
Novel Ultrastructural Insights into the Clear-Cell Carcinoma of the Pancreas: A Case Report
 , , ,
, , ,  , , and
, , and 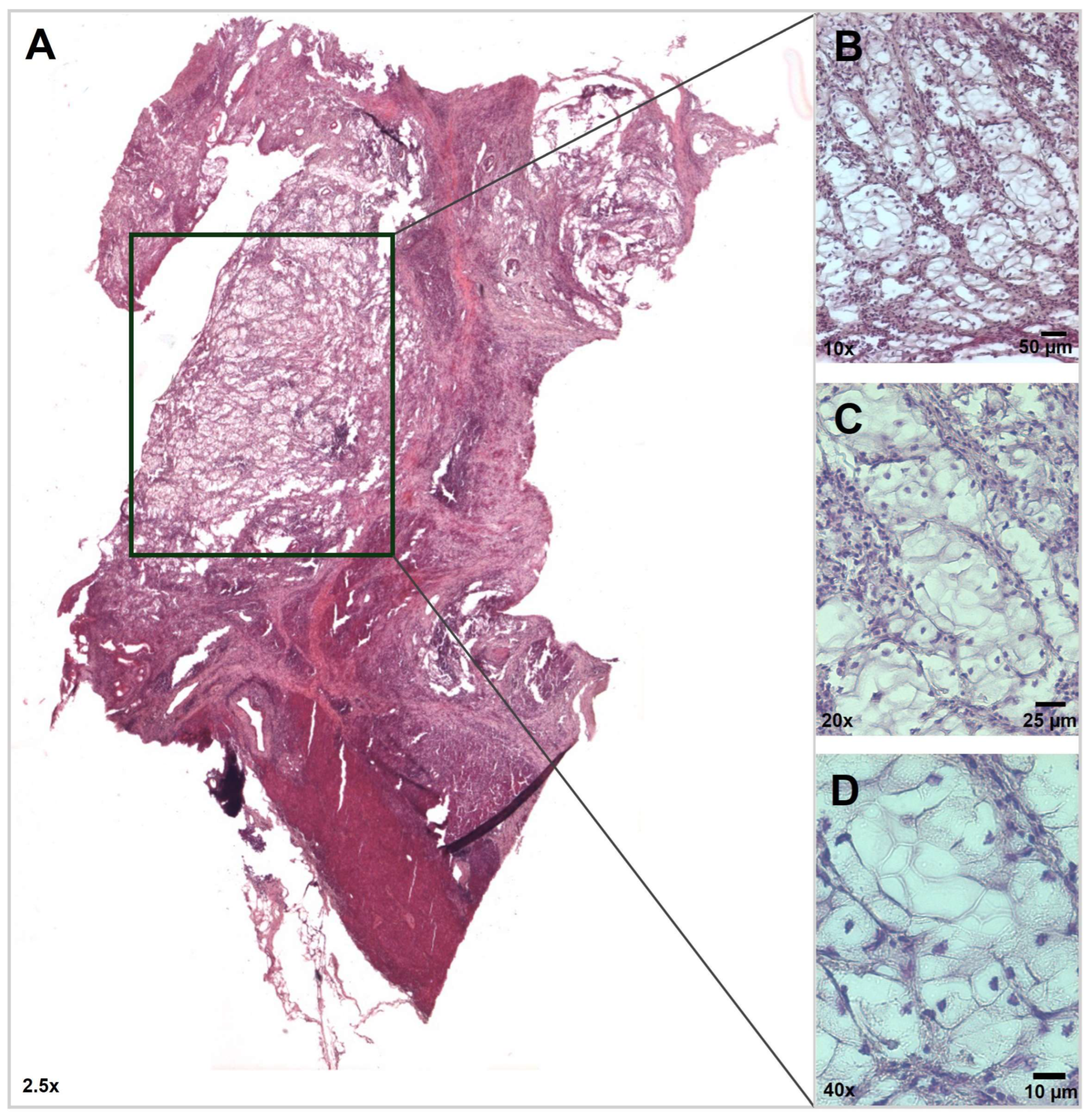{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Patient Report
3. Results
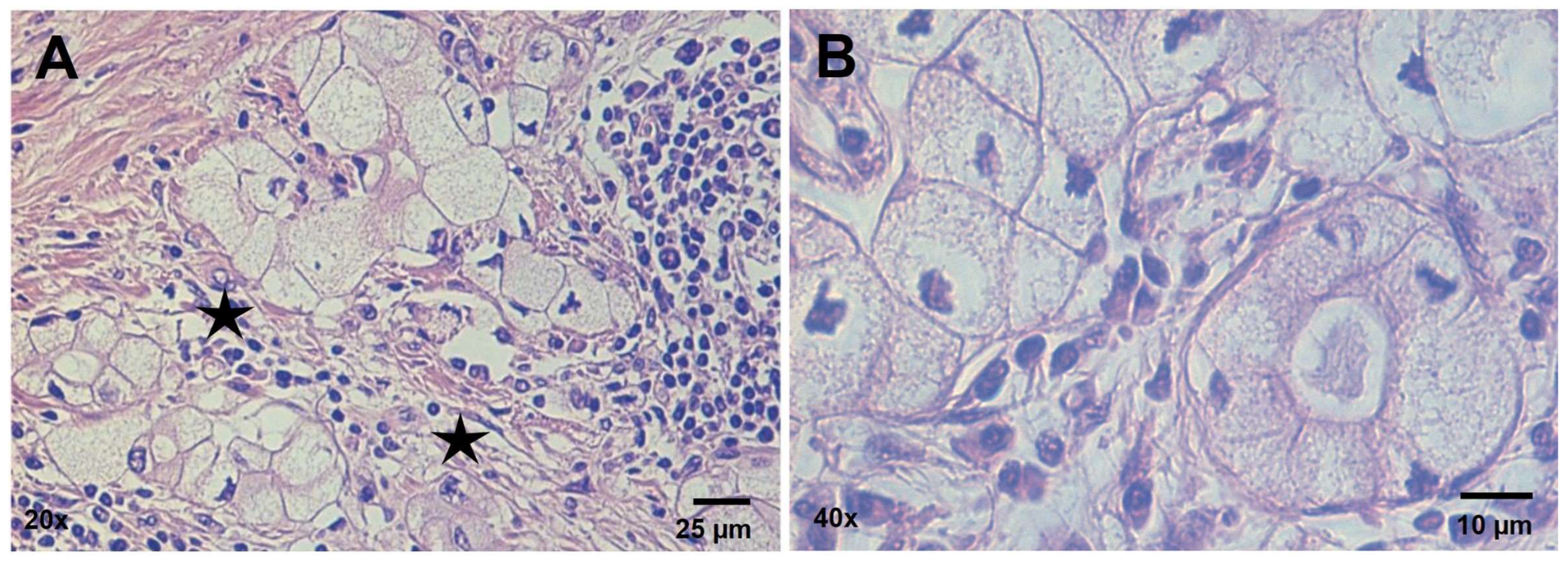
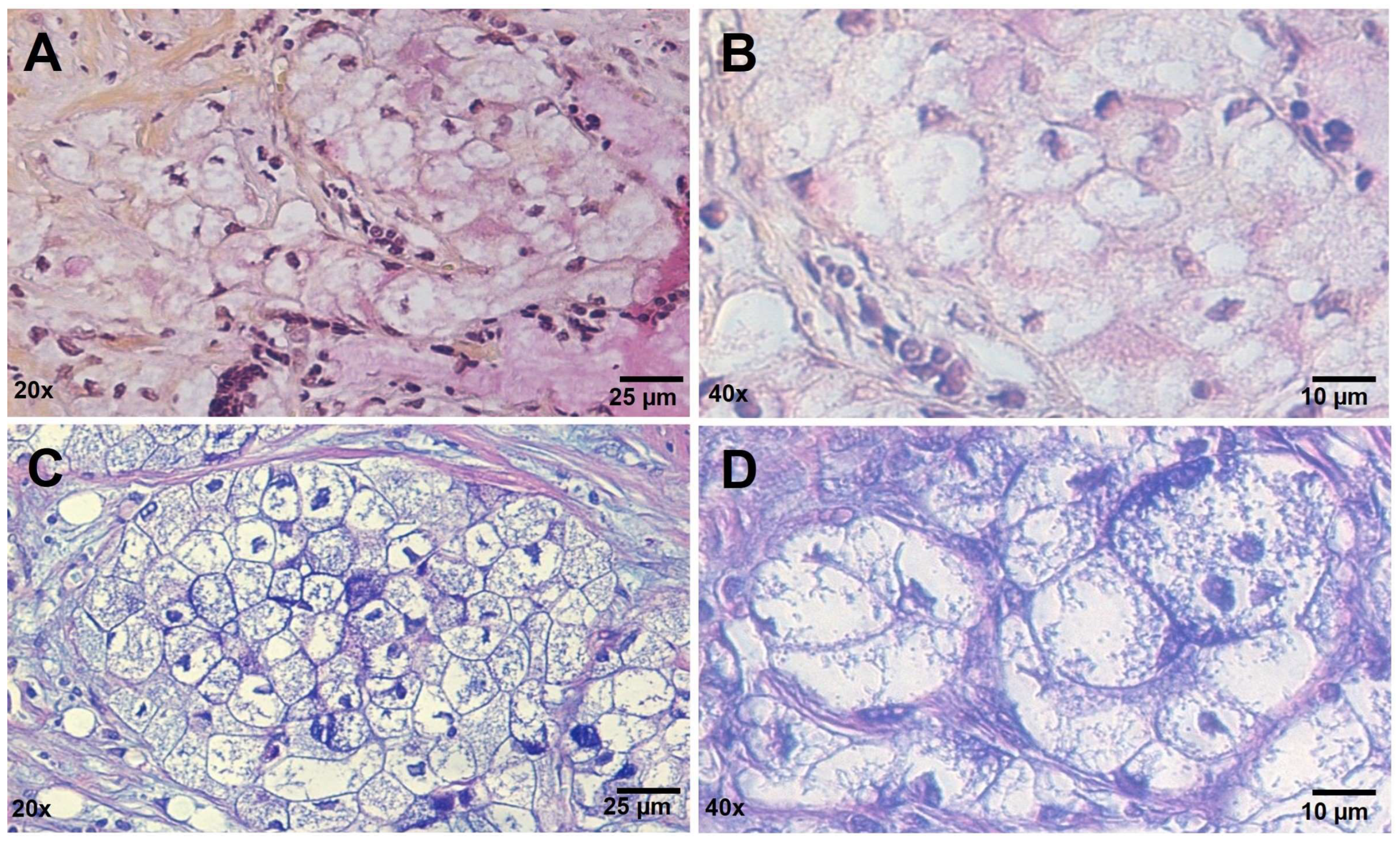
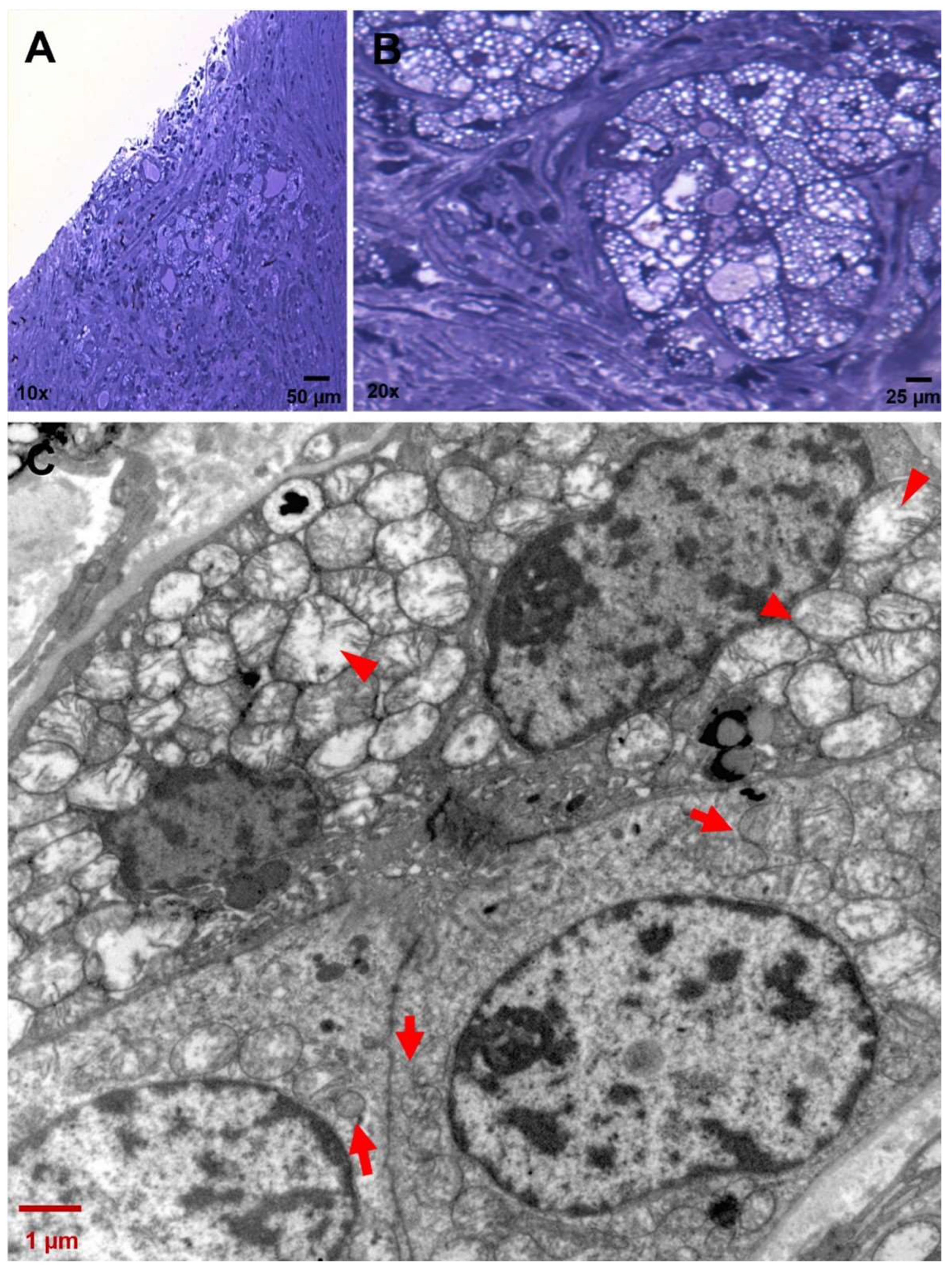
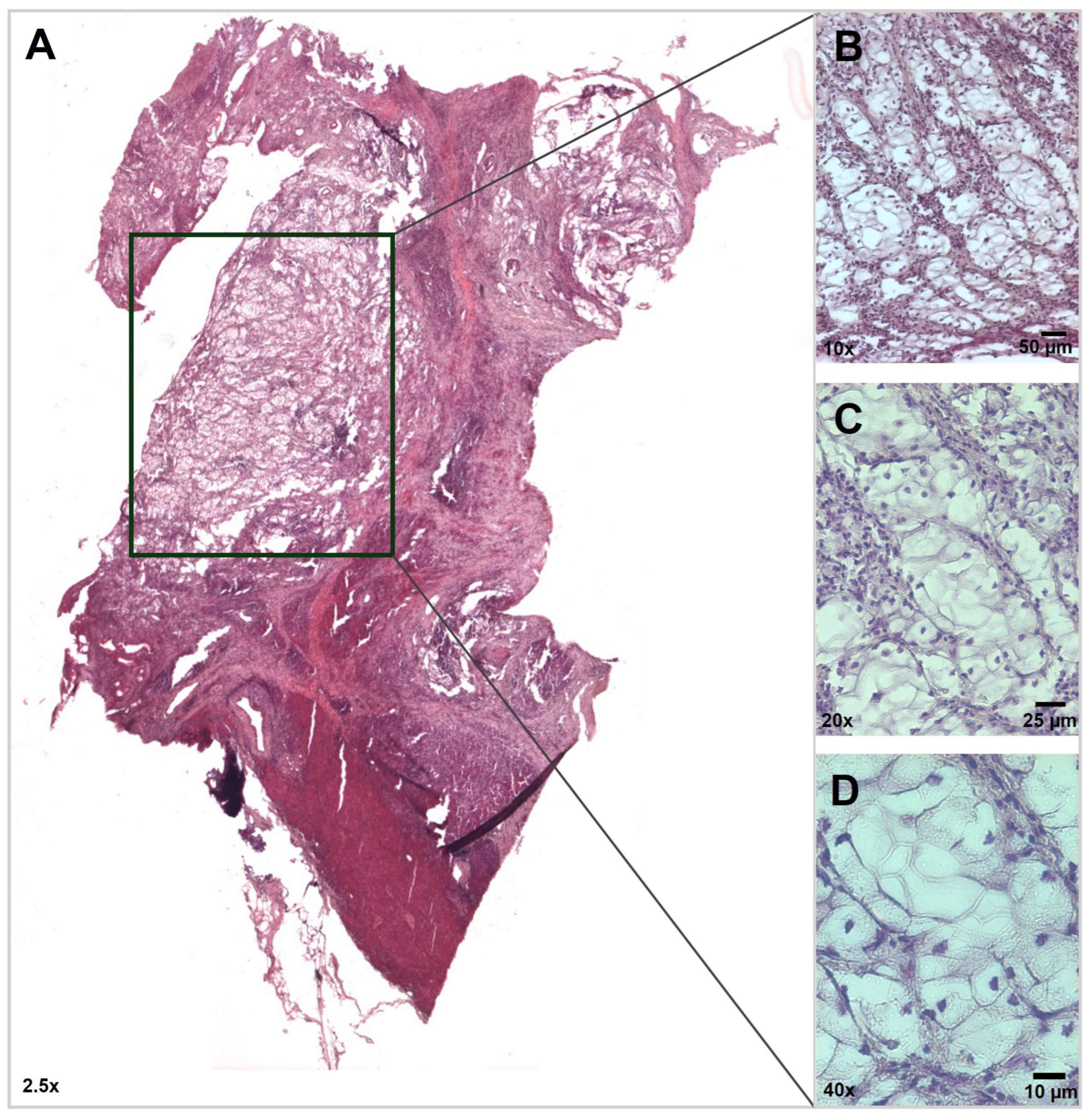
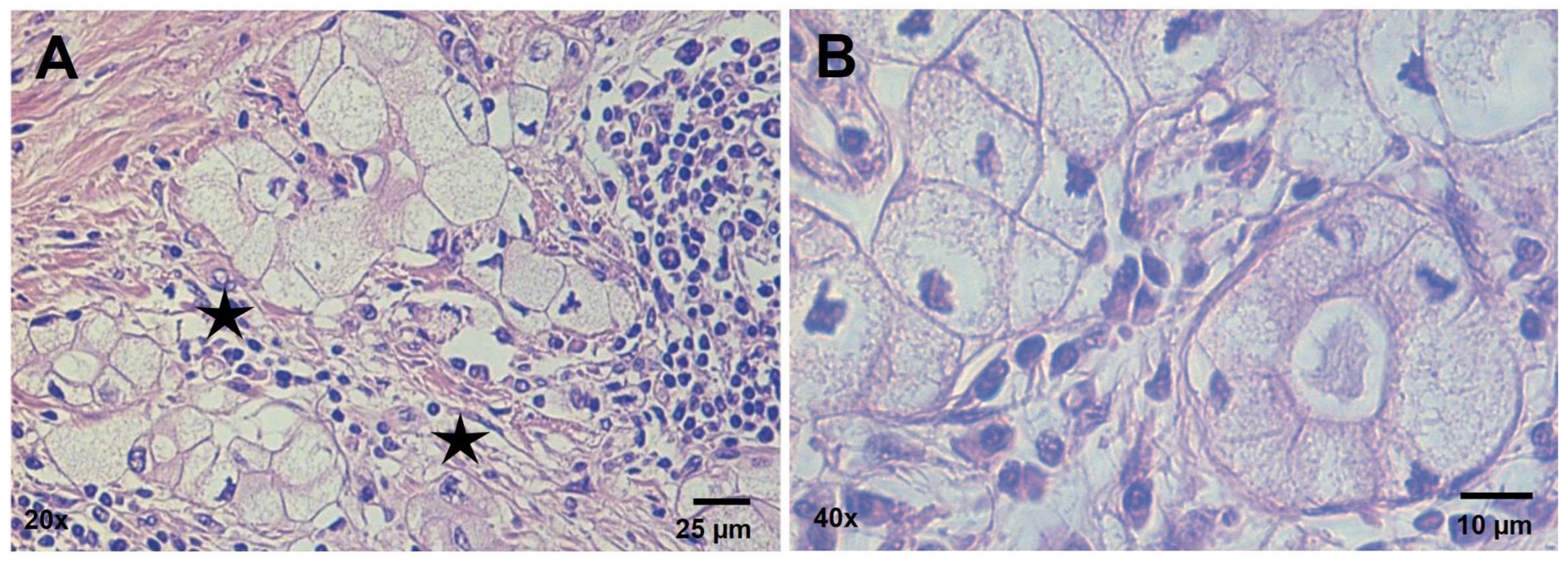
3.1. Histopathology Findings
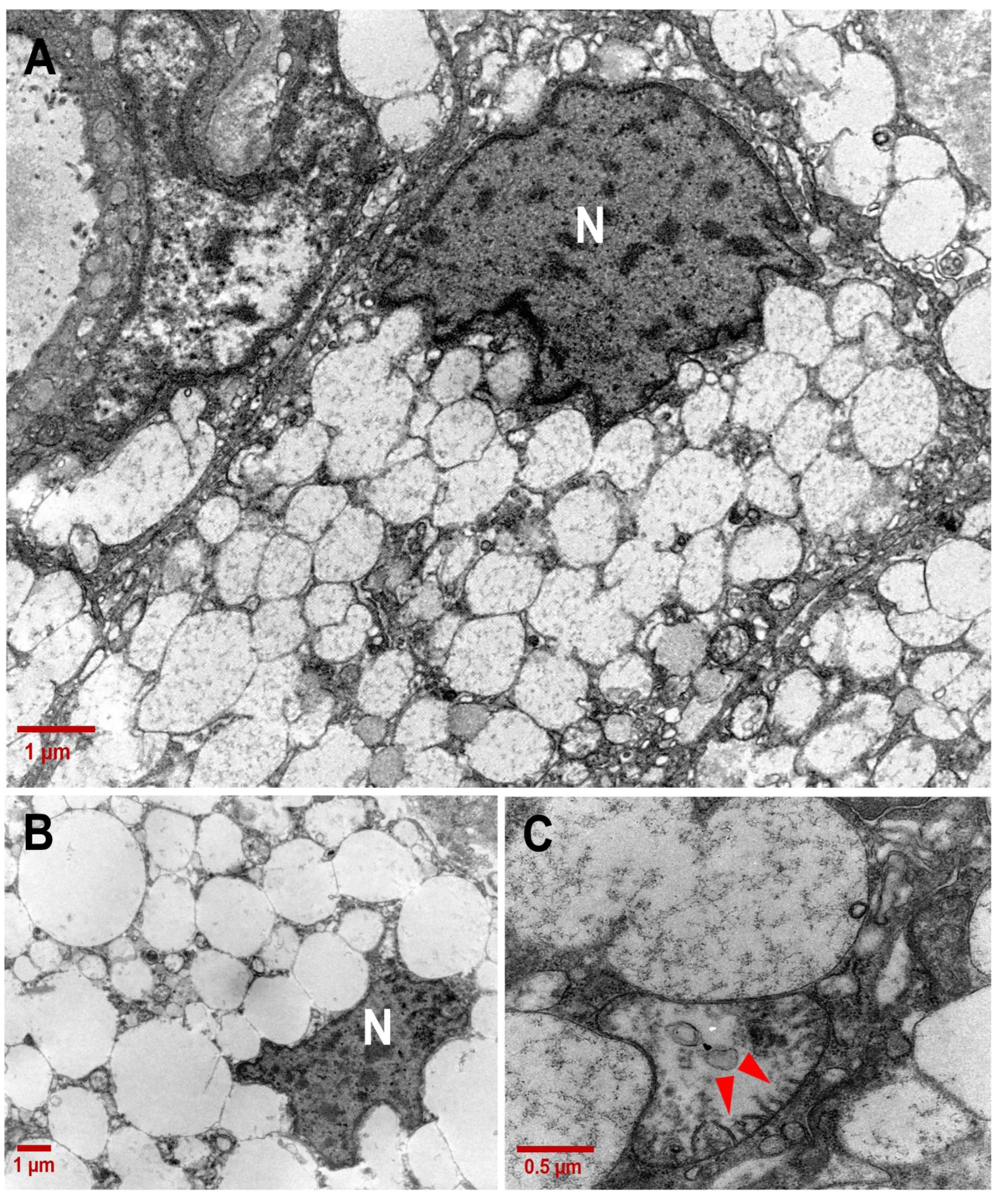
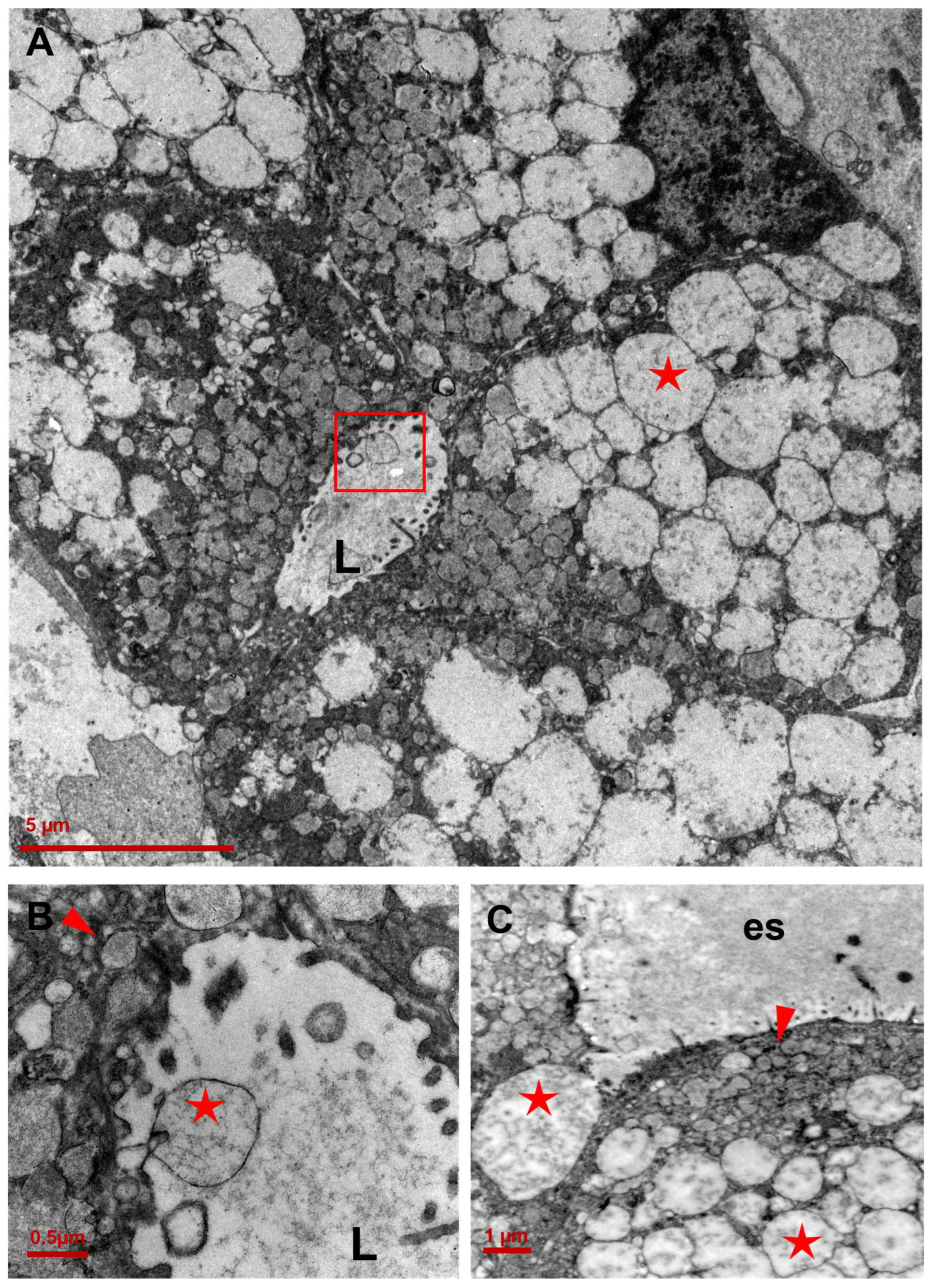
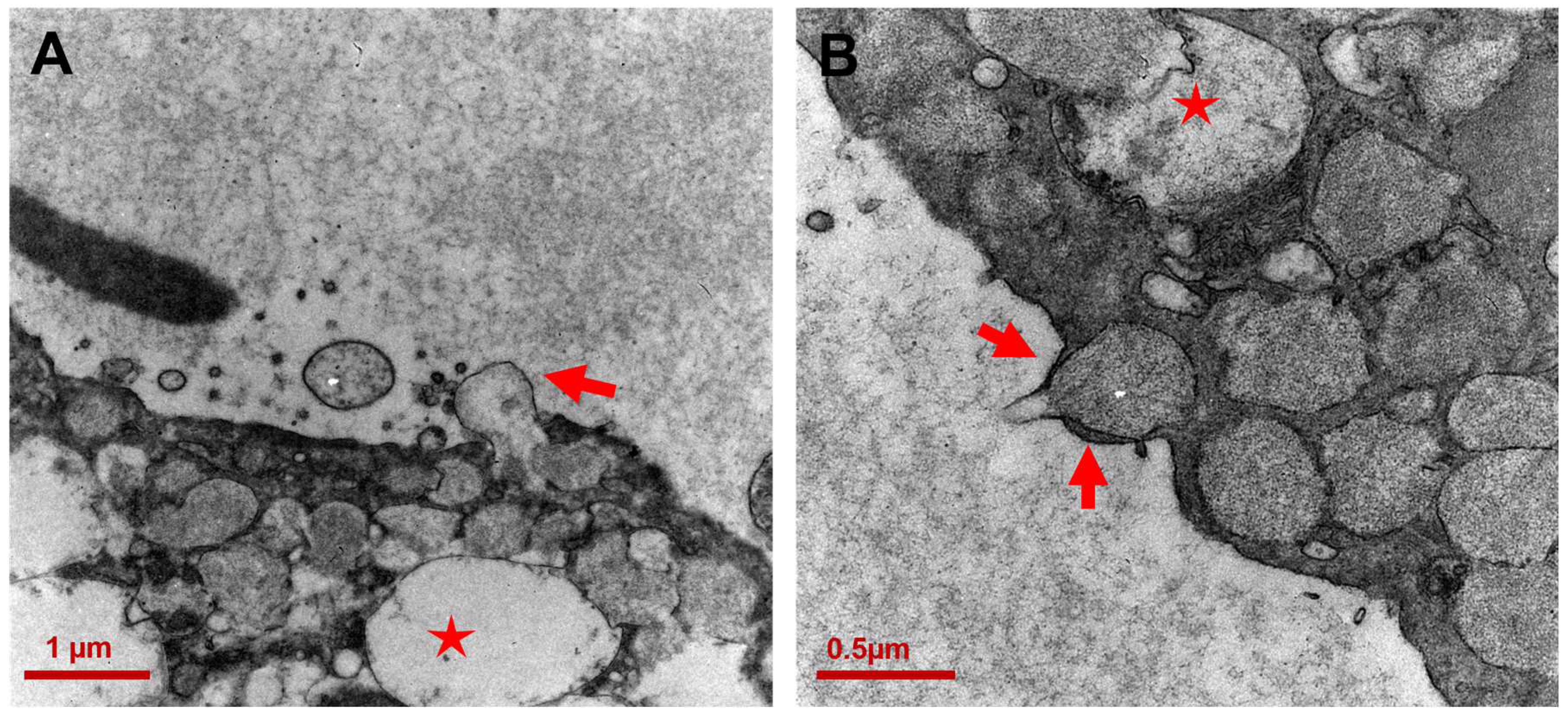
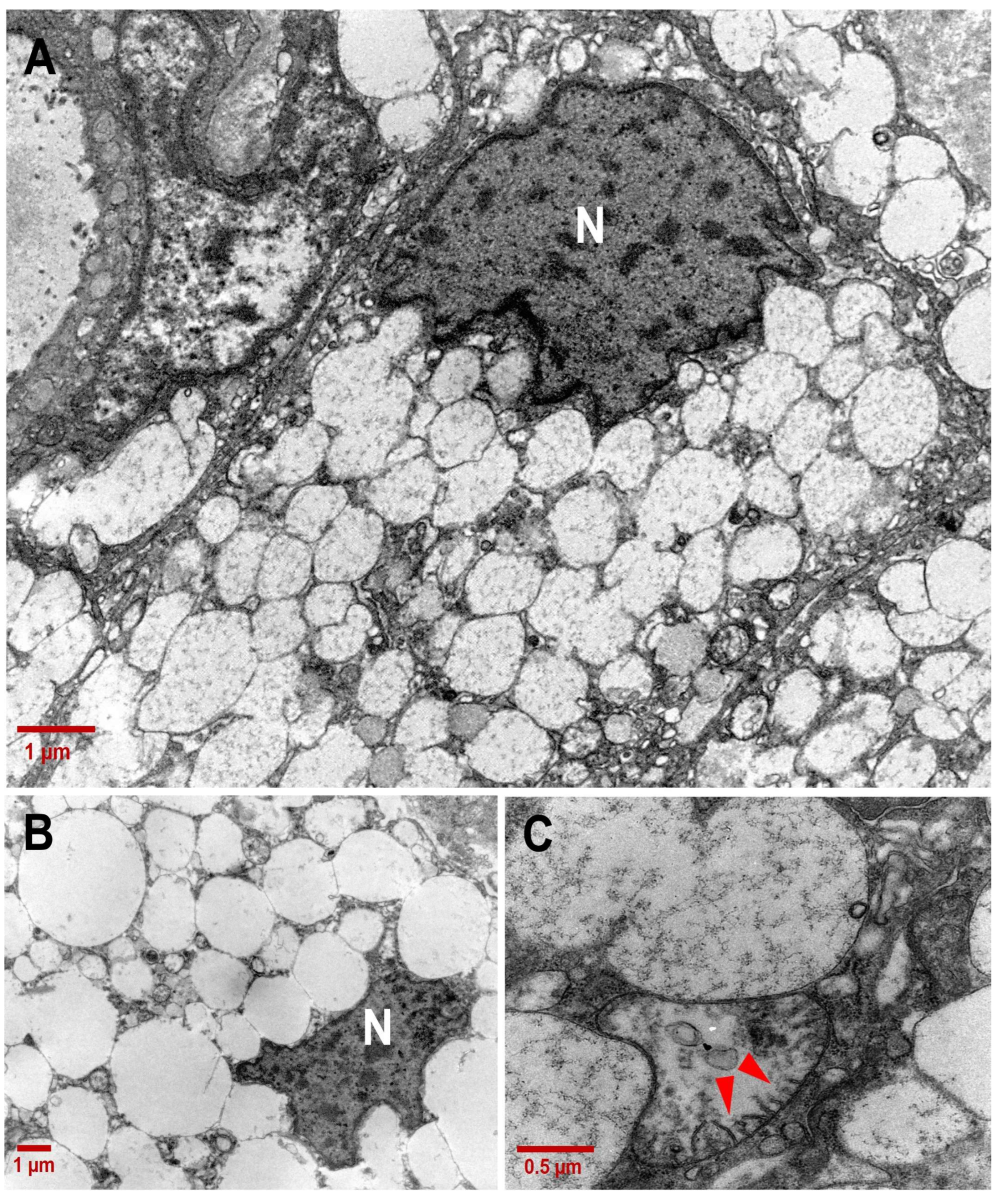
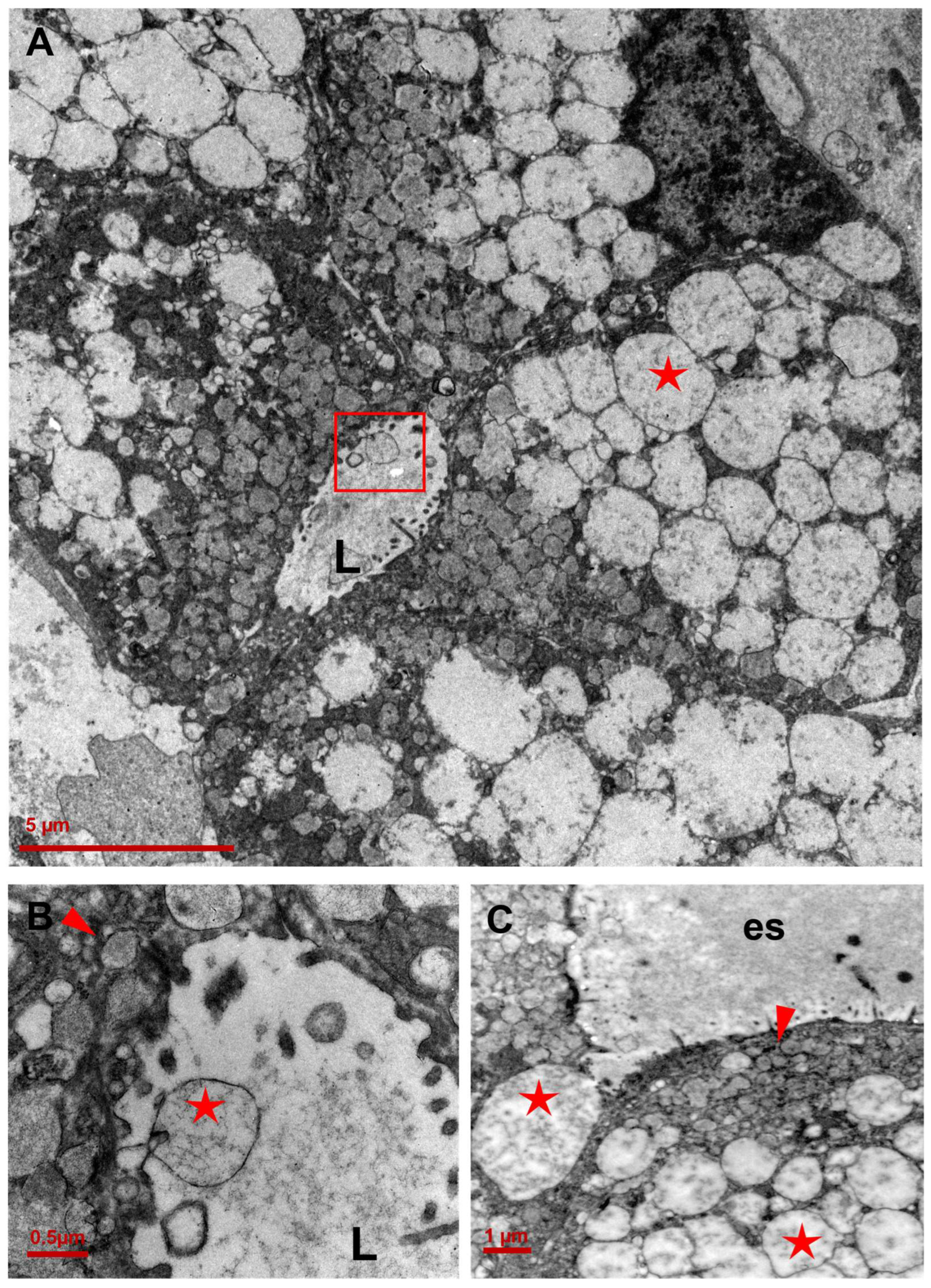
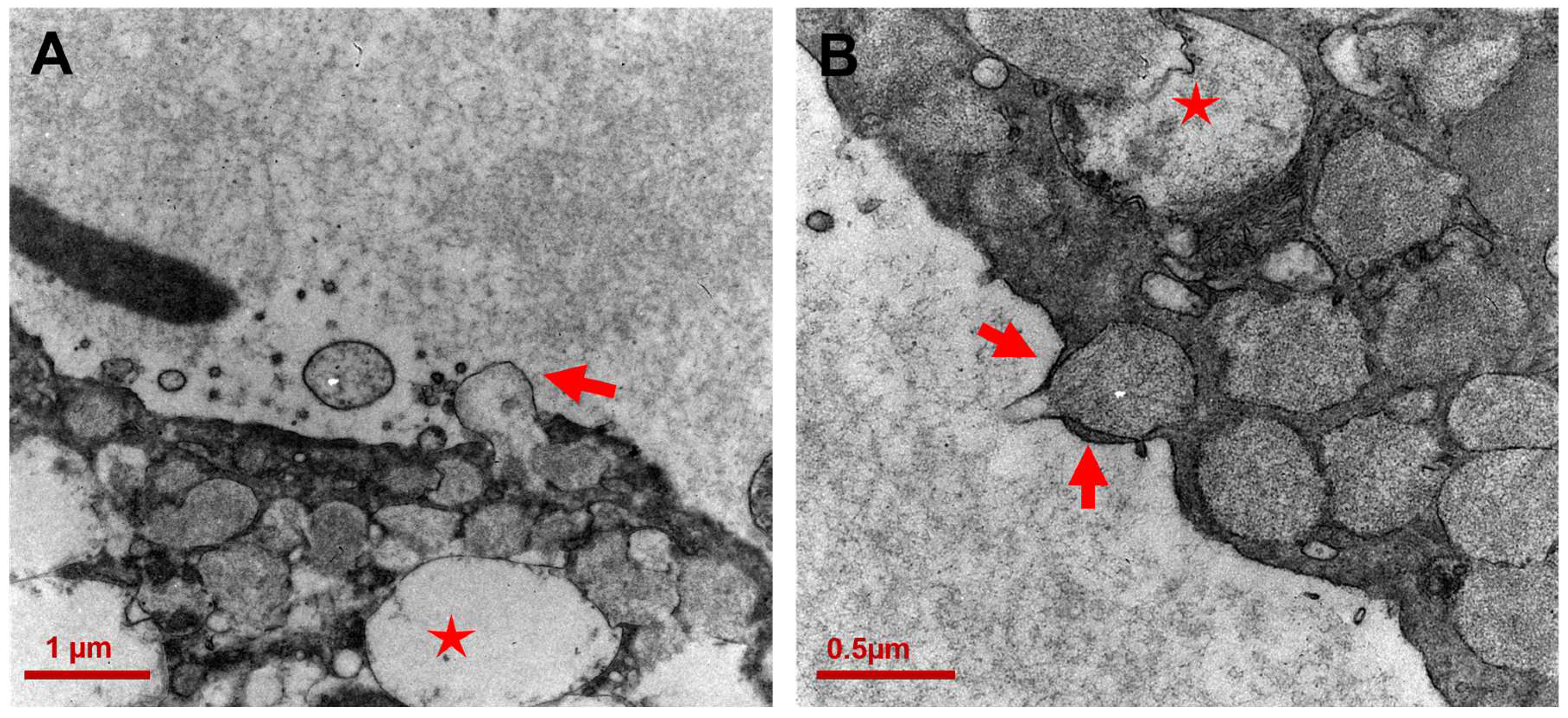
3.2. Ultrastructure of CCCP Cells
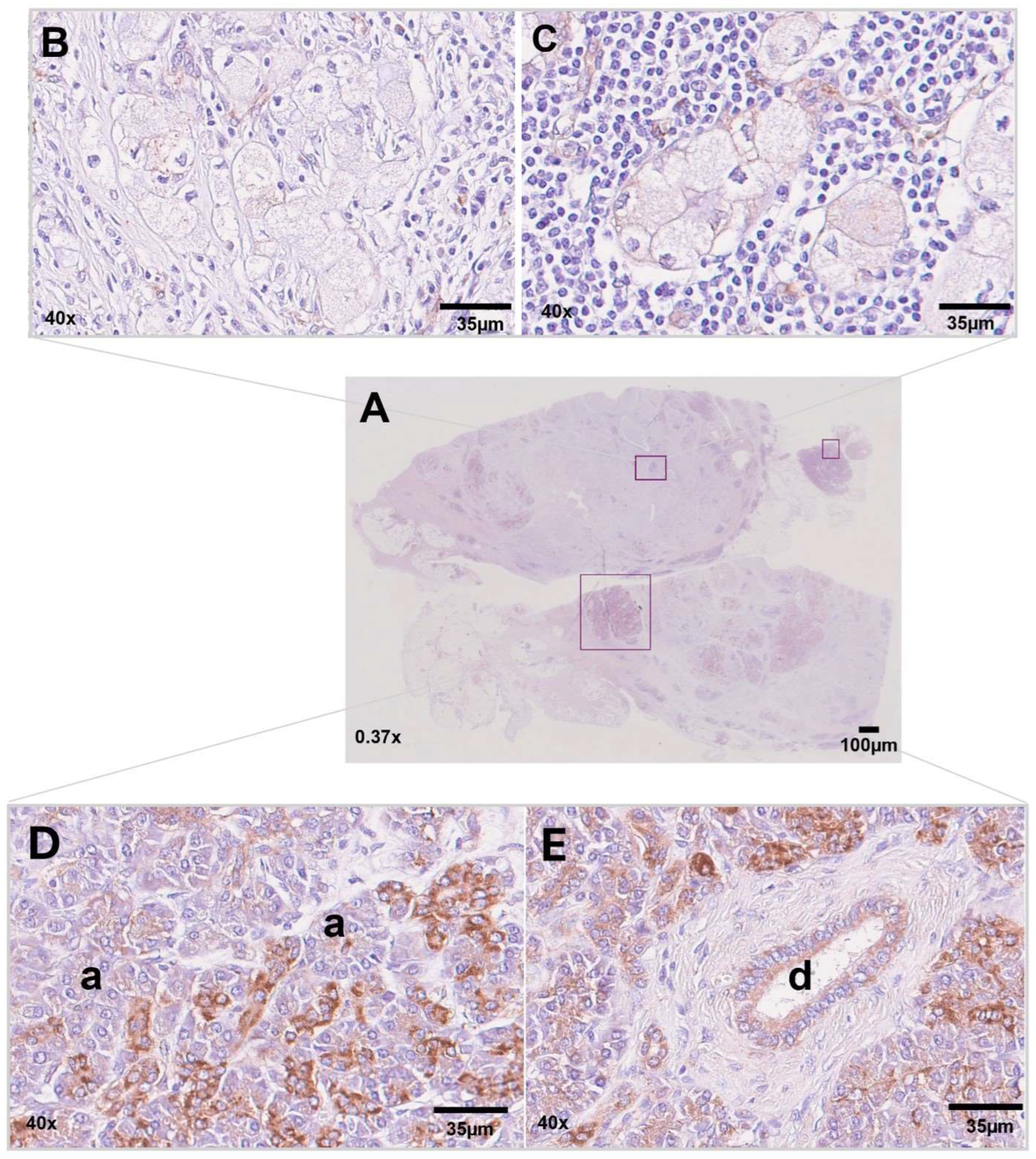
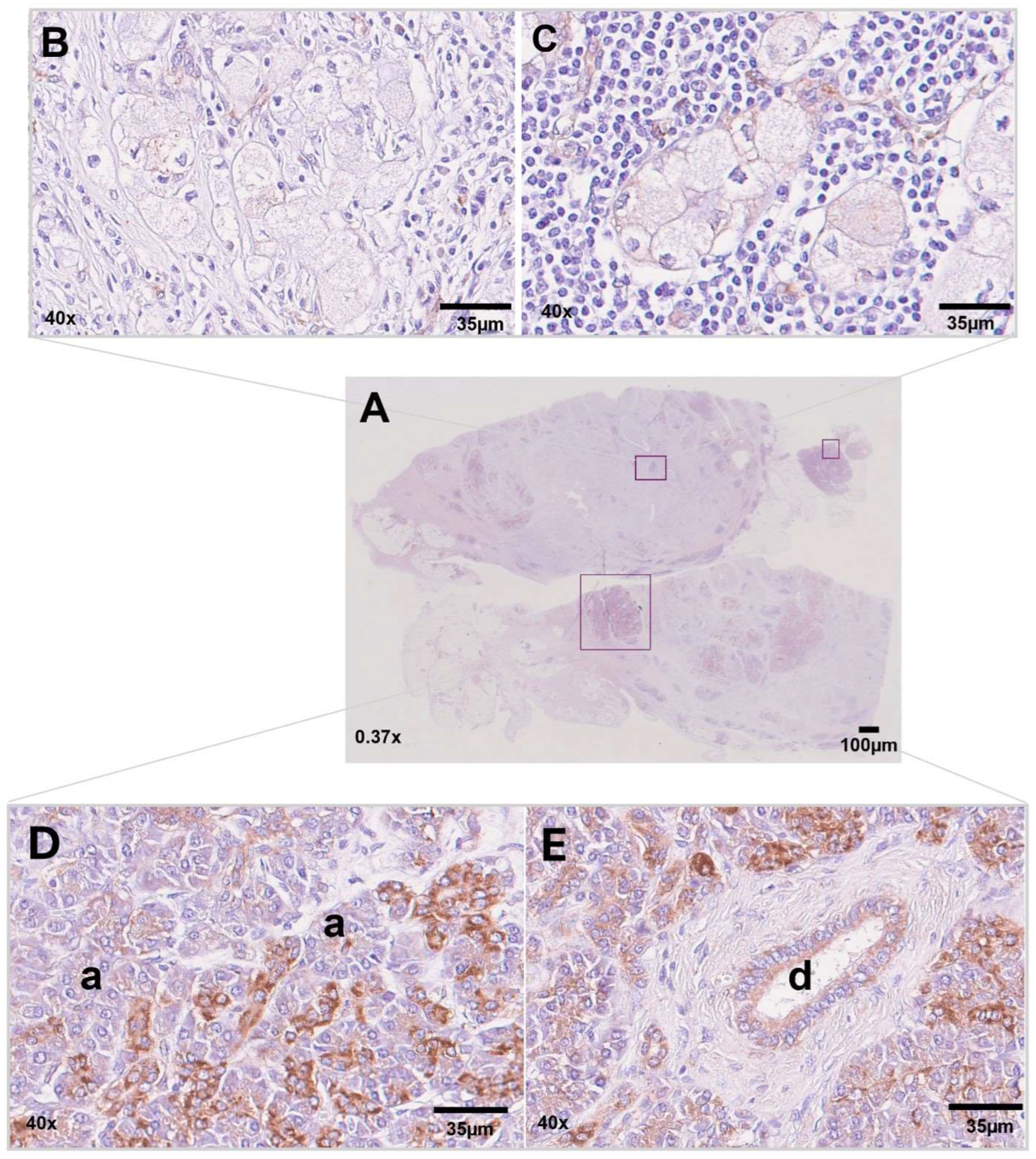
3.3. Immunohistochemistry for NDUFA4L2
4. Discussion
5. Materials and Methods
5.1. Light Microscopy
5.2. Immunohistochemistry (IHC)
5.3. Transmission Electron Microscopy (TEM)
