Abstract
The structure and diversity of microbial communities developing in the combined gradient of temperature (44–19 °C), as well as concentration of oxygen (0–10 mg/L) and hydrogen sulfide (33–0.7 mg/L), were studied in the thermal sulfide spring on the coast of Northern Lake Baikal. The predominance of bacteria participating in sulfur and nitrogen cycles and significant changes in the composition of microbial communities were noted at changing physicochemical conditions. Thiovirga sp. (sulfur-oxidizing bacteria, up to 37%) and Azonexus sp. (nitrogen-fixing bacteria, up to 43%) were dominant at high temperatures and concentrations of hydrogen sulfide in two hydrotherms. In addition, a significant contribution of the Rhodocyclaceae family (up to 51%) which is involved in the denitrification processes, and Acetoanaerobium sp. (up to 20%) fixing carbon oxide were found in the spring water. In the stream, mainly oxygenic cyanobacteria (up to 56%) developed at a temperature of 33 °C, in the presence of hydrogen sulfide and oxygen. In addition, sulfur bacteria of the genus Thiothrix (up to 48%) found in epibiotic communities of benthic animals of Lake Baikal were present here. Thiothrix sp. formed massive fouling in the zone of mixing lake and thermal waters with a significant contribution of hydrogen-oxidizing bacteria of the genus Hydrogenophaga (up to 22.5%). As well as chemolitho- and phototrophic bacteria, chemoorganotrophs (phyla Firmicutes, Chloroflexi, Desulfobacterota, Nitrospirota, Fibrobacterota, etc.) have been identified in all communities. The chemical parameters of water in spring and coastal zones indicate a significant change in the composition of thermal waters occurring with the participation of diverse microbial communities that contribute to the assimilation of inorganic components of mineral thermal waters.
1. Introduction
Hot springs are widespread throughout the world, most commonly in the areas of high volcanic and geothermal activity. Their physical and chemical features depend on the particulars of the surrounding territory, geological composition and the hydrochemical processes both on surface and deeper in the crust. The study of hot springs became particularly active after the discovery of previously unknown thermophilic microorganisms inhabiting their waters [1]. A number of papers shows that diversity and composition of a microbial community depends on the spring type [2,3,4,5]; typically, chemolithoautotrophic metabolism is associated with extreme temperatures and pH, and the concentration of inorganic ions.
Yellowstone National Park hot springs (YNP, USA) are among the most well-studied. For example, in three hot alkaline springs of the Heart Lake Geyser Basin (southern YNP) the dominant bacterial community changed from moderately thermophilic and photosynthetic members (Cyanobacteria and Chloroflexi) at 44 °C to a mixed photosynthetic and thermophilic community (Deinococcus-Thermus) at 63 °C, and further to a non-photosynthetic thermophilic community at 75 °C. The archaeal community was predominantly a methanogenic community in the 44 and 63 °C springs, and a thermophilic community in the 75 °C spring [2]. Composition of microbial community in thermal springs with pH < 4 was significantly different from that in alkaline ones: in the former, cyanobacteria were not detected at all, while archaea and bacteria were dominated by those using H2, S0 and Fe2+ as electron donors [3,4].
In Russia, hot springs are present in volcanically active Kuril Islands and Kamchatka, as well as in areas of tectonic activity (such as Buryatia, Altai Krai, Khabarovsk krai and others) [5]. Most well-studied ones are located in the Uzon caldera, where a relatively small area hosts all major types of geothermal activity. The high temperature of these springs leads to the development of photoautotrophic microorganisms utilizing reduced sulfur compounds, hydrogen and other volcanic gases [6]. Archaeal nitrification, carboxydotrophy and iron oxidation are predominant, but methanogenic, sulfur- and sulfate-reducing microorganisms were observed as well [7]. Most springs are dominated by chemoautotrophic phylum Aquificae, which form massive grey-white growths [8]. Sulfide hot spring Termofil’nyi (55–75 °C, pH 6.2) contains white growths of sulfur bacterium Thermotrix thiopara and deposits of native sulfur [9,10]. At temperatures below 60 °C, anaerobic phototrophs and cyanobacteria start appearing as well. With further decrease in temperature and sulfur concentration, accompanied by oxygen concentration increase, cyanobacteria become predominant and form massive algobacterial mats with various other bacteria [7].
Northern Caucasus (Russia) contains multiple thermal springs, most of them moderately warm. Some of these springs were shown to host sulfur-oxidizing and phototrophic bacteria [11]. A unique Nitrospira strain was isolated and shown to perform the complete nitrification process [12]. Despite their diversity and popularity, hydrothermal springs in this area remain poorly studied.
The Baikal rift zone (BRZ, Russia) also contains a number of thermal springs, typically alkaline with temperatures below 50 °C [13]. The vents of all hydrothermal waters with temperatures below 70 °C host massive microbial mats; these vary in their composition, but mostly consist of cyanobacteria [14,15,16]. Many thermal springs of BRZ also contain a significant number of microorganisms involved in the sulfur cycle. Usually, the areas of mixture between sulfur-rich thermal water and colder river or lake water are colonized at temperatures below 40 °C by massive growths of colorless sulfur bacteria from the genus Thiothrix. Where iron appears in significant amounts, these are joined by microaerophilic iron(II)-oxidizers with less abundant iron(III)-reducers, sulphate-reducers and phototrophic and nitrate-reducing iron(II)-oxidizers [17].
Several hot springs are close to the coastline of Lake Baikal (East Siberia, Russia) with a varying distance to the lake itself. The hot spring in Zmeinaya bay is just several meters away from the water line. Several vents are present, two of which are capped and used as hot baths. As thermal waters move to the lake along the outflow, they gradually cool down and form morphologically different microbial communities. Cyanobacterial mats form within the outflow, while white bacterial growths are more common on rocks and plants near the edge of water. Previously, only cyanobacteria, methanotrophs and methylotrophs have been studied in this location [18,19].
The goal of this work was to study the diversity and structure of bacterial communities within a hot spring and the littoral area of Zmeinaya bay under the gradient of oxygen and sulfide concentrations, temperature and other abiotic factors using NGS technologies, as well as to find out whether hot spring microorganisms are present in the littoral area of Lake Baikal.
2. Materials and Methods
2.1. Characteristics of the Study and Sampling Areas
Samples of water and bacterial mats were taken in July 2019 at various sites in Zmeinaya bay: in baths at the vents (B1, B2); along the outflow leading from vent to Lake Baikal (O8, O11); in littoral area 2–3 m away from water edge, where white growths were found on rocks (C3, C5, C6); and vascular plants (C4, C9); and over the black sediment (C7) (Figure 1). Bacterial mats forming along the outflow (O11) were sampled in layer-wise manner, separately isolating white (O11/1), white-green (11/2), purple (O11/3), and blue-green (O11/4) layers. Sample C12 was taken from Lake Baikal 10 m away from the coast, where no bacterial growths were observed.
