
Impact of Serotonergic 5HT1A and 5HT2A Receptor Activation on the Respiratory Response to Hypercapnia in a Rat Model of Parkinson’s Disease

 , , and
, , and 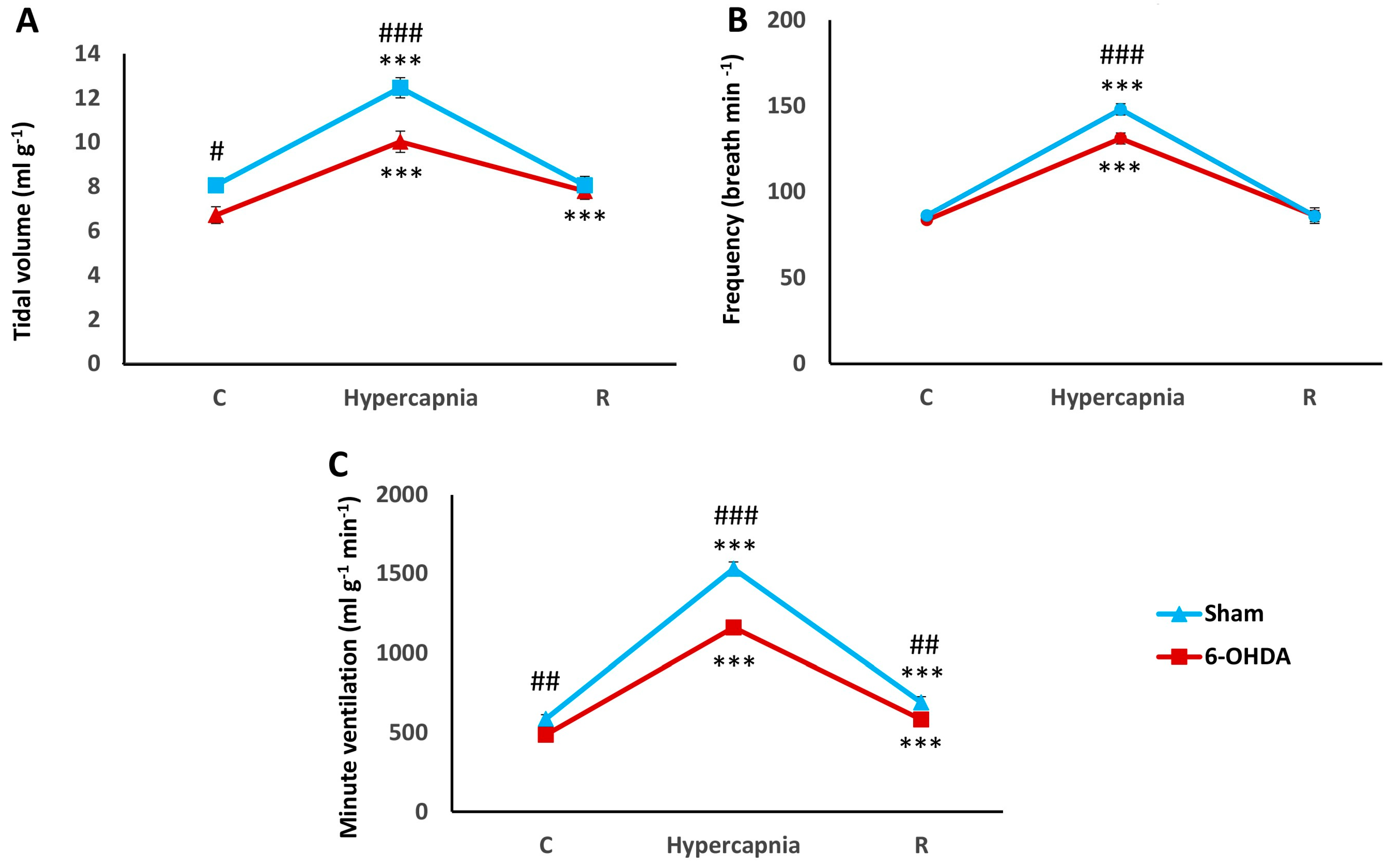{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Hypercapnic