
Plant Conservation Practitioners Can Benefit from Neutral Genetic Diversity




Abstract
:1. Introduction
2. Bridging the Gap between Conservation Geneticists and Conservation Practitioners: Some Examples
3. Expected Scenarios According to Levels of Neutral Genetic Parameters: A Simplified Proposal
4. A Prioritization Framework for Conservation Strategies Based on Neutral Genetic Diversity Scenarios
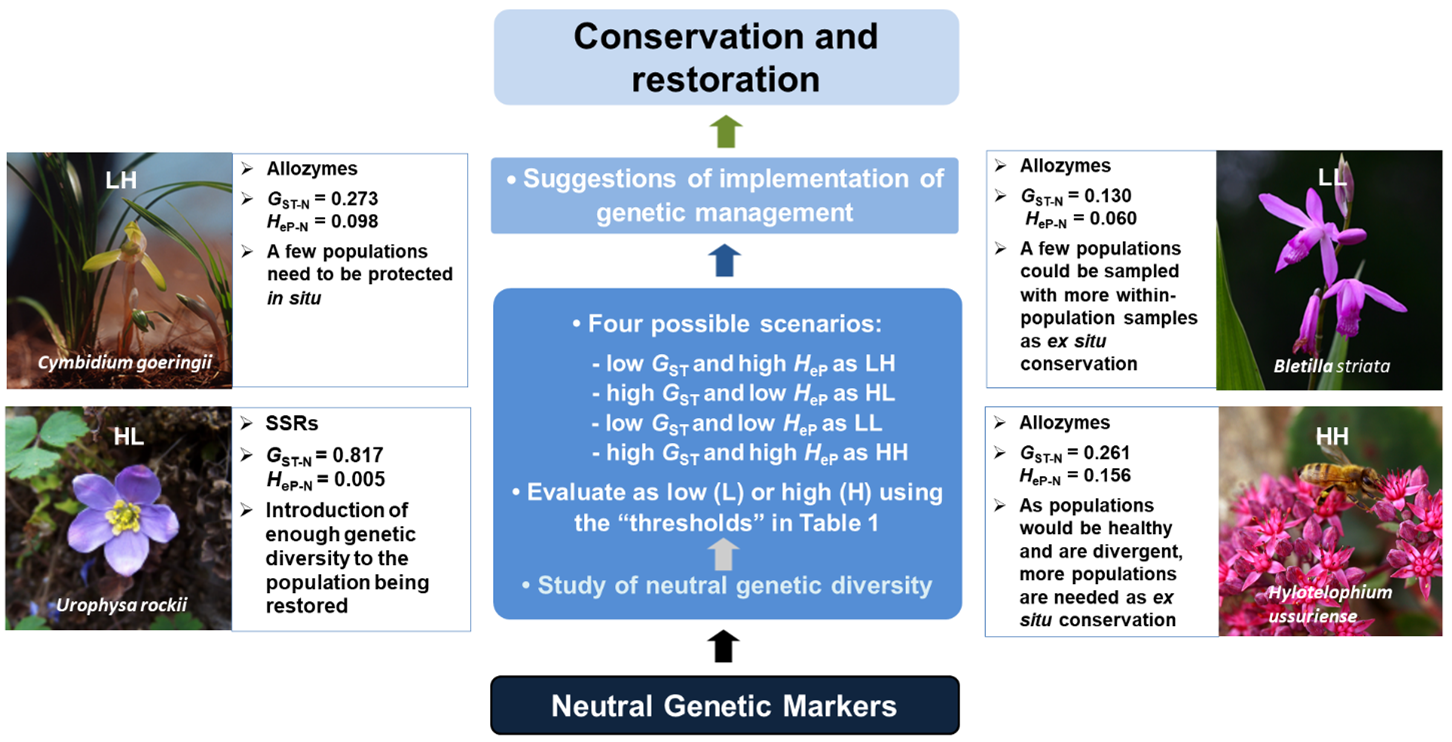
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harrison, K.A.; Pavlova, A.; Telonis-Scott, M.; Sunnucks, P. Using Genomics to Characterize Evolutionary Potential for Conservation of Wild Populations. Evol. Appl. 2014, 7, 1008–1025. [Google Scholar] [CrossRef] [PubMed]
- DeWoody, J.A.; Harder, A.M.; Mathur, S.; Willoughby, J.R. The Long-Standing Significance of Genetic Diversity in Conservation. Mol. Ecol. 2021, 30, 4147–4154. [Google Scholar] [CrossRef]
- Campbell, R.K. Soils, Seed-Zone Maps, and Physiography: Guidelines for Seed Transfer of Douglas-Fir in South-Western Oregon. For. Sci. 1991, 37, 973–986. [Google Scholar]
- Rehfeldt, G.E. Genetic Variation, Climate Models and the Ecological Genetics of Larix occidentalis. For. Ecol. Manag. 1995, 78, 21–37. [Google Scholar] [CrossRef]
- Mazer, S.M.; LeBuhn, G. Genetic Variation in Life-History Traits: Heritability Estimates within and Genetic Differentiation among Populations. In Life History Evolution in Plants; Vuorisalo, T.O., Mutikainen, P.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 86–171. [Google Scholar]
- Willi, Y.; Van Buskirk, J.; Hoffmann, A.A. Limits to the Adaptive Potential of Small Populations. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 433–458. [Google Scholar] [CrossRef]
- Shirk, R.Y.; Hamrick, J.L. Multivariate Adaptation but no Increase in Competitive Ability in Invasive Geranium carolinianum L. (Geraniaceae). Evolution 2014, 68, 2945–2959. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Tang, F.; Wei, N.; Yao, X. Molecular and Quantitative Trait Variation within and among Small Fragmented Populations of the Endangered Plant Species Psilopeganum sinense. Ann. Bot. 2014, 113, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, S.P.; Forester, B.R.; Latch, E.K.; Aitken, S.N.; Hoban, S. Guidelines for Planning Genomic Assessment and Monitoring of Locally Adaptive Variation to Inform Species Conservation. Evol. Appl. 2018, 11, 1035–1052. [Google Scholar] [CrossRef] [PubMed]
- Vimmerstedt, J.P. Southern Appalachian White Pine Plantations Site, Volume, and Yield; USDA Forest Service, Southeastern Forest Experimental Station: Asheville, NC, USA, 1962; pp. 1–15.
- Anekonda, T.S.; Lomas, M.C.; Adams, W.T.; Kavanagh, K.L.; Aitken, S.N. Genetic Variation in Drought Hardiness of Coastal Douglas-Fir Seedlings from British Columbia. Can. J. For. Res. 2002, 32, 1701–1716. [Google Scholar] [CrossRef] [Green Version]
- Arend, M.; Kuster, T.; Günthardt-Goerg, M.S.; Dobbertin, M. Provenance-Specific Growth Responses to Drought and Air-Warming in Three European Oak Species (Quercus robur, Q. petraea and Q. pubescens). Tree Physiol. 2011, 31, 287–297. [Google Scholar] [CrossRef] [Green Version]