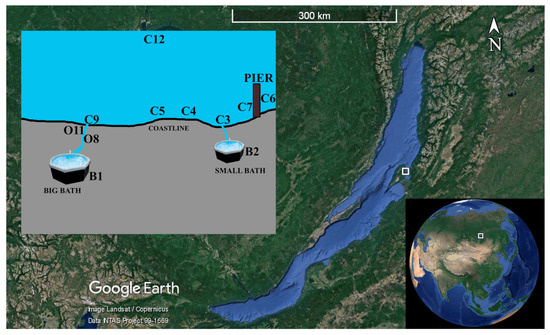
Figure 1.
Schematic map of the study area and sampling sites in Zmeinaya bay (Lake Baikal, Russia). Maps were taken from the Google Earth 7.3.4.8248 Pro software https://www.google.com/intl/ru/earth/versions/#earth-pro (accessed on 12 October 2021).
2.2. Physical and Chemical Parameters of Sites
Chemical analysis was carried out as described previously [20]. Anion concentrations were measured by means of liquid chromatography on a Milichrom-2A chromatograph with a relative error of 5–10%. The samples for cation analysis were stored in polystyrene vials pre-acidified with 50 µL of ultrapure concentrated HNO3 and stored at 4 °C prior to analysis. Cation (Ca2+, Mg2+) concentrations were determined by means of atomic absorption on an AAS-30 Carl Zeiss Jena spectrophotometer, as well as by flame emission methods with a relative error of 3–5% [21]. Concentrations of O2 were measured by Winkler method, hydrogen sulfide—by spectrophotometry [22]. Physicochemical characteristics (pH, Eh, T) were measured with a WTW pH 3310 device at all sampling sites. The main characteristics are presented in Supplementary Table S1.
2.3. DNA Extraction and Sequencing
Water samples from both baths (5 L) were filtered using nitrocellulose filters (25 mm diameter, 0.2 µm pore size; “Millipore”, Darmstadt, Germany) using a diaphragm pump. Filters were placed in a TE buffer (10 mM Tris-HCl, pH 7.4; 1 mM EDTA, pH 8.0) and frozen at −20 °C. DNA from filters and bacterial fouling was extracted according to the modified method of phenol-chloroform extraction [23]. The extracted DNA was kept at −70 °C for further use. For PCR amplification of the bacterial 16S rRNA gene fragments, including the V2–V3 variable region, B_V2F (5′-AGTGGCGGACGGGTGAGTAA) and B_V3R (5′-CCGCGGCTGCTGGCAC) primers were used [24]. Sequencing was performed at the Illumina MiSeq platform in (Collective Instrumental Centre of All-Russian Research Institute of Agricultural Microbiology, Saint-Petersburg). Raw sequences are available in the NCBI SRA archive under the BioProject number PRJNA801752.
2.4. Bioinformatics Analysis
Paired-end sequencing reads were trimmed and filtered in Trimmomatic version 0.39 [25] to remove positions in which more than 25% of the reads had bases with relative quality scores < 20. Then the R1 and R2 sequences were merged into contigs, filtered by size and average quality score (≥25), aligned and checked for chimeras against SILVA 138_1 databases (http://www.arb-silva.de (accessed on 8 Octomber 2021)) using Mothur [26]. Thereafter, gaps were re-moved and sequences were tested for presence of respective forward/reverse primers, allowing two mismatches between forward and reverse primer and sequence. Further rRNA sequence processing was performed according to MiSeq SOP recommendations [27]. The filtered sequences were aligned, clustered into OTUs (operational taxonomic units) at genetic distances of 0.03 and identified taxonomically using the SILVA 138.1 databases. Rarefaction curves were calculated for each sample based on the identified OTUs using the package Mothur. All calculations were performed with HPC-cluster «Akademik V.M. Matrosov» (Irkutsk).
The OTUs were randomly resampled to normalize the reads for each sample, then the richness (ACE, Chao) and diversity estimators (Simpson’s inverse index) were calculated with using web-based tool https://www.microbiomeanalyst.ca (accessed on 8 October 2021). The application http://bioinformatics.psb.ugent.be/webtools/Venn/ (accessed on 23 September 2021) was used to search for overlapping OTUs. The metabolic functions were predicted using the phylogenetic investigation of communities’ reconstruction of unobserved states (PICRUSt2) pipeline (http://picrust.github.io/picrust/ (accessed on 8 Octomber 2021)) using the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. The Mantel test was used to evaluate the correlation of environmental factors and microbial community structure and was calculated using R (9999 permutations).
Before statistical analysis, all data on the concentration of ions in samples, as well as OTU relative abundance, were normalized by unitization with a zero minimum [(x−min)/range]. Comparisons of environmental parameters, alpha and beta diversity of microbial communities using principal coordinate analysis (PCA), boxplots, heat maps and nonmetric multidimensional scaling (NMDS), as well as statistical tests, were performed in R with the vegan, stats, cluster, pheatmap, ggplot2, and ggbiplot packages [28].
3. Results
3.1. Chemical Composition
Multidimensional scaling of chemical and physical environmental values using principal coordinate analysis (PCA) detected three major clusters, excluding sample C12 (Figure 2). This latter sample was taken from Lake Baikal 10 m away from the coast, and its chemical composition is similar to the pelagic waters of the lake [29].
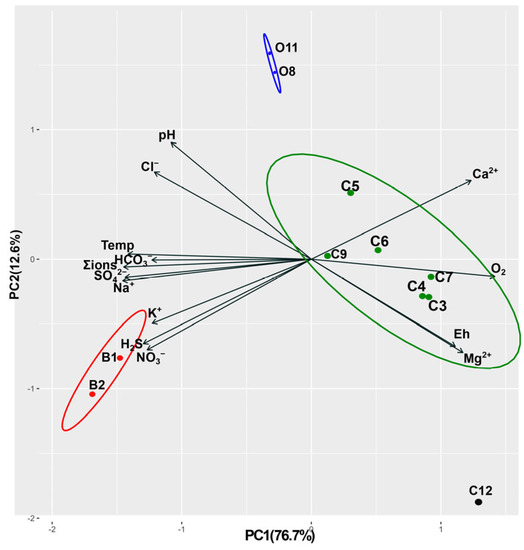
Figure 2.
Principal components analysis (PCA) plot describing the differences of water samples based on the concentration of sum of ions, nutrients, the oxygen concentration (O2), pH, Eh, T.