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Park, W.; Chawla, A.; O’Reilly, E.M. Pancreatic Cancer: A Review. JAMA 2021, 326, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Krebs, A.M.; Mitschke, J.; Lasierra Losada, M.; Schmalhofer, O.; Boerries, M.; Busch, H.; Boettcher, M.; Mougiakakos, D.; Reichardt, W.; Bronsert, P.; et al. The EMT-activator Zeb1 is a key factor for cell plasticity and promotes metastasis in pancreatic cancer. Nat. Cell Biol. 2017, 19, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Giansante, V.; Stati, G.; Sancilio, S.; Guerra, E.; Alberti, S.; Di Pietro, R. The Dual Role of Necroptosis in Pancreatic Ductal Adenocarcinoma. Int. J. Mol. Sci. 2023, 24, 12633. [Google Scholar] [CrossRef] [PubMed]
- Truong, L.H.; Pauklin, S. Pancreatic Cancer Microenvironment and Cellular Composition: Current Understandings and Therapeutic Approaches. Cancers 2021, 13, 5028. [Google Scholar] [CrossRef] [PubMed]
- Bazzichetto, C.; Luchini, C.; Conciatori, F.; Vaccaro, V.; Di Cello, I.; Mattiolo, P.; Falcone, I.; Ferretti, G.; Scarpa, A.; Cognetti, F.; et al. Morphologic and Molecular Landscape of Pancreatic Cancer Variants as the Basis of New Therapeutic Strategies for Precision Oncology. Int. J. Mol. Sci. 2020, 21, 8841. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.A.; Greenhalf, B.; Ellis, I.; Sina-Frey, M.; Rieder, H.; Korte, B.; Gerdes, B.; Kress, R.; Ziegler, A.; Raeburn, J.A.; et al. BRCA2 germline mutations in familial pancreatic carcinoma. J. Natl. Cancer Inst. 2003, 95, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.; Pfeiffer, P.; Vilgrain, V.; Lamarca, A.; Seufferlein, T.; O’Reilly, E.M.; Hackert, T.; Golan, T.; Prager, G.; Haustermans, K.; et al. Pancreatic cancer: ESMO Clinical Practice Guideline for diagnosis, treatment and follow-up. Ann. Oncol. 2023, 34, 987–1002. [Google Scholar] [CrossRef]
- Nagtegaal, I.D.; Odze, R.D.; Klimstra, D.; Paradis, V.; Rugge, M.; Schirmacher, P.; Washington, K.M.; Carneiro, F.; Cree, I.A. The 2019 WHO classification of tumours of the digestive system. Histopathology 2020, 76, 182–188. [Google Scholar] [CrossRef]
- Cubilla, A.L.; Fitzgerald, P.J. Cancer (non-endocrine) of the pancreas. A suggested classification. Monogr. Pathol. 1980, 21, 82–110. [Google Scholar]