- Gary, L.A.; Grat, T.; Mbogga, M.S.; Chen, P.-Y.; Hamann, A. Assisted Migration to Address Climate Change: Recommendations for Aspen Reforestation in Western Canada. Ecol. Appl. 2011, 21, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefévre, F.; Lenormand, T.; Yeaman, S.; Whetten, R. Potential for Evolutionary Responses to Climate Change-Evidence from Tree Populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef] [Green Version]
- George, J.; Grabner, M.; Karanitsch-Ackerl, S.; Mayer, K.; Weißenbacher, L.; Schueler, S. Genetic Variation, Phenotypic Stability, and Repeatability of Drought Response in European Larch Throughout 50 years in a Common Garden Experiment. Tree Physiol. 2017, 37, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sork, V.L. Genomic Studies of Local Adaptation in Natural Plant Populations. J. Hered. 2018, 109, 3–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.T.; Song, B.-H. Plant Adaptation to Climate Change—Where Are We? J. Syst. Evol. 2020, 58, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Capblancq, T.; Fitzpatrick, M.C.; Bay, R.A.; Exposito-Alonso, M.; Keller, S.R. Genomic Prediction of (Mal)Adaptation Across Current and Future Climatic Landscapes. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 245–269. [Google Scholar] [CrossRef]
- Mahony, C.R.; MacLachlan, I.R.; Lind, B.M.; Yoder, J.B.; Wang, T.; Aitken, S.N. Evaluating Genomic Data for Management of Local Adaptation in a Changing Climate: A lodgepole Pine Case Study. Evol. Appl. 2019, 13, 116–131. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Fournier, P.; Lewthwaite, J.M.M.; Mooers, A.Ø. Do We Need to Identify Adaptive Genetic Variation When Prioritizing Populations for Conservation? Conserv. Genet. 2021, 22, 205–216. [Google Scholar] [CrossRef]
- Funk, W.C.; Forester, B.R.; Converse, S.J.; Darst, C.; Morey, S. Improving Conservation Policy with Genomics: A Guide to Integrating Adaptive Potential into U.S. Endangered Species Act Decisions for Conservation Practitioners and Geneticists. Conserv. Genet. 2018, 20, 115–134. [Google Scholar] [CrossRef]
- García-Dorado, A.; Caballero, A. Neutral Genetic Diversity as a Useful Tool for Conservation Biology. Conserv. Genet. 2021, 22, 541–545. [Google Scholar] [CrossRef]
- Kardos, M.; Armstrong, E.; Fitzpatrick, S.; Hauser, S.; Hedrick, P.; Miller, J.; Tallmon, D.A.; Funk, W.C. The Crucial Role of Genome-Wide Genetic Variation in Conservation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Teixeira, J.C.; Huber, C.D. The Inflated Significance of Neutral Genetic Diversity in Conservation Genetics. Proc. Natl. Acad. Sci. USA 2021, 118, e2015096118. [Google Scholar] [CrossRef]
