
QTL Analysis for Rice Quality-Related Traits and Fine Mapping of qWCR3


 ,
, 
Abstract
1. Introduction
2. Results
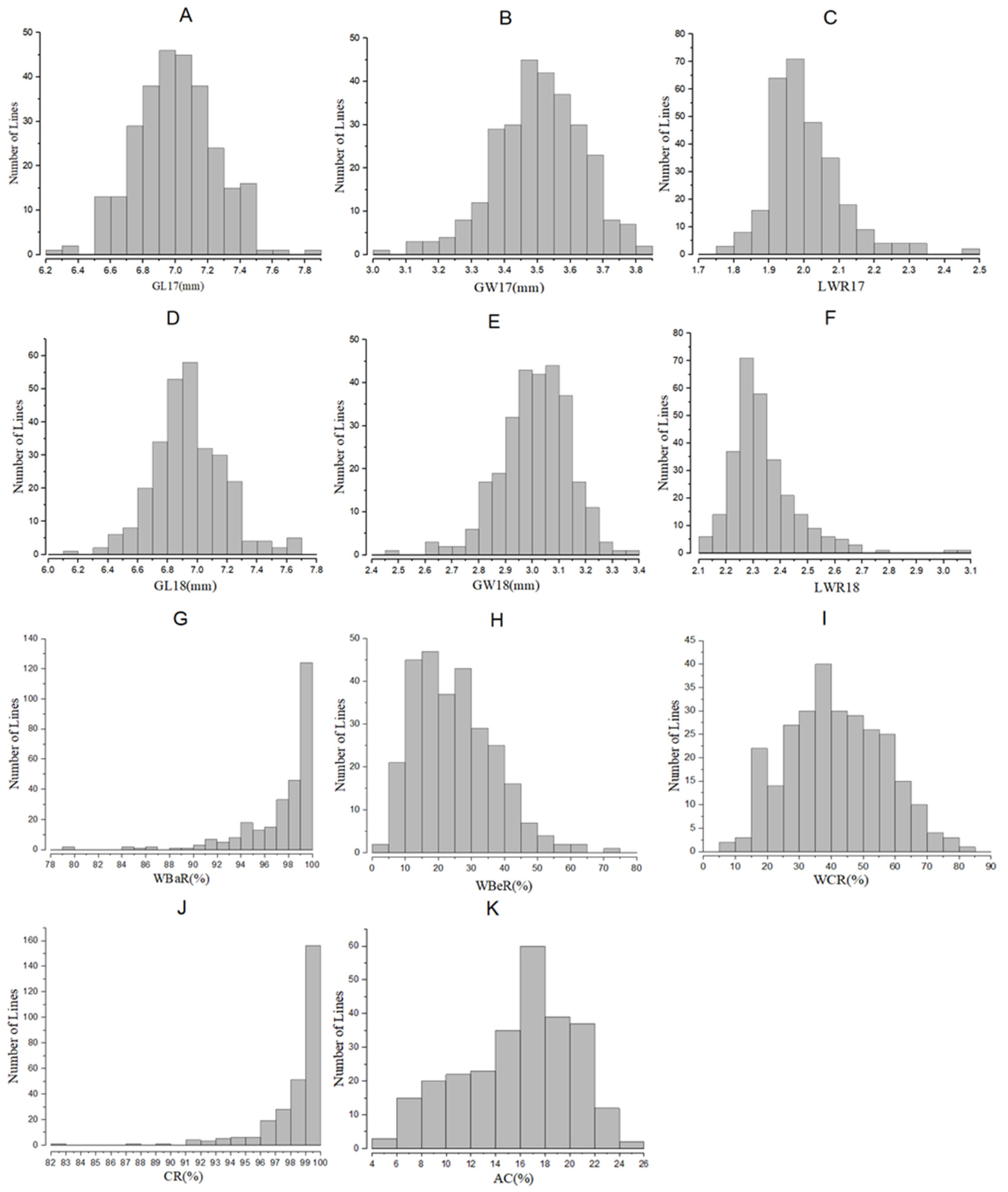
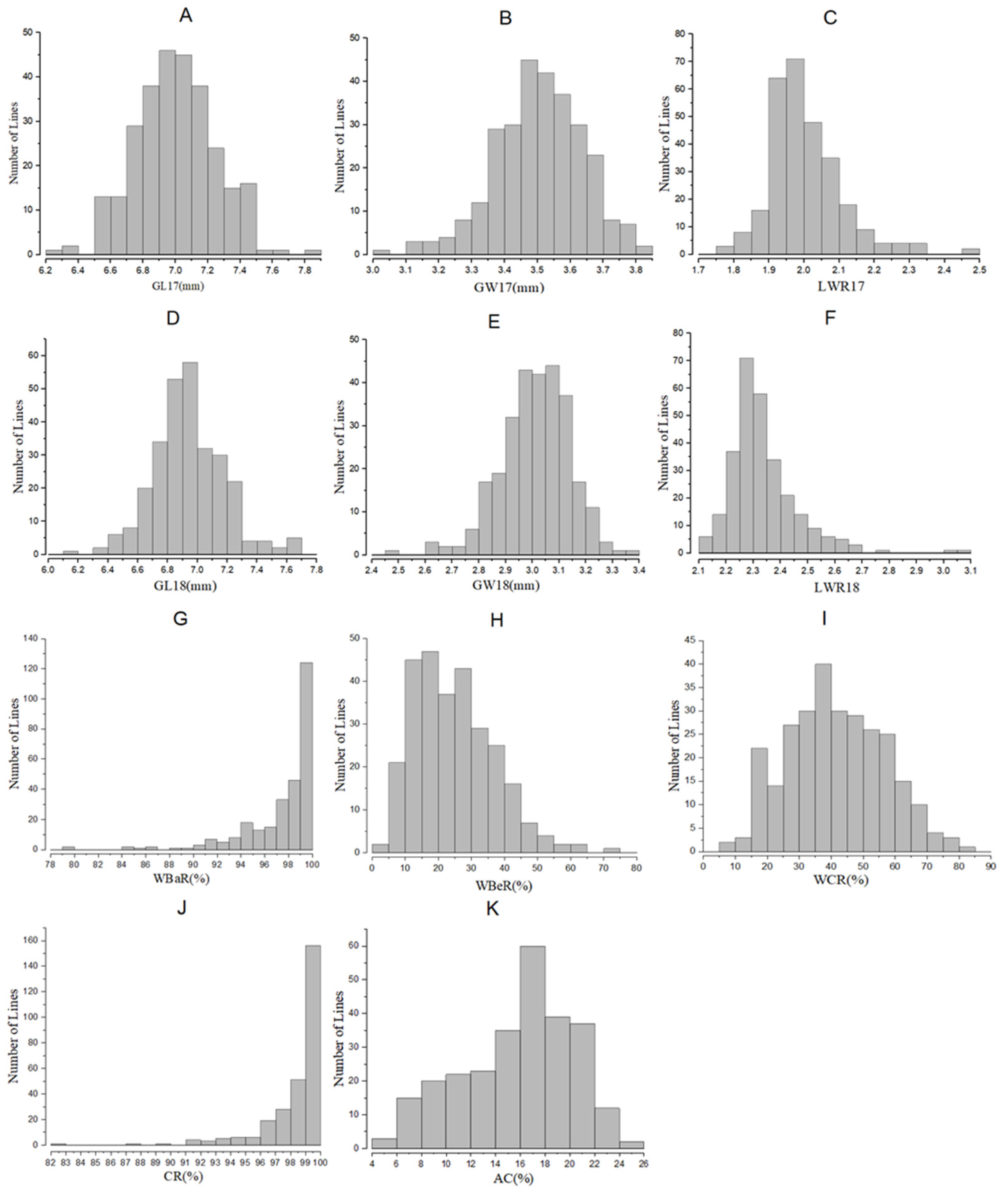
2.1. Phenotype Variation in BC3F2 and BC3F2:3 Populations
2.2. Construction of Genetic Linkage Map
2.3. 37 QTLs Were Detected in BC3F2 and BC3F2:3 Populations
2.4. Genetic Effect Validation of Seven QTLs
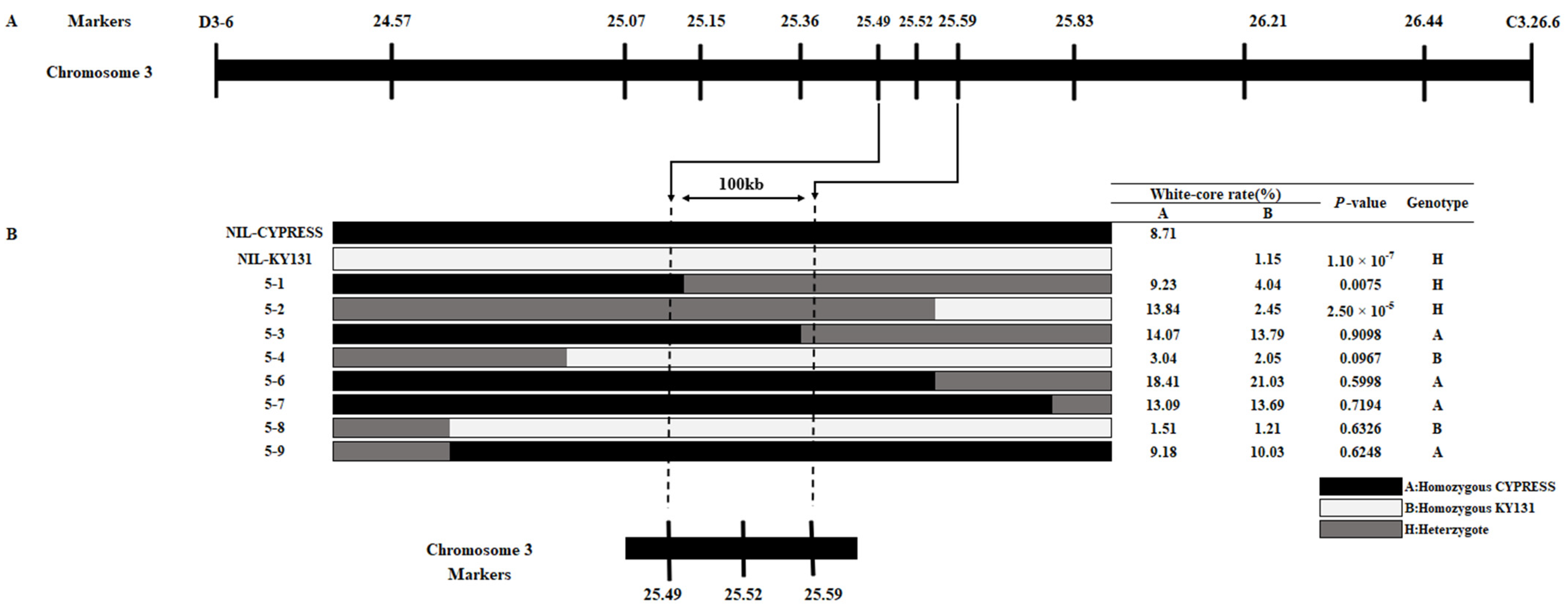
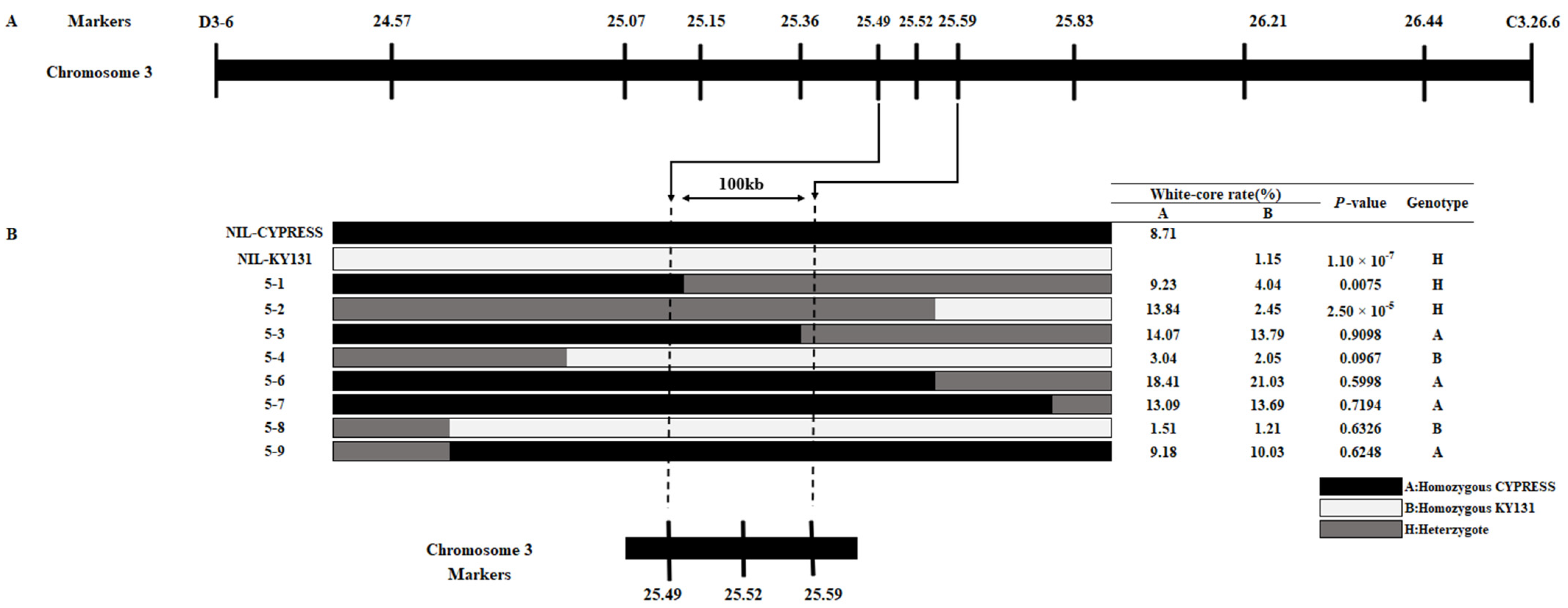
2.5. qWCR3 Was Fine-Mapped to a 100 kb Region
2.6. LOC_Os03g45210 Could Be the Candidate Genes of qWCR3
3. Discussion
3.1. Cloned Genes in the QTL Mapping Intervals
3.2. Four Chalkiness QTLs Were Newly Found
3.3. LOC_Os03g45210 Could Be a New Gene for Rice Chalkiness
4. Materials and Methods
4.1. Plant Materials and Field Experiment
4.2. Phenotyping and Statistical Analysis
4.3. Genotyping and QTL Analysis
4.4. Progeny Testing Analysis
4.5. Genetic Map Construction and QTL Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wing, R.; Purugganan, M.; Zhang, Q. The rice genome revolution: From an ancient grain to Green Super Rice. Nat. Rev. Genet. 2018, 19, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Qian, Q.; Liu, Q.; Yan, M.; Liu, X.; Yan, C.; Liu, G.; Gao, Z.; Tang, S.; Zeng, D.; et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Xia, D.; He, Y. Rice grain quality-traditional traits for high quality rice and health-plus substances. Mol. Breed. 2020, 40, 1. [Google Scholar] [CrossRef]
- Fitzgerald, M.; McCouch, S.; Hall, R. Not just a grain of rice: The quest for quality. Trends Plant Sci. 2009, 14, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhang, C.; Li, Q.; Liu, Q. Genetic control of grain appearance quality in rice. Biotechnol. Adv. 2022, 60, 108014. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Xing, Y.; Li, J.; Yu, S.; Xu, C.; Zhang, Q. Genetic bases of appearance quality of rice grains in Shanyou 63, an elite rice hybrid. Theor. Appl. Genet. 2000, 101, 823–829. [Google Scholar] [CrossRef]
