



Obligation to Enhance OBIS Data for Sea- and Shorebirds of the Americas



Abstract
1. Introduction
2. Methods
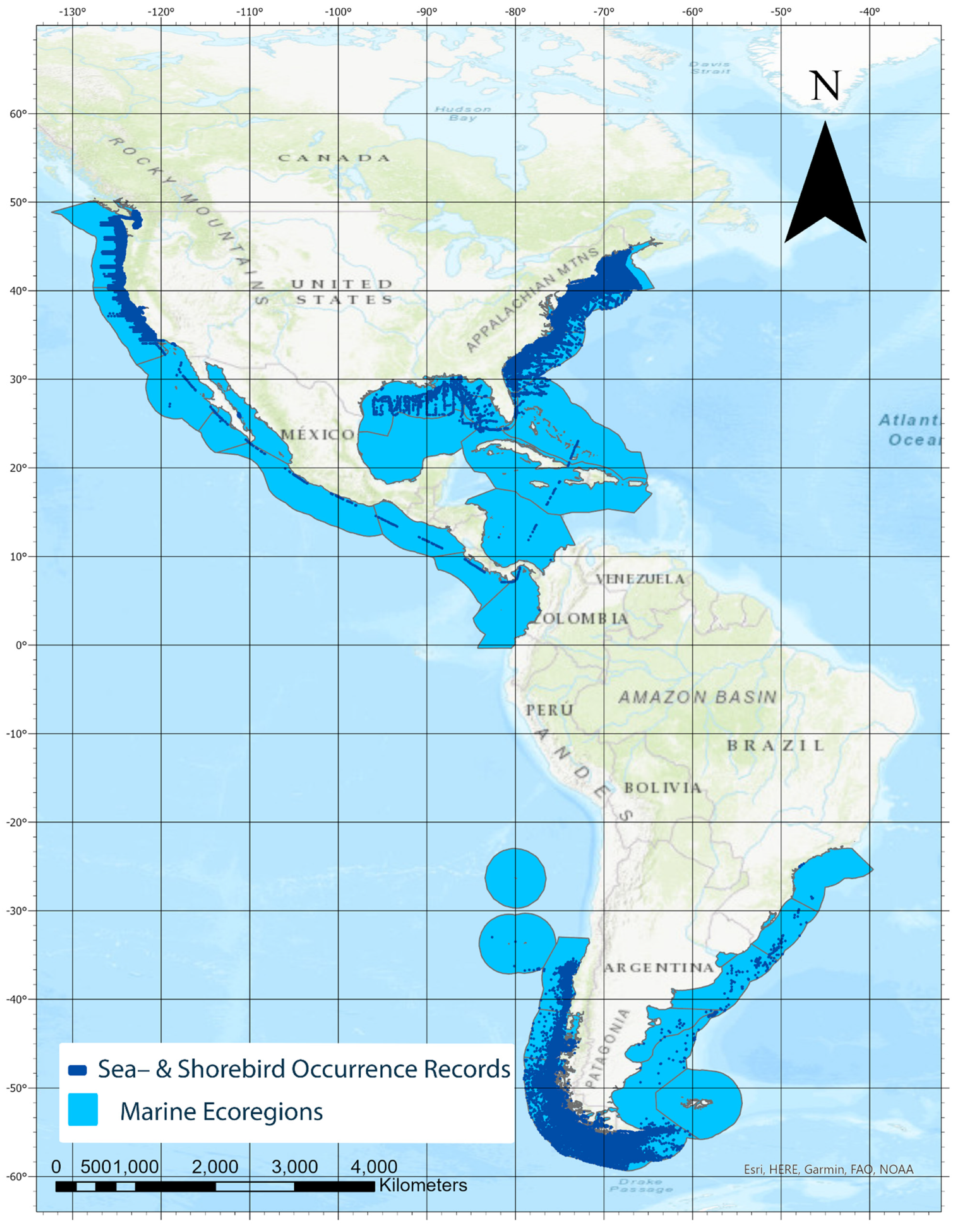
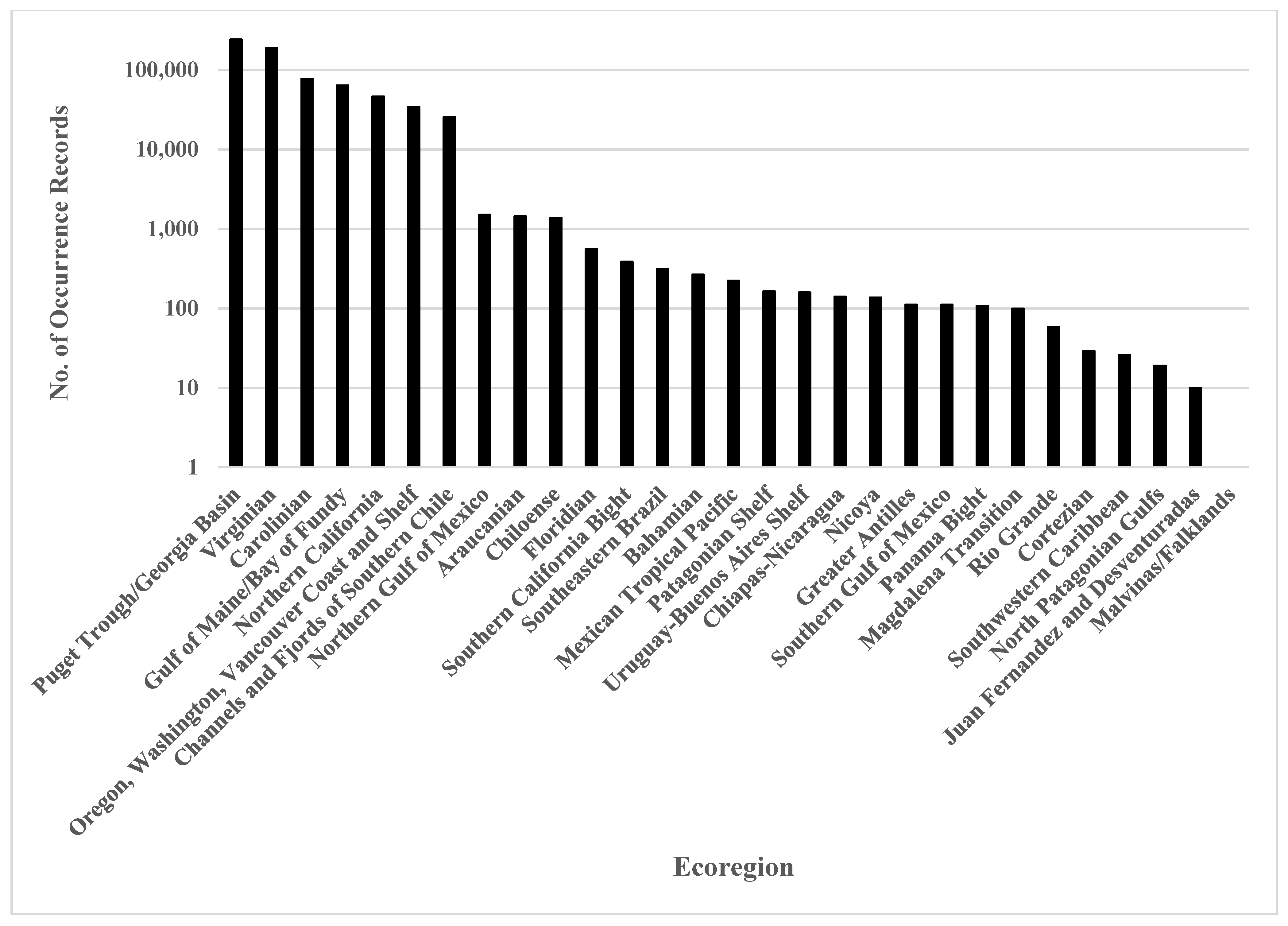
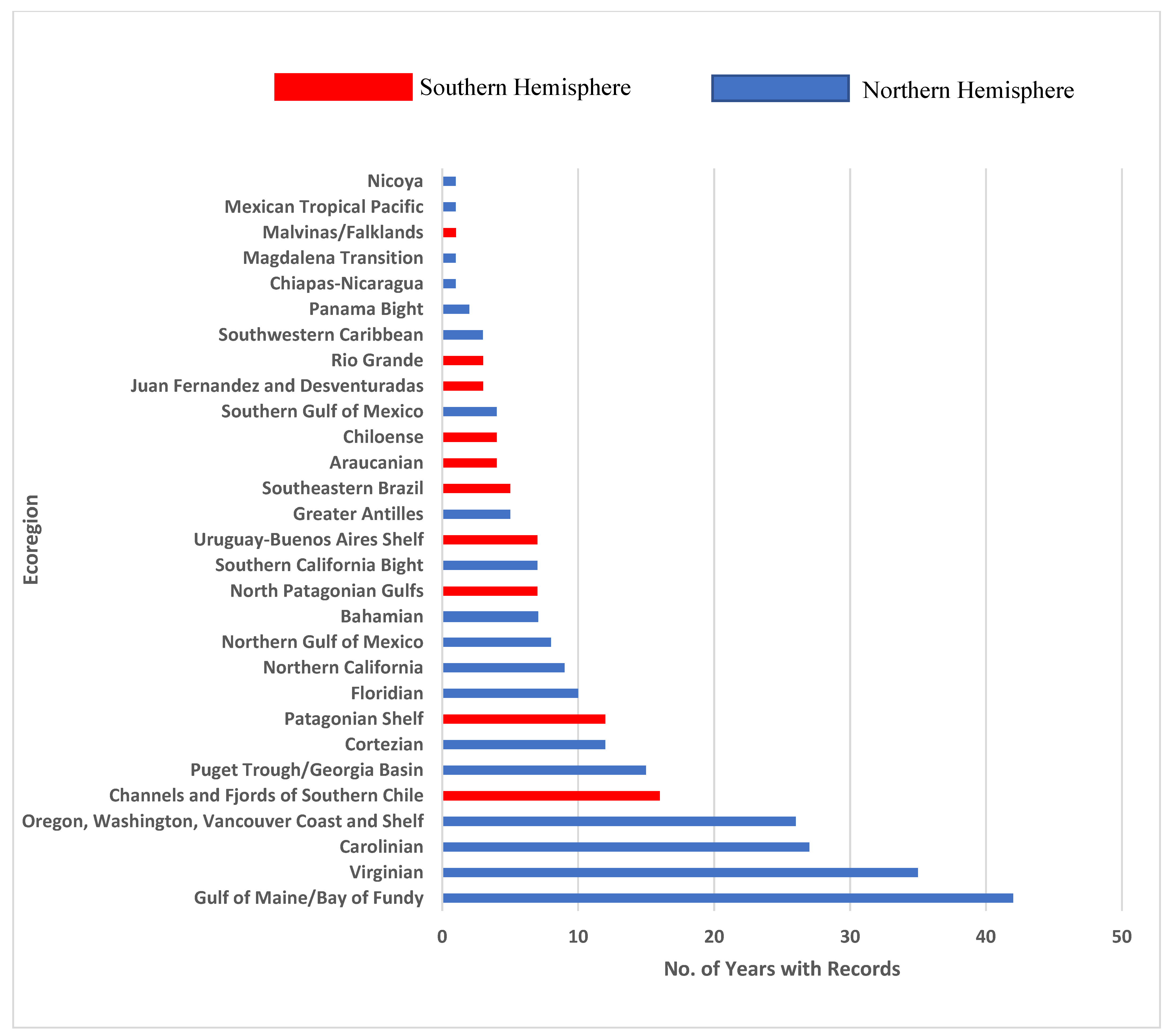
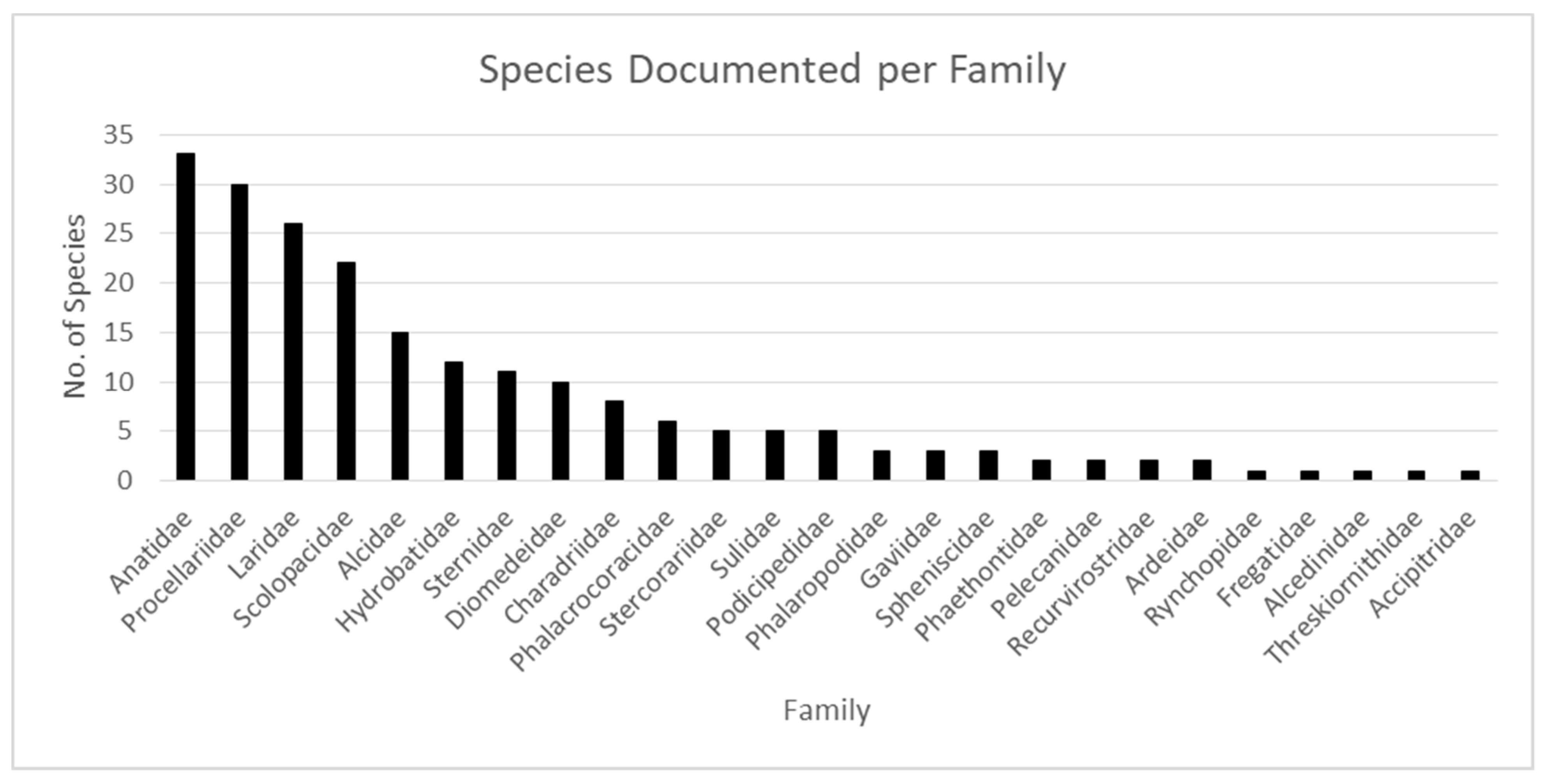
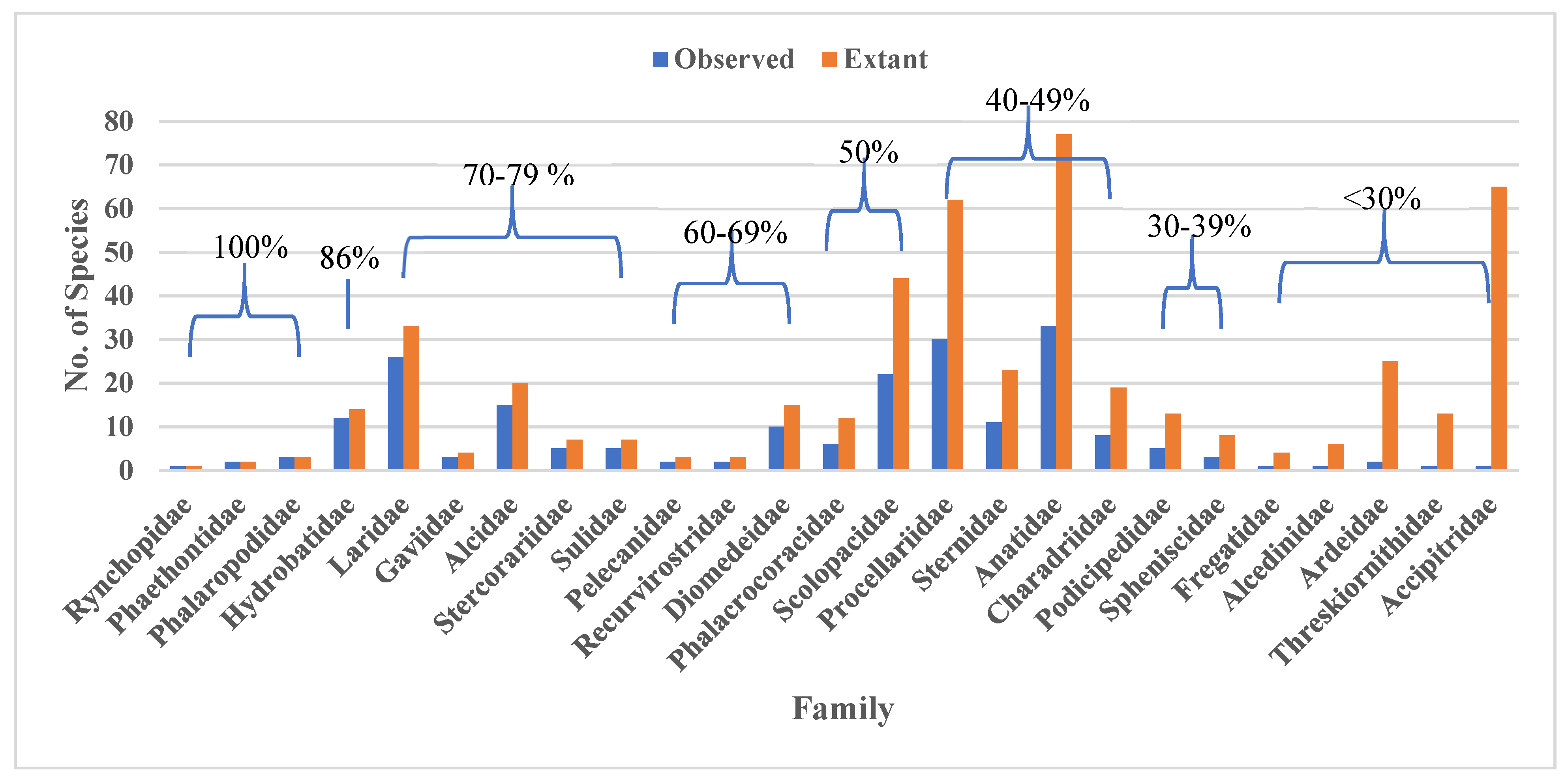
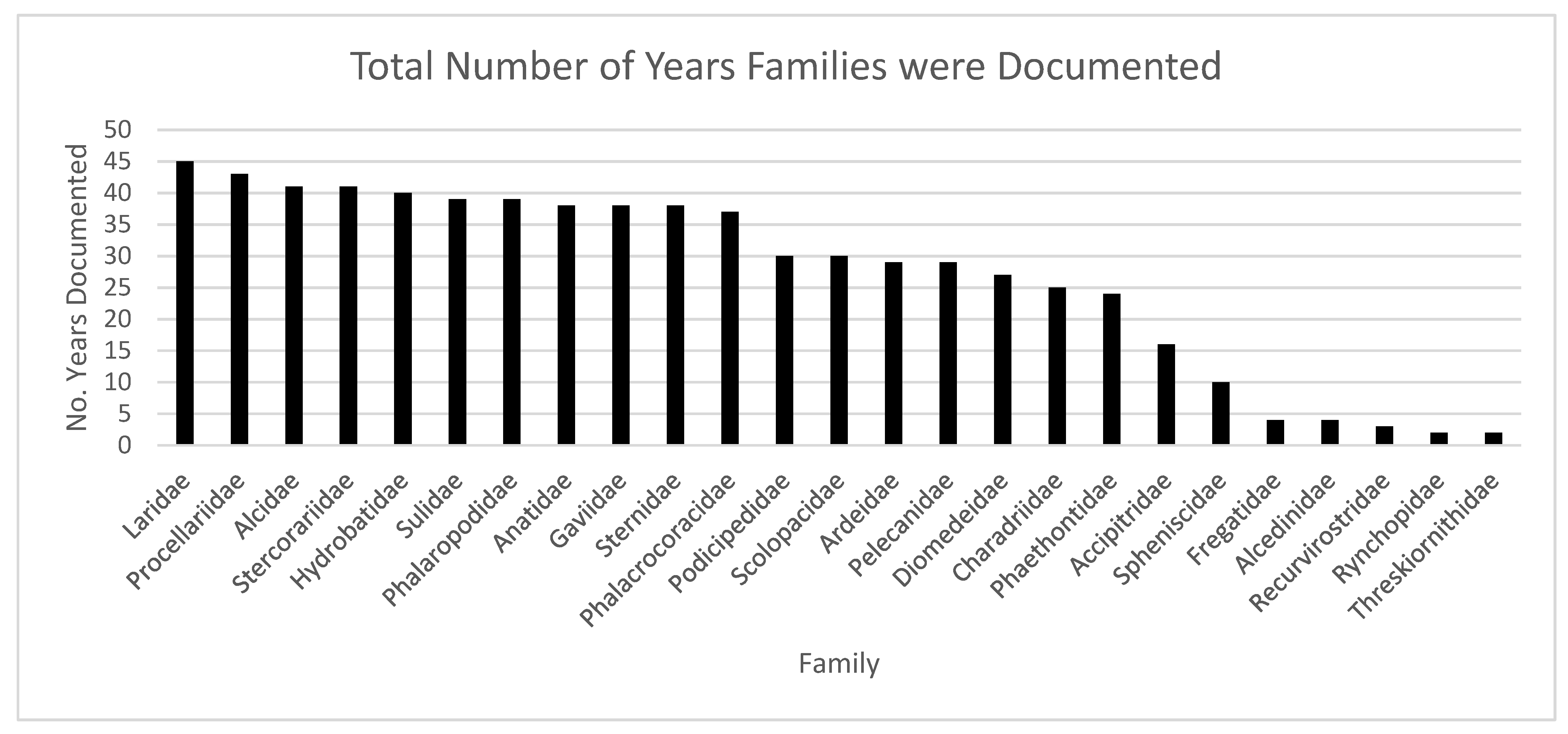
3. Results
4. Discussion
5. Conclusions
- The OBIS platform can provide baseline data for sea- and shorebird distributions in future studies for several North American marine ecoregions.