- Ottewell, K.M.; Bickerton, D.C.; Byrne, M.; Lowe, A.J. Bridging the Gap: A genetic Assessment Framework for Population-Level Threatened Plant Conservation Prioritization and Decision-Making. Divers. Distrib. 2016, 22, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Godefroid, S.; Piazza, C.; Rossi, G.; Buord, S.; Stevens, A.-D.; Aguraiuja, R.; Cowell, C.; Weekley, C.W.; Voggk, G.; Iriondo, J.M.; et al. How Successful are Plant Species Reintroductions? Biol. Conserv. 2011, 144, 672–682. [Google Scholar] [CrossRef]
- Hoban, S.M.; Hauffe, H.C.; Pérez-Espona, S.; Arntzen, J.W.; Bertorelle, G.; Bryja, J.; Frith, K.; Gaggiotti, O.; Galbusera, P.; Godoy, J.; et al. Bringing Genetic Diversity to the Forefront of Conservation Policy and Management. Conserv. Genet. Resour. 2013, 5, 593–598. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.R.; Dussex, N.; van Heezik, Y. Bridging the Conservation Genetics Gap by Identifying Barriers to Implementation for Conservation Practitioners. Glob. Ecol. Conserv. 2017, 10, 231–242. [Google Scholar] [CrossRef]
- Britt, M.; Haworth, S.E.; Johnson, J.B.; Martchenko, D.; Shafer, A.B. The Importance of Non-Academic Coauthors in Bridging the Conservation Genetics Gap. Biol. Conserv. 2018, 218, 118–123. [Google Scholar] [CrossRef]
- Hulme, P.E. Bridging the Knowing-Doing Gap: Know-Who, Know-What, Know-Why, Know-How and Know-When. J. Appl. Ecol. 2014, 51, 1131–1136. [Google Scholar] [CrossRef]
- Rivers, M.C.; Brummitt, N.A.; Lughadha, E.N.; Meagher, T.R. Do Species Conservation Assessments Capture Genetic Diversity? Glob. Ecol. Conserv. 2014, 2, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Haig, S.M.; Miller, M.P.; Bellinger, R.; Draheim, H.M.; Mercer, D.M.; Mullins, T.D. The Conservation Genetics Juggling Act: Integrating Genetics and Ecology, Science and Policy. Evol. Appl. 2016, 9, 181–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian, Y.; Bollmann, K.; Brang, P.; Hein, C.; Olschewski, R.; Rigling, A.; Stofer, S.; Holderegger, R. How to Close the Science-Practice Gap in Nature Conservation? Information Sources Used by Practitioners. Biol. Conserv. 2019, 235, 93–101. [Google Scholar] [CrossRef]
- Holderegger, R.; Balkenhol, N.; Bolliger, J.; Engler, J.O.; Gugerli, F.; Hochkirch, A.; Nowak, C.; Segelbacher, G.; Wider, A.; Zachos, F.E. Conservation Genetics: Linking Science with Practice. Mol. Ecol. 2019, 28, 3848–3856. [Google Scholar] [CrossRef] [Green Version]
- Hoban, S.; Bruford, M.; D’Urban Jackson, J.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjögren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic Diversity Targets and Indicators in the CBD post-2020 Global Biodiversity Framework must be Improved. Biol. Conserv. 2020, 248, 108654. [Google Scholar] [CrossRef]
- Knight, A.T.; Cowling, R.M.; Rouget, M.; Balmford, A.; Lombard, A.T.; Campbell, B.M. Knowing but Not Doing: Selecting priority Conservation Areas and the Research–Implementation Gap. Conserv. Biol. 2008, 22, 610–617. [Google Scholar] [CrossRef]
- Dubois, N.S.; Gomez, A.; Carlson, S.; Russell, D. Bridging the Research-Implementation Gap Requires Engagement from Practitioners. Conserv. Sci. Pract. 2019, 2, e134. [Google Scholar] [CrossRef] [Green Version]