- Chun, A.; Song, J.; Kim, K.; Lee, H. Quality of head and chalky rice and deterioration of eating quality by chalky rice. J. Crop Sci. Biotechnol. 2009, 12, 239–244. [Google Scholar] [CrossRef]
- Tang, S.; Zhang, H.; Liu, W.; Dou, Z.; Zhou, Q.; Chen, W.; Wang, S.; Ding, Y. Nitrogen fertilizer at heading stage effectively compensates for the deterioration of rice quality by affecting the starch-related properties under elevated temperatures. Food Chem. 2019, 277, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Xia, D.; Zhou, H.; Cheng, S.; Wang, Y.; Li, M.; Gao, G.; Zhang, Q.; Li, X.; He, Y. Fine mapping of qWCR7, a grain chalkiness QTL in rice. Mol. Breed. 2021, 41, 68. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, C.; Xing, Y.; Yun, P.; Luo, L.; Yan, B.; Peng, B.; Xie, W.; Wang, G.; Li, X.; et al. Chalk5 encodes a vacuolar H(+)-translocating pyrophosphatase influencing grain chalkiness in rice. Nat. Genet. 2014, 46, 398–404. [Google Scholar] [CrossRef]
- Wu, B.; Yun, P.; Zhou, H.; Xia, D.; Gu, Y.; Li, P.; Yao, J.; Zhou, Z.; Chen, J.; Liu, R.; et al. Natural variation in WHITE-CORE RATE 1 regulates redox homeostasis in rice endosperm to affect grain quality. Plant Cell 2022, 34, 1912–1932. [Google Scholar] [CrossRef] [PubMed]
- Mikami, I.; Uwatoko, N.; Ikeda, Y.; Yamaguchi, J.; Hirano, H.; Suzuki, Y.; Sano, Y. Allelic diversification at the wx locus in landraces of Asian rice. Theor. Appl. Genet. 2008, 116, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Bello, B.; Hou, Y.; Zhao, J.; Jiao, G.; Wu, Y.; Li, Z.; Wang, Y.; Tong, X.; Wang, W.; Yuan, W.; et al. NF-YB1-YC12-bHLH144 complex directly activates Wx to regulate grain quality in rice (Oryza sativa L.). Plant Biotechnol. J. 2019, 17, 1222–1235. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Cai, X.; Wang, Z.; Hong, M. An interaction between a MYC protein and an EREBP protein is involved in transcriptional regulation of the rice Wx gene. J. Biol. Chem. 2003, 278, 47803–47811. [Google Scholar] [CrossRef] [PubMed]
