiation of follicular helper and T helper 1 cells. Immunity 2014, 40, 367–377. [Google Scholar] [CrossRef]
- Rath, S.; Prangley, E.; Donovan, J.; Demarest, K.; Wingreen, N.S.; Meir, Y.; Korennykh, A. Concerted 2-5A-mediated mRNA decay and transcription reprogram protein synthesis in the dsRNA response. Mol. Cell 2019, 75, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.M.; Moon, S.L.; Matheny, T.; Parker, R. RNase L reprograms translation by widespread mRNA turnover escaped by antiviral mRNAs. Mol. Cell 2019, 75, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Dupuis-Maurin, V.; Brinza, L.; Baguet, J.; Plantamura, E.; Schicklin, S.; Chambion, S.; Macari, C.; Tomkowiak, M.; Deniaud, E.; Leverrier, Y.; et al. Overexpression of the transcription factor Sp1 activates the OAS-RNAse L-RIG-I pathway. PLoS ONE 2015, 10, e0118551. [Google Scholar] [CrossRef]
- Ma, Q.; Li, J.; Zhou, H.; Tong, W.; Chen, Y. The function of RNase L and its degradation mechanism in cardiac acute ischemic injury. Apoptosis 2020, 25, 400–411. [Google Scholar] [CrossRef]
- Gao, Y.; Vasic, R.; Song, Y.; Teng, R.; Liu, C.; Gbyli, R.; Biancon, G.; Nelakanti, R.; Lobben, K.; Kudo, E.; et al. m(6)A modification prevents formation of endogenous double-stranded RNAs and deleterious innate immune responses during hematopoietic development. Immunity 2020, 52, 1007–1021. [Google Scholar] [CrossRef]
- Wu, J.; You, Q.; Lyu, R.; Qian, Y.; Tao, H.; Zhang, F.; Cai, Y.; Jiang, N.; Zheng, N.; Chen, D.; et al. Folate metabolism negatively regulates OAS-mediated antiviral innate immunity via ADAR3/endogenous dsRNA pathway. Metabolism 2023, 143, 155526. [Google Scholar] [CrossRef]
- Kiang, J.G.; Smith, J.T.; Anderson, M.N.; Swift, J.M.; Christensen, C.L.; Gupta, P.; Balakathiresan, N.; Maheshwari, R.K. Hemorrhage exacerbates radiation effects on survival, leukocytopenia, thrombopenia, erythropenia, bone marrow cell depletion and hematopoiesis, and inflammation-associated microRNAs expression in kidney. PLoS ONE 2015, 10, e0139271. [Google Scholar] [CrossRef]
- Dainiak, N. Hematologic consequences of exposure to ionizing radiation. Exp. Hematol. 2002, 30, 513–528. [Google Scholar] [CrossRef]
- Long, T.M.; Chakrabarti, A.; Ezelle, H.J.; Brennan-Laun, S.E.; Raufman, J.P.; Polyakova, I.; Silverman, R.H.; Hassel, B.A. RNase-L deficiency exacerbates experimental colitis and colitis-associated cancer. Inflamm. Bowel Dis. 2013, 19, 1295–1305. [Google Scholar] [CrossRef]
- Bisbal, C.; Silhol, M.; Laubenthal, H.; Kaluza, T.; Carnac, G.; Milligan, L.; Le Roy, F.; Salehzada, T. The 2′–5′ oligoadenylate/RNase L/RNase L inhibitor pathway regulates both MyoD mRNA stability and muscle cell differentiation. Mol. Cell Biol. 2000, 20, 4959–4969. [Google Scholar] [CrossRef]
- Dai, J.M.; Sun, D.C.; Lin, R.X.; Yang, J.; Lou, S.; Wang, S.Q. Microarray analysis of differentially expressed genes in mouse bone marrow tissues after ionizing radiation. Int. J. Radiat. Biol. 2006, 82, 511–521. [Google Scholar] [CrossRef]
- Ezelle, H.J.; Malathi, K.; Hassel, B.A. The roles of RNase-L in antimicrobial immunity and the cytoskeleton-associated innate response. Int. J. Mol. Sci. 2016, 17, 74. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Banerjee, S.; Franchi, L.; Loo, Y.M.; Gale, M., Jr.; Núñez, G.; Silverman, R.H. RNase L activates the NLRP3 inflammasome during viral infections. Cell Host Microbe 2015, 17, 466–477. [Google Scholar] [CrossRef]
- Pinsonneault, R.L.; Vacek, P.M.; O’Neill, J.P.; Finette, B.A. Induction of V(D)J-mediated recombination of an extrachromosomal substrate following exposure to DNA-damaging agents. Environ. Mol. Mutagen. 2007, 48, 440–450. [Google Scholar] [CrossRef]
