
Vegetation Ecology of Debris-Covered Glaciers (DCGs)—Site Conditions, Vegetation Patterns and Implications for DCGs Serving as Quaternary Cold- and Warm-Stage Plant Refugia

 ,
, 
{kind=link}
{kind=link}
{kind=link}
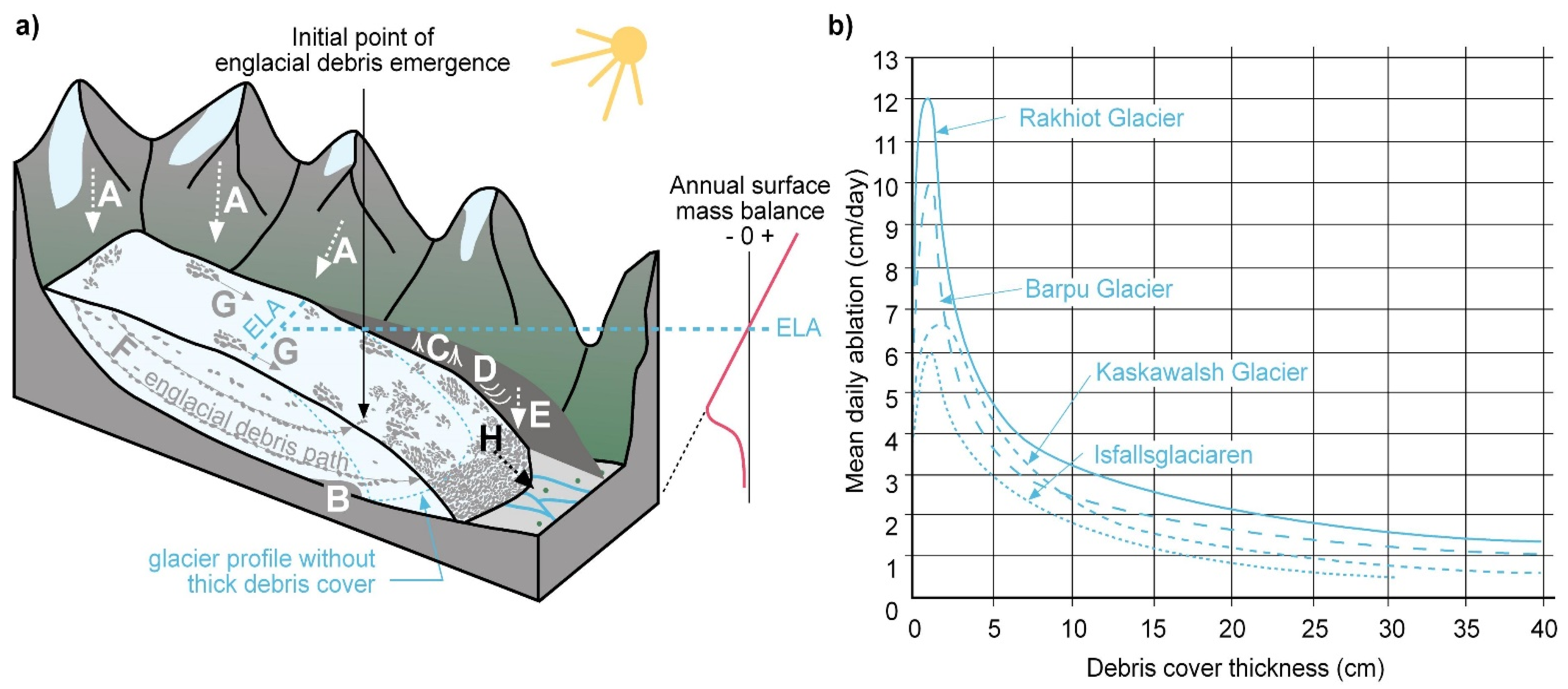{kind=link}
{kind=link}
{kind=link}
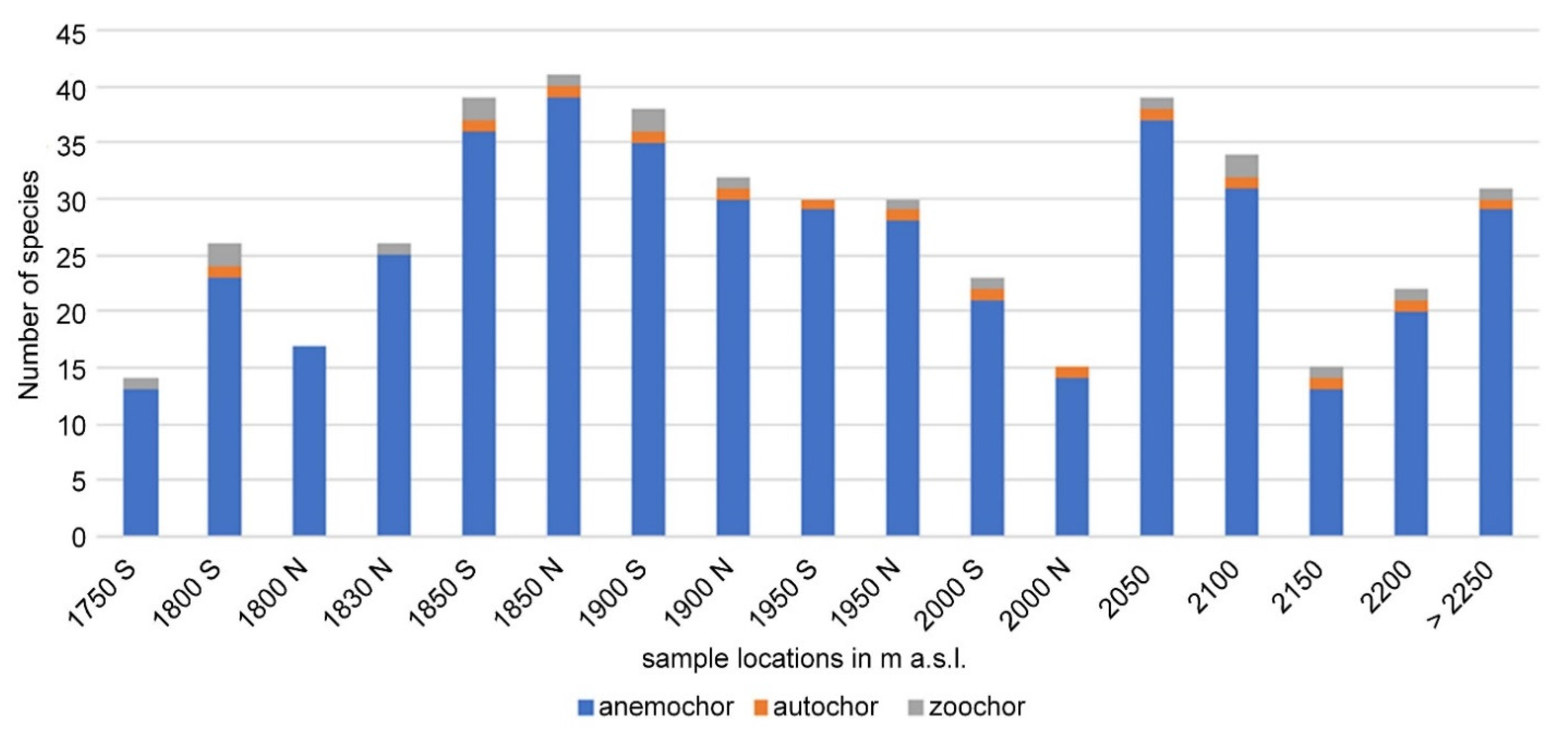{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Origin of Debris Cover and Its Effect on Glacier Movement and Glacier Mass Balance
3. DCGs as Habitats for Plants
3.1. Physical Setting of DCGs
3.2. Source Areas and Dispersal Pathways of Plants Colonizing DCGs
3.3. Spatiotemporal Plant Diversity Patterns on DCGs
4. DCGs as Potential Cold Stage Refugia in the Past
5. The Role of DCGs and Other Cold Rocky Landforms as Refugia under Current Climate Warming
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Materials and Methods
References
- Scherler, D.; Wulf, H.; Gorelick, N. Global assessment of supraglacial debris-cover extents. Geophys. Res. Lett. 2018, 45, 11798–11805. [Google Scholar] [CrossRef] [Green Version]
- Herreid, S.; Pellicciotti, F. The state of rock debris covering Earth’s glaciers. Nat. Geosci. 2020, 13, 621–627. [Google Scholar] [CrossRef]
- Zemp, M.; Frey, H.; Gärtner-Roer, I.; Nussbaumer, S.; Hoelzle, M.; Paul, F.; Haeberli, W.; Denzinger, F.; Ahlstrom, A.P.; Anderson, B.; et al. Historically unprecedented global glacier decline in the early 21st century. J. Glaciol. 2015, 61, 745–762. [Google Scholar] [CrossRef] [Green Version]
- Hugonnet, R.; McNabb, R.; Berthier, E.; Menounos, B.; Nuth, C.; Girod, L.; Farinotti, D.; Huss, M.; Dussaillant, I.; Brun, F.; et al. Accelerated global glacier mass loss in the early twenty-first century. Nature 2021, 592, 726–731. [Google Scholar] [CrossRef]
- Farinotti, D.; Immerzeel, W.W.; de Kok, R.J.; Quincey, D.J.; Dehecq, A. Manifestations and mechanisms of the Karakoram glacier. Anomaly. Nat. Geosci. 2020, 13, 8–16. [Google Scholar] [CrossRef]
- Haeberli, W.; Noetzli, J.; Arenson, L.; Delaloye, R.; Gärtner-Roer, I.; Gruber, S.; Isaksen, K.; Kneisel, C.; Krautblatter, M.; Phillips, M. Mountain permafrost: Development and challenges of a young research field. J. Glaciol. 2011, 56, 1043–1058. [Google Scholar] [CrossRef] [Green Version]
- McColl, S.T. Paraglacial rock-slope stability. Geomorphology 2012, 153–154, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Stoffel, M.; Huggel, C. Effects of climate change on mass movements in mountain environments. Prog. Phys. Geog. 2012, 36, 421–439. [Google Scholar] [CrossRef]
- Wahrhaftig, C.; Cox, A. Rock glaciers in the Alaska Range. Geol. Soc. Am. Bull. 1959, 70, 383–436. [Google Scholar] [CrossRef]
- Barsch, D. Rockglaciers. Indicators for the Present and Former Geoecology in High Mountain Environments; (Springer Series in Physical Environment Book Series); Springer: Berlin, Germany, 1996; Volume 16. [Google Scholar]
- Haeberli, W. Modern Research Perspectives Relating to Permafrost Creep and Rock Glaciers: A Discussion. Permafr. Periglac. 2000, 11, 290–293. [Google Scholar] [CrossRef]