- More observations, especially from Central and South American ecoregions and from the Caribbean Sea, should be shared via open-access databases like OBIS for monitoring and ecological analyses within and across those marine ecoregions.
- Capacity development focused on best practices for seabird and environmental observations, including standardized meta-data formatting and sharing of observations via open databases like OBIS is important to improve the scientific understanding of sea- and shorebirds, as well as providing opportunities for scientists to contribute to the growth and well-being of open-access data initiatives.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Biosketch
References
- Dunnet, G.M.; Crisp, D.J.; Conan, G.; Bourne, W.R.P. Oil pollution and seabird populations. Philos. Trans. R. Soc. B Biol. Sci. 1982, 297, 413–427. [Google Scholar] [CrossRef]
- Valle, C.A.; Cruz, F.; Cruz, J.B.; Merlen, G.; Coulter, M.C. The impact of the 1982–1983 El-Nino-Southern Oscillation on seabirds in the Galapagos-Islands, Equador. J. Geophys. Res.-Oceans 1987, 92, 14437–14444. [Google Scholar] [CrossRef]
- Martini, R.; Mangini, P.R.; Lange, R.R. Seabirds health and conservation medicine in Brazil. J. Nat. Cons. 2022, 69, 126238. [Google Scholar] [CrossRef]
- Hortal, J.; de Bello, F.; Diniz-Filho, J.A.F.; Lewinsohn, T.M.; Lobo, J.M.; Ladle, R.J. Seven shortfalls that beset large-scale knowledge of biodiversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 523–549. [Google Scholar] [CrossRef]
- Orgeret, F.; Thiebault, A.; Kovacs, K.M.; Lydersen, C.; Hindell, M.A.; Thompson, S.A.; Sydeman, W.J.; Pistorius, P.A. Climate change impacts on seabirds and marine mammals: The importance of study duration, thermal tolerance and generation time. Ecol. Lett. 2022, 25, 218–239. [Google Scholar] [CrossRef]
- Morera-Pujol, V.; Catry, P.; Magalhães, M.; Péron, C.; Reyes-González, J.M.; Granadeiro, J.P.; Militão, T.; Dias, M.P.; Oro, D.; Dell’Omo, G.; et al. Methods to detect spatial biases in tracking studies caused by differential representativeness of individuals, populations and time. Divers. Distrib. 2022, 00, 1–20. [Google Scholar] [CrossRef]
- Ocean Biogeographic Information System (OBIS). Intergovernmental Oceanographic Commission of UNESCO. Available online: https://obis.org (accessed on 23 October 2022).