- Taft, H.R.; McCoskey, D.N.; Miller, J.M.; Pearson, S.K.; Coleman, M.A.; Fletcher, N.K.; Mittan, C.S.; Meek, M.H.; Barbosa, S. Research–Management Partnerships: An Opportunity to Integrate Genetics in Conservation Actions. Conserv. Sci. Pract. 2020, 2, e218. [Google Scholar] [CrossRef]
- Mable, B.K. Conservation of Adaptive Potential and Functional Diversity: Integrating Old and New Approaches. Conserv. Genet. 2019, 20, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Cook, C.N.; Beever, E.A.; Thurman, L.L.; Thompson, L.M.; Gross, J.E.; Whiteley, A.R.; Nicotra, A.B.; Szymanski, J.A.; Botero, C.A.; Hall, K.R.; et al. Supporting the Adaptive Capacity of Species through more Effective Knowledge Exchange with Conservation Practitioners. Evol. Appl. 2021, 4, 1969–1979. [Google Scholar] [CrossRef]
- Nishihiro, J.; Uesugi, S.R.; Takagawa, I.; Washitani, I. Toward the Restoration of a Sustainable Population of a Threatened Aquatic Plant, Nymphoides peltata: Integrated Genetic/Demographic Studies and Practices. Biol. Conserv. 2009, 142, 1906–1912. [Google Scholar] [CrossRef]
- Ibáñez, O.; Calero, C.; Mayol, M.; Rosselló, J.A. Isozyme uniformity in a wild extinct insular plant, Lysimachia minoricensis J.J. Rodr. (Primulaceae). Mol. Ecol. 1999, 8, 813–817. [Google Scholar] [CrossRef]
- Laikre, L.; Hoban, S.; Bruford, M.W.; Segelbacher, G.; Allendorf, F.W.; Gajardo, G.; González Rodríguez, A.; Hedrick, P.W.; Heuertz, M.; Hohenlohe, P.A.; et al. Post-2020 Goals Overlook Genetic Diversity. Science 2020, 367, 1083–1085. [Google Scholar] [CrossRef] [Green Version]
- Hamrick, J.L.; Godt, M.J.W. Allozyme Diversity in Plant Species. In Plant Population Genetics, Breeding and Germplasm Resources; Brown, A.H.D., Clegg, M.T., Kahler, A.L., Weir, B.S., Eds.; Sinauer: Sunderland, MA, USA, 1989; pp. 43–63. [Google Scholar]
- Hamrick, J.L.; Godt, M.J.W. Effects of Life History Traits on Genetic Diversity in Plant Species. Philos. Trans. R. Soc. B 1996, 351, 1291–1298. [Google Scholar]
- Hamrick, J.L.; Godt, M.J.W. Sherman-Broyles, S.L. Factors Influencing Levels of Genetic Diversity in Woody Plant Species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- Petit, R.; Hampe, A. Some Evolutionary Consequences of Being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.Y.; Son, S.; Herrando-Moraira, S.; Tang, C.Q.; Maki, M.; Kim, Y.D.; López-Pujol, J.; Hamrick, J.L.; Chung, M.G. Incorporating Differences between Genetic Diversity of Trees and Herbaceous Plants in Conservation Strategies. Conserv. Biol. 2020, 34, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Nybom, H. Comparison of Different Nuclear DNA Markers for Estimating Intraspecific Genetic Diversity in Plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef]
- Guries, R.P.; Ledig, F.T. Genetic Diversity and Population Structure in pitch pine (Pinus rigida Mill.). Evolution 1982, 36, 387–402. [Google Scholar] [CrossRef]
- Houston, D.B.; Houston, D.R. Allozyme Genetic Diversity among Fagus grandifolia Trees Resistant or Susceptible to Beech Bark Disease in Natural Populations. Can. J. For. Res. 2000, 30, 778–789. [Google Scholar] [CrossRef]