- Berthling, I. Beyond confusion: Rock glaciers as cryo-conditioned landforms. Geomorphology 2011, 131, 98–106. [Google Scholar] [CrossRef]
- Whalley, B.; Matsuoka, N.; Sik, A.; Kereszturi, A.; Hargitai, H. Rock Glacier and Debris-Covered Glacier. In Encyclopedia of Planetary Landforms; Hargitai, H., Kereszturi, A., Eds.; Springer: New York, NY, USA, 2015; pp. 1811–1828. [Google Scholar] [CrossRef]
- Kirkbride, M.P. Debris-Covered Glaciers. In Encyclopedia of Snow, Ice and Glaciers; Singh, V.P., Singh, P., Haritashya, U.K., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 190–192. [Google Scholar] [CrossRef]
- Johnson, P.G. Glacier—Rock glacier transition in the Southwest Yukon Territory, Canada. Arct. Alp. Res. 1980, 12, 195–204. [Google Scholar] [CrossRef]
- Whalley, B.; Martin, H. Rock glaciers: II models and mechanisms. Prog. Phys. Geog. 1992, 16, 127–186. [Google Scholar] [CrossRef]
- Anderson, R.; Anderson, L.; Armstrong, W.; Rossi, M.; Crump, S. Glaciation of alpine valleys: The glacier—Debris-covered glacier—Rock glacier continuum. Geomorphology 2018, 311, 127–142. [Google Scholar] [CrossRef]
- Fountain, A.G.; Raymond, C.F.; Nakawo, M. International Workshop examines debris-covered glaciers. Eos 2001, 82, 261–262. [Google Scholar]
- Nicholson, L.; Benn, D.I. Calculating ice melt beneath a debris layer using meteorological data. J. Glaciol. 2006, 52, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Reid, T.; Brock, B. An energy-balance model for debris-covered glaciers including heat conduction through the debris layer. J. Glaciol. 2010, 56, 903–916. [Google Scholar] [CrossRef] [Green Version]
- Evatt, G.; Abrahams, I.; Heil, M.; Mayer, C.; Kingslake, J.; Michell, S.; Fowler, A.; Clark, C. Glacial melt under a porous debris layer. J. Glaciol. 2015, 61, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Naegeli, K.; Huss, M. Sensitivity of mountain glacier mass balance to changes in bare-ice albedo. Ann. Glaciol. 2017, 58, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Fyffe, C.L.; Brock, B.W.; Kirkbride, M.P.; Mair, D.W.F.; Arnold, N.S.; Smiraglia, C.; Diolaiuti, G.; Diotri, F. Do debris-covered glaciers demonstrate distinctive hydrological behaviour compared to clean glaciers? J. Hydrol. 2019, 570, 584–597. [Google Scholar] [CrossRef]
- Ferguson, J.C.; Vieli, A. Modelling steady states and the transient response of debris-covered glaciers. Cryosphere 2021, 15, 3377–3399. [Google Scholar] [CrossRef]
- Huo, D.; Chi, Z.; Ma, A. Modeling Surface Processes on Debris-Covered Glaciers: A Review with Reference to the High Mountain Asia. Water 2021, 13, 101. [Google Scholar] [CrossRef]
- Mayr, E.; Hagg, W. Debris-Covered Glaciers. In Geomorphology of Proglacial Systems. Geography of the Physical Environment; Heckmann, T., Morche, D., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Rounce, D.; Hock, R.; McNabb, R.; Millan, R.; Sommer, C.; Braun, M.; Malz, P.; Maussion, F.; Mouginot, J.; Seehaus, T.; et al. Distributed Global Debris Thickness Estimates Reveal Debris Significantly Impacts Glacier Mass Balance. Geophys. Res. Lett. 2021, 48, e2020GL091311. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A. Brief Communication: Thinning of debris-covered and debris-free glaciers in a warming climate. Cryosphere 11 2017, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Salerno, F.; Thakuri, S.; Tartari, G.; Nuimura, T.; Sunako, S.; Sakai, A.; Fujita, K. Debris-covered glacier anomaly? Morphological factors controlling changes in the mass balance, surface area, terminus position, and snow line altitude of Himalayan glaciers. Earth Planet Sci. Lett. 2017, 471, 19–31. [Google Scholar] [CrossRef]
- Huo, D.; Bishop, M.P.; Bush, A.B.G. Understanding Complex Debris-Covered Glaciers: Concepts, Issues, and Research Directions. Front. Earth Sci. 2021, 9, 358. [Google Scholar] [CrossRef]
- Pfeffer, W.T.; Arendt, A.A.; Bliss, A.; Bolch, T.; Cogley, J.G.; Gardner, A.S.; Hagen, J.-O.; Hock, R.; Kaser, G.; Kienholz, C.; et al. The Randolph Glacier Inventory: A globally complete inventory of glaciers. J. Glaciol. 2014, 60, 537–552. [Google Scholar] [CrossRef] [Green Version]
- RGI Consortium. Randolph Glacier Inventory—A Dataset of Global Glacier Outlines: Version 6.0: Technical Report, Global Land Ice Measurements from Space; Digital Media; RGI Consortium: Boulder, CO, USA, 2017. [Google Scholar] [CrossRef]
- WGMS; NSIDC. World Glacier Inventory. Compiled and Made Available by the World Glacier Monitoring Service; Updated 2012; Zurich, Switzerland, and The National Snow and Ice Data Center: Boulder, CO, USA, 1989. [Google Scholar] [CrossRef]
- Raup, B.; Racoviteanu, A.; Khalsa, S.J.S.; Helm, C.; Armstrong, R.; Arnaud, Y. The GLIMS geospatial glacier database: A new tool for studying glacier change. Glob. Planet Change 2007, 56, 101–110. [Google Scholar] [CrossRef]
- GLIMS; NSIDC. Global Land Ice Measurements from Space Glacier Database; Compiled and made available by the international GLIMS community and the National Snow and Ice Data Center, updated 2018; National Snow and Ice Data Center: Boulder, CO, USA, 2005. [Google Scholar] [CrossRef]
- GlaThiDa Consortium. Glacier Thickness Database 3.1.0; World Glacier Monitoring Service: Zurich, Switzerland, 2020. [Google Scholar] [CrossRef]
- Welty, E.; Zemp, M.; Navarro, F.; Huss, M.; Fürst, J.J.; Gärtner-Roer, I.; Landmann, J.; Machguth, H.; Naegeli, K.; Andreassen, L.M.; et al. Worldwide version-controlled database of glacier thickness observations. Earth Syst. Sci. Data 2020, 12, 3039–3055. [Google Scholar] [CrossRef]
- Paul, F.; Huggel, C.; Kääb, A. Combining satellite multispectral image data and a digital elevation model for mapping debris-covered glaciers. Remote Sens. Environ. 2004, 89, 510–518. [Google Scholar] [CrossRef]
- Brenning, A.; Trombotto, D. Logistic regression modeling of rock glacier and glacier distribution: Topographic and climatic controls in the semi-arid Andes. Geomorphology 2006, 81, 141–154. [Google Scholar] [CrossRef]
- Brenning, A.; Grasser, M.; Friend, D.A. Statistical estimation and generalized additive modeling of rock glacier distribution in the San Juan Mountains, Colorado, USA. J. Geophys. Res. Earth 2007, 112, F02S15. [Google Scholar] [CrossRef]
