
A Principal Components Analysis and Functional Annotation of Differentially Expressed Genes in Brain Regions of Gray Rats Selected for Tame or Aggressive Behavior

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
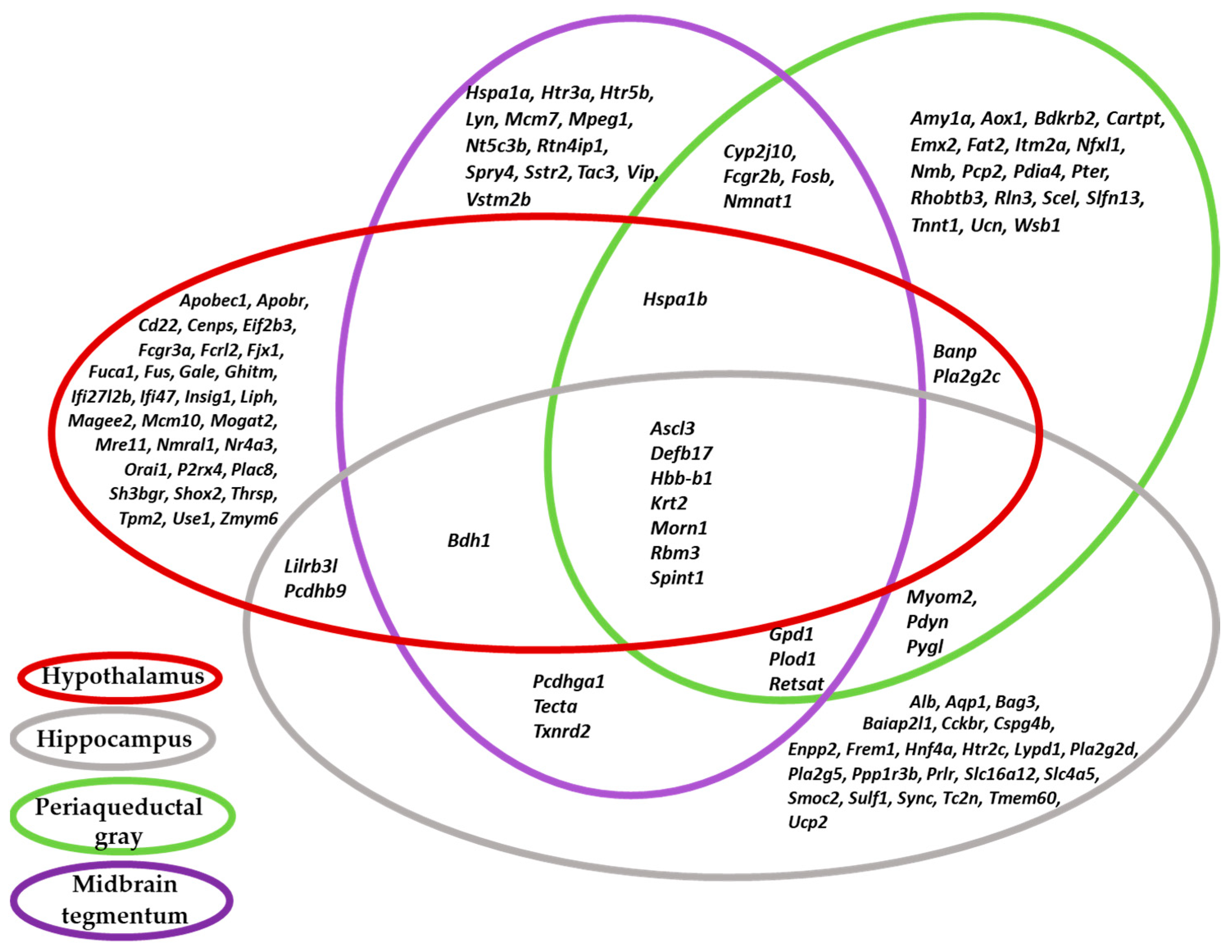
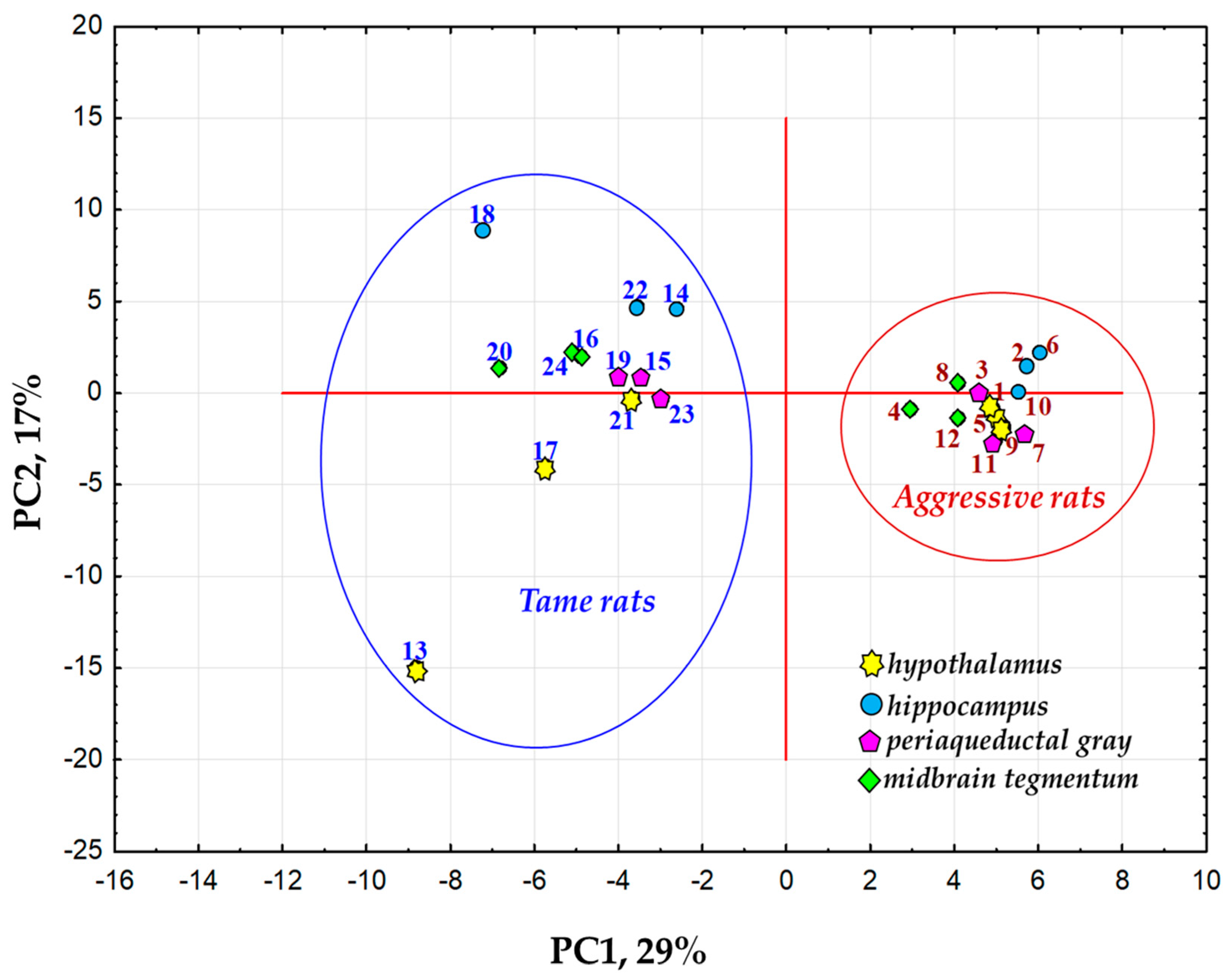
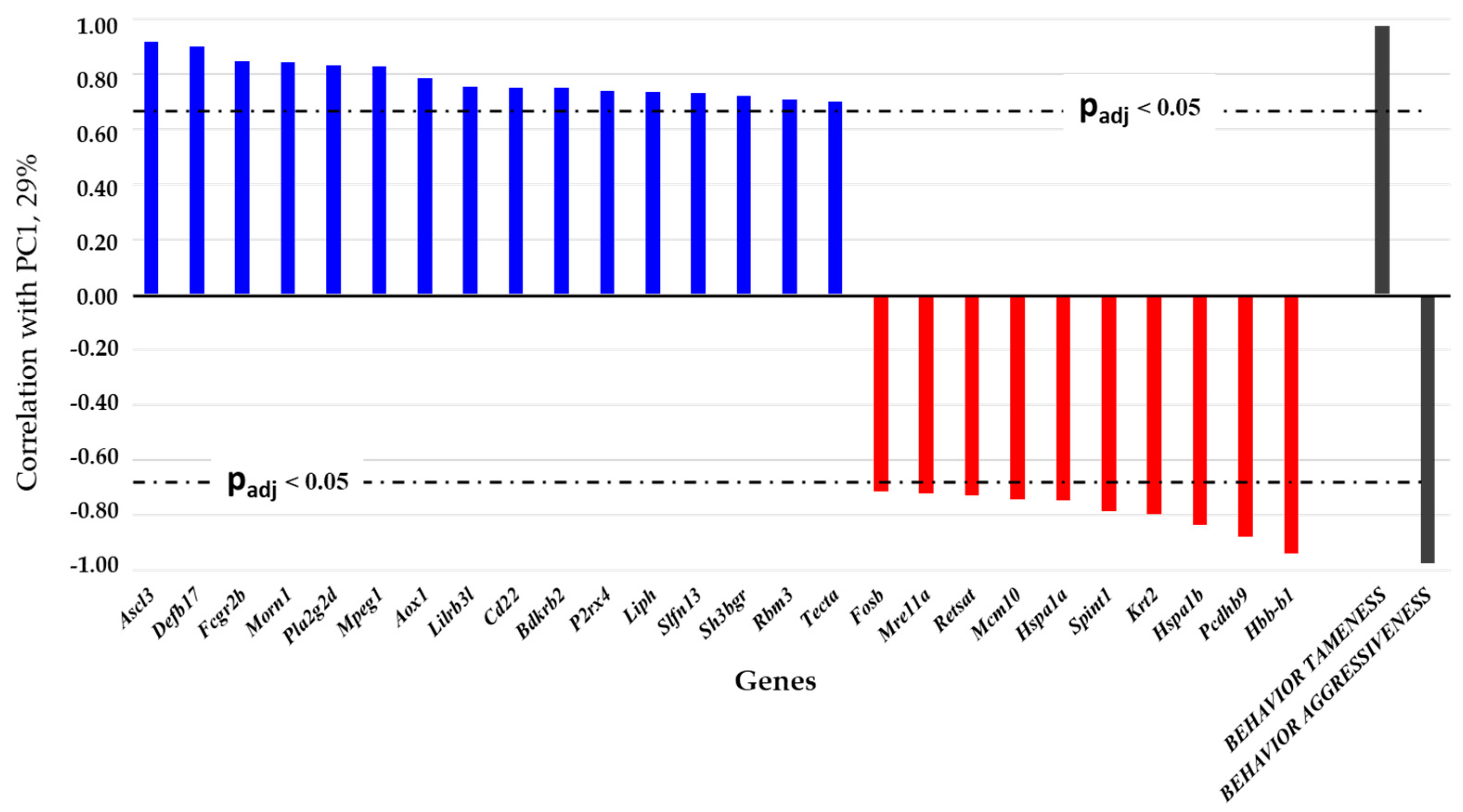
2.1. Examination of the List of DEGs from Brain Samples of Tame and Aggressive Rats by Principal Component Analysis (PCA)
2.2. Functional Annotation of DEGs in the Brain Regions of Tame and Aggressive Rats
2.3. Gene Association Networks Describing Interactions of DEGs in the Brain Regions of Tame and Aggressive Rats
3. Discussion
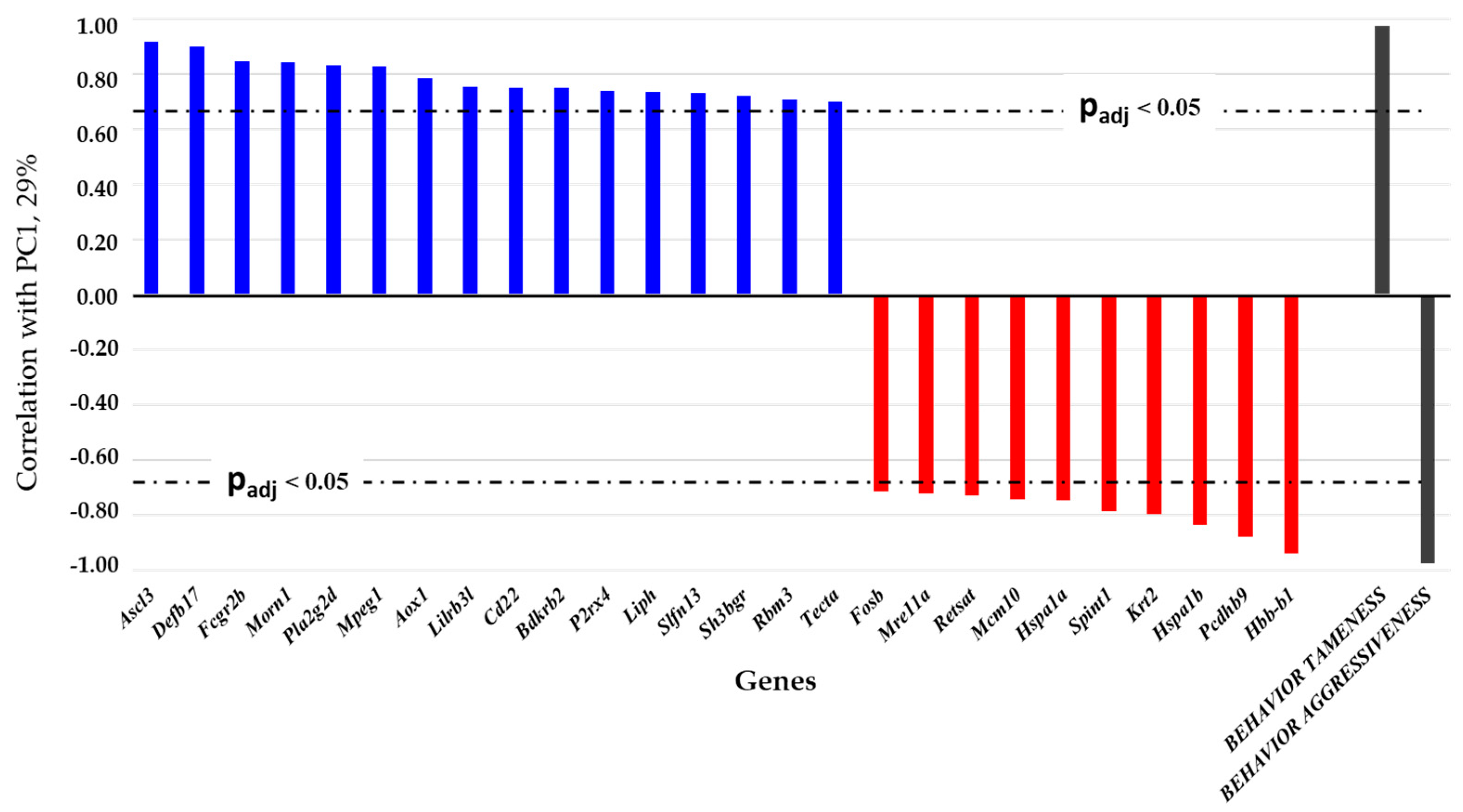
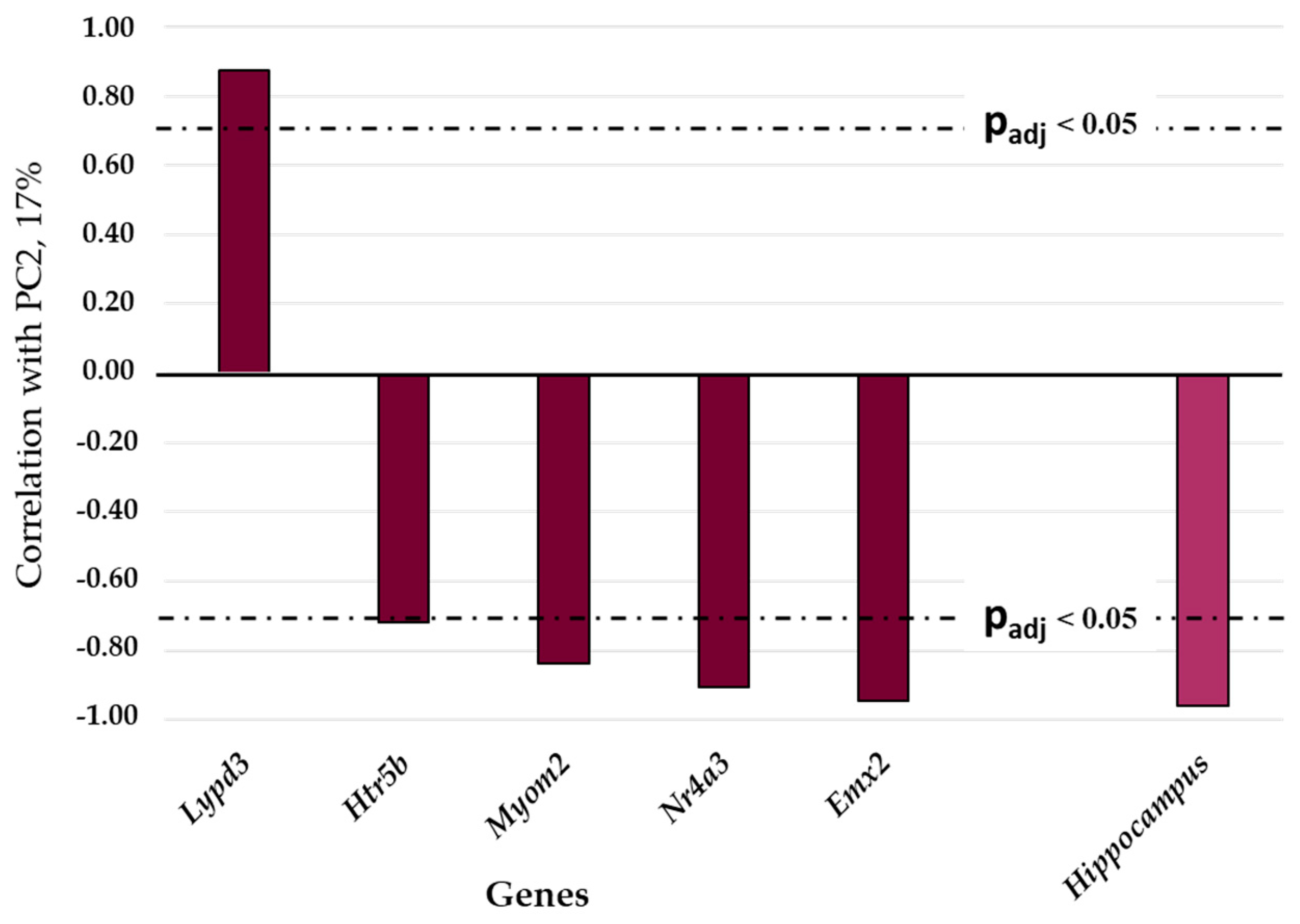
3.1. Relations of DEGs in the Brain Regions of Gray Rats with Behavior According to the PC1 and PC2
3.2. Relations of DEGs in the Brain Regions of Tame and Aggressive Rats with GO and KEGG Terms
4. Materials and Methods
4.1. Materials
4.2. Statistical Analysis
4.3. Functional Analysis of DEGs and Construction of Gene Association Networks
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DEG | differentially expressed gene |
| ECR | extracellular region |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| MTg | tegmental region of the midbrain |
| PAG | periaqueductal gray matter |
| PCA | principal component analysis |
| PC1 | first principal component |
| PC2 | second principal component |
| TF | transcription factor |
References
- Belyaev, D.K.; Plyusnina, I.Z.; Trut, L.N. Domestication in the silver fox (Vulpes fulvus Desm.)—Changes in physiological boundaries of the sensitive period of primary socialization. Appl. Anim. Behav. Sci. 1984, 13, 359–370. [Google Scholar] [CrossRef]
- Zeder, M.A. Pathways to animal domestication. In Biodiversity in Agriculture: Domestication, Evolution and Sustainability; Gepts, P., Famula, T.R., Bettinger, R.L., Bush, S.B., Damania, A.B., McGuire, P.E., Qualset, C.O., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 227–259. [Google