
Habitat Selection Differences of Two Sympatric Large Carnivores in the Southwestern Mountains of China


 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.2.1. Camera Trapping Surveys
2.2.2. Environmental Variables
2.3. Data Analysis
2.3.1. Ensemble Model
2.3.2. Assessment of Habitat Suitability
2.3.3. Assessment of Suitable Habitat Landscapes
2.3.4. Spatial Interactions
3. Results
3.1. Ensemble Model Accuracy Assessment
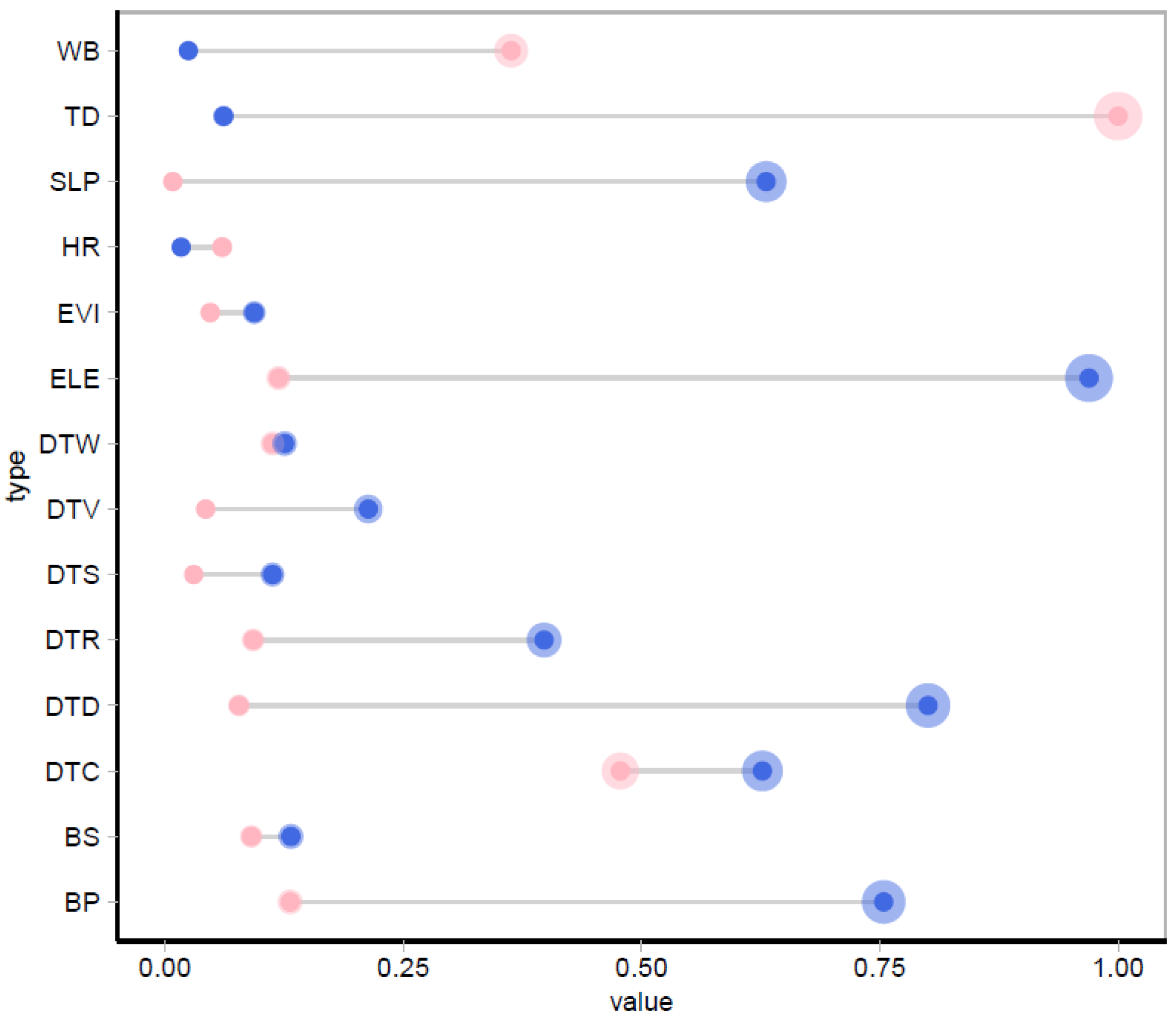
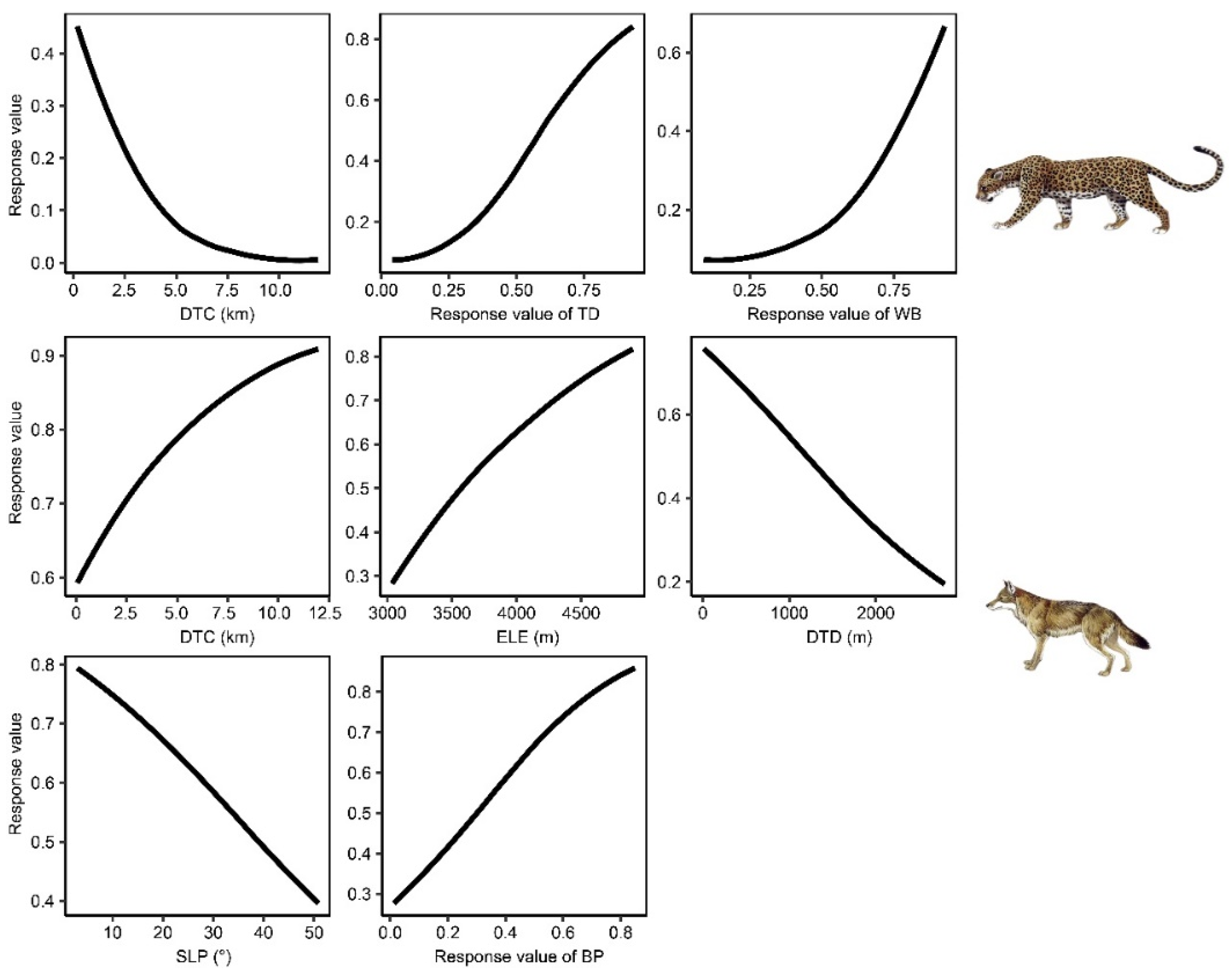
3.2. Environmental Variable Importance Analysis
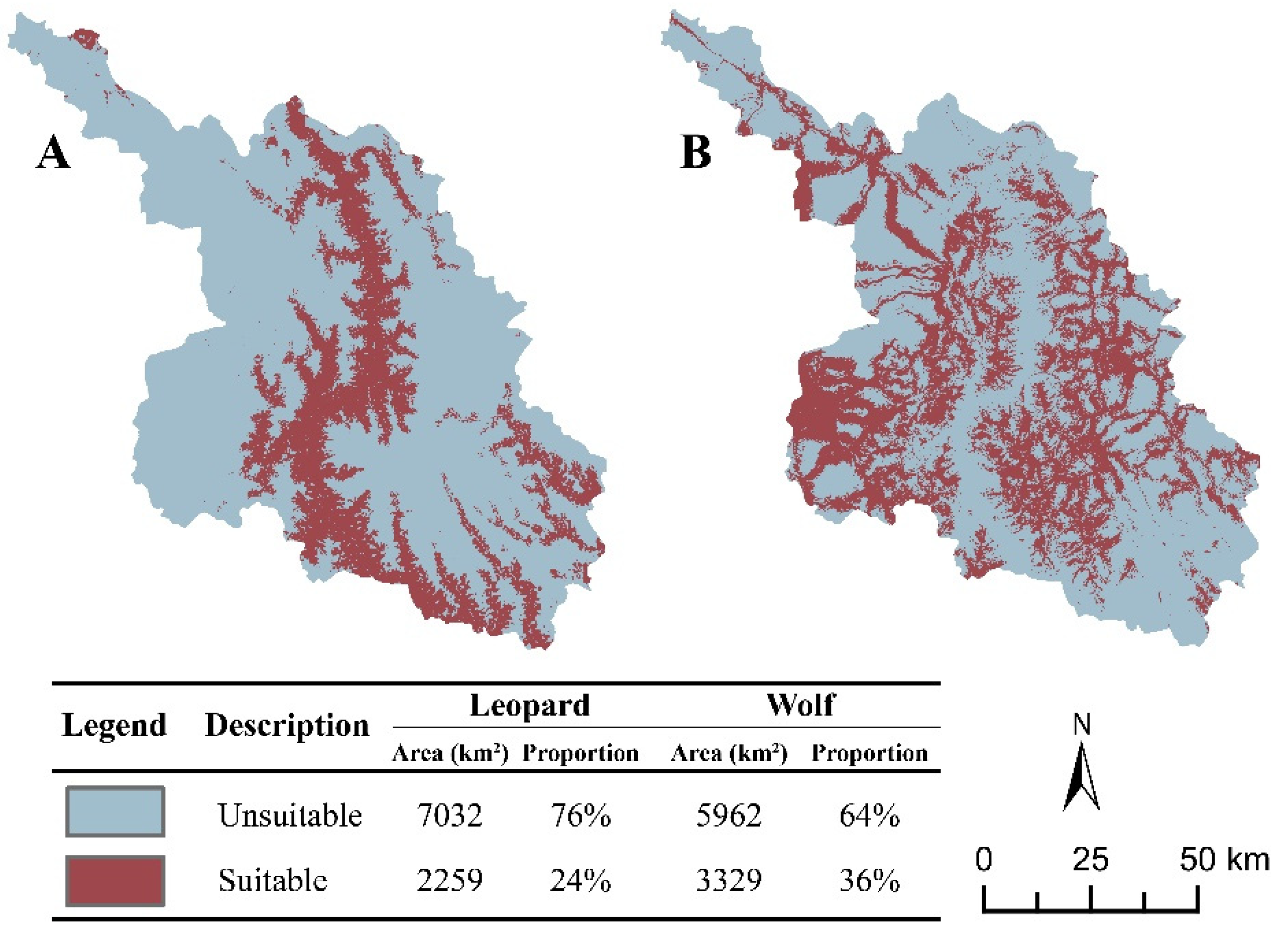
3.3. Habitat Suitability Evaluation
3.4. Habitat Suitability Landscape Evaluation
3.5. Spatial Interactions
4. Discussion
4.1. Divergent Suitable Habitat Selection
4.2. Habitat Fragmentation
4.3. Conclusions and Conservation Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P.; et al. Status and Ecological Effects of the World’s Largest Carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Duo, L.; Li, S.; Wang, T. Competition and coexistence among terrestrial mammalian carnivores. Biodivers. Sci. 2021, 29, 81–97. [Google Scholar] [CrossRef]
- Marske, K.A.; Lanier, H.C.; Siler, C.D.; Rowe, A.H.; Stein, L.R. Integrating Biogeography and Behavioral Ecology to Rapidly Address Biodiversity Loss. Proc. Natl. Acad. Sci. USA 2023, 120, e2110866120. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Din, J.U.; Bosso, L.; Hameed, S.; Kabir, M.; Younas, M.; Nawaz, M.A. Expanding or Shrinking? Range Shifts in Wild Ungulates under Climate Change in Pamir-Karakoram Mountains, Pakistan. PLoS ONE 2021, 16, e0260031. [Google Scholar] [CrossRef]
- Hardin, G. The Competitive Exclusion Principle: An Idea That Took a Century to Be Born Has Implications in Ecology, Economics, and Genetics. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef]