Ventilatory Response (HCVR) in PD and Sham Rats
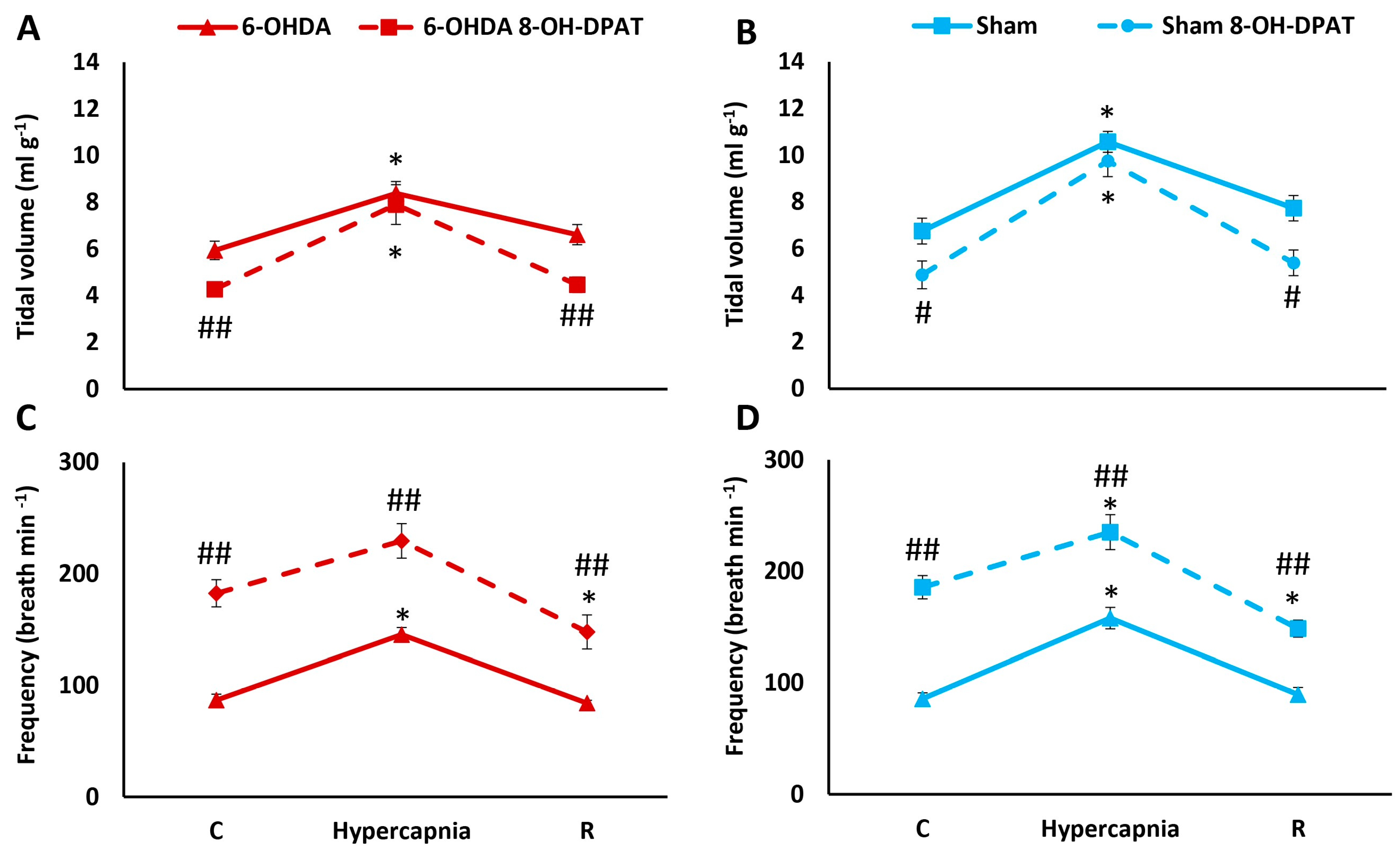
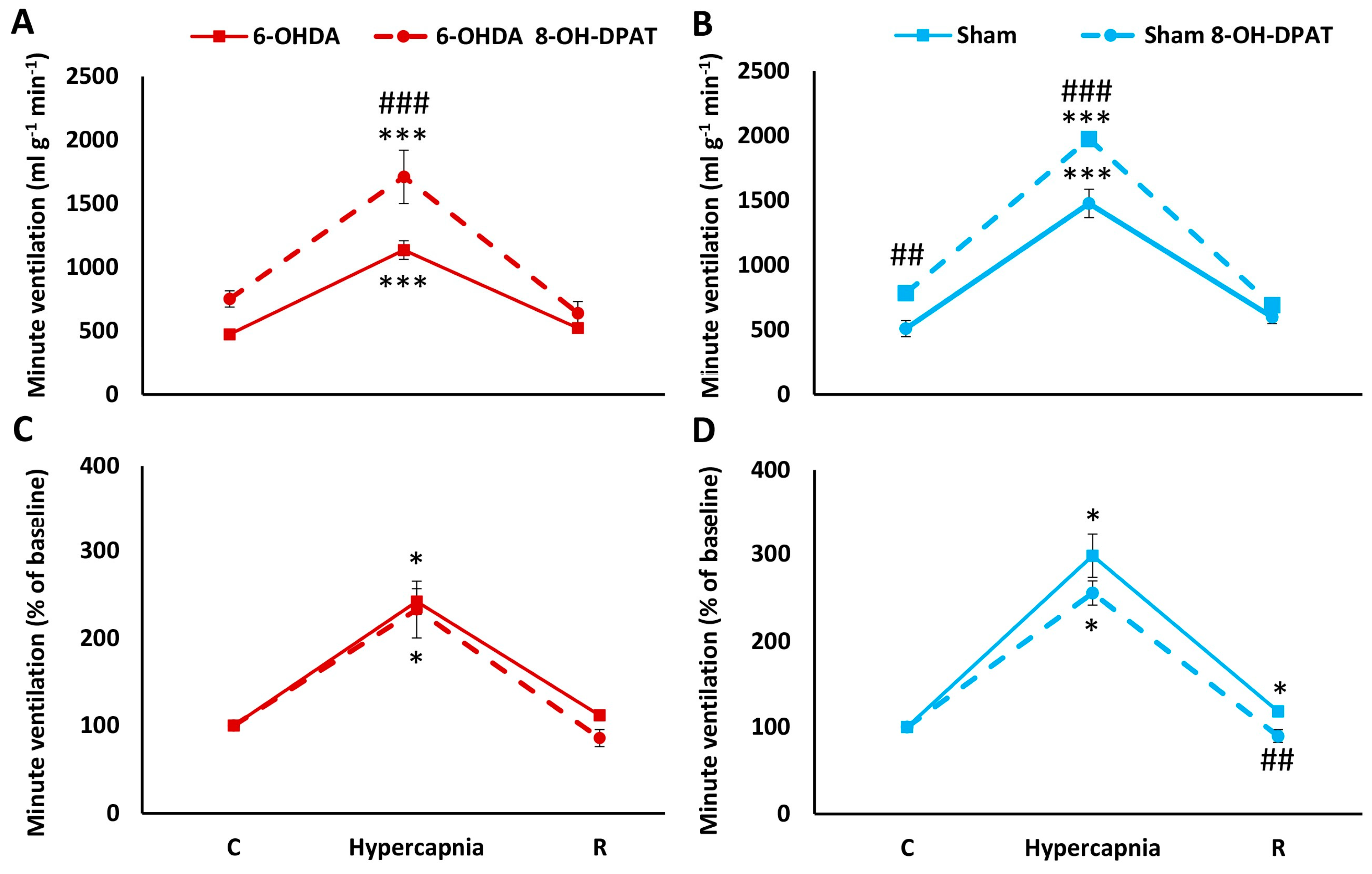
2.2. HCVR in PD and Sham Rats before and after Intraperitoneal (i.p.) Treatment with 8-OH-DPAT