Scholar]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “domestication syndrome” in mammals: A unified explanation based on neural crest cell behavior and genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Rubio, A.O.; Summers, K. Neural crest cell genes and the domestication syndrome: A comparative analysis of selection. PLoS ONE 2022, 17, e0263830. [Google Scholar] [CrossRef]
- Lu, C.; Li, M.; Sun, X.; Li, N.; Wang, W.; Tong, P.; Dai, J. Comparing the hippocampal miRNA expression profiles of wild and domesticated Chinese tree shrews (Tupaia belangeri chinensis). BMC Ecol. Evol. 2021, 21, 12. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Nagayama, H.; Nakaoka, H.; Toyoda, A.; Goto, T.; Koide, T. Combined change of behavioral traits for domestication and gene-networks in mice selectively bred for active tameness. Genes Brain Behav. 2021, 20, e12721. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Slomianka, L.; Farmer, A.J.; Kharlamova, A.V.; Gulevich, R.G.; Herbeck, Y.E.; Trut, L.N.; Wolfer, D.P.; Amrein, I. Selection for tameness, a key behavioral trait of domestication, increases adult hippocampal neurogenesis in foxes. Hippocampus 2015, 25, 963–975. [Google Scholar] [CrossRef]
- Wang, X.; Pipes, L.; Trut, L.N.; Herbeck, Y.; Vladimirova, A.V.; Gulevich, R.G.; Kharlamova, A.V.; Johnson, J.L.; Acland, G.M.; Kukekova, A.V.; et al. Genomic responses to selection for tame/aggressive behaviors in the silver fox (Vulpes vulpes). Proceed. Nat. Acad. Sci. USA 2018, 115, 10398–10403. [Google Scholar] [CrossRef] [PubMed]
- Long, K.; Mao, K.; Che, T.; Zhang, J.; Qiu, W.; Wang, Y.; Tang, Q.; Ma, J.; Li, M.; Li, X. Transcriptome differences in frontal cortex between wild boar and domesticated pig. Anim. Sci. J. 2018, 89, 848–857. [Google Scholar] [CrossRef]
- Lischinsky, J.E.; Lin, D. Neural mechanisms of aggression across species. Nat. Neurosci. 2020, 23, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Plyusnina, I.; Oskina, I. Behavioral and adrenocortical responses to Open-Field test in rats selected for reduced aggressiveness toward humans. Physiol. Behav. 1997, 61, 381–385. [Google Scholar] [CrossRef]