- Odden, M.; Wegge, P.; Fredriksen, T. Do Tigers Displace Leopards? If so Why? Ecol. Res. 2010, 25, 875–881. [Google Scholar] [CrossRef]
- Widodo, F.A.; Imron, M.A.; Sunarto, S.; Giordano, A.J. Carnivores and Their Prey in Sumatra: Occupancy and Activity in Human-Dominated Forests. PLoS ONE 2022, 17, e0265440. [Google Scholar] [CrossRef]
- Ahmad, F.; Nawaz, M.A.; Salim, M.; Rehan, M.; Farhadinia, M.; Bosso, L.; Kabir, M. Patterns of Spatial Distribution, Diel Activity and Human-Bear Conflict of Ursus thibetanus in the Hindu Kush Mountains, Pakistan. Glob. Ecol. Conserv. 2022, 37, e02145. [Google Scholar] [CrossRef]
- Fennell, J.G.; Talas, L.; Baddeley, R.J.; Cuthill, I.C.; Scott-Samuel, N.E. Optimizing Colour for Camouflage and Visibility Using Deep Learning: The Effects of the Environment and the Observer’s Visual System. J. R. Soc. Interface 2019, 16, 20190183. [Google Scholar] [CrossRef]
- Shahsavarzadeh, R.; Hemami, M.-R.; Farhadinia, M.S.; Fakheran, S.; Ahmadi, M. Spatially Heterogeneous Habitat Use across Distinct Biogeographic Regions in a Wide-Ranging Predator, the Persian Leopard. Biodivers. Conserv. 2023, 32, 2037–2053. [Google Scholar] [CrossRef]
- Ngoprasert, D.; Lynam, A.J.; Gale, G.A. Human Disturbance Affects Habitat Use and Behaviour of Asiatic Leopard Panthera pardus in Kaeng Krachan National Park, Thailand. Oryx 2007, 41, 343–351. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- De Lima Filho, J.A.; Vieira, R.J.A.G.; De Souza, C.A.M.; Ferreira, F.F.; De Oliveira, V.M. Effects of Habitat Fragmentation on Biodiversity Patterns of Ecosystems with Resource Competition. Phys. A Stat. Mech. Its Appl. 2021, 564, 125497. [Google Scholar] [CrossRef]