- Molnia, B.F. Satellite Image Atlas of the Glaciers of the World: Alaska; US Geological Survey Professional Paper 1386-K; U.S. Geological Survey: Reston, VA, USA, 2008; 750p.
- Shukla, A.; Gupta, R.P.; Arora, M.K. Delineation of debris-covered glacier boundaries using optical and thermal remote sensing data. Remote Sens. Lett. 2010, 1, 11–17. [Google Scholar] [CrossRef]
- Xie, Z.; Haritashya, U.; Asari, V.; Young, B.; Bishop, M.; Kargel, J. GlacierNet: A Deep-Learning Approach for Debris-Covered Glacier Mapping. IEEE Access 2020, 8, 83495–83510. [Google Scholar] [CrossRef]
- Richter, M.; Fickert, T.; Grüninger, F. Pflanzen auf schuttbedeckten Gletschern—Wandernde Kuriositäten. Geoöko 2004, 25, 225–256. [Google Scholar]
- Fickert, T.; Friend, D.; Grüninger, F.; Molnia, B.; Richter, M. Did Debris-Covered Glaciers Serve as Pleistocene Refugia for Plants? A New Hypothesis Derived from Observations of Recent Plant Growth on Glacier Surfaces. Arct. Antarct. Alp. Res. 2007, 39, 245–257. [Google Scholar] [CrossRef]
- Pelfini, M.; Santilli, M.; Leonelli, G.; Bozzoni, M. Investigating surface movements of debris-covered Miage Glacier (Western Italian Alps) using dendroglaciological analysis. J. Glaciol. 2007, 53, 141–152. [Google Scholar] [CrossRef]
- Caccianiga, M.; Andreis, C.; Diolaiuti, G.; D’Agata, C.; Mihalcea, C.; Smiraglia, C. Alpine debris-covered glaciers as a habitat for plant life. Holocene 2011, 21, 1011–1020. [Google Scholar] [CrossRef]
- Millar, C.I.; Westfall, R.D.; Evenden, A.; Holmquist, J.G.; Schmidt-Gengenbach, J.; Franklin, R.S.; Nachlinger, J.; Delany, D.L. Potential climatic refugia in semi-arid, temperate mountains: Plant and arthropod assemblages associated with rock glaciers, talus slopes, and their forefield wetlands, Sierra Nevada, California, USA. Quatern. Int. 2015, 387, 106–121. [Google Scholar] [CrossRef]
- Tampucci, D.; Citterio, C.; Gobbi, M.; Caccianiga, M. Vegetation outlines of a debris-covered glacier descending below the treeline. Plant Sociol. 2016, 53, 45–54. [Google Scholar] [CrossRef]
- Vezzola, L.; Diolaiuti, G.; D’Agata, C.; Smiraglia, C.; Pelfini, M. Assessing glacier features supporting supraglacial trees: A case study of the Miage debris-covered Glacier (Italian Alps). Holocene 2016, 26, 1138–1148. [Google Scholar] [CrossRef]
- Gobbi, M.; Isaia, M.; De Bernardi, F. Arthropod colonisation of a debris-covered glacier. Holocene 2011, 21, 343–349. [Google Scholar] [CrossRef]
- Gobbi, M.; Ballarin, F.; Brambilla, M.; Compostella, C.; Isaia, M.; Losapio, G.; Maffioletti, C.; Seppi, R.; Tampucci, D.; Caccianiga, M. Life in harsh environments: Carabid and spider trait types and functional diversity on a debris-covered glacier and along its foreland. Ecol. Entomol. 2017, 42, 838–848. [Google Scholar] [CrossRef]
- Gobbi, M. Global warning: Challenges, threats and opportunities for ground beetles (Coleoptera: Carabidae) in high altitude habitats. Acta Zool. Hung. 2020, 66, 5–20. [Google Scholar] [CrossRef]