- Fisher, M.R.; Rivera-Reyes, A.; Bloch, N.B.; Schatz, D.G.; Bassing, C.H. Immature lymphocytes inhibit Rag1 and Rag2 transcription and V(D)J recombination in response to DNA double-strand breaks. J. Immunol. 2017, 198, 2943–2956. [Google Scholar] [CrossRef] [PubMed]
- Toompuu, M.; Kärblane, K.; Pata, P.; Truve, E.; Sarmiento, C. ABCE1 is essential for S phase progression in human cells. Cell Cycle 2016, 15, 1234–1247. [Google Scholar] [CrossRef]
- Ge, C.; Su, F.; Fu, H.; Wang, Y.; Tian, B.; Liu, B.; Zhu, J.; Ding, Y.; Zheng, X. RNA profiling reveals a common mechanism of histone gene downregulation and complementary effects for radioprotectants in response to ionizing radiation. Dose Response 2020, 18, 1559325820968433. [Google Scholar] [CrossRef] [PubMed]
- Hauer, M.H.; Seeber, A.; Singh, V.; Thierry, R.; Sack, R.; Amitai, A.; Kryzhanovska, M.; Eglinger, J. Histone degradation in response to DNA damage enhances chromatin dynamics and recombination rates. Nat. Struct. Mol. 2017, 24, 99–107. [Google Scholar] [CrossRef]
- Goldstein, M.; Derheimer, F.A.; Tait-Mulder, J.; Kastan, M.B. Nucleolin mediates nucleosome disruption critical for DNA double-strand break repair. Proc. Natl. Acad. Sci. USA 2013, 110, 16874–16879. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Pan, B.; Zhu, J.; Fu, H.; Zheng, X. RNase L facilitates the repair of DNA double-strand breaks through the nonhomologous end-joining pathway. FEBS Lett. 2019, 593, 1190–1200. [Google Scholar] [CrossRef] [PubMed]
- Salehzada, T.; Cambier, L.; Vu Thi, N.; Manchon, L.; Regnier, L.; Bisbal, C. Endoribonuclease L (RNase L) regulates the myogenic and adipogenic potential of myogenic cells. PLoS ONE 2009, 4, e7563. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rath, P.C. Expression of mRNA and protein-protein interaction of the antiviral endoribonuclease RNase L in mouse spleen. Int. J. Biol. Macromol. 2014, 69, 307–318. [Google Scholar] [CrossRef]
- Krause, D.; Mullins, J.M.; Penafiel, L.M.; Meister, R.; Nardone, R.M. Microwave exposure alters the expression of 2-5A-dependent RNase. Radiat. Res. 1991, 127, 164–170. [Google Scholar] [CrossRef]
- Tsai, M.H.; Cook, J.A.; Chandramouli, G.V.; DeGraff, W.; Yan, H.; Zhao, S.; Coleman, C.N.; Mitchell, J.B.; Chuang, E.Y. Gene expression profiling of breast, prostate, and glioma cells following single versus fractionated doses of radiation. Cancer Res. 2007, 67, 3845–3852. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Chen, G.; Algehainy, N.; Zeng, C.; Liu, C.; Liu, H.; Liu, W.; Stacey, D.; Zhou, A. RNase L is involved in liposaccharide-induced lung inflammation. Viruses 2020, 12, 73. [Google Scholar] [CrossRef]
- Li, D.; Swaminathan, S. Human IFIT proteins inhibit lytic replication of KSHV: A new feed-forward loop in the innate immune system. PLoS Pathog. 2019, 15, e1007609. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Cheng, W.; Qu, W.; Shao, G.; Liu, S. Antibiotic alleviates radiation-induced intestinal injury by remodeling microbiota, reducing inflammation, and inhibiting fibrosis. ACS Omega 2020, 5, 2967–2977. [Google Scholar] [CrossRef]
- Schaue, D.; McBride, W.H. Links between innate immunity and normal tissue radiobiology. Radiat. Res. 2010, 173, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Bilous, N.I.; Abramenko, I.V.; Chumak, A.A.; Dyagil, I.S.; Martina, Z.V. Stereotyped cases in Ukrainian cohort of chronic lymphocytic leukemia patients depending on the ionizing radiation exposure. Probl. Radiac. Med. Radiobiol. 2022, 27, 307–323. [Google Scholar] [CrossRef] [PubMed]