- Ahmadi, M.; Balouchi, B.N.; Jowkar, H.; Hemami, M.-R.; Fadakar, D.; Malakouti-Khah, S.; Ostrowski, S. Combining Landscape Suitability and Habitat Connectivity to Conserve the Last Surviving Population of Cheetah in Asia. Divers. Distrib. 2017, 23, 592–603. [Google Scholar] [CrossRef]
- Liu, X. Overlapping Dietary and Food Ecological Niches of Three Species of Canids in Eastern Inner Mongolia. Master’s Thesis, Qufu Normal University, Qufu, China, 2009. [Google Scholar]
- Zafar-ul Islam, M.; Volmer, R.; Al Boug, A.; Shehri, A.A.; Gavashelishvili, A. Modelling the Effect of Competition for Prey and Poaching on the Population of the Arabian Leopard, Panthera pardus nimr, in Saudi Arabia (Mammalia: Felidae). Zool. Middle East 2020, 66, 95–106. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, B.; Gu, X.; Song, D.; Yang, B. Camera Trapping Reveals Area of Conservation Significance for Large and Medium-Sized Mammals on the Eastern Tibetan Plateau. Oryx 2022, 56, 137–144. [Google Scholar] [CrossRef]
- Qiao, J. Distribution and Behavior of Leopard (Panthera pardus), Snow Leopard (Panthera unica) and Gray Wolf (Canis lupus) in Gongga Mountain, Sichuan Province. Master’s Thesis, Sichuan Agricultural University, Chengdu, China, 2022. [Google Scholar]
- Kamino, L.H.Y.; Stehmann, J.R.; Amaral, S.; De Marco, P.; Rangel, T.F.; de Siqueira, M.F.; De Giovanni, R.; Hortal, J. Challenges and Perspectives for Species Distribution Modelling in the Neotropics. Biol. Lett. 2011, 8, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gao, H.; Teng, L.; Su, Y.; Wang, X.; Kong, Y. Habitat suitability assessment of blue sheep in Helan Mountain based on MAXENT modeling. Acta Ecol. Sin. 2013, 33, 7243–7249. [Google Scholar] [CrossRef]
- Qi, Z.; Xu, W.; Xiong, X.; Ou, Y.; Zheng, H.; Gan, D. Assessment of potential habitat for Ursus thibetanus in the Qinling Mountains. Biodivers. Sci. 2011, 19, 343–352. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, Q.; Wang, B.; Gong, X.; Duan, C.; Zhang, Y. The habitat suitability evaluation of Salweenia bouffordiana based on MaxEnt model. Acta Ecol. Sin. 2020, 40, 6077–6085. [Google Scholar] [CrossRef]