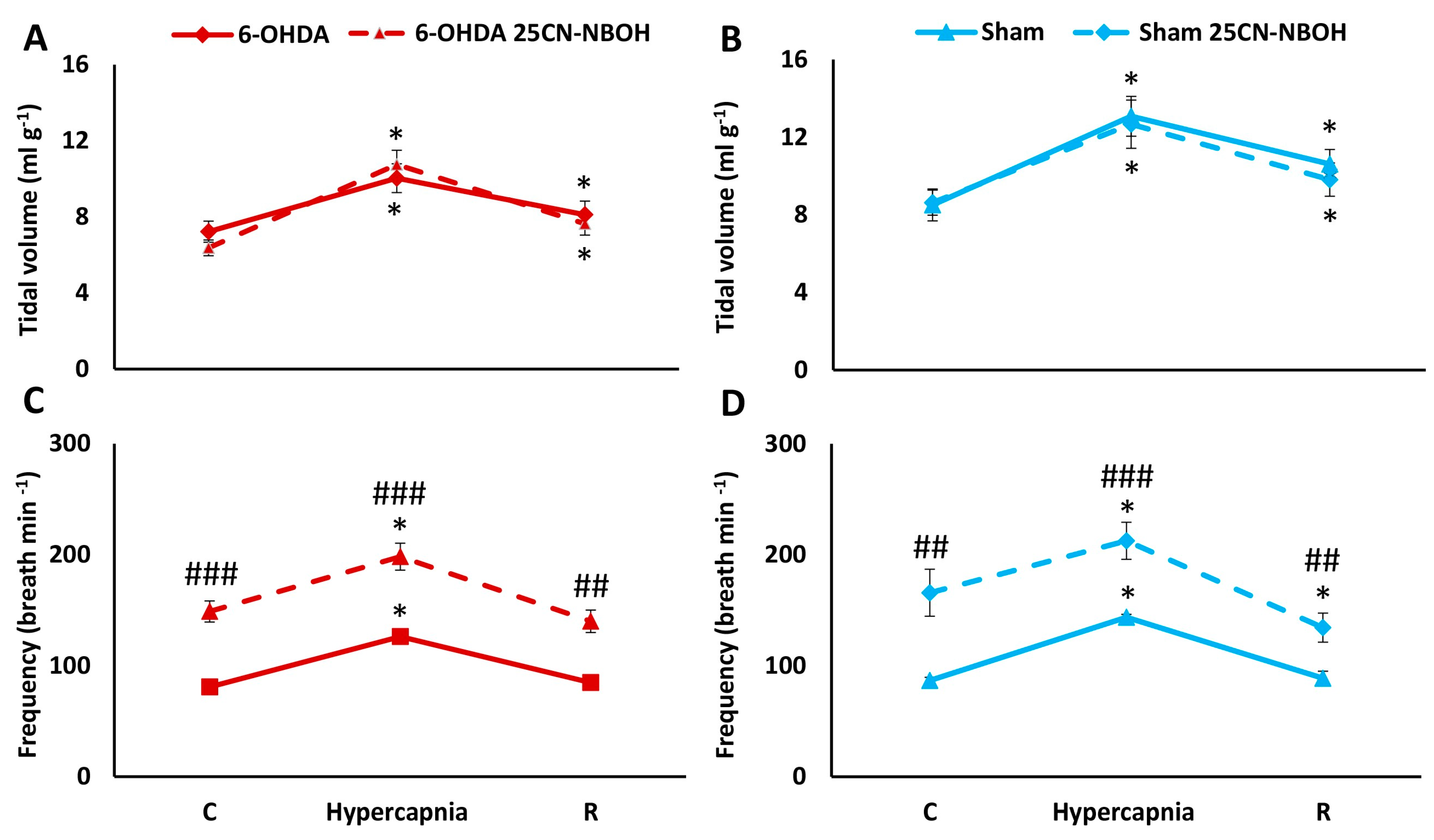
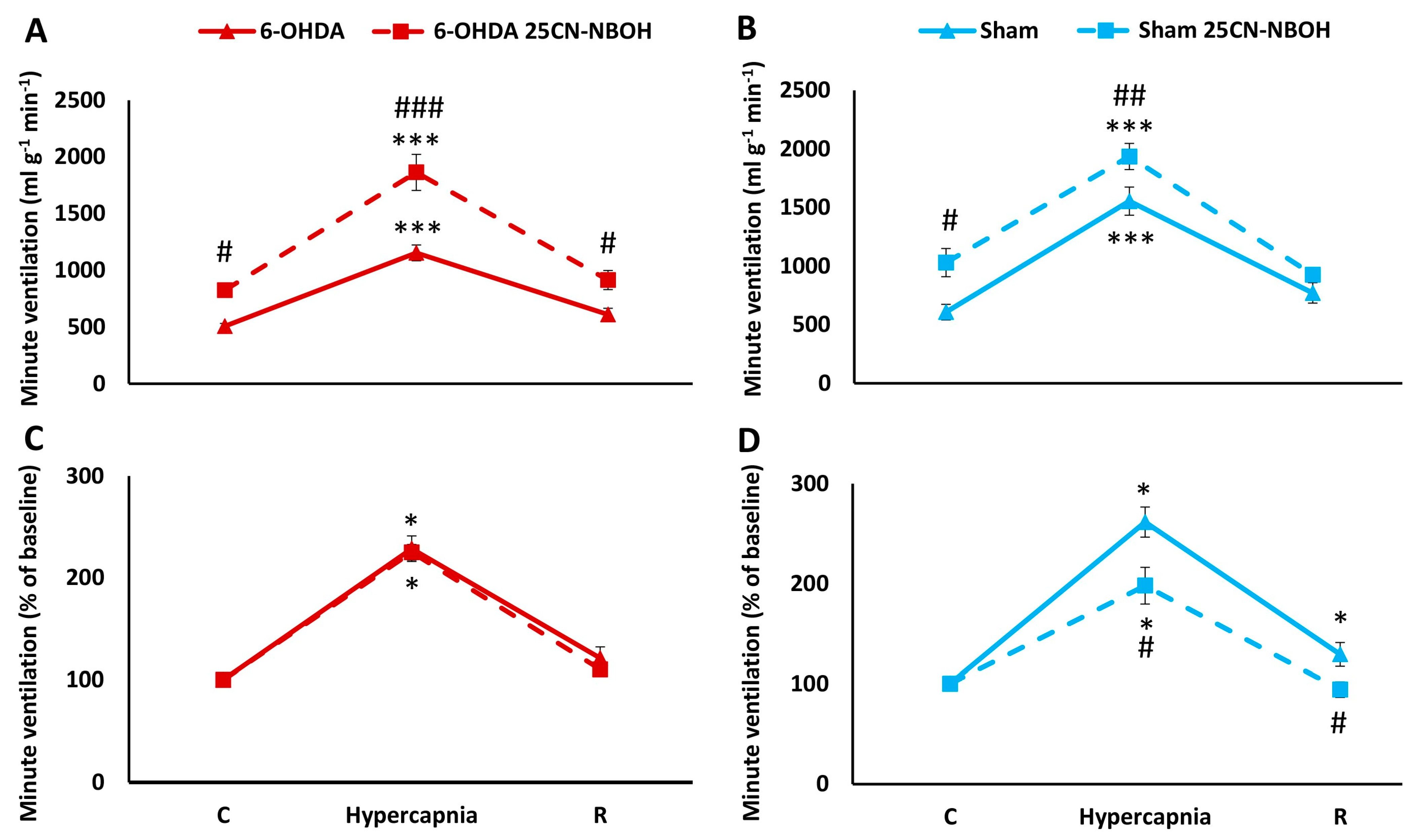
2.3. HCVR in PD and Sham Rats before and after i.p. Treatment with 25CN-NBOH
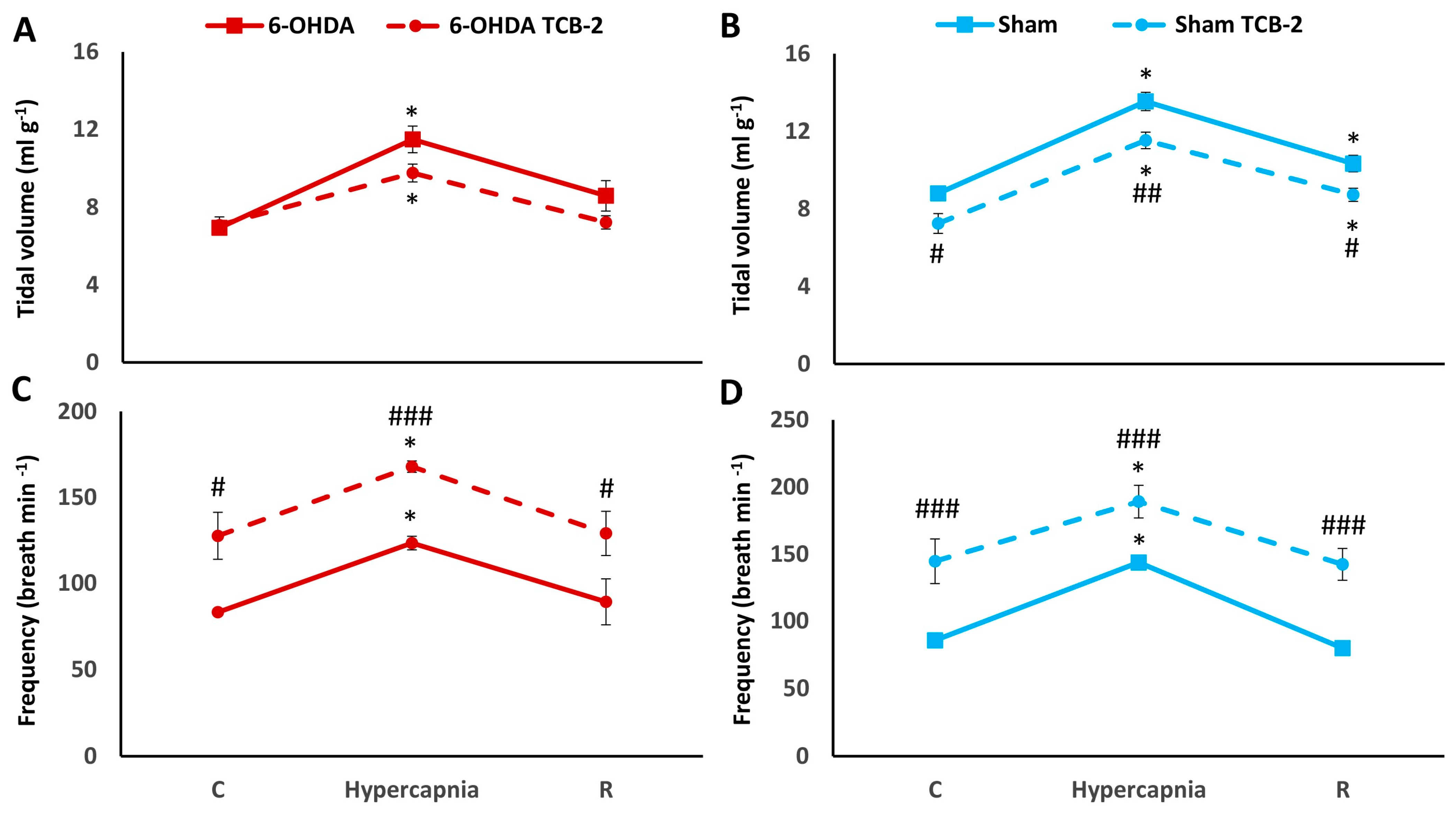
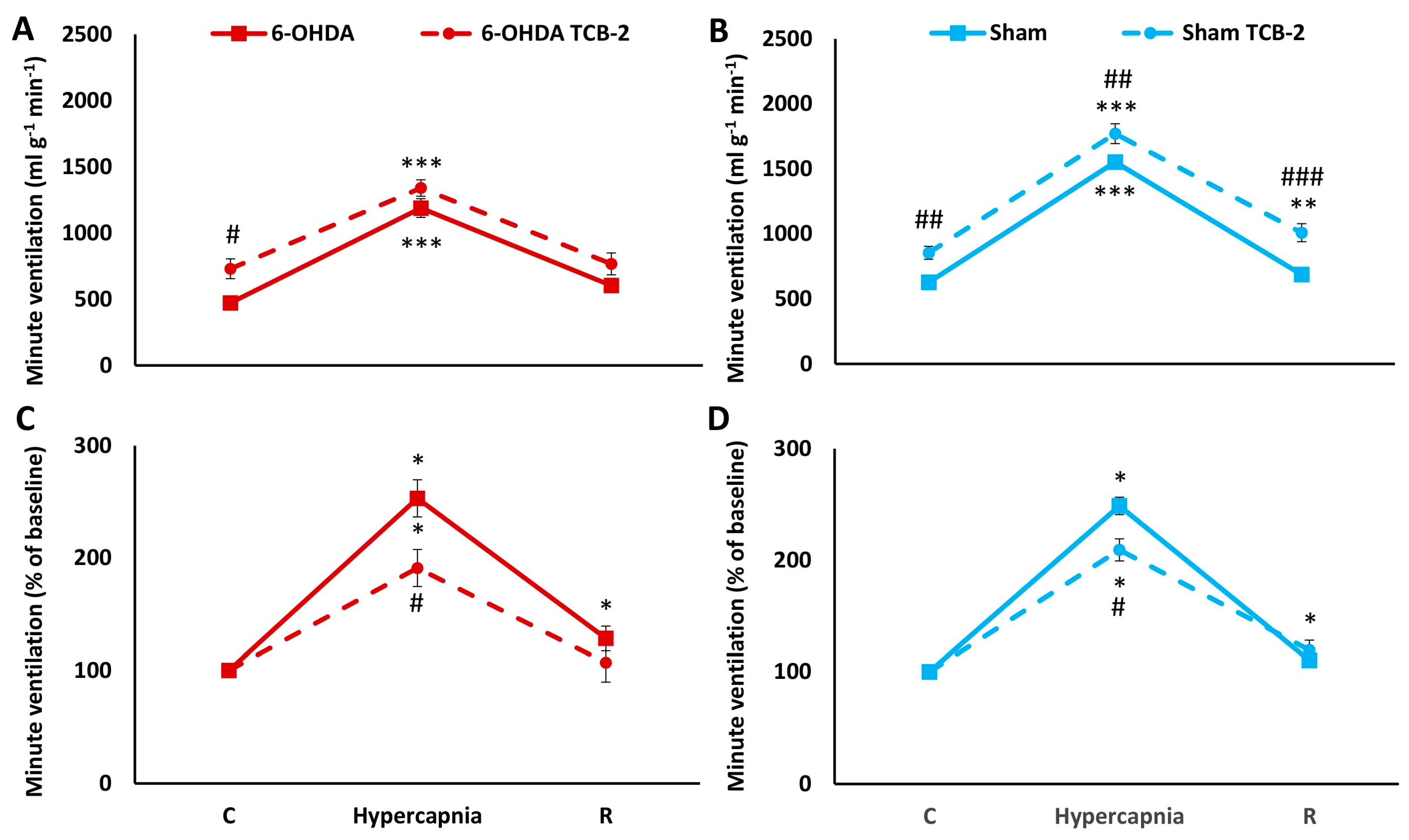
