

Mangrove Rehabilitation and Brachyuran Crab Biodiversity in Ranong, Thailand



Abstract
:1. Introduction
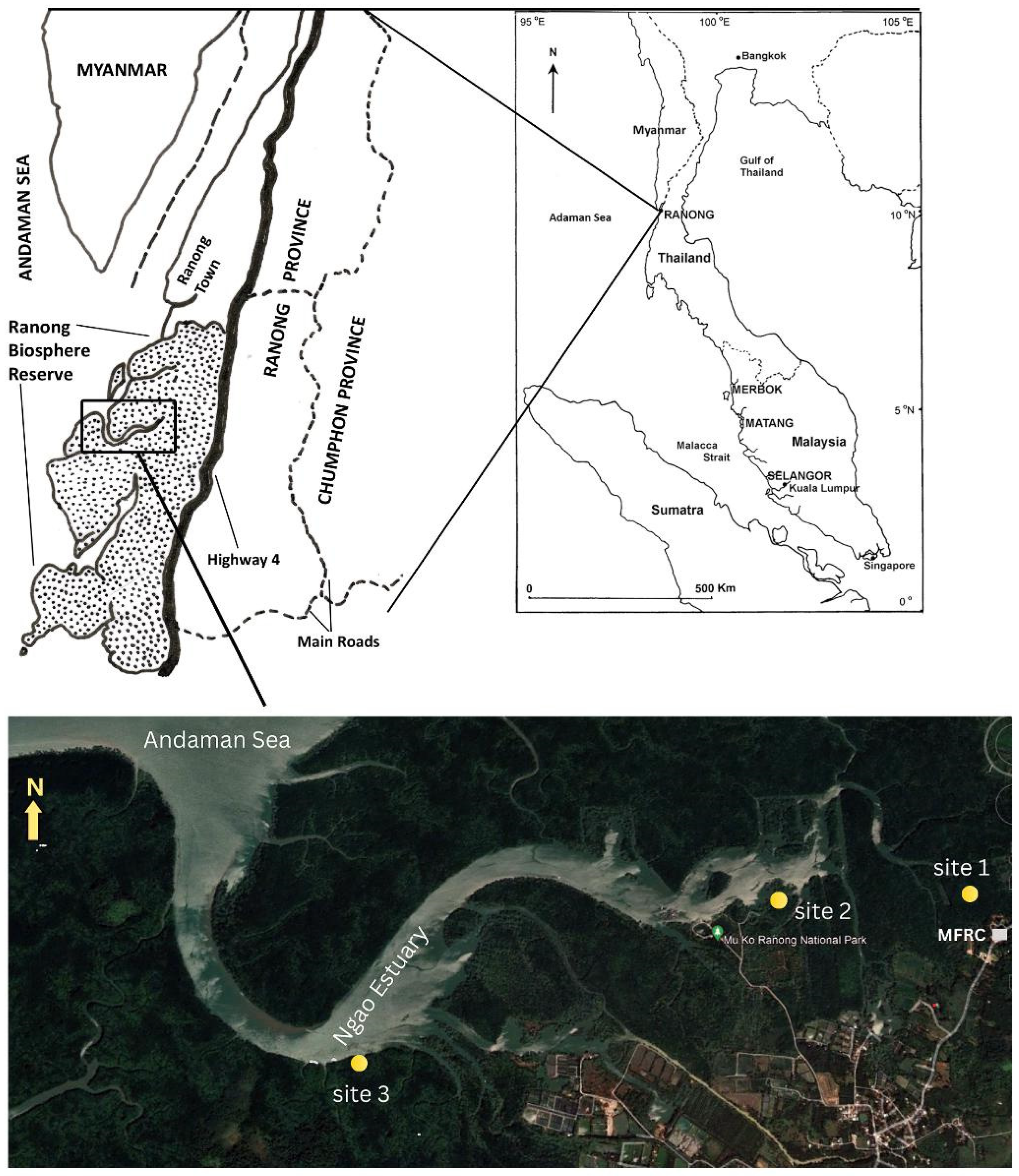
2. Materials and Methods
3. Results
3.1. Vegetation
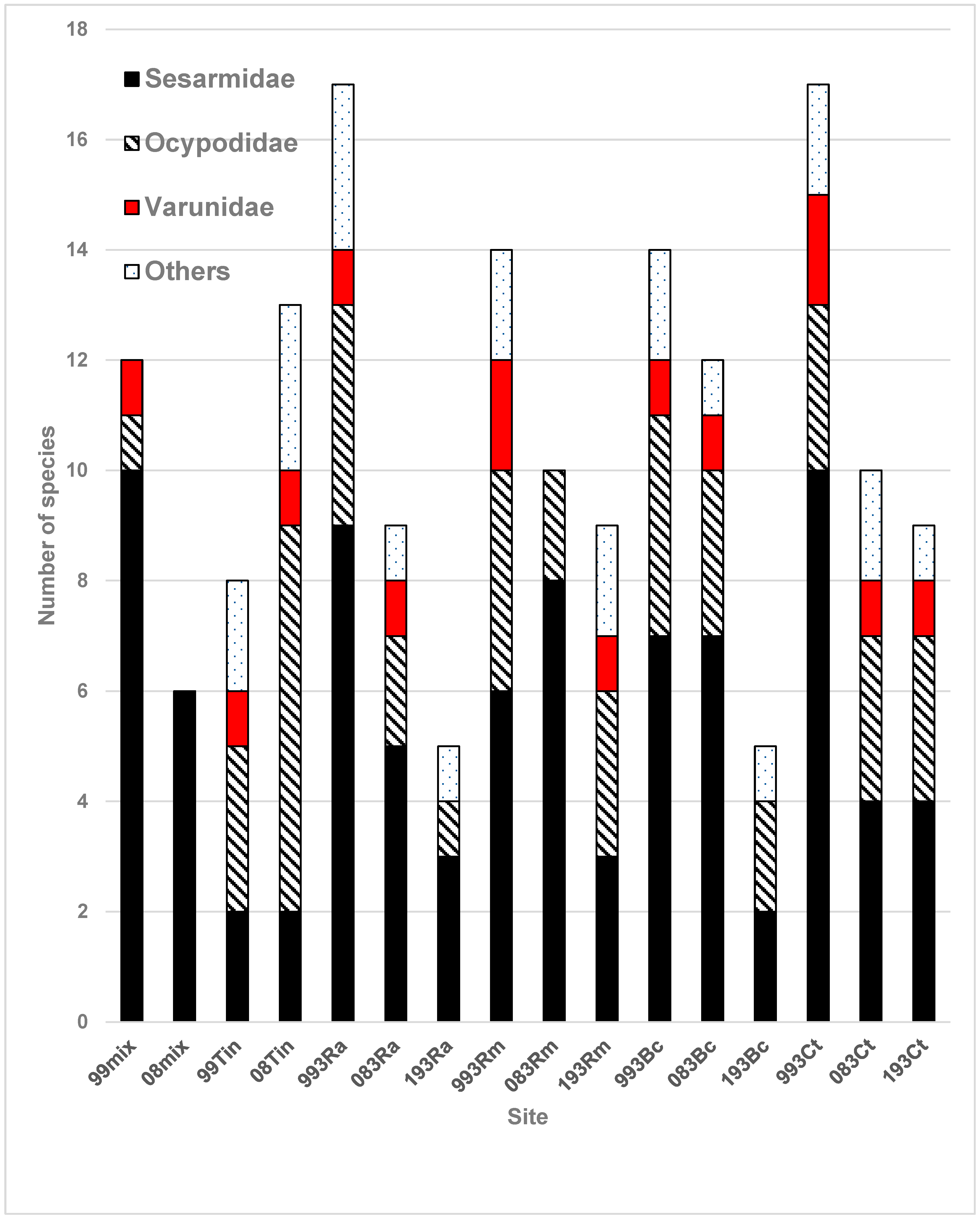
3.2. Brachyuran Crabs
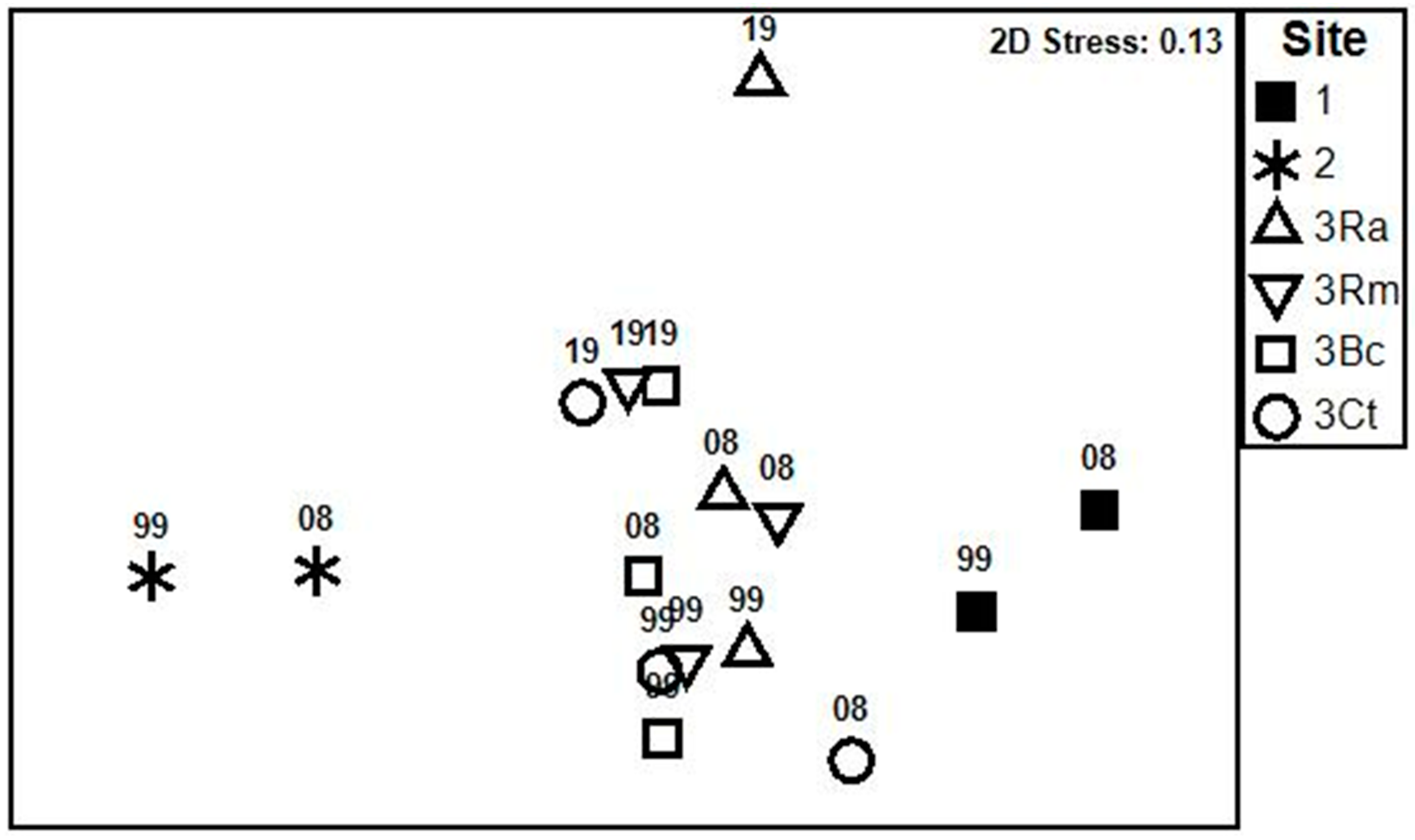
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bryan-Brown, D.N.; Connolly, R.M.; Richards, D.R.; Adame, F.; Friess, D.A.; Brown, C.J. Global trends in mangrove forest fragmentation. Sci. Rep. 2020, 10, 7117. [Google Scholar] [CrossRef] [PubMed]
- Richards, D.R.; Friess, D.A. Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proc. Natl. Acad. Sci. USA 2016, 113, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Food and Agricultural Organization of the United Nations. Mangrove Management in Thailand, Malaysia and Indonesia; FAO Environment Paper 4; Food and Agriculture Organization of the United Nations: Rome, Italy, 1985; 60p. [Google Scholar]
- Aksornkoae, S. Ecology and Management of Mangroves; International Union for Conservation of Nature and Natural Resources: Bangkok, Thailand, 1993; 192p. [Google Scholar]
- Macintosh, D.J.; Ashton, E.C. Growth and carbon stocks in four mangrove species planted on a former charcoal concession site in Ranong, Thailand. Carbon Footpr. 2023, 2, 14. [Google Scholar] [CrossRef]
- Bunting, P.; Rosenqvist, A.; Hilarides, L.; Lucas, R.M.; Thomas, N.; Tadono, T.; Worthington, T.A.; Spalding, M.D.; Murray, N.J.; Rebelo, L.-M. Global Mangrove Extent Change 1996–2020: Global Mangrove Watch Version 3.0. Remote Sens. 2022, 14, 3657. [Google Scholar] [CrossRef]
- Macintosh, D.J.; Suárez, E.L.; Sidik, F.; Pham, T.T.; Polgar, G.; Nightingale, M.; Valderrábano, M. IUCN Red List of Ecosystems, Mangroves of the Sunda Shelf. EcoEvoRxiv 2023. [Google Scholar] [CrossRef]