- Plyusnina, I.Z.; Oskina, I.N.; Tibeikina, M.A.; Popova, N.K. Cross-fostering effects on weight, exploratory activity, acoustic startle reflex and corticosterone stress response in Norway gray rats selected for elimination and for enhancement of aggressiveness towards human. Behav. Genet. 2009, 39, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Plyusnina, I.Z.; Solov’eva, M.Y.; Oskina, I.N. Effect of domestication on aggression in gray Norway rats. Behav. Genet. 2011, 41, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.S.; Kozhemyakina, R.V.; Plyusnina, I.F.; Kulikov, A.V.; Popova, N.K. Serotonin 5-HT1A receptor in infancy-onset aggression: Comparison with genetically defined aggression in adult rats. Behav. Brain. Res. 2013, 243, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, D.C.; Popova, N.K.; Plyusnina, I.; Velichko, I.L.; Campbell, D.; Blanchard, R.J.; Nikulina, J.; Nikulina, E.M. Defensive reactions of “Wild-Type” and “Domesticated” wild rats to approach and contact by a threat stimulus. Aggress. Behav. 1994, 20, 387–397. [Google Scholar] [CrossRef]
- Plyusnina, I.Z.; Shchepina, O.A.; Os’kina, I.N.; Trut, L.N. Some features of learning in the Morris water test in rats selected for responses to humans. Neurosci. Behav. Physiol. 2008, 38, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Gulevich, R.; Kozhemyakina, R.; Shikhevich, S.; Konoshenko, M.; Herbeck, Y. Aggressive behavior and stress response after oxytocin administration in male Norway rats selected for different attitudes to humans. Physiol. Behav. 2019, 199, 210–218. [Google Scholar] [CrossRef]
- Albert, F.W.; Shchepina, O.; Winter, C.; Römpler, H.; Teupser, D.; Palme, R.; Ceglarek, U.; Kratzsch, J.; Sohr, R.; Trut, L.N.; et al. Phenotypic differences in behavior, physiology and neurochemistry between rats selected for tameness and for defensive aggression towards humans. Horm. Behav. 2008, 53, 413–421. [Google Scholar] [CrossRef]