- Ancillotto, L.; Mori, E.; Bosso, L.; Agnelli, P.; Russo, D. The Balkan Long-Eared Bat (Plecotus kolombatovici) Occurs in Italy—First Confirmed Record and Potential Distribution. Mamm. Biol. 2019, 96, 61–67. [Google Scholar] [CrossRef]
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A Comparison between Ensemble and MaxEnt Species Distribution Modelling Approaches for Conservation: A Case Study with Egyptian Medicinal Plants. Ecol. Inform. 2020, 60, 101150. [Google Scholar] [CrossRef]
- Brito, J.C.; Crespo, E.G.; Paulo, O.S. Modelling Wildlife Distributions: Logistic Multiple Regression vs Overlap Analysis. Ecography 1999, 22, 251–260. [Google Scholar] [CrossRef]
- Qing, J.; Yang, Z.; He, K.; Zhang, Z.; Gu, X.; Yang, X.; Zhang, W.; Yang, B.; Qi, D.; Dai, Q. The Minimum Area Requirements (MAR) for Giant Panda: An Empirical Study. Sci. Rep. 2016, 6, 37715. [Google Scholar] [CrossRef]
- Swets, J. Measuring the Accuracy of Diagnostic Systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the Accuracy of Species Distribution Models: Prevalence, Kappa and the True Skill Statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Richmond, O.M.W.; Hines, J.E.; Beissinger, S.R. Two-Species Occupancy Models: A New Parameterization Applied to Co-Occurrence of Secretive Rails. Ecol. Appl. 2010, 20, 2036–2046. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhao, X.; Han, B.; Wang, T.; Mou, P.; Ge, J.; Feng, L. Spatiotemporal Patterns of Amur Leopards in Northeast China: Influence of Tigers, Prey, and Humans. Mamm. Biol. 2018, 92, 120–128. [Google Scholar] [CrossRef]