- Macintosh, D.J.; Suárez, E.L.; Aung, T.; Friess, D.A.; Nightingale, M.; Valderrábano, M. IUCN Red List of Ecosystems, Mangroves of the Andaman. EcoEvoRxiv 2023. [Google Scholar] [CrossRef]
- Wantongchai, P. Ranong Biosphere Reserve; Department of Marine and Coastal Resources, Ministry of Natural Resources and Development: Bangkok, Thailand, 2020; 36p.
- Macnae, W. A General Account of the Fauna and Flora of Mangrove Swamps and Forests in the Indo-West-Pacific Region. Adv. Mar. Biol. 1969, 6, 73–270. [Google Scholar] [CrossRef]
- Ashton, E.C.; Hogarth, P.J.; Macintosh, D.J. A comparison of brachyuran crab community structure at four mangrove locations under different management systems along the Melaka Straits-Andaman sea coast of Malaysia and Thailand. Estuaries 2003, 26, 1461–1471. [Google Scholar] [CrossRef]
- Saher, N.U.; Qureshi, N.A. Diversity and distribution of mangrove crabs in three intertidal areas of Balochistan, Pakistan. Pak. J. Mar. Sci. 2011, 20, 27–36. [Google Scholar]
- Ashton, E.C.; Macintosh, D.J.; Hogarth, P.J. A baseline study of the diversity and community ecology of crab and molluscan macrofauna in the Sematan mangrove forest, Sarawak, Malaysia. J. Trop. Ecol. 2003, 19, 127–142. [Google Scholar] [CrossRef]
- Rummell, A.J.; Borland, H.P.; Leon, J.X.; Henderson, C.J.; Gilby, B.L.; Ortodossi, N.L.; Mosman, J.D.; Gorissen, B.; Olds, A.D. Fish and crustaceans provide early indicators of success in wetland restoration. Restor. Ecol. 2023, 31, e13952. [Google Scholar] [CrossRef]
- Steinke, T.D.; Rajh, A.; Holland, A.J. The feeding behaviour of the red mangrove crab Sesarma meinerti De Man, 1887 (Crustacea: Decapoda: Grapsidae) and its effect on the degradation of mangrove leaf litter. S. Afr. J. Mar. Sci. 1993, 13, 151–160. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Blaber, S.J.M.; Bouillon, S.; Green, P.; Haywood, M.; Kirton, L.G.; Meynecke, J.-O.; Pawlik, J.; Penrose, H.M.; Sasekumar, A.; et al. The habitat function of mangroves for terrestrial and marine fauna: A review. Aquat. Bot. 2008, 89, 155–185. [Google Scholar] [CrossRef]