- Heyne, H.O.; Lautenschlager, S.; Nelson, R.; Besnier, F.; Rotival, M.; Cagan, A.; Kozhemyakina, R.; Plyusnina, I.Z.; Trut, L.; Carlborg, Ö.; et al. Genetic influences on brain gene expression in rats selected for tameness and aggression. Genetics 2014, 198, 1277–1290. [Google Scholar] [CrossRef]
- Ilchibaeva, T.V.; Tsybko, A.S.; Kondaurova, E.M.; Kovetskaya, A.I.; Kozhemyakina, R.V.; Naumenko, V.S. Expression patterns of serotonin receptors 1a and 7 in the brain of rats with genetically determined fear-induced aggressive behavior or the lack of aggression. Neurochem. J. 2020, 14, 180–186. [Google Scholar] [CrossRef]
- Chadaeva, I.; Ponomarenko, P.; Kozhemyakina, R.; Suslov, V.; Bogomolov, A.; Klimova, N.; Shikhevich, S.; Savinkova, L.; Oshchepkov, D.; Kolchanov, N.A.; et al. Domestication explains two-thirds of differential gene expression variance between domestic and wild animals; the remaining one-third reflects intraspecific and interspecific variation. Animals 2021, 11, 2667. [Google Scholar] [CrossRef]
- Oshchepkov, D.; Chadaeva, I.; Kozhemyakina, R.; Zolotareva, K.; Khandaev, B.; Sharypova, E.; Ponomarenko, P.; Bogomolov, A.; Klimova, N.V.; Shikhevich, S.; et al. Stress reactivity, susceptibility to hypertension, and differential expression of genes in hypertensive compared to normotensive patients. Int. J. Mol. Sci. 2022, 23, 2835. [Google Scholar] [CrossRef]
- Shikhevich, S.; Chadaeva, I.; Khandaev, B.; Kozhemyakina, R.; Zolotareva, K.; Kazachek, A.; Oshchepkov, D.; Bogomolov, A.; Klimova, N.V.; Ivanisenko, V.A.; et al. Differentially expressed genes and molecular susceptibility to human age-related diseases. Int. J. Mol. Sci. 2023, 24, 3996. [Google Scholar] [CrossRef]
- Oshchepkov, D.; Chadaeva, I.; Kozhemyakina, R.; Shikhevich, S.; Sharypova, E.; Savinkova, L.; Klimova, N.V.; Tsukanov, A.; Levitsky, V.G.; Markel, A.L. Transcription factors as important regulators of changes in behavior through domestication of gray rats: Quantitative data from RNA sequencing. Int. J. Mol. Sci. 2022, 23, 12269. [Google Scholar] [CrossRef]