Cluster I included water samples from the baths (B1 and B2), which featured high temperatures (42.6° and 39 °C), absence of oxygen, high concentration of hydrogen sulfide (33 mg/L) and total ion concentration (up to 426 mg/L), pH 9.1–9.3, Eh = −422 mV.
Cluster II contained samples along the outflow (O8, O11) where green cyanobacterial mats with white and purple inclusions were observed. In this area, water temperature was lower, at 33 °C, hydrogen sulfide concentration was only 1.4 mg/L, and total mineralization was 242 mg/L. Water Eh (−426 mV) was close to bath samples and points to reducing conditions. The water pH was more alkaline than it was at the vent, reaching 9.6.
Finally, cluster III consisted of samples taken in coastal Lake Baikal where hydrothermal waters mix with those of the lake (C3–C9). Water temperature in this area varied between 18° and 24 °C, oxygen concentration increased to 9 mg/L, hydrogen sulfide concentration decreased to 0.7 mg/L and total mineralization to 74–220 mg/L. The water pH was still alkaline at 8.3–8.9, and Eh varied between −153 and −230 mV.
3.2. Alpha Diversity Differences among Sites
Sequencing produced a total of 255,271 reads, which were separated into 922 OTUs from 29 phyla. Rarefaction curves for all samples reached plateau (Figure S1). The total abundance of OTUs observed in 16S sequencing data is closest to real diversity in bath samples (ACE = 89 and sobs = 79 in B2). The number of OTUs observed here is lower than in other libraries, more so in the smaller bath (B2, Chao1 index 89) than in the larger one (B1, Chao index 123) (Figure 3). The distribution is most even in the second cluster and sample C7 (inverse Simpson index 8.2). The highest number of OTUs was observed in sample C7 (415 OTUs).
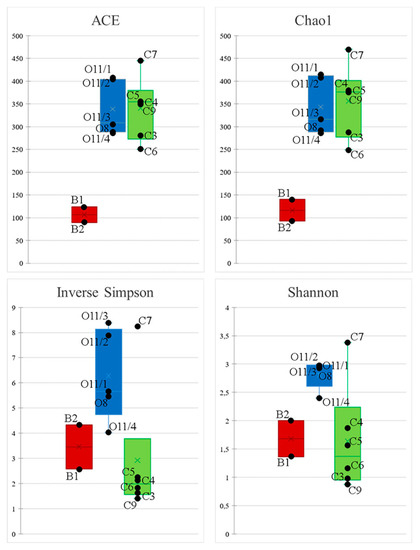
Figure 3.
Box plots show bacterial alpha diversity (ACE and Chao1, Shannon Index and inverse Simpson Index); variation across samples based on non-rarefied data (for a cluster distance of 0.03).
3.3. Beta Diversity Differences between Sites
Clustering samples by their taxonomic composition at phylum or genus rank with unweighted pair group method with arithmetic mean based on Bray-Curtis distance measures generated three clusters identical to those observed from abiotic data. However, sample C7, with its high diversity, is somewhat distant from the rest of cluster III (Figure 4).
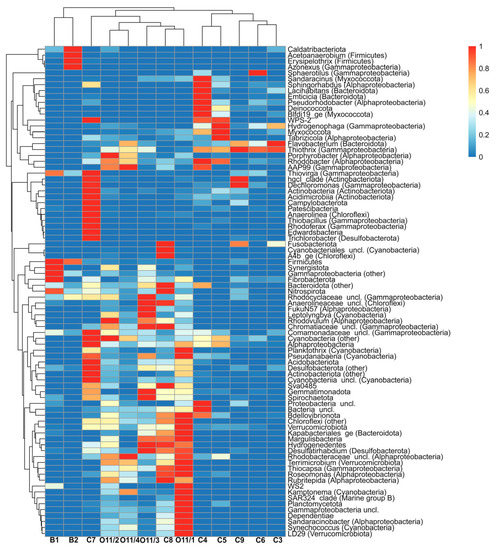
Figure 4.
Heat map demonstrating the distribution of bacterial phylum and dominant genus.
Microbial communities of cluster I (B1 and B2) were dominated by Gammaproteobacteria (91% and 70%, respectively, Figure 5). This class was predominantly represented by uncultured members of the family Rhodocyclaceae (51% and 14%, respectively). This family is metabolically diverse, including photoautotrophic, photoheterotrophic and photoorganotrophic bacteria. The large bath (B1) community was also dominated by sulfur oxidizer genus Thiovirga, family Halothiobacillaceae (37%), which only reached approx. 10% abundance in the small bath (B2). In contrast, the small bath community contained numerous nitrogen-fixing Azonexus, family Rhodocyclaceae (43%), as well as some Firmicutes (now Bacillota [30]) from genera Thermobrachium (3.3%) and Acetoanaerobium (20%). Thermobrachium are proteolytic, while Acetoanaerobium produce acetate from H2 and CO2. Phylum Bacteroidota was the third most abundant in these two baths (2.7 and 11%), but it was mostly represented by unclassified and uncultured species. In minor abundances (under 1%), cluster I samples also contained Chloroflexi, Desulfobacterota, Patescibacteria, Nitrospira and Fibrobacteria. Some sequences of Verrucomicrobia, Synergistota, Spirochaeota and Caldatribacteriota were observed as well.
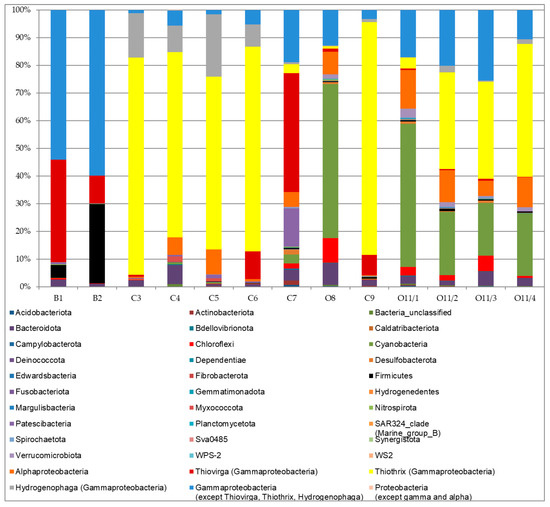
Figure 5.
Taxonomic composition of bacterial communities in different sites of Zmeinaya bay.
Cluster II communities, which develop under lower temperature and hydrogen sulfide concentration, were different: autotrophic cyanobacteria became the dominant phylum (8–56% and 11/1–52%). Out of cyanobacteria, genus Synechococcus was most abundant, reaching 39% in O11/1, and genus Cyanobium was most abundant (present in all samples with abundances up to 0.3%). These communities also included the representatives of genera Leptolyngbya (16% in O11/3), Kamptonema (3.6% in O11/1), Pseudanabaena (up to 1%), Limnothrix and others.