2.4. HCVR in PD and Sham Rats before and after i.p. Treatment with TCB-2
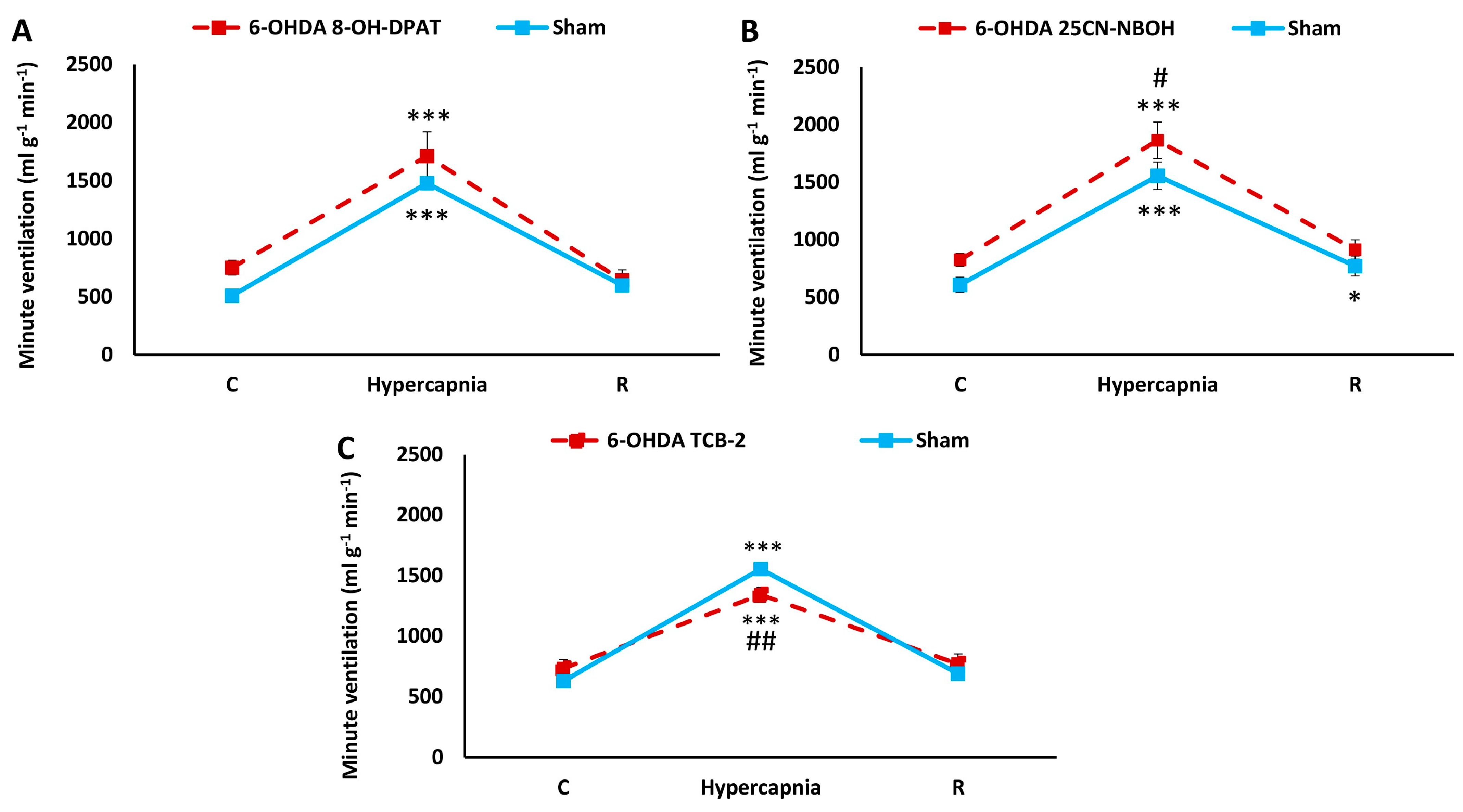
2.5. Comparison of HCVR in Sham and PD Rats after i.p. Treatment with 25CN-NBOH, TCB-2, and 8-OH-DPAT
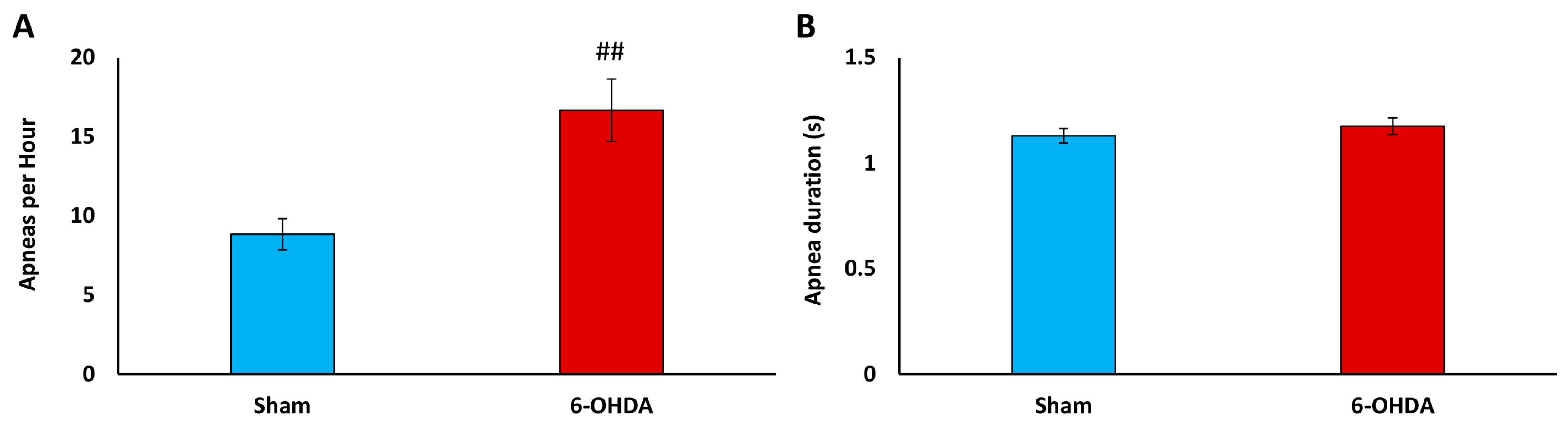