- Custodio, R.J.P.; Botanas, C.J.; de la Peña, J.B.; Dela Peña, I.J.; Kim, M.; Sayson, L.V.; Abiero, A.; Ryoo, Z.Y.; Kim, B.N.; Kim, H.J.; et al. Overexpression of the thyroid hormone-responsive (Thrsp) gene in the striatum leads to the development of inattentive-like phenotype in mice. Neuroscience 2018, 390, 141–150. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Schadt, E.E. variancePartition: Interpreting drivers of variation in complex gene expression studies. BMC Bioinform. 2016, 17, 483. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Jyothi, A.K.; Thotakura, B.; Priyadarshini, S.C.; Patil, S.; Poojari, M.S.; Subramanian, M. Paternal stress alters synaptic density and expression of Gap-43, Grin1, M1 and Syp genes in the hippocampus and cortex of offspring of stress-induced male rats. Morphologie 2023, 107, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Wonnacott, S.; Bailey, S.J.; Bailey, C.P. Sex differences in brain region-specific activation of c-Fos following kappa opioid receptor stimulation or acute stress in mice. Int. J. Mol. Sci. 2023, 24, 15098. [Google Scholar] [CrossRef] [PubMed]
- Sato, D.X.; Rafati, N.; Ring, H.; Younis, S.; Feng, C.; Blanco-Aguiar, J.A.; Rubin, C.J.; Villafuerte, R.; Hallböök, F.; Carneiro, M.; et al. Brain transcriptomics of wild and domestic rabbits suggests that changes in dopamine signaling and ciliary function contributed to evolution of tameness. Genome Biol. Evol. 2020, 12, 1918–1928. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, F.; Lo, L.-C.; Johnson, J.E.; Auerbach, A.; Anderson, D.J.; Joyner, A.L. Mammalian achaete-scute homolog 1 is required for the early development of olfactory and autonomic neurons. Cell 1993, 75, 463–476. [Google Scholar] [CrossRef]
- Tomita, K.; Moriyoshi, K.; Nakanishi, S.; Guillemot, F.; Kageyama, R. Mammalian achaete-scute and atonal homologs regulate neuronal versus glial fate determination in the central nervous system. EMBO J. 2000, 19, 5460–5472. [Google Scholar] [CrossRef]
- Sanes, D.H. Development of the Nervous System, 3rd ed.; Sanes, D., Reh, T., Harris, W., Eds.; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar] [CrossRef]