Class Gammaproteobacteria also showed a significant abundance in 16S rRNA libraries for this cluster (between 15% and 62%). Besides colorless sulfur bacteria (0.9–48%), purple sulfur bacteria Allochromatium and Thiocapsa (both up to 5.1%), as well as other minor genera, were observed. Pigmented Alphaproteobacteria that actively participate in sulfur and nitrogen cycles also play a significant role in these communities. In particular, red-colored Rubritepida (up to 1.1%) can oxidize reduced sulfur compounds to sulphate. Furthermore, we detected a remarkable abundance of unclassified purple non-sulfur bacteria (up to 7.8%) and members of genus Rhodovulum (up to 0.6%). Finally, phyla SAR324 (Marine_group_B) and Margulisbacteria were observed in small amounts. The latter are genetically close to cyanobacteria and have not been previously observed in Lake Baikal. They are thought to be involved in the formation of aerobic lifestyle [31].
The final cluster contained samples taken from the area where lake and hot spring water mix, creating an environment rich in both oxygen and hydrogen sulfide. These communities were heavily dominated by phylum Proteobacteria (96–98%), with genus Thiothrix comprising from 62% to 85% of all reads. Another abundant genus was hydrogen-oxidizing Hydrogenophaga (22.5% in C5). This cluster was the only one to feature methylotrophic Methylophilus and Methylotenera (also involved in nitrate reduction), although at abundances below 0.3%. In addition, we observed numerous Alphaproteobacteria, including aerobic anoxygenic phototrophs Tabrizicola (up to 2.4%), phototrophic Rhodobacter (up to 1.1%) and Cereibacter (under 0.1%), and chemoorganotrophic Roseomonas (under 0.1%).
Communities from all three clusters also contained members of phyla Bacteroidota (0.7–6.9%), Nitrospirota (up to 0.4%), and Desulfobacterota (under 1.8%). Clusters II and III also contained members of Fusobacteriota и Gemmatimonadota (both around 0.1%).
3.4. Comparative Community Analysis
Using the Venn diagrams application, we compared bacterial communities from each sample, taking into account OTUs. According to this tool, 49 OTUs were unique for bath samples, and 19 of those were shared between B1 and B2 (Supplementary Table S2). Most of these belonged to genus Thiovirga and one is from genus Desulfomicrobium (Desulfobacteriota). B1 had six unique OTUs, two of which belonged to Brevinema (Spirochaeota) and Tepidomonas (Gammaproteobacteria). B2 had 29 unique OTUs, nine of which belonged to genus Azonexus and five to Acetonanaerobium. OTUs shared between the cluster II communities belonged to the phylum Dependentiae (formerly known as TM6). According to metagenomic data, parasitism is widespread in this lineage [32] and its presence is significantly higher in mineral nitrogen-treated plots [33]. Samples O11/1-O11/4 had 32 unique OTUs, three of which belong to genera Thermoflexibacter (Bacteroidota), Ochrobactrum, and Elioraea (Alphaproteobacteria). Members of phylum Fibrobacterota, capable of degrading plant-based cellulose, were shared between large bath and cluster III. Finally, a total of 13 unique OTUs were detected in cluster III. The highest number of unique OTUs, 78, was observed in community C7. Only one OTU, chemolithoautotrophic sulfur-oxidizing Thiovirga sp., was shared between this sample and all others.
3.5. Predicted Metabolic Functions
The putative metabolic functions of the microbial communities developing in Zmeinaya bay were predicted from 16S rDNA amplicon sequence data using the PICRUSt2 tool (Figure 6). The estimated KEGG orthologues were found to be involved in the main pathways of metabolism. Most of the 60 pathways were associated with the metabolism of energy, carbohydrates, amino acids, lipids and glycan biosynthesis. Xenobiotic biodegradation was found in a small number of studied samples and was linked to the degradation of benzoate. The predicted energy metabolic pathways in the bacterial communities from hot spring in Zmeinaya bay involved oxidative phosphorylation and the metabolism of sulfur, methane and nitrogen. Methane metabolism was found in both baths and in the outflow, and metabolism of sulphur and nitrogen were found in microbial communities of the coastal area. However, in the small bath (B2) community, the nitrogen metabolism pathway was predicted. Carbon fixation in prokaryotes was found in almost every sample, but the carbon fixation pathway in photosynthetic organisms was found only in microbial communities of the small bath (B2). Secondary metabolite pathways for streptomycin, monobactam and prodigiosin were predicted for microbial communities from clusters II and III. Terpenoid backbone biosynthesis was predicted only for both baths (B1, B2). Metabolism of cofactors and vitamins also was found in microbial communities of baths and C7.
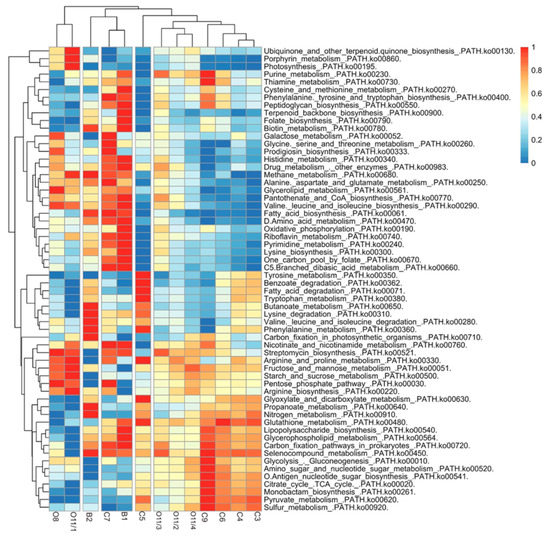
Figure 6.
Heat map of the top 60 abundant putative pathways of microbial communities from the hot spring area in Zmeinaya bay. The pathways are presented according to KEGG.
3.6. Interaction between Microbial Diversity and Abiotic Factors
Analysis of Spearman correlations between relative abundances of major taxa (phyla, families and genera) and various abiotic environmental factors has detected multiple significant correlations, both positive and negative. The negative effect on some taxa was caused by temperature, pH, and concentrations of K+ and hydrogen sulfide (Figure S2). Significant positive correlations were observed between: genera Tabrizicola, Rhodobacter, Sphingorhabdus and nitrate availability; genera Thiocapsa, Desulfatirhabdium and Ca2+; and genera Thiocapsa, Trichlorobacter and Eh. Phylum Deinococcota, genera Sphaerotilus (Gammaproteobacteria) and Lacihabitans (Bacteroidota) correlated negatively with Ca2+ concentration. The latter genus also shows negative correlation with Eh and positive correlation with temperature and hydrogen sulfide concentration.