- Grassle, J.F. The Ocean Biogeographic Information System (OBIS): An on-line, worldwide atlas for accessing, modeling and mapping marine biological data in a multidimensional geographic context. Oceanography 2000, 13, 5–7. [Google Scholar] [CrossRef]
- Provoost, P.; Appeltans, W.; Lambert, A. OBIS Github. Available online: https://github.com/iobis (accessed on 1 October 2021).
- Wieczorek, J.; Bloom, D.; Guralnick, R.; Blum, S.; Döring, M.; Giovanni, R.; Robertson, T.; Vieglais, D. Darwin Core: An evolving community-developed biodiversity data standard. PLoS ONE 2012, 7, e29715. [Google Scholar] [CrossRef]
- eBird: An Online Database of Bird Distribution and Abundance [Web Application]. eBird, Cornell Lab of Ornithology, Ithaca, New York. Available online: http://www.ebird.org (accessed on 24 November 2022).
- Seabird Tracking Database. BirdLife International. Available online: http://www.seabirdtracking.org (accessed on 27 November 2022).
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef]
- Price, M.H. Mastering ArcGIS Pro, 1st ed.; McGraw-Hill Education: New York, NY, USA, 2020; ISBN 978-1-260-58733-3. [Google Scholar]
- Digital Aerial Baseline Survey of Marine Wildlife in Support of Offshore Wind Energy—OPA 2016. Available online: http://seamap.env.duke.edu/dataset/1817 (accessed on 27 October 2022).
- OBIS-SEAMAP: The World Data Center for Marine Mammal, Seabird, Sea Turtle, Shark & Ray Distributions. Available online: https://seamap.env.duke.edu/ (accessed on 27 October 2022).
- PIROP Northwest Pacific 1965–1992. Available online: https://seamap.env.duke.edu/dataset/280 (accessed on 1 December 2022).
- SEFSC Caribbean Survey 1995 Dataset. Available online: https://seamap.env.duke.edu/dataset/11 (accessed on 1 December 2022).
- Tracy, D.M.; Schamel, D.; Dale, J. Red Phalarope (Phalaropus Fulicarius), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Albatrosses (Diomedeidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Auks, Murres, and Puffins (Alcidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Cormorants and Shags (Phalacrocoracidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Ducks, Geese, and Waterfowl (Anatidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Frigatebirds (Fregatidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Grebes (Podicipedidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Gulls, Terns, and Skimmers (Laridae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Hawks, Eagles, and Kites (Accipitridae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Herons, Egrets, and Bitterns (Ardeidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Ibises and Spoonbills (Threskiornithidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Kingfishers (Alcedinidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Loons (Gaviidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Northern Storm-Petrels (Hydrobatidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Pelicans (Pelecanidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Penguins (Spheniscidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Plovers and Lapwings (Charadriidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Sandpipers and Allies (Scolopacidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Shearwaters and Petrels (Procellariidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Stilts and Avocets (Recurvirostridae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Tropicbirds (Phaethontidae), Version 1.0. In Birds of the World; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Sculenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Cornell Lab of Ornithology, Birds of the World. Available online: https://birdsoftheworld.org/bow/home (accessed on 1 October 2021).
- About OBIS. Available online: https://obis.org/about/ (accessed on 27 October 2022).