- Wilson, K.A. Ecology of mangrove crabs: Predation, physical factors and refuges. Bull. Mar. Sci. 1989, 44, 263–273. [Google Scholar]
- Lee, J.J. A conceptual model of marine detrital decomposition and the organisms associated with the process. In Advances in Aquatic Microbiology; Droop, M.R., Jannasch, H.W., Eds.; Academic Press: London, UK, 1980; pp. 257–291. [Google Scholar]
- Robertson, A.I.; Daniel, P.A. The influence of crabs on litter processing in high intertidal mangrove forests in tropical Australia. Oecologia 1989, 78, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y. Ecological role of grapsid crabs in mangrove ecosystems: A review. Mar. Freshw. Res. 1998, 49, 335–343. [Google Scholar] [CrossRef]
- Macintosh, D.J. Predation of fiddler crabs (Uca spp.) in estuarine mangroves. In Proceedings of the Symposium on Mangrove and Estuarine Vegetation, Serdang, Selangor, Malaysia, 22–28 August 1978. [Google Scholar]
- Shanij, K.; Praveen, V.P.; Suresh, S.; Oommen, M.M.; Nayar, T.S. Tree climbing and temporal niche shifting: An anti-predatory strategy in the mangrove crab Parasesarma plicatum (Latreille, 1803). Curr. Sci. 2016, 111, 1201–1207. [Google Scholar] [CrossRef]
- Warren, J.H.; Underwood, A.J. Effects of burrowing crabs on the topography of mangrove swamps in New South Wales. J. Exp. Mar. Biol. Ecol. 1986, 102, 223–235. [Google Scholar] [CrossRef]
- Frith, D.W.; Tantanasiriwong, R.; Bhatia, O. Zonation of Macrofauna on a Mangrove Shore, Phuket Island; Phuket Marine Biological Center, Research Bulletin: Phuket, Thailand, 1976; Volume 10, 37p. [Google Scholar]