- Zeng, D.; Yan, M.; Wang, Y.; Liu, X.; Qian, Q.; Li, J. Du1, encoding a novel Prp1 protein, regulates starch biosynthesis through affecting the splicing of Wxb pre-mRNAs in rice (Oryza sativa L.). Plant Mol. Biol. 2007, 65, 501–509. [Google Scholar] [CrossRef]
- Heang, D.; Sassa, H. Antagonistic actions of HLH/bHLH proteins are involved in grain length and weight in rice. PLoS ONE 2012, 7, e31325. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Tong, H.; Xiao, Y.; Che, R.; Xu, F.; Hu, B.; Liang, C.; Chu, J.; Li, J.; Chu, C. Activation of Big Grain1 significantly improves grain size by regulating auxin transport in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11102–11107. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Nakagawa, H.; Tomita, C.; Shimatani, Z.; Ohtake, M.; Nomura, T.; Jiang, C.; Dubouzet, J.; Kikuchi, S.; Sekimoto, H.; et al. BRASSINOSTEROID UPREGULATED1, encoding a helix-loop-helix protein, is a novel gene involved in brassinosteroid signaling and controls bending of the lamina joint in rice. Plant Physiol. 2009, 151, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Hua, L.; Zhu, Z.; Tan, L.; Zhao, X.; Zhang, W.; Liu, F.; Fu, Y.; Cai, H.; Sun, X.; et al. GAD1 Encodes a Secreted Peptide That Regulates Grain Number, Grain Length, and Awn Development in Rice Domestication. Plant Cell 2016, 28, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, C.; Xing, Y.; Jiang, Y.; Luo, L.; Sun, L.; Shao, D.; Xu, C.; Li, X.; Xiao, J.; et al. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 2011, 43, 1266–1269. [Google Scholar] [CrossRef]
- Tong, H.; Liu, L.; Jin, Y.; Du, L.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice. Plant Cell 2012, 24, 2562–2577. [Google Scholar] [CrossRef] [PubMed]