- Abramenko, I.; Bilous, N.; Chumak, A.; Davidova, E.; Kryachok, I.; Martina, Z.; Nechaev, S.; Dyagil, I.; Bazyka, D.; Bebeshko, V. Chronic lymphocytic leukemia patients exposed to ionizing radiation due to the Chernobyl NPP accident—With focus on immunoglobulin heavy chain gene analysis. Leuk. Res. 2008, 32, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Aloni-Grinstein, R.; Schwartz, D.; Rotter, V. Accumulation of wild-type p53 protein upon gamma-irradiation induces a G2 arrest-dependent immunoglobulin kappa light chain gene expression. Embo J. 1995, 14, 1392–1401. [Google Scholar] [CrossRef]
- Vriesendorp, H.M.; Halliwell, R.E.; Johnson, P.M.; Fey, T.A.; McDonough, C.M. Immunoglobulin levels in dogs after total-body irradiation and bone marrow transplantation. Transplantation 1985, 39, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Karasik, A.; Jones, G.D.; DePass, A.V.; Guydosh, N.R. Activation of the antiviral factor RNase L triggers translation of non-coding mRNA sequences. Nucleic Acids Res. 2021, 49, 6007–6026. [Google Scholar] [CrossRef]
- Preite, S.; Gomez-Rodriguez, J.; Cannons, J.L.; Schwartzberg, P.L. T and B-cell signaling in activated PI3K delta syndrome: From immunodeficiency to autoimmunity. Immunol. Rev. 2019, 291, 154–173. [Google Scholar] [CrossRef]
- Yi, X.; Zeng, C.; Liu, H.; Chen, X.; Zhang, P.; Yun, B.S.; Jin, G.; Zhou, A. Lack of RNase L attenuates macrophage functions. PLoS ONE 2013, 8, e81269. [Google Scholar] [CrossRef]
- Jaufmann, J.; Carevic, M.; Tümen, L.; Eliacik, D.; Schmitt, F.; Hartl, D.; Beer-Hammer, S. Enhanced IgG(1) -mediated antibody response towards thymus-dependent immunization in CXCR1-deficient mice. Immun. Inflamm. Dis. 2021, 9, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, S.; Fujimoto, K.; Jang, M.H.; Yang, B.G.; Jung, Y.J.; Nishiyama, M.; Sato, S.; Tsujimura, T.; Yamamoto, M.; Yokota, Y.; et al. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nat. Immunol. 2008, 9, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.; Han, F.; Ge, C.; Mao, W.; Chen, L.; Hu, H.; Chen, G.; Lang, Q.; Fang, C. OmicStudio: A composable bioinformatics cloud platform with real-time feedback that can generate high-quality graphs for publication. iMeta 2023, 2, e85. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, K.; Li, H.; Tai, F.; Duan, J.; Wang, Q.; Zhai, R.; Fu, H.; Ge, C.; Zheng, X. Unraveling the Role of RNase L Knockout in Alleviating Immune Response Activation in Mice Bone Marrow after Irradiation. Int. J. Mol. Sci. 2024, 25, 2722. https://doi.org/10.3390/ijms25052722
Ding K, Li H, Tai F, Duan J, Wang Q, Zhai R, Fu H, Ge C, Zheng X. Unraveling the Role of RNase L Knockout in Alleviating Immune Response Activation in Mice Bone Marrow after Irradiation. International Journal of Molecular Sciences. 2024; 25(5):2722. https://doi.org/10.3390/ijms25052722
Chicago/Turabian StyleDing, Kexin, Hujie Li, Fumin Tai, Junzhao Duan, Qiong Wang, Rui Zhai, Hanjiang Fu, Changhui Ge, and Xiaofei Zheng. 2024. "Unraveling the Role of RNase L Knockout in Alleviating Immune Response Activation in Mice Bone Marrow after Irradiation" International Journal of Molecular Sciences 25, no. 5: 2722. https://doi.org/10.3390/ijms25052722
APA StyleDing, K., Li, H., Tai, F., Duan, J., Wang, Q., Zhai, R., Fu, H., Ge, C., & Zheng, X. (2024). Unraveling the Role of RNase L Knockout in Alleviating Immune Response Activation in Mice Bone Marrow after Irradiation. International Journal of Molecular Sciences, 25(5), 2722. https://doi.org/10.3390/ijms25052722