- Macintosh, D.J.; Aksornkoae, S.; Vannucci, M.; Field, C.D.; Clough, B.F.; Kjerfve, B.; Paphavasit, N.; Wattayakorn, G. Final Report of the Integrated Multidisciplinary Survey and Research Programme of the Ranong Mangrove Ecosystem; UNDP/UNESCO regional project: Research and its application in the management of the mangroves of Asia and the Pacific; Funny Publishing Limited Partnership: Bangkok, Thailand, 1991; 183p. [Google Scholar]
- Macintosh, D.J.; Ashton, E.C.; Havanon, S. Mangrove rehabilitation and intertidal biodiversity: A study in the Ranong mangrove ecosystem, Thailand. Estuar. Coast. Shelf Sci. 2002, 55, 331–345. [Google Scholar] [CrossRef]
- Ng, P.K.L.; Guinot, D.; Davie, P.J.F. Systema Brachyurorum: Part 1. An annotated checklist of extant brachyuran crabs of the world. Raffles Bull. Zool. 2008, 17, 1–286. [Google Scholar]
- Shih, H.-T.; Ng, P.K.L.; Davie, P.J.F.; Schubart, C.D.; Türkay, M.; Naderloo, R.; Jones, D.; Liu, M.-Y. Systematics of the family Ocypodidae Rafinesque, 1815 (Crustacea: Brachyura), based on phylogenetic relationships, with a reorganization of subfamily rankings and a review of the taxonomic status of Uca Leach, 1814, sensu lato and its subgenera. Raffles Bull. Zool. 2016, 64, 139–175. [Google Scholar]
- Shih, H.-T.; Chan, B.K.K. Systematics and biogeography of fiddler crabs—A special issue in Zoological studies. Zool. Stud. 2022, 61, 64. [Google Scholar] [CrossRef]
- Shih, H.-T.; Wong, K.J.H.; Chan, B.K.K.; Nguyen, T.D.; Do, V.T.; Ngo, X.Q.; Hsu, P.-Y. Diversity and Distribution of Fiddler Crabs (Crustacea: Brachyura: Ocypodidae) in Vietnam. Zool. Stud. 2022, 61, 66. [Google Scholar] [CrossRef]
- Rahim, N.H.A.; Shuib, S.; Yahya, K. Rapid baseline assessment of crab abundance and species richness in mangroves using a video recording method. IOP Conf. Ser. Earth Environ. Sci. 2021, 736, 012056. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423, 623–656. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; 190p. [Google Scholar]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; Primer-E Ltd.: Plymouth, UK, 2001. [Google Scholar]