- Benítez-Burraco, A.; Uriagereka, J.; Nataf, S. The genomic landscape of mammal domestication might be orchestrated by selected transcription factors regulating brain and craniofacial development. Dev. Genes Evol. 2023, 233, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Yang, J.; Wang, G.; Wang, C.; Zhang, H. Bioinformatic analysis of gene expression profiles of pituitary gonadotroph adenomas. Oncol. Lett. 2018, 15, 1655–1663. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, J.; Liang, S.; Sun, M.; Duan, J. Low-dose exposure of silica nanoparticles induces neurotoxicity via neuroactive ligand-receptor interaction signaling pathway in zebrafish embryos. Int. J. Nanomed. 2020, 15, 4407–4415. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, P.; Conte, C.; Albi, E. The Multiple roles of sphingomyelin in Parkinson’s disease. Biomolecules 2021, 11, 1311. [Google Scholar] [CrossRef] [PubMed]
- Prasolova, L.A.; Gerbek, Y.E.; Gulevich, R.G.; Shikhevich, S.G.; Konoshenko, M.Y.; Kozhemyakina, R.V.; Oskina, I.N.; Plyusnina, I.Z. The effects of prolonged selection for behavior on the stress response and activity of the reproductive system of male grey mice (Rattus norvegicus). Russ. J. Genet. 2014, 50, 959–966. [Google Scholar] [CrossRef]
- Dityatev, A.; Rusakov, D.A. Molecular signals of plasticity at the tetrapartite synapse. Curr. Opin. Neurobiol. 2011, 21, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Tallima, H.; El Ridi, R. Arachidonic acid: Physiological roles and potential health benefits—A review. J. Adv. Res. 2017, 11, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Raleigh, M.J.; McGuire, M.T.; Brammer, G.L.; Pollack, D.B.; Yuwiler, A. Serotonergic mechanisms promote dominance acquisition in adult male vervet monkeys. Brain Res. 1991, 559, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Olivier, B. Serotonin and Aggression. Ann. N.Y. Acad. Sci. 2010, 1036, 382–392. [Google Scholar] [CrossRef]