- Tampucci, D.; Azzoni, R.S.; Boracchi, P.; Citterio, C.; Compostella, C.; Diolaiuti, G.; Isaia, M.; Marano, G.; Smiraglia, C.; Gobbi, M.; et al. Debris-covered glaciers as habitat for plant and arthropod species: Environmental framework and colonization pattern. Ecol. Complex 2017, 32, 42–52. [Google Scholar] [CrossRef]
- Valle, B.; Ambrosini, R.; Caccianiga, M.; Gobbi, M. Ecology of the cold-adapted species Nebria germari (Coleoptera: Carabidae): The role of supraglacial stony debris as refugium during the current interglacial period. Acta Zool. Hung. 2020, 66, 199–220. [Google Scholar] [CrossRef]
- Franzetti, A.; Tatangelo, V.; Gandolfi, I.; Bertolini, V.; Bestetti, G.; Diolaiuti, G.; D’Agata, C.; Mihalcea, C.; Smiraglia, C.; Ambrosini, R. Bacterial community structure on two alpine debris-covered glaciers and biogeography of Polaromonas phylotypes. ISME J. 2013, 7, 1483–1492. [Google Scholar] [CrossRef] [Green Version]
- Losapio, G.; Jordán, F.; Caccianiga, M.; Gobbi, M. Structure-dynamic relationship of plant–insect networks along a primary succession gradient on a glacier foreland. Ecol. Model. 2015, 314, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Brighenti, S.; Hotaling, S.; Finn, D.A.; Fountain, A.G.; Hayashi, M.; Herbst, D.; Saros, J.E.; Tronstad, L.M.; Millar, C.I. Rock glaciers and related cold rocky landforms: Overlooked climate refugia for mountain biodiversity. Glob. Change Biol. Vol. 2021, 27, 1504–1517. [Google Scholar] [CrossRef]
- Matthews, J.A. The Ecology of Recently-Deglaciated Terrain. A Geoecological Approach to Glacier Forelands and Primary Succession; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Erschbamer, B.; Caccianiga, M.S. Glacier forelands: Lessons of plant population and community development. Prog. Bot. 2017, 78, 259–284. [Google Scholar]
- Lüdi, W. Die Pflanzengesellschaften des Lauterbrunnentales und ihre Sukzession; Beiträge zur Geobotanischen Landesaufnahme der Schweiz; Verlag: Zürich, Switzerland, 1921; Volume 9, pp. 1–364. [Google Scholar]
- Negri, G. La vegetazione delle morene del Ghiacciaio del Lys (Monte Rosa). Boll. Del Com. Glaciol. Ital. 1934, 15, 105–172. [Google Scholar]
- Negri, G. Osservazioni di U. Monterin su alcuni casi di invasioni delle morene galleggianti dei ghiacciai del Monte Rosa da parte della vegetazione. Nuovo G. Bot. Ital. 1935, 42, 699–712. [Google Scholar]
- Negri, G. Nuovi dati sull’invasione delle morene galleggianti dei ghiacciai alpini da parte della vegetazione. Nuovo G. Bot. Ital. 1942, 49, 448–459. [Google Scholar]
- Valbusa, U. Florula di due isole glaciali del Rutor con appendice morenica epiglaciale. Nuovo G. Bot. Ital. 1937, 44, 705–714. [Google Scholar]
- Porter, P.R.; Evans, A.J.; Hodson, A.; Lowe, A.T.; Crabtree, M.D. Sediment-moss interactions on a temperate glacier: Falljökull, Iceland. Ann. Glaciol. 2008, 48, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Russell, I.C. An expedition to Mount St. Elias. Natl. Geogr. 1891, 3, 53–191. [Google Scholar]
- Russell, I.C. Second Expedition to Mount St. Elias; U.S. Geological Survey 13th Annual Report Part 2; US Government Printing Office: Washington, DC, USA, 1893; pp. 1–91.