- Karanth, K.U.; Srivathsa, A.; Vasudev, D.; Puri, M.; Parameshwaran, R.; Kumar, N.S. Spatio-Temporal Interactions Facilitate Large Carnivore Sympatry across a Resource Gradient. Proc. R. Soc. B 2017, 284, 20161860. [Google Scholar] [CrossRef]
- Wei, W. The Suitability Evaluation of Amur Tiger (Panthera tigris altaica) and Amur Leopard (Panthera pardus orientalis) Habitat Based on Remote Sensing and GIS. Master’s Thesis, JiLin University, Changchun, China, 2019. [Google Scholar]
- Han, L. Habitat Selectivities and Seasonal Variations of Wolves (Canis lupus) in Mount Kalamaili Ungulate Nature Reserve, Xinjiang, China. Master’s Thesis, Xinjiang University, Urumqi, China, 2015. [Google Scholar]
- Allen, W.L.; Cuthill, I.C.; Scott-Samuel, N.E.; Baddeley, R. Why the Leopard Got Its Spots: Relating Pattern Development to Ecology in Felids. P. Roy. Soc. B-Biol. Sci. 2010, 278, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Bannasch, D.L.; Kaelin, C.B.; Letko, A.; Loechel, R.; Hug, P.; Jagannathan, V.; Henkel, J.; Roosje, P.; Hytonen, M.K.; Lohi, H.; et al. Dog Colour Patterns Explained by Modular Promoters of Ancient Canid Origin. Nat. Ecol. Evol. 2021, 5, 1415. [Google Scholar] [CrossRef] [PubMed]
- Botts, R.T.; Eppert, A.A.; Wiegman, T.J.; Blankenship, S.R.; Rodriguez, A.; Wagner, A.P.; Ullrich, S.E.; Allen, G.R.; Garley, W.M.; Asselin, E.M.; et al. Does Moonlight Increase Predation Risk for Elusive Mammals in Costa Rica? Trop. Conserv. Sci. 2020, 13, 194008292095240. [Google Scholar] [CrossRef]
- Palomares, F.; Caro, T.M. Interspecific Killing among Mammalian Carnivores. Am. Nat. 1999, 153, 492–508. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, Y.; Xia, F.; Bu, H.; Liu, Y.; Shen, X.; Li, S. Free-Ranging Livestock Altered the Spatiotemporal Behavior of the Endangered North Chinese Leopard (Panthera pardus japonensis) and Its Prey and Intensified Human–Leopard Conflicts. Integr. Zool. 2023, 18, 143–156. [Google Scholar] [CrossRef]