The Mantel test based on the Bray–Curtis method was used to evaluate the effect of environmental factors on the microbial community structure near the hot spring in Zmeinaya bay. All factors positively correlated with the bacterial community in studied area, among which the correlations of sodium and sulfate-ions were the most significant (Table 1, R = 0.814 and R = 0.786, p < 0.05). The effect of environmental factors on the microbial community structure also is shown with using NMDS (Figure S3).

Table 1.
Mantel analysis of the relationship between environmental factors and microbial community from the area of hot spring in Zmeinaya bay.
4. Discussion
As our study shows, the water of hot spring Zmeinaya bay is the closest in temperature to other thermal springs of the BRZ (for example, Davsha) and Caucasus region [11], but has higher concentration of hydrogen sulfide and more alkaline pH. It is also obvious that neither temperature nor hydrogen sulfide concentration are constant [13,34,35], likely due to changing trajectory and speed of fluid flows. Previously, the effect of fluid flows on the microbial communities was shown in a study of sediment microbes in Posolsk Bank [36]. It is possible that thermal waters of the two Zmeiny vents pass through different subsurface materials, taking with them whatever gases and autochtonous microorganisms are present in these subterranean habitats. This could explain the different composition of dominant taxa in two baths: one (B1) was dominated by sulfur-oxidizing Thiovirga and the other (B2) by nitrogen-fixing Azonexus, although the vents are close to each other and contain chemically similar water. We should also note that all samples contained OTUs of Thiovirga, also observed in a sublacustrine thermal spring of Yellowstone Lake with similar temperature and hydrochemistry [37].
Both the abiotic factors and microbial communities have changed along the way to the lake littoral; the latter varied both in composition and predicted metabolic capacity. As our data show, the community composition is most affected by changing temperature and concentrations of hydrogen sulfide and oxygen. This conclusion is supported both by statistically significant correlation value (Figure S2) and by similar observations in other hot springs of the world [2,6,7,37,38]. Communities in hot and highly mineralized waters (outflow) were dominated by nitrogen fixers, acetogenic bacteria and aerobic sulfur oxidizers perhaps associated with the autotrophic denitrification [39]. These were predominantly mesophilic anaerobic or microaerophilic chemo- or organotrophic bacteria, most of whom utilize sulfur compounds in their metabolism. As the water moves towards the lake, it cools down and somewhat decreases in mineralization, which leads to increasing diversity and changing dominant taxa. Mat-forming cyanobacteria become major players, and a variety of bacteria involved in mat formation may take part in increasing water alkalinity. Major metabolic processes in this area are photosynthesis, nitrogen fixation and sulfide oxidation. Finally, the water and the community it hosts change once again after mixing with cold, oxygen-rich and poorly mineralized waters of Lake Baikal. Now, the communities are dominated by hydrogen-reducing bacteria and consumers of organic matter. In similar ecotopes, the oxidation of sulfur compounds is the main oxygen sink, which is compatible with our data and potentially explains the low abundance of sulfate-reducing and methanotrophic bacteria within the spring.
Microbial communities inhabiting the hot spring in Zmeinaya bay from vent to water edge are similar in their diversity and structure to microbiomes that form in other sulfide springs under temperature gradient. Typically, sulfide-rich waters below 50° are dominated by cyanobacteria and sulfur bacteria. 16S rRNA libraries of the spring we studied contained various cyanobacterial taxa. In particular, we have observed members of the family Leptolyngbyaceae at 44 °C and 33 mg/L sulfide concentration; thus, this family may have wider tolerance than previously thought. At lower temperature and sulfide concentrations (33 °C and 3 mg/L, respectively), genera Synechococcus and Leptolyngbya thrive and form layered deposits, including colorless sulfur bacteria. It is possible that the latter use cyanobacteria-produced oxygen rather than rely on atmospheric supply. The littoral area near the spring, where geothermal and lacustrine waters mix and cool to 19 °C, is dominated by colorless sulfur bacteria Thiothrix and trichome-forming cyanobacterial genera Calothrix, Ancylothrix and Limnothrix. White thread-like growths were previously observed near multiple BRZ hot spring at water temperature below 38 °C: Bauntovski, Kotelnikovsky, Bolsherechensky and others [14,40,41] and sulfide concentration below 41.5 mg/L [42]. It is also important to note that Thiothrix was also detected in epibiotic consortia on oligochaetes from Gorevoi Utes oil seep and ostracodes from Goloustnoe methane seep in Lake Baikal [43,44], where sulfide concentration was below the detection threshold using the same methods as here. These data show that genus Thiothrix has a wide range of acceptable temperatures, oxygen and hydrogen sulfide concentrations; they also do not depend on the light.
All clusters and thus all areas contained bacteria that are capable of disproportionating thiosulfate and elemental sulfur (phylum Nitrospirota), as well as those that reduce sulfates to sulfides and degrade organics to acetate and CO2 (phylum Desulfobacterota). Members of phylum SAR324 found in cluster II may play a role in sulfur cycle, as well. According to metapangenomic data, these bacteria exhibit high metabolic plasticity: they use sulfur-based chemolithoautotrophy in the absence of light and are photoheterotrophic when light is available [45]. Phylum Margulisbacteria has also been detected in cluster II. This phylum is known to be involved in sulfur oxidation and hydrogen cycle; it is also closely related to Cyanobacteria [46].
Since the sulfur cycle is linked to the iron cycle, littoral microbial communities contain a remarkable abundance of heterotrophic iron bacteria Sphaerotilus sp. (Gammaproteobacteria) that accumulate sulfur granules in their cells [47]. Microbial mats formed by simultaneously Thiothrix and Sphaerotilus are usually observed in the activated sludge of treatment facilities and in the surface layers of fresh water heavily contaminated with agricultural waste. Growths of Sphaerotilus natans and Thiothrix sp. were originally discovered in sulfur springs of Northern Caucasus, with the former dominating under low concentration of hydrogen sulfide and disappearing under higher ones [48]. Another sulfur-oxidizing bacterium in our dataset is Sideroxydans sp., which is detected in sample C7.