- Xia, K.; Ou, X.; Tang, H.; Wang, R.; Wu, P.; Jia, Y.; Wei, X.; Xu, X.; Kang, S.; Kim, S.; et al. Rice microRNA osa-miR1848 targets the obtusifoliol 14α-demethylase gene OsCYP51G3 and mediates the biosynthesis of phytosterols and brassinosteroids during development and in response to stress. New Phytol. 2015, 208, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.; Yu, X.; Zhou, K.; et al. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 17043. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Sun, S.; Yao, J.; Wang, C.; Yu, S.; Xu, C.; Li, X.; Zhang, Q. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 19579–19584. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Wang, L.; Mao, H.; Shao, L.; Li, X.; Xiao, J.; Ouyang, Y.; Zhang, Q. A G-protein pathway determines grain size in rice. Nat. Commun. 2018, 9, 851. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, G.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.; Zeng, L.; Xu, E.; Xu, J.; et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Zhang, L.; Gampala, S.; Zhu, S.; Song, W.; Chong, K.; Wang, Z. Functions of OsBZR1 and 14-3-3 proteins in brassinosteroid signaling in rice. Proc. Natl. Acad. Sci. USA 2007, 104, 13839–13844. [Google Scholar] [CrossRef] [PubMed]
- Si, L.; Chen, J.; Huang, X.; Gong, H.; Luo, J.; Hou, Q.; Zhou, T.; Lu, T.; Zhu, J.; Shangguan, Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Fang, J.; Ou, S.; Gao, S.; Zhang, F.; Du, L.; Xiao, Y.; Wang, H.; Sun, X.; Chu, J.; et al. Variations in CYP78A13 coding region influence grain size and yield in rice. Plant Cell Environ. 2015, 38, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.; Mieda, K.; Ando, T.; Kono, I.; Yano, M.; Kitano, H.; Iwasaki, Y. The SMALL AND ROUND SEED1 (SRS1/DEP2) gene is involved in the regulation of seed size in rice. Genes Genet. Syst. 2010, 85, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Kearney, L.; Flashman, E. Targeting plant cysteine oxidase activity for improved submergence tolerance. Plant J. 2022, 109, 779–788. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.; Giuntoli, B.; Giorgi, F.; Voesenek, L.; Perata, P.; van Dongen, J. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Tanabata, T.; Shibaya, T.; Hori, K.; Ebana, K.; Yano, M. SmartGrain: High-throughput phenotyping software for measuring seed shape through image analysis. Plant Physiol. 2012, 160, 1871–1880. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Corke, H.; Sun, M. Nucleotide diversity in starch synthase IIa and validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice (Oryza sativa L.). Theor. Appl. Genet. 2006, 113, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Agetsuma, M.; Kitano, H.; Yoshimura, A.; Matsuoka, M.; Jacobsen, S.E.; Ashikari, M. A metastable DWARF1 epigenetic mutant affecting plant stature in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 11218–11223. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.; Lincoln, S.; Newberg, L. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Basten, C.; Zeng, Z. Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State. 2006. Available online: http://statgen.ncsu.edu/qtlcart/WQTLCart.htm (accessed on 8 April 2024).


{kind=link}
{kind=link}
{kind=link}
| Trait | Parents | 2017HN BC3F2 | 2018WH BC3F2:3 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| KY131 | Cypress | Mean | SD | MIN | MAX | Mean | SD | MIN | MAX | |
| GL (mm) | 6.81 | 8.67 | 6.99 | 0.29 | 5.37 | 7.87 | 6.94 | 0.01 | 6.14 | 7.67 |
| GW (mm) | 2.88 | 2.52 | 3.50 | 0.13 | 3.05 | 3.84 | 3.00 | 0.01 | 2.47 | 3.38 |
| LWR | 2.39 | 3.46 | 2.01 | 0.10 | 1.79 | 2.46 | 2.33 | 0.01 | 2.11 | 3.08 |
| BR (%) | 22.06 | 0.99 | 97.48 | 3.32 | 79.21 | 100 | ||||