- Xinning, S.; Dazhao, S.; Qiaowen, H.; Sheng, L.; Meng, Y. Fast Surveys and Molecular Diet Analysis of Carnivores Based on Fecal DNA and Metabarcoding. Biodivers. Sci. 2019, 27, 543. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GLM | MaxEnt | Ensemble Model | |||
|---|---|---|---|---|---|---|
| AUC | TSS | AUC | TSS | AUC | TSS | |
| Leopard | 0.87 | 0.69 | 0.75 | 0.63 | 0.81 | 0.67 |
| Wolf | 0.78 | 0.69 | 0.63 | 0.65 | 0.71 | 0.67 |
| Species | PLAND | PD | ED | LSI | AI | CONNECT | PAFRAC |
|---|---|---|---|---|---|---|---|
| Leopard | 27.65 | 0.45 | 11.78 | 54.75 | 96.82 | 0.11 | 1.34 |
| Wolf | 52.14 | 1.99 | 35.09 | 118.24 | 94.94 | 0.04 | 1.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, T.; Chen, X.; Pan, H.; Jin, Y.; Zhang, X.; Xiang, Y.; Song, D.; Yang, B.; Zhang, L. Habitat Selection Differences of Two Sympatric Large Carnivores in the Southwestern Mountains of China. Diversity 2023, 15, 968. https://doi.org/10.3390/d15090968
Tian T, Chen X, Pan H, Jin Y, Zhang X, Xiang Y, Song D, Yang B, Zhang L. Habitat Selection Differences of Two Sympatric Large Carnivores in the Southwestern Mountains of China. Diversity. 2023; 15(9):968. https://doi.org/10.3390/d15090968
Chicago/Turabian StyleTian, Tengteng, Xing Chen, Han Pan, Yuyi Jin, Xiaodian Zhang, Yang Xiang, Dazhao Song, Biao Yang, and Li Zhang. 2023. "Habitat Selection Differences of Two Sympatric Large Carnivores in the Southwestern Mountains of China" Diversity 15, no. 9: 968. https://doi.org/10.3390/d15090968
APA StyleTian, T., Chen, X., Pan, H., Jin, Y., Zhang, X., Xiang, Y., Song, D., Yang, B., & Zhang, L. (2023). Habitat Selection Differences of Two Sympatric Large Carnivores in the Southwestern Mountains of China. Diversity, 15(9), 968. https://doi.org/10.3390/d15090968