- Tarr, R.S.; Martin, L. Alaskan Glacier Studies; National Geographic Society: Washington, DC, USA, 1914. [Google Scholar]
- Sharp, R.P. The latest major advance of Malaspina Glacier, Alaska. Geogr. Rev. 1958, 48, 16–26. [Google Scholar] [CrossRef]
- Stephens, F.R. A forest ecosystem on a glacier in Alaska. Arctic 1969, 22, 441–444. [Google Scholar] [CrossRef]
- Rampton, V. Neoglacial fluctuations of the Natazhat and Klutlan glaciers, Yukon Territory, Canada. Can. J. Earth Sci. 1970, 7, 1236–1263. [Google Scholar] [CrossRef]
- Post, A.; Streveler, G. The tilted forest: Glaciological-geological implications of vegetated neoglacial ice at Lituya Bay, Alaska. Quat. Res. 1976, 6, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Birks, H.J.B. The present flora and vegetation of the moraines of the Klutlan Glacier, Yukon Territory, Canada: A study in plant succession. Quat. Res 1980, 1, 60–86. [Google Scholar] [CrossRef]
- Benn, D.I.; Evans, D.J.A. Glaciers and Glaciation; Routledge: New York, NY, USA, 2010; 802p. [Google Scholar]
- Veblen, T.T.; Ashton, D.H.; Rubulis, S.; Lorenz, D.; Cortes, M. Nothofagus Stand Development on In-Transit Moraines, Casa Pangue Glacier, Chile. Arct. Alp. Res. 1989, 21, 144–155. [Google Scholar] [CrossRef]
- Miehe, G. Langtang Himal—Flora und Vegetation als Klimazeiger und -zeugen im Himalaya; Dissertationes Botanicae 158; Borntaeger: Berlin, Germany; Stuttgart, Germany, 1990; 529p. [Google Scholar]
- Hambrey, M.; Quincey, D.; Glasser, N.; Reynolds, J.; Richardson, S.; Clemmens, S. Sedimentological, geomorphological and dynamic context of debris-mantled glaciers, Mount Everest (Sagarmatha) region, Nepal. Quat. Sci Rev. 2009, 28, 2361–2389. [Google Scholar] [CrossRef]
- Janke, J.; Bellisario, A.; Ferrando, F. Classification of debris-covered glaciers and rock glaciers in the Andes of central Chile. Geomorphology 2015, 241, 98–121. [Google Scholar] [CrossRef]
- Evatt, G.; Mayer, C.; Mallinson, A.; Abrahams, I.; Heil, M.; Nicholson, L. The secret life of ice sails. J. Glaciol. 2017, 63, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Fowler, A.C.; Mayer, C. The formation of ice sails. Geophys Astro Fluid 2017, 111, 411–428. [Google Scholar] [CrossRef]
- Ravanel, L.; Magnin, F.; Deline, P. Impacts of the 2003 and 2015 summer heat waves on permafrost-affected rock-walls in the Mont Blanc massif. Sci. Total. Environ. 2017, 609, 132–143. [Google Scholar] [CrossRef]
- Mollaret, C.; Hilbich, C.; Pellet, C.; Flores-Orozco, A.; Delaloye, R.; Hauck, C. Mountain permafrost degradation documented through a network of permanent electrical resistivity tomography sites. Cryosphere 2019, 13, 2557–2578. [Google Scholar] [CrossRef] [Green Version]
- Hartmeyer, I.; Delleske, R.; Keuschnig, M.; Krautblatter, M.; Lang, A.; Schrott, L.; Otto, J.-C. Current glacier recession causes significant rockfall increase: The immediate paraglacial response of deglaciating cirque walls. Earth Surf. Dynam. 2020, 8, 729–751. [Google Scholar] [CrossRef]
- Van Woerkom, T.; Steiner, J.F.; Kraaijenbrink, P.D.A.; Miles, E.S.; Immerzeel, W.W. Sediment supply from lateral moraines to a debris-covered glacier in the Himalaya. Earth Surf. Dynam. 2019, 7, 411–427. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.S.; Anderson, R.S. Modeling debris-covered glaciers: Response to steady debris deposition. Cryosphere 2016, 10, 1105. [Google Scholar] [CrossRef] [Green Version]
- Fleischer, F.; Otto, J.-C.; Junker, R.; Hölbling, D. Evolution of debris cover on glaciers of the Eastern Alps, Austria, between 1996 and 2015. Earth Surf. Proc. Land. 2021, 46, 1673–1691. [Google Scholar] [CrossRef]
- Mattson, L.E.; Gardner, J.S.; Young, G.Y. Ablation on Debris Covered Glaciers: An Example from the Rakhiot Glacier, Punjab, Himalaya. In Snow and Glacier Hydrology, Proceedings of the Kathmandu Symposium, Kathmandu, Nepal, 16–21 November 1992; IAHS Publ. no. 218; IAHS: Wallingford, UK, 1993; pp. 289–296. [Google Scholar]
- Boxall, K.; Willis, I.; Giese, A.; Liu, Q. Quantifying Patterns of Supraglacial Debris Thickness and Their Glaciological Controls in High Mountain Asia. Front. Earth Sci. 2021, 9, 504. [Google Scholar] [CrossRef]
- Mayer, C.; Licciulli, C. The concept of steady state, cyclicity and debris unloading of debris-covered glaciers. Front. Earth Sci. 2021, 9, 710276. [Google Scholar] [CrossRef]
- Anderson, L.; Anderson, R. Debris thickness patterns on debris-covered glaciers. Geomorphology 2018, 311, 1–12. [Google Scholar] [CrossRef]
- Moore, P.L. Numerical Simulation of Supraglacial Debris Mobility: Implications for Ablation and Landform Genesis. Front. Earth Sci. 2021, 9, 595. [Google Scholar] [CrossRef]