| WCR (%) | 3.82 | 3.16 | 41.38 | 15.08 | 7.66 | 80.72 | ||||
| WBR (%) | 2.96 | 1.56 | 24.91 | 12.27 | 3.72 | 74.89 | ||||
| CR (%) | 24.42 | 4.98 | 98.41 | 2.24 | 82.67 | 100 | ||||
| AC (%) | 16.67 | 25.06 | 15.81 | 4.49 | 5.63 | 24.51 | ||||
| GL (mm) | GW (mm) | LWR | BR (%) | WCR (%) | WBR (%) | CR (%) | |
|---|---|---|---|---|---|---|---|
| GW (mm) | 0.20 ** | ||||||
| LWR | 0.50 ** | −0.74 ** | |||||
| BR (%) | 0.06 | 0.01 | 0.04 | ||||
| WCR (%) | −0.04 | 0.28 ** | −0.28 ** | 0.14 * | |||
| WBR (%) | −0.12 * | 0.24 ** | −0.29 ** | −0.01 | 0.55 ** | ||
| CR (%) | 0.03 | 0.08 | −0.04 | 0.93 ** | 0.22 ** | 0.13 * | |
| AC (%) | −0.04 | −0.01 | −0.02 | 0.03 | 0.24 ** | 0.01 | 0.04 |
| Traits | QTL | Chr | Interval | BC3F2 | BC3F2:3 | ||||
|---|---|---|---|---|---|---|---|---|---|
| LOD | ADD | V (%) | LOD | ADD | V (%) | ||||
| GL | qGL3 | 3 | T3-1-RM545 | 11.46 | 1.291 | 20.60 | |||
| qGL4 | 4 | Z4-20.6-Z4-26.5 | 3.06 | −0.188 | 4.85 | ||||
| qGL6 | 6 | Z6-6.0-TT6-1 | 4.81 | −0.140 | 5.88 | ||||
| qGL7.1 | 7 | RM481-Z7-5.4 | 3.69 | 0.125 | 4.55 | ||||
| qGL7.2 | 7 | T7-3-RM478 | 6.72 | −0.342 | 9.12 | 8.74 | −0.316 | 12.82 | |
| qGL7.3 | 7 | RM478-Z7-28.3 | 4.03 | −0.252 | 7.07 | ||||
| qGL8 | 8 | RM515-Z8-25.3 | 4.44 | −0.178 | 6.61 | ||||
| GW | qGW1 | 1 | RM486-A1.40.39 | 2.96 | 0.097 | 4.58 | |||
| qGW3 | 3 | T3-1-RM545 | 7.87 | 0.608 | 17.52 | ||||
| qGW5.1 | 5 | RM405-TT5-1 | 3.49 | 0.156 | 10.03 | ||||
| qGW5.2 | 5 | Z5-5.1-Z5-7.2 | 6.35 | 0.139 | 13.03 | ||||
| qGW7.1 | 7 | RM346-RM478 | 5.83 | 0.196 | 12.50 | ||||
| qGW7.2 | 7 | T7-3-RM478 | 5.36 | 0.140 | 8.94 | ||||
| LWR | qLWR3 | 3 | Z3-14.8-Z3-17.6 | 3.83 | −0.087 | 4.10 | |||
| qLWR5 | 5 | Z5-5.1-Z5-7.2 | 6.20 | −0.095 | 8.82 | 10.56 | −0.179 | 21.13 | |
| qLWR7.1 | 7 | RM346-RM478 | 25.80 | −0.237 | 33.26 | 17.73 | −0.246 | 26.86 | |
| qLWR7.2 | 7 | T7-3-RM346 | 10.19 | −0.203 | 13.20 | ||||
| qLWR9 | 9 | 9.8.0-Z9-10.4 | 3.72 | −0.275 | 14.78 | ||||