- Godt, M.J.W.; Hamrick, J.L. Genetic Diversity and Morphological Differentiation in Liatris helleri (Asteraceae), a Threatened Plant Species. Biodiver. Conserv. 1996, 5, 461–471. [Google Scholar] [CrossRef]
- Broyles, S.B.; Wyatt, R. Allozyme Diversity and Genetic Structure in Southern Appalachian Populations of Asclepias exaltata. Syst. Bot. 1993, 18, 18–30. [Google Scholar] [CrossRef]
- Chung, M.Y.; Chung, M.G. Allozyme Diversity and Population Structure in Korean Populations of Cymbidium goeringii (Orchidaceae). J. Plant Res. 1999, 112, 139–144. [Google Scholar] [CrossRef]
- Bucharova, A.; Bossdorf, O.; Hölzel, N.; Kollman, J.; Prasse, R.; Durka, W. Mix and Match: Regional Admixture Provenancing Strikes a Balance among Different Seed-Searching Strategies for Ecological Restoration. Conserv. Genet. 2019, 20, 7–17. [Google Scholar] [CrossRef]
- Godt, M.J.W.; Hamrick, J.L.; Bratton, S. Genetic Diversity in a Threatened Wetland Species, Helonias bullata (Liliaceae). Conserv. Biol. 1995, 9, 596–604. [Google Scholar] [CrossRef]
- Xie, D.-F.; Li, M.-J.; Tan, J.-B.; Price, M.; Xiao, Q.-Y.; Zhou, S.-D.; Yu, Y.; He, X.J. Phylogeography and Genetic Effects of Habitat Fragmentation on Endemic Urophysa (Ranunculaceae) in Yungui Plateau and Adjacent Regions. PLoS ONE 2017, 12, e0186378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Pujol, J.; Zhang, F.-M.; Ge, S. Population Genetics and Conservation of the Critically Endangered Clematis acerifolia (Ranunculaceae). Can. J. Bot. 2005, 83, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.Y.; López-Pujol, J.; Chung, M.G. Population History of the Two Carnivorous Plants Drosera peltata var. nipponica and Drosera rotundifolia (Droseraceae) in Korea. Am. J. Bot. 2013, 11, 2231–2239. [Google Scholar] [CrossRef]
- Boys, J.; Cherry, M.; Dayanandan, S. Microsatellite Analysis Reveals Genetically Disjunct Populations of Red Pine (Pinus resinosa, Pinaceae). Am. J. Bot. 2005, 92, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Wang, Y.; Korpelainen, H.; Li, C. Allozyme Variation in Natural Populations of Picea Asperata. Silva Fenn. 2005, 39, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.Y.; López-Pujol, J.; Chung, M.G. Low Genetic Diversity in Marginal Populations of Bletilla striata (Orchidaceae) in Southern Korea: Insights into Population History and Implications for Conservation. Biochem. Syst. Ecol. 2013, 46, 88–96. [Google Scholar] [CrossRef]
- Chung, M.Y.; López-Pujol, J.; Chung, M.G. Comparative Genetic Structure between Sedium ussuriense and S. kamtschaticum (Crassulaceae). Am. J. Bot. 2014, 101, 946–956. [Google Scholar] [CrossRef]
- Xie, D.-F.; Cheng, R.-Y.; Fu, X.; Zhang, X.-Y.; Price, M.; Lan, Y.-L.; Wang, C.-B.; He, X.-J. A Combined Morphological and Molecular Evolutionary Analysis of Karst-Environment Adaptation for the Genus Urophysa (Ranunculaceae). Front. Plant Sci. 2021, 12, 667988. [Google Scholar] [CrossRef]