- de Boer, S.F.; Koolhaas, J.M. 5-HT1A and 5-HT1B receptor agonists and aggression: A pharmacological challenge of the serotonin deficiency hypothesis. Europ. J. Pharmac. 2005, 526, 125–139. [Google Scholar] [CrossRef]
- Audero, E.; Mlinar, B.; Baccini, G.; Skachokova, Z.K.; Corradetti, R.; Gross, C. Suppression of serotonin neuron firing increases aggression in mice. J. Neurosci. 2013, 33, 8678–8688. [Google Scholar] [CrossRef]
- Zamponi, G. Targeting voltage-gated calcium channels in neurological and psychiatric diseases. Nat. Rev. Drug. Discov. 2016, 15, 19–34. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic. Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic. Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Term | Gene Symbols | NGENE | % | padj |
|---|---|---|---|---|---|
| KEGG PATHWAY | Neuroactive ligand-receptor interaction | Bdkrb2,Cckbr,Htr2c,Nmb,Pdyn,Prlr,P2rx4,Rln3,Sstr2,Tac3,Ucn,Vip | 12 | 10.9 | 0.005 |
| UP KW PTM | Cleavage on pair of basic residues | Alb, Cartpt,Enpp2,Pdyn,Rln3,Tac3,Ucn,Vip | 8 | 7.3 | 0.01 |
| GO TERM BP DIRECT | Arachidonic acid secretion | Bdkrb2,Nmb,Pla2g2c,Pla2g2d,Pla2g5 | 5 | 4.5 | 0.01 |
| UP KW BIOLOGICAL PROCESS | Lipid metabolism | Aox1,Bdh1,Fuca1,Enpp2,Insig1,Liph,Mogat2,Pla2g2c,Pla2g2d,Pla2g5, Retsat, Thrsp | 12 | 10.9 | 0.01 |
| GO TERM CC DIRECT | Extracellular region | Alb,Defb17,Enpp2,Hspa1a,Hspa1b,Liph,Nmb,Pla2g2c,Pla2g2d,Pla2g5,Pdyn,Rln3,Tac3,Tecta,Ucn,Vip | 16 | 14.5 | 0.025 |
| UP KW DOMAIN | Signal | Alb,Cartpt,Cd22,Cspg4b, Defb17,Enpp2,Fat2,Fcgr3a,Fcgr2b,Fcrl2,Fjx1, Frem1,Fuca1,Htr2c,Htr3a,Lilrb3l,Liph,Lypd1, Mpeg1,Nmb,Pcdhga1, Pdia4, Pdyn,Pla2g2c,Pla2g2d,Pla2g5,Plod1, Prlr,Retsat,Rln3,Smoc2,Spint1,Sulf1,Tac3,Tecta, Ucn,Vip, Vstm2b | 38 | 34.5 | 0.025 |
| UP KW PTM | Disulfide bond | Alb,Bdkrb2,Cartpt,Cckbr,Defb17,Enpp2,Fat2,Fcgr3a,Fcgr2b,Fosb,Htr2c,Htr3a,Itm2a,Lilrb3l,Liph,Lypd1,Pla2g2c,Pla2g2d,Pla2g5,Pdia4,Pdyn,Prlr,P2rx4,Rln3,Sstr2,Tecta, Txnrd2 | 27 | 24.5 | 0.05 |
| Group | Tame vs. Aggressive Rats Brain Region | |||
|---|---|---|---|---|
| Hypothalamus | Hippocampus | PAG | MTg | |