- Jumpponen, A.; Väre, H.; Mattson, K.; Ohtonen, R.; Trappe, J. Characterization of ‘safe sites’ for pioneers in primary succession on recently deglaciated terrain. J. Ecol. 2003, 87, 98–105. [Google Scholar] [CrossRef]
- Marcante, S.; Erschbamer, B.; Buchner, O.; Neuner, G. Heat tolerance of early developmental stages of glacier foreland species in the growth chamber and in the field. Plant Ecol. 2014, 215, 747–758. [Google Scholar] [CrossRef] [Green Version]
- Fickert, T.; Grüninger, F. High-speed colonization of bare ground—Permanent plot studies on primary succession of plants in recently deglaciated glacier forelands. Land Degrad. Dev. 2018, 29, 2668–2680. [Google Scholar] [CrossRef]
- Eichel, J. Vegetation succession and biogeomorphic interactions in glacier forelands: Landform and Sediment Dynamics in Recently Deglaciated Alpine Landscapes. In Geomorphology of Proglacial Systems; Heckmann, T., Morche, D., Eds.; Springer: Cham, Switzerland, 2019; pp. 327–349. [Google Scholar] [CrossRef]
- Woodward, F.I.; McKee, I.F. Vegetation and climate. Environ. Int. 1991, 17, 535–546. [Google Scholar] [CrossRef]
- Barry, R.G. Mountain Weather and Climate, 3rd ed.; Cambridge University Press: Cambridge, UK, 2008; 532p. [Google Scholar]
- Tackenberg, O.; Stöcklin, J. Wind dispersal of alpine plant species: A comparison with lowland species. J. Veg. Sci. 2008, 19, 109–118. [Google Scholar] [CrossRef]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.A.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.-P.; et al. Flora Indicativa: Ökologische Zeigerwerte und Biologische Kennzeichen zur Flora der Schweiz und der Alpen; Haupt: Bern, Germany, 2010; 376p. [Google Scholar]
- Biek, D. Flora of Mount Rainier National Park; Oregon State University Press: Corvallis, OR, USA, 2000; 506p. [Google Scholar]
- Fischer, M.A.; Adler, W.; Oswald, K. Exkursionsflora für Österreich, Liechtenstein und Südtirol; Land Oberösterreich, Biologiezentrum der OÖ Landesmuseen: Linz, Germany, 2005; 1392p. [Google Scholar]
- Körner, C. Alpine Plant Life, 2nd ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2003; 344p. [Google Scholar]
- Bradshaw, A.D. Understanding the fundamentals of succession. In Primary Succession on Land; Miles, J., Walton, D.W.H., Eds.; Blackwell Scientific Publications: Oxford, UK, 1993; pp. 1–3. [Google Scholar]
- Emmer, A.; Klimeš, J.; Hölbling, D.; Abad, L.; Draebing, D.; Skalák, P.; Štěpánek, P.; Zahradníček, P. Distinct types of landslides in moraines associated with the post-LIA glacier thinning: Observations from the Kinzl Glacier, Huascarán, Peru. Sci. Total Environ. 2020, 739, 139997. [Google Scholar] [CrossRef] [PubMed]
- Gottfried, M.; Pauli, H.; Futschik, A.; Akhalkatsi, M.; Barančok, P.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, M.R.F.; et al. Continent-wide response of mountain vegetation to climate change. Nat. Clim. Change 2012, 2, 111–115. [Google Scholar] [CrossRef]
- Lawrence, D.B. Primary versus secondary succession at Glacier Bay National Monument, southeastern Alaska. In Proceedings of the First Conference on Scientific Research in the National Parks, Orleans, LA, USA, 9–12 November 1976; Transactions and Proceedings Series No. 5.; United States Department of the Interior, National Park Service: Washington, DC, USA, 1979; pp. 213–224. [Google Scholar]
- Fischer, A. Personal communication on subsurface ice on Jamtalferner; Silvretta: Tyrol, Austria, 2021. [Google Scholar]
- Cannone, N.; Diolaiuti, G.; Guglielmin, M.; Smiraglia, C. Accelerating climate change impacts on alpine glacier forefield ecosystems in the European Alps. Ecol. Appl. 2008, 18, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.; Fickert, T.; Schwaizer, G.; Patzelt, G.; Groß, G. Vegetation dynamics in Alpine glacier forelands tackled from space. Sci. Rep. 2019, 9, 13918. [Google Scholar] [CrossRef] [Green Version]
- Gould, S.J. Is uniformitarianism necessary? Am. J. Sci. 1965, 263, 223–228. [Google Scholar] [CrossRef]
- Ives, J.D. Biological refugia and the nunatak hypothesis. In Arctic and Alpine Environments; Ives, J.D., Barry, R.G., Eds.; Methuen: London, UK, 1974; pp. 605–636. [Google Scholar]
- Stehlik, I. Nunataks and peripheral refugia for alpine plants during Quaternary glaciation in the middle parts of the Alps. Bot. Helv. 2000, 110, 25–30. [Google Scholar]
- Brochmann, C.; Gabrielsen, T.; Nordal, I.; Landvik, J.; Elven, R. Glacial Survival or tabula rasa? The History of North Atlantic Biota Revisited. Taxon 2003, 52, 417–450. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalen, M. Refugia revisited: Individualistic response of species in space and time. Proc. R. Soc. B: Biol. Sci. 2009, 277, 661–671. [Google Scholar] [CrossRef] [Green Version]
- Birks, H.J.B.; Willis, K.J. Alpines, trees, and refugia in Europe. Plant Ecol. Divers. 2008, 1, 147–160. [Google Scholar] [CrossRef]