Besides primary producers, the microbial communities of Zmeinaya bay contain bacteria involved in various destruction processes. The littoral area hosts the members of family Saprospiraceae (phylum Bacteroidota) capable of algicidal activity against cyanobacteria, lytic destruction of some bacterial cells, and degradation of complex macromolecules. Clusters II and III contain predatory bacteria Bdellovibrio (Bdellovibrionota) that consume external biopolymers such as proteins and nucleic acids. Members of phylum Patescibacteria are present as well, and the genomes of this group suggest the capacity for degrading organic matter [49]. Previously, Patescibacteria (CPR) were only present significantly in the deep aphotic layer of Lake Baikal [50]. Almost all samples also contain a small abundance (up to 0.04%) of cellulose destructor phylum Fibrobacterota, which was never before documented in the lake itself, but present in the microbial community of BRZ hot spring Kuchiger [51].
The baths in Zmeinaya bay are a popular tourist destination, particularly during the summer. Thus, it is not surprising that some opportunistic pathogens are present in studied microbial communities. In particular, the smaller bath contains genus Erysipelothrix (5.5%, phylum Firmicutes), which includes skin pathogens. This genus is also present near the larger bath (0.01%) and along the coastline (up to 0.14%). It is interesting that the majority of potential pathogens have been detected in cluster III, presumably due to the more tolerable temperature. Microbial communities of this cluster contain predatory phylum Myxococcota at 0.1–19% abundance. Members of this phylum take part in sulphate and nitrite reduction, and some of them are pathogenic for humans. The same communities also include Roseomonas sp. (Alphaproteobacteria), some strains of which are opportunistic pathogens. Other members of this genus have also been found in Baikal sediment cores [52] and epilithic biofilms on quartz [53].
Thus, the waters of hot spring in Zmeinaya bay contain a peculiar microbial community whose metabolic potential is based on chemolithoautotrophs utilizing sulfur and nitrogen compounds as an energy source. On the way from spring issue to Lake Baikal, the community composition repeatedly shifts, from chemoautotroph-dominated baths to photoautotrophs in the outflow and chemoheterotrophs near outflow mouth. In turn, the bacterial community affects the chemical composition of spring water. This effect of the microbiome on water chemistry has also been described in the lake itself, although it has a completely different microbial community with different metabolic capacities. It is obvious that some bacteria are moved from the spring to the lake, but their habitat is localized and does not extend far beyond outflow mouth, presumably due to lower temperature and oligotrophic conditions.
5. Conclusions
It is obvious that the mineralized water of the geothermal spring in Zmeinaya bay has a local effect on the ecosystem of Lake Baikal, which is quickly neutralized by the dissolution of geothermal water in the lake and consumption of N2, CO2, H2, H2S and other reduced sulfur compounds by microbial communities. These bacteria serve as a sort of filter, preventing toxic compounds from reaching the wider Baikal ecosystem. Gradients of temperature, sulfide concentration and oxygen concentration lead to the formation of multiple distinct bacterial communities in a small patch of lake coast. We should also note the high number of sequences with low homology to studied species, suggesting high potential for the discovery of novel bacteria.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15020298/s1, Figure S1: Rarefaction curves plotting the number observed OTUs as a function of the number of sequences defined at genetic distance level of 0.03; Figure S2: A heat map (with Spearman’s rank correlation coefficient) showing correlation between dominant OTUs (97% identity) with abiotic factors. Colors indicate the r values of Spearman’s rank correlation coefficient. The influence of a factor was assumed to be reliable if the p-value of the Spearman statistics was <0.05; Figure S3: Non-metric multidimensional scaling (NMDS) based on Bray-Curtis dissimilarities shows relationships between the relative bacterial abundance of each sample and environmental parameters; Table S1: Physical and chemical parameters of water in Zmeinaya bay (concentration of ion-mg/L). Table S2: Total and unique OTUs in samples.
Author Contributions
Conceptualization, T.Z. and S.C.; methodology, A.Z.; software, S.B.; validation, S.B.; formal analysis, S.C.; investigation, I.E., T.P.; writing—original draft preparation, S.C.; writing—review and editing, T.Z.; visualization, S.B.; supervision, T.Z.; project administration, T.Z. All authors have read and agreed to the published version of the manuscript.
Funding
Expeditionary and chemical research were supported by the State Project of the Ministry of Science and Higher Education of the Russian Federation No. 0279-2021-0006 (121032300223-1); molecular biological analysis was supported by the Russian Science Foundation Grant No. 22-14-00084.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All raw sequences are available in the NCBI SRA archive under the BioProject number PRJNA801752.
Acknowledgments