2.6. Apnea Index Analysis
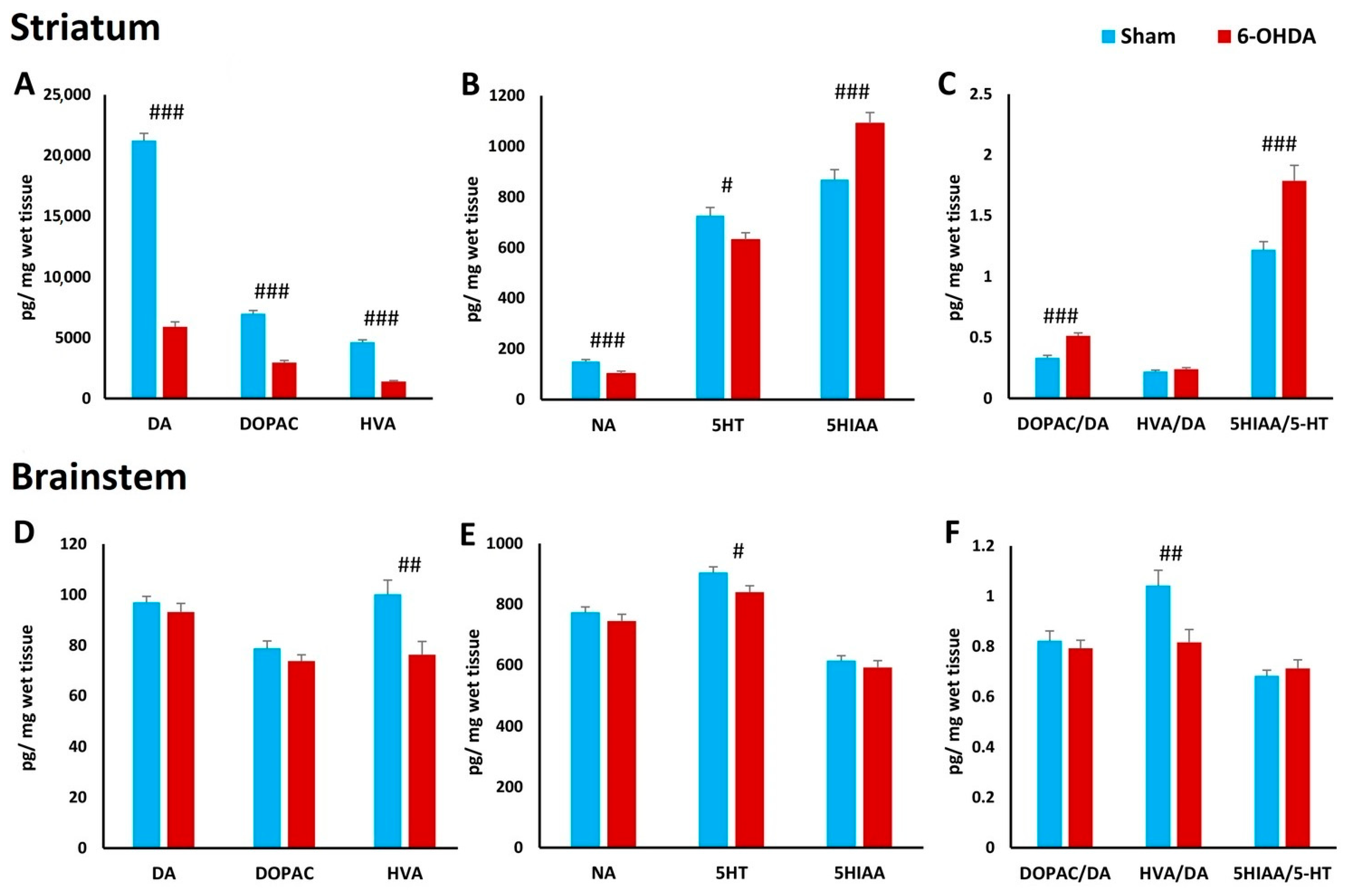
2.7. Concentration of Biogenic Amines in Striatum and Brainstem
3. Discussion
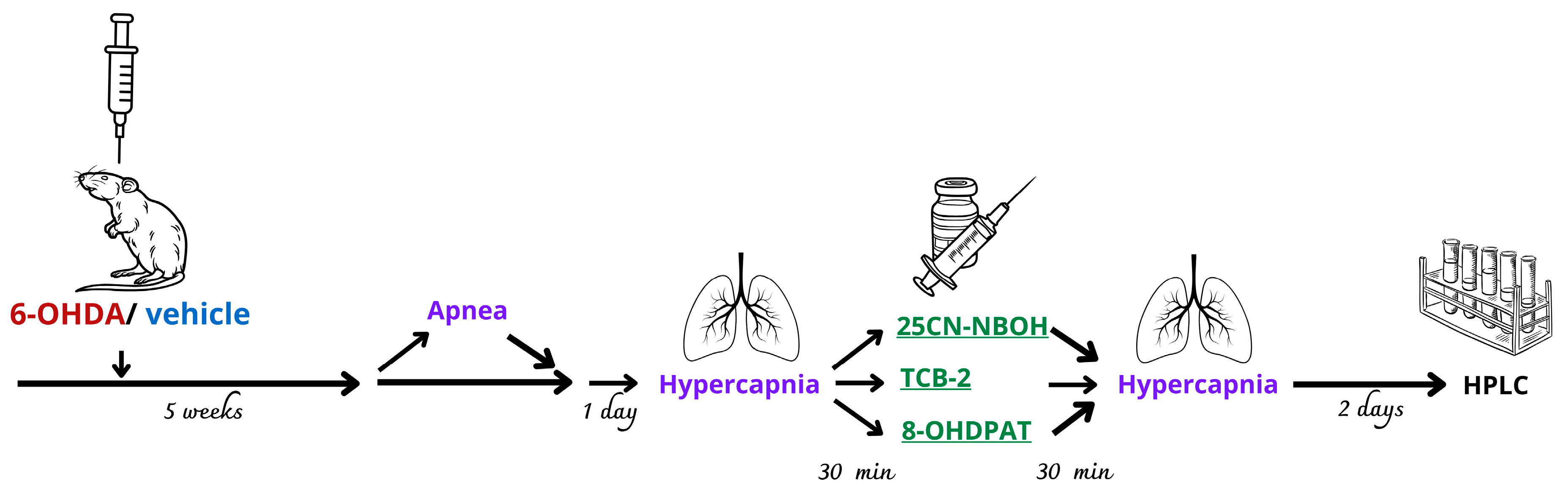
4. Material and Methods
4.1. Animals
4.2. Experimental Groups and Drug Treatment
- 25CN-NBOH at a dose of 1 mg kg−1 dissolved in water (Sham n = 7, 6-OHDA n = 7);
- TCB-2 at a dose of 1 mg kg−1 dissolved in water (Sham n = 7, 6-OHDA n = 7);
- 8-OH-DPAT at a dose of 0.5 mg kg−1 dissolved in water (Sham n = 6, 6-OHDA n = 6).