- Auer, T.; Barker, S.; Borgmann, K.; Charnoky, M.; Childs, D.; Curtis, J.; Davies, I.; Downie, I.; Fink, D.; Fredericks, T.; et al. EOD—EBird Observation Dataset; Cornell Lab of Ornithology: Ithaca, NY, USA, 2022. [Google Scholar] [CrossRef]
- Boakes, E.H.; McGowan, P.J.K.; Fuller, R.A.; Chang-Qing, D.; Clark, N.E.; O’Connor, K.; Mace, G.M. Distorted views of biodiversity: Spatial and temporal bias in species occurrence data. PLOS Biol. 2010, 8, e1000385. [Google Scholar] [CrossRef] [PubMed]
- Kot, C.Y.; Fujioka, E.; Hazen, L.J.; Best, B.D.; Read, A.J.; Halpin, P.N. Spatio-temporal gap analysis of OBIS-SEAMAP project data: Assessment and way forward. PLoS ONE 2010, 5, e12990. [Google Scholar] [CrossRef] [PubMed]
- Leles, S.G.; Mitra, A.; Flynn, K.J.; Tillmann, U.; Stoecker, D.; Jeong, H.J.; Burkholder, J.; Hansen, P.J.; Caron, D.A.; Gilbert, P.M.; et al. Sampling bias misrepresents the biogeographical significance of constitutive mixotrophs across global oceans. Glob. Ecol. Biogeogr. 2019, 28, 418–428. [Google Scholar] [CrossRef]
- Pardo, I.; Pata, M.P.; Gómez, D.; García, M.B. A novel method to handle the effect of uneven sampling effort in biodiversity databases. PLoS ONE 2013, 8, e52786. [Google Scholar] [CrossRef]
- Ruete, A. Displaying bias in sampling effort of data accessed from biodiversity databases using ignorance maps. Biodivers. Data J. 2015, 3, e5361. [Google Scholar] [CrossRef] [PubMed]
- Munira, A.N.; Nurul Salmi, A.L.; Shahrul Anuar, M.S.; Mohd Abdul Muin, M.A.; Amirrudin, A.; Juliani, S. Nur. Diversity and temporal distribution of birds in rice-growing landscape, northern peninsular Malaysia. Sains Malays. 2014, 43, 513–520. [Google Scholar]
- Dias, M.P.; Oppel, S.; Bond, A.L.; Carneiro, A.P.B.; Cuthbert, R.J.; González-Solís, J.; Wanless, R.M.; Glass, T.; Lascelles, B.; Small, C.; et al. Using globally threatened pelagic birds to identify priority sites for marine conservation in the South Atlantic Ocean. Biol. Conserv. 2017, 211, 76–84. [Google Scholar] [CrossRef]
- Hyrenbach, K.D.; Veit, R.R.; Weimerskirch, H.; Metzl, N.; Hunt, G.L. Community structure across a large-scale ocean productivity gradient: Marine bird assemblages of the southern Indian Ocean. Deep-Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 1129–1145. [Google Scholar] [CrossRef]
- Perry, J.; Kutt, A.; Perkins, G.; Vanderduys, E.; Colman, N. A bird survey method for Australian tropical savannas. Emu-Austral Ornithol. 2012, 112, 261–266. [Google Scholar] [CrossRef]
- Powers, K. Pelagic distributions of marine birds off the northeastern United States. In NOAA Technical Memorandum; NMFS-F/NEC-27; NOAA: Washington, DC, USA, 1983; pp. 1–201. [Google Scholar]
- Brunet, M.; Jones, P. Data rescue initiatives: Bringing historical climate data into the 21st century. Clim. Res. 2011, 47, 29–40. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Diniz-Filho, J.A.F.; Jaramillo, C.A.; Soeller, S.A. Climate, niche conservatism, and the global bird diversity gradient. Am. Nat. 2007, 170, S16–S27. [Google Scholar] [CrossRef] [PubMed]
- Oro, D. Seabirds and climate: Knowledge, pitfalls, and opportunities. Front. Ecol. Evol. 2014, 2, 79. [Google Scholar] [CrossRef]
- Knockaert, C.; Tyberghein, L.; Goffin, A.; Vanhaecke, D.; Ong’anda, H.; Wakwabi, E.O.; Mees, J. Biodiversity data rescue in the framework of a long-term Kenya-Belgium cooperation in marine sciences. Sci. Data 2019, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.; Nost, E.; Lemelin, A.; Lave, R.; Dillon, L. Practicing environmental data justice: From DataRescue to Data Together. Geo. Geogr. Environ. 2018, 5, e00061. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.; da Silva Santos, L.B.; Bourne, P.E.; et al. The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef] [PubMed]
- Bayraktarov, E.; Ehmke, G.; O’Connor, J.; Burns, E.L.; Nguyen, H.A.; McRae, L.; Possingham, H.P.; Lindenmayer, D.B. Do big unstructured biodiversity data mean more knowledge? Front. Ecol. Evol. 2019, 6, 239. [Google Scholar] [CrossRef]
- Croxall, J.P.; Butchart, S.H.M.; Lascelles, B.; Stattersfield, A.J.; Sullivan, B.; Symes, A.; Taylor, P. Seabird conservation status, threats and priority actions: A global assessment. Bird Conserv. Int. 2012, 22, 1–34. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Jablonski, D. The tropics as a source of evolutionary novelty through geological time. Nature 1993, 364, 142–144. [Google Scholar] [CrossRef]