- Somerfield, P.J.; Clarke, K.R.; Gorley, R.N. Analysis of similarities (ANOSIM) for 2-way layouts using a generalised ANOSIM statistic, with comparative notes on Permutational Multivariate Analysis of Variance (PERMANOVA). Austral Ecol. 2021, 46, 911–926. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aus. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Similarity-based testing for community pattern: The 2-way layout with no replication. Mar. Biol. 1994, 118, 167–176. [Google Scholar] [CrossRef]
- Mukri, M.S.; Shuhaida, S. The habitat preferences of mangrove crabs in different mangrove forests of Penang, North Peninsular Malaysia. IOP Conf Ser. Earth Environ. Sci. 2021, 736, 012062. [Google Scholar] [CrossRef]
- Aksornkoae, S.; Paphavasit, N.; Wattayakorn, G. Mangroves of Thailand: Present status of conservation, use and management. In The Economic and Environmental Values of Mangrove Forests and Their Present State of Conservation in the South-East Asia/Pacific Region; Clough, B., Ed.; Mangrove Ecosystems Technical Reports 1; International Society for Mangrove Ecosystems: Okinawa, Japan, 1993; pp. 83–133. [Google Scholar]
- Shih, H.T.; Hsu, J.W.; Wong, K.J.H.; Ng, N.K. Review of the mudflat varunid crab genus Metaplax (Crustacea, Brachyura, Varunidae) from East Asia and northern Vietnam. ZooKeys 2019, 877, 1–29. [Google Scholar] [CrossRef]
- Herbon, C.M.; Nordhaus, I. Experimental determination of stable carbon and nitrogen isotope fractionation between mangrove leaves and crabs. Mar. Ecol. Prog. Ser. 2013, 490, 91–105. [Google Scholar] [CrossRef]