- Li, X.; Ding, X.; Chu, B.; Zhou, Q.; Ding, G.; Gu, S. Genetic Diversity Analysis and Conservation of the Endangered Chinese Endemic Herb Dendrobium officinale Kimura et Migo (Orchidaceae) Based on AFLP. Genetica 2008, 133, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.G.; Kang, S.S. Allozyme Genetic and Clonal Diversity within Populations of Chimaphila japonica and Pyrola japonica (Pyrolaceae). Isr. J. Plant Sci. 1996, 44, 259–271. [Google Scholar] [CrossRef]
- Chung, M.G. Allozyme Diversity in the Endangered Herb Lycoris sanguinea Var. koreana (Amaryllidaceae). Ann. Bot. Fenn. 1999, 36, 1–7. [Google Scholar]
- Gibson, J.P.; Hamrick, J.L. Genetic Diversity and Structure in Pinus pungens (Table Mountain Pine) Populations. Can. J. For. Res. 1991, 21, 635–642. [Google Scholar] [CrossRef]
- He, J.S.; Li, Z.Z.; Huang, H.W. Allozymic Genetic Diversity in Manglietia patungensis, an Endangered Species, and Its Conservation Strategies. Biodivers. Sci. 2005, 13, 27–35, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Shore, J.S. Tetrasomic Inheritance and Isozyme Variation in Turnera ulmifolia vars. elegans Urb. and intermedia Urb. (Turneraceae). Heredity 1991, 66, 305–312. [Google Scholar] [CrossRef]
- Griffin, S.R.; Barrett, S.C.H. Post-Glacial History of Trillium grandiflorum (Melanthiaceae) in Eastern North America: Inferences from Phylogeography. Am. J. Bot. 2004, 91, 465–473. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Parameter | Annuals | Herbaceous Perennials | Woody Perennials |
|---|---|---|---|
| Allozyme | |||
| NP, NS | 226, 226 | 228, 215 | 213/214 |
| %PP-N | 29.4 b | 27.5 b | 47.9 a |
| %PS-N | 49.2 b | 42.8 b | 62.7 a |
| AP-N | 1.45 b | 1.38 b | 1.74 a |
| AS-N | 2.02 ab | 1.75 b | 2.16 a |
| HeP-N | 0.101 b | 0.096 b | 0.144 a |
| HeS-N | 0.154 b | 0.125 c | 0.170 a |
| GST-N | 0.355 a | 0.256 b | 0.089 c |
| Parameter | Annuals | Short-lived perennials | Long-lived perennials b |
| RAPD c | |||
| N | 6 | 17 | 37 |
| Hpop | 0.13 b | 0.20 ab | 0.25 a |
| N | 10 | 45 | 60 |
| ΦST-N | 0.62 a | 0.41 b | 0.25 c |
| N | 2 | 18 | 24 |
| GST-N | 0.47 a | 0.32 ab | 0.19 b |
| SSR | |||
| N | 15 | 29 | 59 |
| HeP-N | 0.46 b | 0.55 b | 0.68 a |
| N | 4 | 12 | 17 |
| GST-N | 0.40 a | 0.31 ab | 0.19 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, M.Y.; Son, S.; López-Pujol, J.; Mao, K.; Chung, M.G. Plant Conservation Practitioners Can Benefit from Neutral Genetic Diversity. Diversity 2021, 13, 552. https://doi.org/10.3390/d13110552
Chung MY, Son S, López-Pujol J, Mao K, Chung MG. Plant Conservation Practitioners Can Benefit from Neutral Genetic Diversity. Diversity. 2021; 13(11):552. https://doi.org/10.3390/d13110552
Chicago/Turabian StyleChung, Mi Yoon, Sungwon Son, Jordi López-Pujol, Kangshan Mao, and Myong Gi Chung. 2021. "Plant Conservation Practitioners Can Benefit from Neutral Genetic Diversity" Diversity 13, no. 11: 552. https://doi.org/10.3390/d13110552
APA StyleChung, M. Y., Son, S., López-Pujol, J., Mao, K., & Chung, M. G. (2021). Plant Conservation Practitioners Can Benefit from Neutral Genetic Diversity. Diversity, 13(11), 552. https://doi.org/10.3390/d13110552