4.3. Surgery and 6-OHDA/Vehicle Injection
4.4. Ventilation Measurements
4.5. Apnea Measurement
4.6. High-Performance Liquid Chromatography (HPLC) Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahsas, G.; Chisti, W.; Verma, A.; Agrawal, N.; Bansal, K. The Role of the Serotonergic System of the Brain in the Pathogenesis of Parkinson’s Disease. Neurochem. J. 2023, 17, 30–41. [Google Scholar] [CrossRef]
- Huot, P.; Fox, S.H.; Brotchie, J.M. The serotonergic system in Parkinson’s disease. Prog. Neurobiol. 2011, 95, 163–212. [Google Scholar] [CrossRef] [PubMed]
- Politis, M.; Loane, C. Serotonergic dysfunction in Parkinson’s disease and its relevance to disability. Sci. World J. 2011, 11, 1726–1734. [Google Scholar] [CrossRef] [PubMed]
- Hilaire, G.; Voituron, N.; Menuet, C.; Ichiyama, R.M.; Subramanian, H.H.; Dutschmann, M. The role of serotonin in respiratory function and dysfunction. Respir. Physiol. Neurobiol. 2010, 174, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Szereda-Przestaszewska, M.; Kaczyńska, K. Serotonin and substance P: Synergy or competition in the control of breathing. Auton. Neurosci. 2020, 225, 102658. [Google Scholar] [CrossRef] [PubMed]
- Hodges, M.R.; Richerson, G.B. Medullary serotonin neurons and their roles in central respiratory chemoreception. Respir. Physiol. Neurobiol. 2010, 173, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Hodges, M.R.; Tattersall, G.J.; Harris, M.B.; McEvoy, S.D.; Richerson, D.N.; Deneris, E.S.; Johnson, R.L.; Chen, Z.-F.; Richerson, G.B. Defects in breathing and thermoregulation in mice with near-complete absence of central serotonin neurons. J. Neurosci. 2008, 28, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Nattie, E.E.; Li, A.; Richerson, G.; Lappi, D.A. Medullary serotonergic neurones and adjacent neurones that express neurokinin-1 receptors are both involved in chemoreception in vivo. J. Physiol. 2004, 556, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Brust, R.D.; Corcoran, A.E.; Richerson, G.B.; Nattie, E.; Dymecki, S.M. Functional and developmental identification of a molecular subtype of brain serotonergic neuron specialized to regulate breathing dynamics. Cell Rep. 2014, 9, 2152–2165. [Google Scholar] [CrossRef]
- Mateika, J.H.; Komnenov, D.; Pop, A.; Kuhn, D.M. Genetic depletion of 5-HT increases central apnea frequency and duration and dampens arousal but does not impact the circadian modulation of these variables. J. Appl. Physiol. 2019, 126, 1–10. [Google Scholar] [CrossRef]
- Kubin, L.; Mann, G.L. Hypoglossal motoneurons are endogenously activated by serotonin during the active period of circadian cycle. Respir. Physiol. Neurobiol. 2018, 248, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Baille, G.; De Jesus, A.M.; Perez, T.; Devos, D.; Dujardin, K.; Charley, C.M.; Defebvre, L.; Moreau, C. Ventilatory Dysfunction in Parkinson’s Disease. J. Park. Dis. 2016, 6, 463–471. [Google Scholar] [CrossRef]
- Guilherme, E.M.; Moreira, R.D.F.C.; De Oliveira, A.; Ferro, A.M.; DILorenzo, V.A.P.; Gianlorenço, A.C.L. Respiratory Disorders in Parkinson’s Disease. J. Park. Dis. 2021, 11, 993–1010. [Google Scholar] [CrossRef] [PubMed]
- Kaczyńska, K.; Orłowska, M.E.; Andrzejewski, K. Respiratory Abnormalities in Parkinson’s Disease: What Do We Know from Studies in Humans and Animal Models? Int. J. Mol. Sci. 2022, 23, 3499. [Google Scholar] [CrossRef] [PubMed]
- Aquino, Y.C.; Cabral, L.M.; Miranda, N.C.; Naccarato, M.C.; Falquetto, B.; Moreira, T.S.; Takakura, A.C. Respiratory disorders of Parkinson’s disease. J. Neurophysiol. 2022, 127, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewski, K.; Jampolska, M.; Zaremba, M.; Joniec-Maciejak, I.; Boguszewski, P.M.; Kaczyńska, K. Respiratory pattern and phrenic and hypoglossal nerve activity during normoxia and hypoxia in 6-OHDA-induced bilateral model of Parkinson’s disease. J. Physiol. Sci. 2020, 70, 16. [Google Scholar] [CrossRef] [PubMed]
- Tintner, R.; Manian, P.; Gauthier, P.; Jankovic, J. Pleuropulmonary fibrosis after long-term treatment with the dopamine agonist pergolide for Parkinson Disease. Arch. Neurol. 2005, 62, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewski, K.; Kaczyńska, K.; Zaremba, M. Serotonergic system in hypoxic ventilatory response in unilateral rat model of Parkinson’s disease. J. Biomed. Sci. 2017, 24, 24. [Google Scholar] [CrossRef] [PubMed]
- Tuppy, M.; Barna, B.F.; Alves-dos-Santos, L.; Britto, L.R.G.; Chiavegatto, S.; Moreira, T.S.; Takakura, A. Respiratory deficits in a rat model of Parkinson’s disease. Neuroscience 2015, 297, 194–204. [Google Scholar] [CrossRef]
- Taylor, N.C.; Li, A.; Nattie, E.E. Medullary serotonergic neurones modulate the ventilatory response to hypercapnia, but not hypoxia in conscious rats. J. Physiol. 2005, 566, 543–557. [Google Scholar] [CrossRef]
- Corcoran, A.E.; Richerson, G.B.; Harris, M.B. Serotonergic mechanisms are necessary for central respiratory chemoresponsiveness in situ. Respir. Physiol. Neurobiol. 2013, 186, 214–220. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salloum, A.; Rowley, J.A.; Mateika, J.H.; Chowdhuri, S.; Omran, Q.; Badr, M.S. Increased propensity for central apnea in patients with obstructive sleep apnea: Effect of nasal continuous positive airway pressure. Am. J. Respir. Crit. Care Med. 2010, 181, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Aung, O.; Amorim, M.R.; Mendelowitz, D.; Polotsky, V.Y. Revisiting the Role of Serotonin in Sleep-Disordered Breathing. Int. J. Mol. Sci. 2024, 25, 1483. [Google Scholar] [CrossRef]
- Teran, F.A.; Massey, C.A.; Richerson, G.B. Serotonin neurons and central respiratory chemoreception: Where are we now? Prog. Brain Res. 2014, 209, 207–233. [Google Scholar] [CrossRef]
- Gonye, E.C.; Bayliss, D.A. Criteria for central respiratory chemoreceptors: Experimental evidence supporting current candidate cell groups. Front. Physiol. 2023, 14, 1241662. [Google Scholar] [CrossRef]
- De Moreno, V.S.; Bícego, K.C.; Szawka, R.E.; Anselmo-Franci, J.A.; Gargaglioni, L.H. Serotonergic mechanisms on breathing modulation in the rat locus coeruleus. Pflug. Arch. 2010, 459, 357–368. [Google Scholar] [CrossRef]
- Levitt, E.S.; Hunnicutt, B.J.; Knopp, S.J.; Williams, J.T.; Bissonnette, J.M. A selective 5-HT1a receptor agonist improves respiration in a mouse model of Rett syndrome. J. Appl. Physiol. 2013, 115, 1626–1633. [Google Scholar] [CrossRef]
- Cummings, K.J. Eupnea and gasping in vivo are facilitated by the activation of 5-HT2A receptors. J. Neurophysiol. 2021, 125, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Joyal, K.G.; Petrucci, A.N.; Littlepage-Saunders, M.V.; Boodhoo, N.A.; Wendt, L.H.; Buchanan, G.F. Selective Serotonin Reuptake Inhibitors and 5-HT2 Receptor Agonists Have Distinct, Sleep-state Dependent Effects on Postictal Breathing in Amygdala Kindled Mice. Neuroscience 2023, 513, 76–95. [Google Scholar] [CrossRef]