This study was carried out using the equipment of the Core Centrum “Genomic Technologies, Proteomics and Cell Biology” in ARRIAM. The authors would like to thank Irkutsk Supercomputer Center SB RAS for providing access to HPC-cluster “Akademik V.M. Matrosov”. S.C. is grateful to O. Kotsar for help with Figure 1.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brock, T.D.; Freeze, H. Thermus aquaticus, a nonsporulating extreme thermophile. J. Bacteriol. 1969, 98, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Bowen De Leon, K.; Gerlach, R.; Peyton, B.M.; Fields, M.W. Archaeal and bacterial communities in three alkaline hot springs in Heart Lake Geyser Basin, Yellowstone National Park. Front. Microbiol. 2013, 4, 330. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.M.; Beard, B.L.; Beukes, N.J.; Klein, C.; O’Leary, J.M. Ancient geochemical cycling in the Earth as inferred from Fe isotope studies of banded iron formations from the Transvaal Craton. Contrib. Mineral. Petrol. 2003, 144, 523–547. [Google Scholar] [CrossRef]
- Beam, J.P.; Jay, Z.J.; Schmid, M.C.; Rusch, D.B.; Romine, M.F.; Jennings, R.M.; Kozubal, M.A.; Tringe, S.G.; Wagner, M.; Inskeep, W.P. Ecophysiology of an uncultivated lineage of Aigarchaeota from an oxic, hot spring filamentous ‘streamer’ community. ISME J. 2016, 10, 210–224. [Google Scholar] [CrossRef]
- Kochetkova, T.V.; Podosokorskaya, O.A.; Elcheninov, A.G.; Kublanov, I.V. Diversity of thermophilic prokaryotes inhabiting Russian natural hot springs. Microbiology 2022, 91, 3–31. (In Russian) [Google Scholar] [CrossRef]
- Zavarzin, G.A.; Karpov, G.A.; Gorlenko, V.M.; Golovacheva, R.S.; Gerasimenko, L.M.; Bonch-Osmolovskaya, E.A.; Orleanskii, V.K. Kaldernye Mikroorganizmy (Caldera Microorganisms); Nauka: Moscow, Russia, 1989; p. 120. (In Russian) [Google Scholar]
- Gorlenko, V.M.; Starynin, D.A.; Bonch-Osmolovskaya, E.A.; Kachalkin, V.I. Production processes in microbial cenoses of the Thermofil’nyi Hot Spring. Microbiology 1987, 56, 872–878. (In Russian) [Google Scholar]
- Reigstad, L.J.; Jorgensen, S.L.; Schleper, C. Diversity and abundance of Korarchaeota in terrestrial hot springs of Iceland and Kamchatka. ISME J. 2010, 4, 346–356. [Google Scholar] [CrossRef]
- Bonch-Osmolovskaya, E.A.; Karpov, G.A. Microbial methane formation in Uzon Caldera hydrothermal vents. Microbiology 1986, 56, 516–518. (In Russian) [Google Scholar]
- Lazareva, E.V.; Anisimova, N.S.; Bryanskaya, A.V.; Ogorodnikova, O.L.; Zhmodik, S.M. Features of Mineral Formation in Microbial Communities Developing along the Outflow of Termofil’nyi Vent (Uzon caldera, Kamchatka). Tr. Kronotsk. Gos. Biosfer. Zapovednika (Proc. Kronotsky State Biosphere Reserve); Mosolov, V.I., Ed.; Kamchatpress: Petropavlovsk-Kamchatsky, Russia, 2012; pp. 143–156. (In Russian) [Google Scholar]
- Chernousova, E.Y. Biodiversity of Filamentous Sulfur Bacteria in Sulfide Ecosystems of the North Caucasus Region: Ecological and Molecular Genetic Aspects. Ph.D. Thesis, Voronezh State University, Voronezh, Russia, 2011. [Google Scholar]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef]
- Borisenko, I.M.; Zamana, L.V. Mineral’nye Vody Buryatskoi ASSR (Mineral Waters of Buryat ASSR); Buryat. Izd.: Ulan-Ude, Russia, 1978; p. 162. [Google Scholar]
- Kompantseva, E.I.; Gorlenko, V.M. Phototrophic communities of some thermal springs of Lake Baikal. Microbiology 1988, 57, 841–846. (In Russian) [Google Scholar]
- Namsaraev, Z.B.; Gorlenko, V.M.; Namsaraev, B.B.; Buriukhaev, S.P.; Iurkov, V.V. The structure and biogeochemical activity of the phototrophic communities from the Bol’sherechenskii alkaline hot spring. Microbiology 2003, 72, 228–238. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Babasanova, O.B.; Barkhutova, D.D.; Namsaraev, Z.B.; Kalashnikova, O.M. Aerobic organothrophic bacteria in hot springs of Baikal region. Bulletin of the Buryat State University. Biol. Geogr. 2007, 3, 112–118. (In Russian) [Google Scholar]
- Koeksoy, E.; Halama, M.; Hagemann, N.; Weigold, P.R.; Laufer, K.; Kleindienst, S.; Byrne, J.M.; Sundman, A.; Hanselmann, K.; Halevy, I.; et al. A case study for late Archean and Proterozoic biogeochemical iron- and sulphur cycling in a modern habitat-the Arvadi Spring. Geobiology 2017, 16, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Sorokovikova, E.G.; Belykh, O.I.; Likhoshvai, E.V. Cyanobacteria of Thermal Springs of the Baikal Rift Zone: Species Composition, Cell Ultrastructure, Silicon Biomineralization; Modern Paleontology, Classical and Modern Methods; Borissiak Paleontological Institute of the Russian Academy of Sciences: Moscow, Russia, 2007; pp. 11–18. [Google Scholar]
- Zelenkina, T.S.; Dagurova, O.P.; Namsaraev, B.B.; Eshinimayev, B.T.; Suzina, N.E.; Trotsenko, Y.A. Aerobic methanotrophs from the coastal thermal springs of Lake Baikal. Microbiology 2009, 78, 492–497. (In Russian) [Google Scholar] [CrossRef]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer: New York, NY, USA, 1991; p. 391. [Google Scholar]
- Baram, G.I.; Vereshchagin, A.L.; Golobokova, L.P. Microcolumn high-performance liquid chromatography with UV detection for the determination of anions in environmental materials. J. Anal. Chem. 1999, 54, 962–965. (In Russian) [Google Scholar]
- Cline, J.D. Spectrophotometric determination of hydrogen sulphide in natural waters. Limnol. Oceanogr. 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Marmur, J. A procedure for the isolation of deoxyribonucleic acid from microorganisms. J. Mol. Biol. 1961, 3, 208–218. [Google Scholar] [CrossRef]
- Sahm, K.; John, P.; Nacke, H.; Wemheuer, B.; Grote, R.; Daniel, R.; Antranikian, G. High abundance of heterotrophic prokaryotes in hydrothermal springs of the Azores as revealed by a network of 16S rRNA gene-based methods. Extremophiles 2013, 17, 649–662. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. 2022. Available online: https://www.R-project.org/ (accessed on 4 October 2022).