- Tannous, T.; Perez Rodriguez, A.L.; Mak, A.W.; Tannous, K.; Keating, M. Primary Clear Cell Carcinoma of the Pancreas: A Systematic Review. Cureus 2021, 13, e15668. [Google Scholar] [CrossRef]
- Kanai, N.; Nagaki, S.; Tanaka, T. Clear cell carcinoma of the pancreas. Acta Pathol. Jpn. 1987, 37, 1521–1526. [Google Scholar] [CrossRef]
- Kim, L.; Liao, J.; Zhang, M.; Talamonti, M.; Bentrem, D.; Rao, S.; Yang, G.Y. Clear cell carcinoma of the pancreas: Histopathologic features and a unique biomarker: Hepatocyte nuclear factor-1beta. Mod. Pathol. 2008, 21, 1075–1083. [Google Scholar] [CrossRef]
- Jamali, M.; Serra, S.; Chetty, R. Adenosquamous carcinoma of the pancreas with clear cell and rhabdoid components. A case report. JOP J. Oncol. Pract. 2007, 8, 330–334. [Google Scholar]
- Lucarelli, G.; Rutigliano, M.; Sallustio, F.; Ribatti, D.; Giglio, A.; Lepore Signorile, M.; Grossi, V.; Sanese, P.; Napoli, A.; Maiorano, E.; et al. Integrated multi-omics characterization reveals a distinctive