
Freshwater Reservoir, Ecological Traps and Source-Sink Dynamics


Abstract
:1. Introduction
2. Materials and Methods
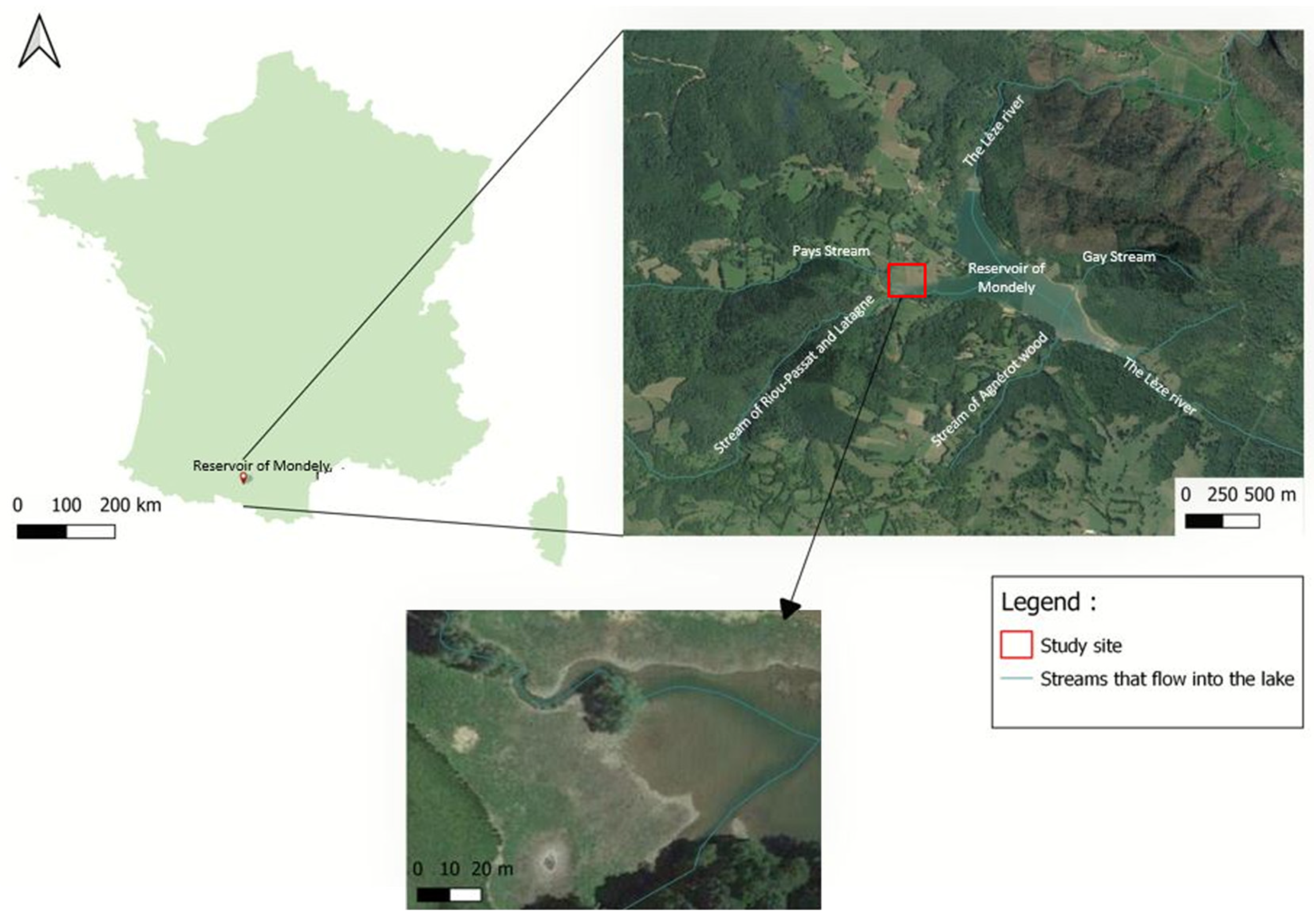
2.1. Study Site
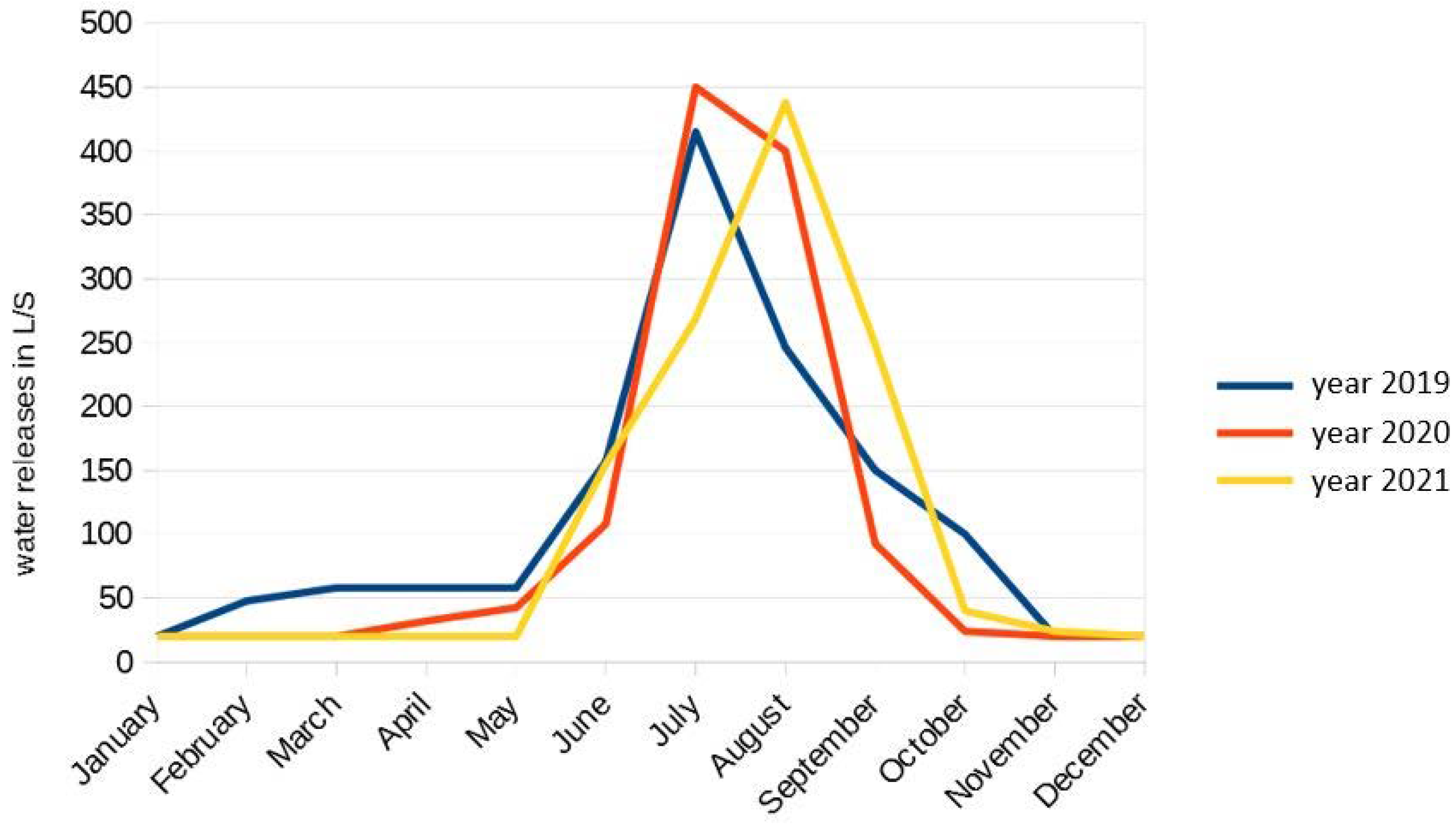
2.2. Sampling Methods
2.3. Odonata Life History Traits
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, I.J.; Abell, R.; Darwall, W.; Thieme, M.L.; Tickner, D.; Timboe, I. The freshwater biodiversity crisis. Science 2018, 362, 1369. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oertli, B. The use of dragonflies in the assessment and monitoring of aquatic habitats. In Dragonflies & Damselflies; Model Organisms for Ecological and Evolutionary Research; Cordoba-Aguilar, A., Ed.; Oxford University Press: Oxford, UK, 2008; pp. 79–95. [Google Scholar]
- Pollard, E. A Method for Assessing Changes in the Abundance of Butterflies. Biol. Cons. 1977, 12, 115–134. [Google Scholar] [CrossRef]
- Waller, J.T.; Willink, B.; Tschol, M.; Svensson, E.I. The odonate phenotypic database, a new open data resource for comparative studies of an old insect order. Sci. Data 2019, 6, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harabiš, F.; Hronková, J. European database of the life-history, morphological and habitat characteristics of dragonflies (Odonata). Eur. J. Ent. 2020, 117, 302–308. [Google Scholar] [CrossRef]
- Charlot, B.; Danflous, S.; Louboutin, B.; Jaulin, S. Liste Rouge des Odonates d’Occitanie; CEN Midi-Pyrénées & OPIE: Toulouse, France, 2018; p. 102. [Google Scholar]
- Corbet, P.S. Dragonflies: Behavior and Ecology of Odonata; Cornell University Press: Ithaca, NY, USA, 2004; p. 829. [Google Scholar]
- Battin, J. When good animals love bad habitats: Ecological traps and the conservation of animal populations. Cons. Biol. 2004, 18, 1482–1491. [Google Scholar] [CrossRef]