- Jablonski, D.; Roy, K.; Valentine, J.W. Out of the tropics: Evolutionary dynamics of the latitudinal diversity gradient. Science 2006, 314, 102–106. [Google Scholar] [CrossRef]
- Kennedy, J.D.; Wang, Z.; Weir, J.T.; Rahbek, C.; Fjeldså, J.; Price, T.D. Into and out of the tropics: The generation of the latitudinal gradient among New World passerine birds. J. Biogeogr. 2014, 41, 1746–1757. [Google Scholar] [CrossRef]
- Smith, B.T.; Seeholzer, G.F.; Harvey, M.G.; Cuervo, A.M.; Brumfield, R.T. A latitudinal phylogeographic diversity gradient in birds. PLOS Biol. 2017, 15, e2001073. [Google Scholar] [CrossRef] [PubMed]
- Chown, S.L.; Gaston, K.J.; Williams, P.H. Global patterns in species richness of pelagic seabirds: The Procellariiformes. Ecography 2006, 21, 342–350. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family Name | Common Name/s | Habitat | # Species | # spp in OBIS | # Occurrence Records | No. of Each Marine Ecoregion with Records |
|---|---|---|---|---|---|---|
| Anatidae | Ducks, Geese | Coastal/inland | 174 | 33 | 158,392 | 3, 10, 17, 19, 22, 29 |
| Laridae | Gulls | Coastal/inland | 51 | 26 | 150,416 | 2, 3, 5, 7–10, 12, 14, 15, 17–20, 22, 24–27, 29 |
| Procellariidae | Petrels, Shearwaters | Pelagic | 96 | 30 | 72,975 | 2–5, 7–12, 14–22, 25–29 |
| Alcidae | Auks, Murres, Puffins | Cold water, coastal | 25 | 15 | 72,386 | 3, 10, 17, 19, 22, 25, 29 |
| Sulidae | Boobies, Gannets | Pelagic | 10 | 5 | 44,381 | 2, 3, 5, 7–10, 14, 15, 18 20, 26, 27, 29 |
| Gaviidae | Loons | Coastal/inland | 5 | 3 | 42,642 | 3, 10, 17, 19, 22, 29 |
| Diomedeidae | Albatrosses | Pelagic | 15 | 10 | 29,730 | 1, 4, 6, 11–13, 16, 17, 19, 21, 23–25, 28 |
| Hydrobatidae | Northern Storm Petrels | Pelagic | 18 | 12 | 28,277 | 2–5, 7–10, 12, 14, 15, 17–20, 25–29 |
| Phalacrocoracidae | Cormorants, Shags | Coastal & pelagic | 40 | 6 | 20,867 | 3, 8, 10, 17–19, 22, 29 |
| Podicipedidae | Grebes | Coastal/inland | 22 | 5 | 20,680 | 3, 17, 19, 22, 29 |
| Phalaropodidae | Phalaropes | Shorebirds | 3 | 2 | 16,425 | 3, 10, 14, 17, 19, 22, 25, 29 |
| Sternidae | Terns | Inland, coastal & pelagic | 45 | 11 | 10,318 | 2, 3, 8–10, 14, 15, 17–20, 22, 24, 26, 29 |
| Ardeidae | Herons, Egrets, Bitterns | Shorebirds | 68 | 2 | 7,526 | 3, 8, 10, 17, 19, 20, 22, 29 |
| Stercorariidae | Skuas | Cold water, coastal | 7 | 5 | 3,202 | 2, 3, 8–10, 14, 17–19, 22, 25, 26, 28, 29 |
| Scolopacidae | Sandpipers | Shorebirds | 97 | 22 | 2,373 | 3, 10, 17, 19, 20, 22, 27, 29 |
| Accipitridae | Hawks, Eagles, Kites | Mostly terrestrial | 250 | 1 | 2,224 | 10, 19, 22, 29 |
| Pelecanidae | Pelicans | Coastal/inland | 8 | 2 | 1,104 | 7, 8, 17–20, 22, 25, 26, 29 |
| Spheniscidae | Penguins | Southern cold waters | 18 | 3 | 712 | 4, 21 |
| Charadriidae | Plovers, Lapwings | Coastal/inland | 68 | 8 | 549 | 3, 7, 10, 19, 20, 22, 24, 29 |
| Phaethontidae | Tropicbirds | Islands & pelagic | 3 | 2 | 157 | 2, 3, 5, 7, 8, 10, 14, 15, 26, 29 |
| Rynchopidae | Skimmers | Coastal | 3 | 1 | 65 | 24, 29 |
| Fregatidae | Frigatebirds | Coastal & pelagic | 5 | 1 | 27 | 2, 3, 5, 8, 14, 15, 20, 27 |
| Recurvirostridae | Stilts, Avocets | Coastal/inland | 9 | 2 | 17 | 3, 17 |
| Alcedinidae | Kingfishers | Coastal/inland | 118 | 1 | 7 | 3, 10, 27 |
| Threskiornithidae | Ibises, Spoonbills | Shorebirds | 36 | 1 | 2 | 3, 27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hartman, S.; Hallock, P.; Muller-Karger, F. Obligation to Enhance OBIS Data for Sea- and Shorebirds of the Americas. Diversity 2022, 14, 1099. https://doi.org/10.3390/d14121099
Hartman S, Hallock P, Muller-Karger F. Obligation to Enhance OBIS Data for Sea- and Shorebirds of the Americas. Diversity. 2022; 14(12):1099. https://doi.org/10.3390/d14121099
Chicago/Turabian StyleHartman, Savannah, Pamela Hallock, and Frank Muller-Karger. 2022. "Obligation to Enhance OBIS Data for Sea- and Shorebirds of the Americas" Diversity 14, no. 12: 1099. https://doi.org/10.3390/d14121099
APA StyleHartman, S., Hallock, P., & Muller-Karger, F. (2022). Obligation to Enhance OBIS Data for Sea- and Shorebirds of the Americas. Diversity, 14(12), 1099. https://doi.org/10.3390/d14121099