metabolic signature and the role of NDUFA4L2 in promoting angiogenesis, chemoresistance, and mitochondrial dysfunction in clear cell renal cell carcinoma. Aging 2018, 10, 3957–3985. [Google Scholar] [CrossRef]
- Wakabayashi, T. Megamitochondria formation—Physiology and pathology. J. Cell. Mol. Med. 2002, 6, 497–538. [Google Scholar] [CrossRef]
- Javadov, S.; Chapa-Dubocq, X.; Makarov, V. Different approaches to modeling analysis of mitochondrial swelling. Mitochondrion 2018, 38, 58–70. [Google Scholar] [CrossRef]
- Lee, C.; Nam, J.S.; Lee, C.G.; Park, M.; Yoo, C.M.; Rhee, H.W.; Seo, J.K.; Kwon, T.H. Analysing the mechanism of mitochondrial oxidation-induced cell death using a multifunctional iridium(III) photosensitiser. Nat. Commun. 2021, 12, 26. [Google Scholar] [CrossRef]
- Sperandio, S.; de Belle, I.; Bredesen, D.E. An alternative, nonapoptotic form of programmed cell death. Proc. Natl. Acad. Sci. USA 2000, 97, 14376–14381. [Google Scholar] [CrossRef]
- Xu, C.C.; Lin, Y.F.; Huang, M.Y.; Zhang, X.L.; Wang, P.; Huang, M.Q.; Lu, J.J. Paraptosis: A non-classical paradigm of cell death for cancer therapy. Acta Pharmacol. Sin. 2023, 45, 223–237. [Google Scholar] [CrossRef]
- Carmona-Carmona, C.A.; Dalla Pozza, E.; Ambrosini, G.; Errico, A.; Dando, I. Divergent Roles of Mitochondria Dynamics in Pancreatic Ductal Adenocarcinoma. Cancers 2022, 14, 2155. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, E.Y.; Khryapenkova, T.G.; Galkina, S.I.; Sukhikh, G.T.; Zorov, D.B. Cytoplasm and organelle transfer between mesenchymal multipotent stromal cells and renal tubular cells in co-culture. Exp. Cell Res. 2010, 316, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Di Giuseppe, M.; Njah, J.; Sala, E.; Shiva, S.; St Croix, C.M.; Stolz, D.B.; Watkins, S.C.; Di, Y.P.; Leikauf, G.D.; et al. Mesenchymal stem cells use extracellular vesicles to outsource mitophagy and shuttle microRNAs. Nat. Commun. 2015, 6, 8472. [Google Scholar] [CrossRef]
- Uspenskaya, Y.A.; Malinovskaya, N.A.; Salmina, A.B. Intercellular Transport of Mitochondria: Molecular Mechanisms and Role in Maintaining Energy Homeostasis in Tissues. Cell Tissue Biol. 2022, 16, 97–113. [Google Scholar] [CrossRef]