| BR | qBR1 | 1 | A1.40.39-RM104 | 2.69 | 6.398 | 4.25 | |||
| qBR2 | 2 | T2-1-T2-2 | 4.37 | 2.790 | 8.94 | ||||
| qBR9 | 9 | 9.2.0-9.8.0 | 3.03 | 2.680 | 3.79 | ||||
| qBR11 | 11 | C11-3.5-11-6.6 | 3.77 | −2.135 | 5.05 | ||||
| WBR | qWBR1.1 | 1 | C1-23.3-RM5 | 3.24 | 6.771 | 4.32 | |||
| qWBR1.2 | 1 | D1-8-B1.40.61 | 4.52 | −9.274 | 5.46 | ||||
| qWBR5 | 5 | Z5-5.1-Z5-7.2 | 4.54 | 8.925 | 6.73 | ||||
| qWBR6 | 6 | RM586-Z6-6.0 | 9.00 | −19.770 | 18.17 | ||||
| WCR | qWCR3 | 3 | D3-6-C3.26.6 | 2.54 | −10.607 | 4.88 | |||
| qWCR5.1 | 5 | Z5-5.1-TT5-1 | 2.98 | 10.873 | 5.75 | ||||
| qWCR5.2 | 5 | TT5-1-TT5-2 | 2.81 | 7.307 | 3.99 | ||||
| qWCR11 | 11 | C11-3.5-11.6.6 | 2.98 | −1.310 | 4.47 | ||||
| qWCR12 | 12 | Z12-19.9-Z12-25.2 | 4.38 | −10.997 | 7.02 | ||||
| CR | qCR1.1 | 1 | Z1-18.0-C1-23.3 | 3.25 | 3.393 | 26.30 | |||
| qCR1.2 | 1 | C1-23.3-RM5 | 4.04 | 1.448 | 5.59 | ||||
| qCR2 | 2 | T2-1-T2-2 | 3.43 | 1.675 | 7.22 | ||||
| qCR9 | 9 | H9.4.7-9.8.0 | 3.48 | 1.880 | 4.57 | ||||
| qCR11 | 11 | 11-6.6-Z11-16.9 | 2.88 | −1.026 | 3.76 | ||||
| AC | qAC4 | 4 | Z4-6.0-Z4-14.3 | 2.52 | 3.687 | 4.01 | |||
| Year | QTL | Cypress Genotype | KY131 Genotype | p Value | ADD |
|---|---|---|---|---|---|
| 2019WH | qLWR7.2 | 2.89 ± 0.17 | 2.58 ± 0.20 | 0.0016 | 0.15 |
| qBR2 (%) | 40.05 ± 20.93 | 61.97 ± 18.97 | 0.033 | −10.96 | |
| qWBR1.2 (%) | 25.65 ± 11.52 | 8.66 ± 7.50 | 0.0019 | 8.50 | |
| 2020WH | qGL7.2 (mm) | 8.42 ± 0.39 | 7.70 ± 0.24 | 0.0059 | 0.36 |
| qGW3 (mm) | 3.15 ± 0.15 | 3.31 ± 0.08 | 0.0069 | −0.08 | |
| qLWR7.2 | 2.80 ± 0.07 | 2.50 ± 0.06 | 5.58 × 10−9 | 0.15 | |
| 2021WH | qBR2 (%) | 29.06 ± 6.19 | 41.81 ± 9.02 | 0.0026 | −6.37 |
| 2021HN | qWCR3(%) | 14.47 ± 5.90 | 2.46 ± 2.99 | 0.001 | 6.01 |
| qWCR11(%) | 3.91 ± 1.26 | 14.86 ± 3.86 | 6.14 × 10−7 | −5.47 | |
| 2022WH | qWCR3(%) | 8.71 ± 4.29 | 1.16 ± 0.75 | 1.09 × 10−7 | 3.78 |