- Szereda-Przestaszewska, M.; Kaczyńska, K. Peripheral 5-HT1A receptors are not essential for increased ventilation evoked by systemic 8-OH-DPAT challenge in anaesthetized rats. Exp. Physiol. 2007, 92, 953–961. [Google Scholar] [CrossRef]
- Radocaj, T.; Mustapic, S.; Prkic, I.; Stucke, A.G.; Hopp, F.A.; Stuth, E.A.E.; Zuperku, E.J. Activation of 5-HT1A receptors in the preBötzinger region has little impact on the respiratory pattern. Respir. Physiol. Neurobiol. 2015, 212–214, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Xu, F. Systemic 8-OH-DPAT challenge causes hyperventilation largely via activating pre-botzinger complex 5-HT1A receptors. Respir. Physiol. Neurobiol. 2022, 296, 103810. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Zhang, Z.; Zhang, C.; Xu, F. 8-OH-DPAT abolishes the pulmonary C-fiber-mediated apneic response to fentanyl largely via acting on 5HT1A receptors in the nucleus tractus solitarius. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R449–R458. [Google Scholar] [CrossRef] [PubMed]
- Kline, A.E.; Wagner, A.K.; Westergom, B.P.; Malena, R.R.; Zafonte, R.D.; Olsen, A.S.; Sozda, C.N.; Luthra, P.; Panda, M.; Cheng, J.P.; et al. Acute treatment with the 5-HT(1A) receptor agonist 8-OH-DPAT and chronic environmental enrichment confer neurobehavioral benefit after experimental brain trauma. Behav. Brain Res. 2007, 177, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.P.; Leary, J.B.; Sembhi, A.; Edwards, C.M.; Bondi, C.O.; Kline, A.E. 5-hydroxytryptamine1A (5-HT1A) receptor agonists: A decade of empirical evidence supports their use as an efficacious therapeutic strategy for brain trauma. Brain Res. 2016, 1640, 5–14. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Vry, J.; Schreiber, R.; Melon, C.; Dalmus, M.; Jentzsch, K.R. 5-HT1A receptors are differentially involved in the anxiolytic- and antidepressant-like effects of 8-OH-DPAT and fluoxetine in the rat. Eur. Neuropsychopharmacol. 2004, 14, 487–495. [Google Scholar] [CrossRef]
- Ślifirski, G.; Król, M.; Turło, J. 5-HT Receptors and the Development of New Antidepressants. Int. J. Mol. Sci. 2021, 22, 9015. [Google Scholar] [CrossRef]
- Kung, H.F.; Frederick, D.; Kim, H.; McElgin, W.; Kung, M.P.; Mu, M.; Mozley, P.D.; Vessotskie, J.M.; Stevenson, D.A.; Kushner, S.A.; et al. In Vivo SPECT Imaging of 5-HTlA Receptors With [1231]p-MPPI in Nonhuman Primates. Synapse 1996, 24, 273–281. [Google Scholar] [CrossRef]
- Peña, F.; Ramirez, J.M. Endogenous activation of serotonin-2A receptors is required for respiratory rhythm generation in vitro. J. Neurosci. 2002, 22, 11055–11064. [Google Scholar] [CrossRef]
- Hodges, M.R.; Wehner, M.; Aungst, J.; Smith, J.C.; Richerson, G.B. Transgenic mice lacking serotonin neurons have severe apnea and high mortality during development. J. Neurosci. 2009, 29, 10341–10349. [Google Scholar] [CrossRef]
- Ptak, K.; Yamanishi, T.; Aungst, J.; Milescu, L.S.; Zhang, R.; Richerson, G.B.; Smith, J.C. Raphé neurons stimulate respiratory circuit activity by multiple mechanisms via endogenously released serotonin and substance P. J. Neurosci. 2009, 29, 3720–3737. [Google Scholar] [CrossRef] [PubMed]
- Brandes, I.F.; Zuperku, E.J.; Stucke, A.G.; Jakovcevic, D.; Hopp, F.A.; Stuth, E.A.E. Serotonergic modulation of inspiratory hypoglossal motoneurons in decerebrate dogs. J. Neurophysiol. 2006, 95, 3449–3459. [Google Scholar] [CrossRef] [PubMed]
- Märcher Rørsted, E.; Jensen, A.A.; Kristensen, J.L. 25CN-NBOH: A Selective Agonist for in vitro and in vivo Investigations of the Serotonin 2A Receptor. ChemMedChem 2021, 16, 3263–3270. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, G.; De Deurwaerdère, P. TCB-2 [(7R)-3-bromo-2,5-dimethoxy-bicyclo[4.2.0]octa-1,3,5-trien-7-yl]methanamine]: A hallucinogenic drug, a selective 5-HT2A receptor pharmacological tool, or none of the above? Neuropharmacology 2018, 142, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Fernandes-Junior, S.A.; Carvalho, K.S.; Moreira, T.S.; Takakura, A.C. Correlation between neuroanatomical and functional respiratory changes observed in an experimental model of Parkinson’s disease. Exp. Physiol. 2018, 103, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.C.; Oliveira, L.M.; Botelho, M.T.; Moreira, T.S.; Takakura, A.C. The involvement of the pathway connecting the substantia nigra, the periaqueductal gray matter and the retrotrapezoid nucleus in breathing control in a rat model of Parkinson’s disease. Exp. Neurol. 2018, 302, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Navailles, S.; De Deurwaerdère, P. Presynaptic control of serotonin on striatal dopamine function. Psychopharmacology 2011, 213, 213–242. [Google Scholar] [CrossRef] [PubMed]
- Gourine, A.V.; Llaudet, E.; Dale, N.; Spyer, K.M. ATP is a mediator of chemosensory transduction in the central nervous system. Nature 2005, 436, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Wenker, I.C.; Kréneisz, O.; Nishiyama, A.; Mulkey, D.K. Astrocytes in the retrotrapezoid nucleus sense H+ by inhibition of a Kir4.1-Kir5.1-like current and may contribute to chemoreception by a purinergic mechanism. J. Neurophysiol. 2010, 104, 3042–3052. [Google Scholar] [CrossRef]
- Joseph, V.; Laouafa, S.; Marcouiller, F.; Roussel, D.; Pialoux, V.; Bairam, A. Progesterone decreases apnoea and reduces oxidative stress induced by chronic intermittent hypoxia in ovariectomized female rats. Exp. Physiol. 2020, 105, 1025–1034. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrzejewski, K.; Orłowska, M.E.; Zaremba, M.; Joniec-Maciejak, I.; Kaczyńska, K. Impact of Serotonergic 5HT1A and 5HT2A Receptor Activation on the Respiratory Response to Hypercapnia in a Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2024, 25, 4403. https://doi.org/10.3390/ijms25084403
Andrzejewski K, Orłowska ME, Zaremba M, Joniec-Maciejak I, Kaczyńska K. Impact of Serotonergic 5HT1A and 5HT2A Receptor Activation on the Respiratory Response to Hypercapnia in a Rat Model of Parkinson’s Disease. International Journal of Molecular Sciences. 2024; 25(8):4403. https://doi.org/10.3390/ijms25084403
Chicago/Turabian StyleAndrzejewski, Kryspin, Magdalena E. Orłowska, Małgorzata Zaremba, Ilona Joniec-Maciejak, and Katarzyna Kaczyńska. 2024. "Impact of Serotonergic 5HT1A and 5HT2A Receptor Activation on the Respiratory Response to Hypercapnia in a Rat Model of Parkinson’s Disease" International Journal of Molecular Sciences 25, no. 8: 4403. https://doi.org/10.3390/ijms25084403
APA StyleAndrzejewski, K., Orłowska, M. E., Zaremba, M., Joniec-Maciejak, I., & Kaczyńska, K. (2024). Impact of Serotonergic 5HT1A and 5HT2A Receptor Activation on the Respiratory Response to Hypercapnia in a Rat Model of Parkinson’s Disease. International Journal of Molecular Sciences, 25(8), 4403. https://doi.org/10.3390/ijms25084403