- Hale, R.; Swearer, S.E. Ecological traps: Current evidence and future directions. Proc. R. Soc. B 2016, 283, 20152647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, B.A.; Hutto, R.L. A framework for understanding ecological traps and an evaluation of existing evidence. Ecology 2006, 87, 1075–1085. [Google Scholar] [CrossRef]
- Robertson, B.A.; Hutto, R.L. Is selectively harvested forest an ecological trap for Olive-sided Flycatchers? Condor 2007, 109, 109–121. [Google Scholar] [CrossRef]
- Hale, R.; Treml, E.A.; Swearer, S.E. Evaluating the metapopulation consequences of ecological traps. Proc. R. Soc. B 2015, 282, 20142930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, P.C. Sources and sinks in population biology. Trends. Ecol. Evol. 1996, 11, 326–330. [Google Scholar] [CrossRef]
- Stevens, V.M.; Turlure, C.; Baguette, M. A meta-analysis of dispersal in butterflies. Biol. Rev. 2010, 85, 625–642. [Google Scholar] [CrossRef] [PubMed]
- Stevens, V.M.; Trochet, A.; Van Dyck, H.; Clobert, J.; Baguette, M. How is dispersal integrated in life-histories: A quantitative analysis with butterflies. Ecol. Lett. 2012, 15, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Manel, S.; Schwartz, M.K.; Luikart, G.; Taberlet, C. Landscape genetics: Combining landscape ecology and population genetics. Trends Ecol. Evol. 2003, 18, 189–197. [Google Scholar] [CrossRef]
- Manel, S.; Holderegger, R. Ten Years of Landscape Genetics. Trends. Ecol. Evol. 2013, 28, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Hanski, I. Metapopulation Ecology; Oxford University Press: Oxford, UK, 1999; p. 324. [Google Scholar]
- Carmignani, J.R.; Roy, A.H. Ecological impacts of water drawdowns on lake littoral zones: A review. Aquat. Sci. 2017, 79, 803–827. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Abundance Class | Reproduction Index |