- Gavin, D.; Fitzpatrick, M.; Gugger, P.; Heath, K.; Rodríguez-Sánchez, F.; Dobrowski, S.; Hampe, A.; Hu, F.; Ashcroft, M.; Bartlein, P.; et al. Climate refugia: Joint inference from fossil records, species distribution models and phylogeography. N. Phytol. 2014, 204, 37–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguinot, J.; Ivy-Ochs, S.; Jouvet, G.; Huss, M.; Funk, M.; Preusser, F. Modelling last glacial cycle ice dynamics in the Alps. Cryosphere 2018, 12, 3265–3285. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Körner, C. Topographically controlled thermal habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Schönwiese, C. Klimaänderungen: Daten, Analysen, Prognosen; Springer: Berlin/Heidelberg, Germany, 1995; 224p. [Google Scholar]
- O’Neill, B.; Kriegler, E.; Ebi, K.; Kemp-Benedict, E.; Riahi, K.; Rothman, D.; van Ruijven, B.; Vuuren, D.; Birkmann, J.; Kok, K.; et al. The roads ahead: Narratives for shared socioeconomic pathways describing world futures in the 21st century. Glob. Env. Chang. 2015, 42, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Zale, R.; Huang, Y.-T.; Bigler, C.; Wood, J.R.; Dalén, L.; Wang, X.-R.; Segerström, U.; Klaminder, J. Growth of plants on the Late Weichselian ice-sheet during Greenland interstadial-1? Quat. Sci Rev 2018, 185, 222–229. [Google Scholar] [CrossRef]
- Wright, H.E.; Stefanova, I. Plant trash in the basal sediments of glacial lakes. Acta Palaeobot. 2004, 44, 141–146. [Google Scholar]
- Gentili, R.; Baroni, C.; Caccianiga, M.; Armiraglio, S.; Ghiani, A.; Citterio, S. Potential warm-stage microrefugia for alpine plants: Feedback between geomorphological and biological processes. Ecol. Complex 2015, 21, 87–99. [Google Scholar] [CrossRef]
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Ecol. 2008, 9, 100–107. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.; Ohlemüller, R.; Roy, D.B.; Thomas, C. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Benito Alonso, J.-L.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.; Georges, D.; Rabatel, A.; Randin, C.; Renaud, J.; Delestrade, A.; Zimmermann, N.; Choler, P.; Thuiller, W. Accounting for tree line shift, glacier retreat and primary succession in mountain plant distribution models. Divers. Distrib. 2014, 20, 1379–1391. [Google Scholar] [CrossRef]
- Cannone, N.; Pignatti, S. Ecological responses of plant species and communities to climate warming: Upward shift or range filling processes? Clim. Change 2014, 123, 201–214. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.C. Climate-related range shifts—A global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Steinbauer, M.; Grytnes, J.A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Engler, R.; Randin, C.F.; Thuiller, W.; Dullinger, S.; Zimmermann, N.E.; Araújo, M.B.; Pearman, P.B.; Lay, G.L.; Piedallu, C.; Albert, C.H.; et al. 21st century climate change threatens mountain flora unequally across Europe. Glob. Change Biol. 2011, 17, 2330–2341. [Google Scholar] [CrossRef]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Change 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Körner, C.; Hiltbrunner, E. Why is the Alpine Flora Comparatively Robust against Climatic Warming? Diversity 2021, 13, 383. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Šmilaur, P. CANOCO for Windows Version 4.5; Biometris, Plant Research International: Wageningen, The Netherlands, 2002. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fickert, T.; Friend, D.; Molnia, B.; Grüninger, F.; Richter, M. Vegetation Ecology of Debris-Covered Glaciers (DCGs)—Site Conditions, Vegetation Patterns and Implications for DCGs Serving as Quaternary Cold- and Warm-Stage Plant Refugia. Diversity 2022, 14, 114. https://doi.org/10.3390/d14020114
Fickert T, Friend D, Molnia B, Grüninger F, Richter M. Vegetation Ecology of Debris-Covered Glaciers (DCGs)—Site Conditions, Vegetation Patterns and Implications for DCGs Serving as Quaternary Cold- and Warm-Stage Plant Refugia. Diversity. 2022; 14(2):114. https://doi.org/10.3390/d14020114
Chicago/Turabian StyleFickert, Thomas, Donald Friend, Bruce Molnia, Friederike Grüninger, and Michael Richter. 2022. "Vegetation Ecology of Debris-Covered Glaciers (DCGs)—Site Conditions, Vegetation Patterns and Implications for DCGs Serving as Quaternary Cold- and Warm-Stage Plant Refugia" Diversity 14, no. 2: 114. https://doi.org/10.3390/d14020114
APA StyleFickert, T., Friend, D., Molnia, B., Grüninger, F., & Richter, M. (2022). Vegetation Ecology of Debris-Covered Glaciers (DCGs)—Site Conditions, Vegetation Patterns and Implications for DCGs Serving as Quaternary Cold- and Warm-Stage Plant Refugia. Diversity, 14(2), 114. https://doi.org/10.3390/d14020114