| Total number of sequence reads (NCBI SRA ID: PRJNA668014) | 219,086,104 | 169,529,658 | 210,128,758 | 182,197,974 |
| Reads mapped to reference rat genome RGSC Rnor_6.0, UCSC Rn6, July 2014 (%) | 184,991,379 (84.4%) | 146,521,467 (88.7%) | 177,608,837 (84.5%) | 158,310,590 (86.9%) |
| Expressed genes identified (at a threshold of at least 10 reads per gene in all 4 brain regions) | 14039 | |||
| Statistically significant DEGs (pADJ < 0.05, Fisher’s Z-test with Benjamini correction) | 44 | 42 | 39 | 42 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chadaeva, I.; Kozhemyakina, R.; Shikhevich, S.; Bogomolov, A.; Kondratyuk, E.; Oshchepkov, D.; Orlov, Y.L.; Markel, A.L. A Principal Components Analysis and Functional Annotation of Differentially Expressed Genes in Brain Regions of Gray Rats Selected for Tame or Aggressive Behavior. Int. J. Mol. Sci. 2024, 25, 4613. https://doi.org/10.3390/ijms25094613
Chadaeva I, Kozhemyakina R, Shikhevich S, Bogomolov A, Kondratyuk E, Oshchepkov D, Orlov YL, Markel AL. A Principal Components Analysis and Functional Annotation of Differentially Expressed Genes in Brain Regions of Gray Rats Selected for Tame or Aggressive Behavior. International Journal of Molecular Sciences. 2024; 25(9):4613. https://doi.org/10.3390/ijms25094613
Chicago/Turabian StyleChadaeva, Irina, Rimma Kozhemyakina, Svetlana Shikhevich, Anton Bogomolov, Ekaterina Kondratyuk, Dmitry Oshchepkov, Yuriy L. Orlov, and Arcady L. Markel. 2024. "A Principal Components Analysis and Functional Annotation of Differentially Expressed Genes in Brain Regions of Gray Rats Selected for Tame or Aggressive Behavior" International Journal of Molecular Sciences 25, no. 9: 4613. https://doi.org/10.3390/ijms25094613
APA StyleChadaeva, I., Kozhemyakina, R., Shikhevich, S., Bogomolov, A., Kondratyuk, E., Oshchepkov, D., Orlov, Y. L., & Markel, A. L. (2024). A Principal Components Analysis and Functional Annotation of Differentially Expressed Genes in Brain Regions of Gray Rats Selected for Tame or Aggressive Behavior. International Journal of Molecular Sciences, 25(9), 4613. https://doi.org/10.3390/ijms25094613