|---|---|---|
| Calopteryx virgo meridionalis | 2 | - |
| Lestes barbarus | 1 | E |
| Lestes viridis | 4 | T |
| Sympecma fusca | 4 | T |
| Ischnura elegans | 2 | M |
| Enallagma cyathigerum | 3 | M |
| Coenagrion puella | 3 | E |
| Erythromma lindenii | 2 | E |
| Pyrrhosoma nymphula | 1 | E |
| Ceriagrion tenellum | 1 | - |
| Platycnemis pennipes | 1 | - |
| Aeshna mixta | 2 | E |
| Aeshna affinis | 2 | T |
| Anax imperator | 1 | E |
| Boyeria irene | 1 | - |
| Cordulegaster boltonii | 1 | E |
| Libellula quadrimaculata | 2 | - |
| Libellula depressa | 1 | E |
| Orthetrum cancellatum | 1 | - |
| Sympetrum striolatum | 4 | T |
| Sympetrum fonscolombii | 3 | T |
| Sympetrum sanguineum | 3 | T |
| Sympetrum meridionale | 1 | M |
| Species | Egg-Laying Strategy | Larval Location | Egg Laying/Hatching Time | Larval Development |
|---|---|---|---|---|
| Lestes barbarus | Out of water, endophytic | On sediments | June–August/4–21 weeks | 4–8 weeks |
| Lestes viridis | Out of water, endophytic | On sediments | August–September/28–32 weeks | 8–12 weeks |
| Sympecma fusca | In water, endophytic | On sediments | April–May/3–6 weeks | 8–12 weeks |
| Ischnura elegans | In water, endophytic | On vegetation | June–September/1–3 weeks | 2–3 weeks/40–50 weeks |
| Enallagma cyathigerum | In water, endophytic | On vegetation | June–August/2–3 weeks | 40–50 weeks |
| Coenagrion puella | In water, endophytic | On sediments | June–July/3–5 weeks | 24–40 weeks |
| Erythromma lindenii | In water, endophytic | On vegetation | June–July/ | 24–48 weeks |
| Ceriagrion tenellum | In water, endophytic | On sediments | July–August/3–4 weeks | 36–72 weeks |
| Aeshna mixta | In water, endophytic | Mixed | August–September/winter diapause (28–36 weeks) | 16–20 weeks |