- Simeone, P.; Bologna, G.; Lanuti, P.; Pierdomenico, L.; Guagnano, M.T.; Pieragostino, D.; Del Boccio, P.; Vergara, D.; Marchisio, M.; Miscia, S.; et al. Extracellular Vesicles as Signaling Mediators and Disease Biomarkers across Biological Barriers. Int. J. Mol. Sci. 2020, 21, 2514. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.N.; Yang, R.Z.; Zheng, H.L.; Jiang, L.S.; Jiang, S.D. NDUFA4L2 Regulated by HIF-1α Promotes Metastasis and Epithelial-Mesenchymal Transition of Osteosarcoma Cells Through Inhibiting ROS Production. Front. Cell Dev. Biol. 2020, 8, 515051. [Google Scholar] [CrossRef]
- Coller, H.A. Is cancer a metabolic disease? Am. J. Pathol. 2014, 184, 4–17. [Google Scholar] [CrossRef]
- Basile, M.; Centurione, L.; Passaretta, F.; Stati, G.; Soritau, O.; Susman, S.; Gindraux, F.; Silini, A.; Parolini, O.; Di Pietro, R. Mapping of the Human Amniotic Membrane: In Situ Detection of Microvesicles Secreted by Amniotic Epithelial Cells. Cell Transplant. 2023, 32, 9636897231166209. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giansante, V.; Di Angelo, L.; Calabrese, C.; De Sanctis, P.; Regi, P.; Martelli, F.M.; Stati, G.; Lattanzio, R.; Alberti, S.; Guerra, E.; et al. Novel Ultrastructural Insights into the Clear-Cell Carcinoma of the Pancreas: A Case Report. Int. J. Mol. Sci. 2024, 25, 4313. https://doi.org/10.3390/ijms25084313
Giansante V, Di Angelo L, Calabrese C, De Sanctis P, Regi P, Martelli FM, Stati G, Lattanzio R, Alberti S, Guerra E, et al. Novel Ultrastructural Insights into the Clear-Cell Carcinoma of the Pancreas: A Case Report. International Journal of Molecular Sciences. 2024; 25(8):4313. https://doi.org/10.3390/ijms25084313
Chicago/Turabian StyleGiansante, Valentina, Luca Di Angelo, Chiara Calabrese, Paolo De Sanctis, Paolo Regi, Filippo Maria Martelli, Gianmarco Stati, Rossano Lattanzio, Saverio Alberti, Emanuela Guerra, and et al. 2024. "Novel Ultrastructural Insights into the Clear-Cell Carcinoma of the Pancreas: A Case Report" International Journal of Molecular Sciences 25, no. 8: 4313. https://doi.org/10.3390/ijms25084313