- Falkner, K.K.; Measures, C.I.; Herbelin, S.E.; Edmond, J.M.; Weiss, R.F. The major and minor element geochemistry of Lake Baikal. Limnol. Oceanogr. 1991, 36, 413–423. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 10. [Google Scholar] [CrossRef] [PubMed]
- Matheus Carnevali, P.B.; Schulz, F.; Castelle, C.J.; Kantor, R.S.; Shih, P.M.; Sharon, I.; Santini, J.M.; Olm, M.R.; Amano, Y.; Thomas, B.C.; et al. Hydrogen-based metabolism as an ancestral trait in lineages sibling to the Cyanobacteria. Nat. Commun. 2019, 10, 463. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Sekiguchi, Y.; Parks, D.H.; Hugenholtz, P.B. Comparative genomics of candidate phylum TM6 suggests that parasitism is widespread and ancestral in this lineage. Mol. Biol. Evol. 2016, 33, 915–927. [Google Scholar] [CrossRef]
- Suproniene, S.; Doyeni, M.O.; Viti, C.; Tilvikiene, V.; Pini, F. Characterization of the Soil Prokaryotic community with respect to time and fertilization with animal waste–based digestate in a humid continental climate. Front. Environ. Sci. 2022, 10, 12. [Google Scholar] [CrossRef]
- Barhutova, D.D. Effect of Environmental Parameters on Bacterial Diversity and Functional Activity of Bacteria-Destructors in Sulfide-Rich Springs of the Baikal Region. Ph.D. Thesis, Banzarov Buryat State University, Ulan-Ude, Russia, 2000. [Google Scholar]
- Plyusnin, A.M.; Zamana, L.V.; Shvartsev, S.L.; Tokarenko, O.G.; Chernyavskii, M.K. Hydrogeochemical peculiarities of the composition of nitric thermal waters in the Baikal Rift Zone. Geol. Geophys. 2013, 54, 647–664. [Google Scholar] [CrossRef]
- Chernitsyna, S.M.; Mamaeva, E.V.; Lomakina, A.V.; Pogodaeva, T.V.; Galach’yants, Y.P.; Bukin, S.V.; Khlystov, O.M.; Zemskaya, T.I.; Pimenov, N.V. Phylogenetic diversity of microbial communities of the Posolsk Bank bottom sediments, Lake Baikal. Microbiology 2016, 85, 672–680. [Google Scholar] [CrossRef]
- Yang, T.; Lyons, S.; Aguilar, C.; Cuhel, R.; Teske, A. Microbial communities and chemosynthesis in Yellowstone Lake sublacustrine hydrothermal vent waters. Front. Microbiol. 2011, 2, 130. [Google Scholar] [CrossRef]
- Martínez, A.; Pibernat, I.; Figueras, J.; García-Gil, J. Structure and composition of freshwater microbial mats from a sulfur spring (“Font Pudosa”, NE Spain). Microbiologia 1997, 13, 45–56. [Google Scholar]
- Lan, L.; Zhao, J.; Wang, S.; Li, X.; Qiu, L.; Liu, S.; Bin Nasry, A.A. NO and N2O accumulation during nitrite-based sulfide-oxidizing autotrophic denitrification. Bioresour. Technol. Rep. 2019, 7, 100190. [Google Scholar] [CrossRef]
- Potapova, Z.M. The Species Composition of Cyanobacteria and Their Ecophysiology in Nitrogen-Rich Thermal Springs of Northern Transbaikal Region. Ph.D. Thesis, Banzarov Buryat State University, Ulan-Ude, Russia, 2010. [Google Scholar]
- Yurkov, V.V.; Gorlenko, V.M.; Mityushina, L.L.; Starynin, D.A. The Effect of Limiting Factors on the Structure of Phototrophic Communities in the Bol’sherechenskie Thermal Springs. Microbiology 1991, 60, 129–138. (In Russian) [Google Scholar]
- Namsaraev, B.B.; Barkhutova, D.D.; Danilova, E.V.; Bryanskaya, A.V.; Buryukhaev, S.P.; Garmaev, E.Z.; Gorlenko, V.M.; Dagurova, O.P.; Dambaev, V.B.; Zaitseva, S.V.; et al. The Geochemical Activity of Microorganisms of the Thermal Springs in the Baikal Rift Zone; Weinstein, M.B., Ed.; Academic Publishing House “Geo”: Novosibirsk, Russia, 2011; p. 302. [Google Scholar]
- Chernitsyna, S.M.; Khalzov, I.A.; Sitnikova, T.Y.; Naumova, T.V.; Khabuev, A.V.; Zemskaya, T.I. Microbial Communities Associated with Bentic Invertebrates of Lake Baikal. Curr. Microbiol. 2021, 78, 3020–3031. [Google Scholar] [CrossRef] [PubMed]
- Khalzov, I.A.; Bukin, S.V.; Zakharenko, A.S.; Chernitsyna, S.M.; Galachyants, Y.P.; Sitnikova, T.Y.; Zemskaya, T.I. Microbial communities associated with the ostracods Candona sp. inhabiting the area of the methane seep Goloustnoye (Lake Baikal). Symbiosis 2021, 85, 163–174. [Google Scholar] [CrossRef]
- Boeuf, D.; Eppley, J.M.; Mende, D.R.; Malmstrom, R.R.; Woyke, T.; DeLong, E.F. Metapangenomics reveals depth-dependent shifts in metabolic potential for the ubiquitous marine bacterial SAR324 lineage. Microbiome 2021, 9, 18. [Google Scholar] [CrossRef]
- Anantharaman, K.; Brown, C.T.; Hug, L.A.; Sharon, I.; Castelle, C.J.; Probst, A.J.; Thomas, B.C.; Singh, A.; Wilkins, M.J.; Karaoz, U.; et al. Thousands of microbial genomes shed light on interconnected biogeochemical processes in an aquifer system. Nat. Commun. 2016, 7, 11. [Google Scholar] [CrossRef]
- Van Veen, W.L.; Mulder, E.G.; Deinema, M.H. The Sphaerotilus−Leptothrix group of bacteria. Microbiol. Rev. 1978, 42, 162–329. [Google Scholar] [CrossRef]
- Chernousova, E.Y.; Gridneva, E.V.; Grabovich, M.Y.; Akimov, V.N.; Dubinina, G.A. Phylogenetic in situ/ex situ analysis of a sulfur mat microbial community from a thermal sulfide spring in the North Caucasus. Microbiology 2008, 77, 219–223. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, C.; Li, W.; Weng, S.; Song, W.; Li, J.; Wang, Y. Functional diversity of microbial communities in inactive seafloor sulfide deposits. FEMS Microbiol. Ecol. 2021, 97, 14. [Google Scholar] [CrossRef]
- Cabello-Yeves, P.J.; Zemskaya, T.I.; Zakharenko, A.S.; Sakirko, M.V.; Ivanov, V.G.; Ghai, R.; Rodriguez-Valera, F. Microbiome of the deep Lake Baikal, a unique oxic bathypelagic habitat. Limnol. Oceanogr. 2020, 65, 1471–1488. [Google Scholar] [CrossRef]
- Yuriev, D.A.; Zaitseva, S.V.; Zhdanova, G.O.; Tolstoy, M.Y.; Barkhutova, D.D.; Vyatchina, O.F.; Konovalova, E.Y.; Stom, D.I. Microbial mat of the thermal springs Kuchiger Republic of Buryatia: Species composition, biochemical properties and electrogenic activity in biofuel cell. IOP Conf. Ser. Earth Environ. Sci. 2018, 121, 022012. [Google Scholar] [CrossRef]
- Andreeva, I.S.; Pechurkina, N.I.; Morozova, O.V.; Ryabchikova, E.I.; Belikov, S.I.; Puchkova, L.I.; Emel’yanova, E.K.; Torok, T.; Repin, V.E. The new eubacterium Roseomonas baikalica sp. nov. isolated from core samples collected by deep-hole drilling of the bottom of Lake Baikal. Microbiology 2007, 76, 487–493. [Google Scholar] [CrossRef]
- Sukhanova, E.V.; Shtykova, Y.R.; Suslova, M.Y.; Pestunova, O.S.; Kostornova, T.Y.; Khanaev, I.V.; Zimens, E.A.; Podlesnaya, G.V.; Parfenova, V.V. Diversity and Physiological and Biochemical Properties of Heterotrophic Bacteria Isolated from Lake Baikal Epilithic Biofilms. Microbiology 2019, 88, 345–357. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).