| Aeshna affinis | Out of water, in mud or endophytic | No data | August/ winter diapause (28–36 weeks) | 12 weeks |
| Anax imperator | In water, endophytic | On vegetation | June–September/3–6 weeks | 12–100 weeks |
| Libellula quadrimaculata | In water, exophytic | Mixed | June–July/2–7 weeks | 52–156 weeks |
| Libellula depressa | In water, exophytic | On sediments | June–July/1–5 weeks | 52–104 weeks |
| Orthetrum cancellatum | In water, exophytic | On sediments | July–August/5–6 weeks | 52–156 weeks |
| Sympetrum striolatum | Out or in water, exophytic | On sediments | August–September/4 weeks or after winter diapause | 20 weeks |
| Sympetrum fonscolombii | In water, exophytic | On sediments | June–September/1–3 weeks | Direct development or winter diapause |
| Sympetrum sanguineum | Out or in water, exophytic | Mixed | July–August/winter diapause | 6–10 weeks |
| Sympetrum meridionale | Out or in water, exophytic | Mixed | August–October/winter diapause | 8–16 weeks |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melendez, N.; Stevens, V.M.; Baguette, M. Freshwater Reservoir, Ecological Traps and Source-Sink Dynamics. Diversity 2022, 14, 1021. https://doi.org/10.3390/d14121021
Melendez N, Stevens VM, Baguette M. Freshwater Reservoir, Ecological Traps and Source-Sink Dynamics. Diversity. 2022; 14(12):1021. https://doi.org/10.3390/d14121021
Chicago/Turabian StyleMelendez, Nina, Virginie M. Stevens, and Michel Baguette. 2022. "Freshwater Reservoir, Ecological Traps and Source-Sink Dynamics" Diversity 14, no. 12: 1021. https://doi.org/10.3390/d14121021
APA StyleMelendez, N., Stevens, V. M., & Baguette, M. (2022). Freshwater Reservoir, Ecological Traps and Source-Sink Dynamics. Diversity, 14(12), 1021. https://doi.org/10.3390/d14121021