| qWCR11(%) | 6.08 ± 4.24 | 12.02 ± 6.46 | 0.0070 | −2.97 |
| Gene | Gene Product Name |
|---|---|
| LOC_Os03g45150 | LTP family protein precursor |
| LOC_Os03g45160 | Hypothetical protein |
| LOC_Os03g45170 | Amino acid permease |
| LOC_Os03g45180 | Expressed protein |
| LOC_Os03g45194 | Oxidoreductase |
| LOC_Os03g45210 | Plant Cysteine oxidase-3 |
| LOC_Os03g45220 | Expressed protein |
| LOC_Os03g45230 | Expressed protein |
| LOC_Os03g45250 | Plant Cysteine oxidase-2 |
| LOC_Os03g45260 | Vesicle transport v-SNARE protein |
| LOC_Os03g45270 | CS domain containing protein |
| LOC_Os03g45280 | Dehydrin |
| LOC_Os03g45290 | Ankyrin repeat domain-containing protein 50 |
| LOC_Os03g45300 | Transposon protein |
| LOC_Os03g45310 | Hypothetical protein |
| LOC_Os03g45320 | Dehydrogenase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, H.; Wang, Y.; Liu, E.; Shi, H.; Gao, G.; Zhang, Q.; Lou, G.; Jiang, G.; He, Y. QTL Analysis for Rice Quality-Related Traits and Fine Mapping of qWCR3. Int. J. Mol. Sci. 2024, 25, 4389. https://doi.org/10.3390/ijms25084389
Liu J, Zhang H, Wang Y, Liu E, Shi H, Gao G, Zhang Q, Lou G, Jiang G, He Y. QTL Analysis for Rice Quality-Related Traits and Fine Mapping of qWCR3. International Journal of Molecular Sciences. 2024; 25(8):4389. https://doi.org/10.3390/ijms25084389
Chicago/Turabian StyleLiu, Jun, Hao Zhang, Yingying Wang, Enyu Liu, Huan Shi, Guanjun Gao, Qinglu Zhang, Guangming Lou, Gonghao Jiang, and Yuqing He. 2024. "QTL Analysis for Rice Quality-Related Traits and Fine Mapping of qWCR3" International Journal of Molecular Sciences 25, no. 8: 4389. https://doi.org/10.3390/ijms25084389
APA StyleLiu, J., Zhang, H., Wang, Y., Liu, E., Shi, H., Gao, G., Zhang, Q., Lou, G., Jiang, G., & He, Y. (2024). QTL Analysis for Rice Quality-Related Traits and Fine Mapping of qWCR3. International Journal of Molecular Sciences, 25(8), 4389. https://doi.org/10.3390/ijms25084389