- Macintosh, D.J. Ecology and productivity of Malaysian mangrove crab populations (Decadopa: Brachyura). In Proceedings of the Asian Symposium on Mangrove Environment, Research and Management; Soepadmo, E., Rao, A.N., Macintosh, D.J., Eds.; University of Malaya and UNESCO: Kuala Lumpur, Malaysia, 1984; pp. 354–377. [Google Scholar]
- Sasekumar, A. Distribution of macrofauna on a Malayan mangrove shore. J. Anim. Ecol. 1974, 43, 51–69. [Google Scholar] [CrossRef]
- Colpo, K.D.; Negreiros-Fransozo, M.L. Morphological diversity of setae on the second maxilliped of fiddler crabs (Decapoda: Ocypodidae) from the southwestern Atlantic coast. Invertebr. Biol. 2013, 132, 38–45. [Google Scholar] [CrossRef]
- Peer, N.; Miranda, N.A.F.; Perissinotto, R. Impact of fiddler crab activity on microphytobenthic communities in a South African mangrove forest. Estuar. Coast. Shelf Sci. 2019, 31, 227. [Google Scholar] [CrossRef]
- Dye, A.H.; Lasiak, T.A. Assimilation efficiencies of fiddler crabs and deposit-feeding gastropods from tropical mangrove sediments. Comp. Biochem. Physiol. Part A Physiol. 1987, 87, 341–344. [Google Scholar] [CrossRef]
- Nielsen, O.I.; Kristensen, E.; Macintosh, D.J. Impact of fiddler crabs (Uca spp.) on rates and pathways of benthic mineralization in deposited mangrove shrimp pond waste. J. Exp. Mar. Biol. Ecol. 2003, 289, 59–81. [Google Scholar] [CrossRef]
- Gillis, L.G.; Snavely, E.; Lovelock, C.; Zimmer, M. Effects of crab burrows on sediment characteristics in a Ceriops australis-dominated mangrove forest. Estuar. Coast. Shelf Sci. 2019, 218, 334–339. [Google Scholar] [CrossRef]
- Smith, N.F.; Wilcox, C.; Lessmann, J.M. Fiddler crab burrowing affects growth and production of the white mangrove (Laguncularia racemosa) in a restored Florida coastal marsh. Mar. Biol. 2009, 156, 2255–2266. [Google Scholar] [CrossRef]
- Malley, D.F. Degradation of mangrove leaf litter by the tropical sesarmid crab Chiromanthes onychophorum. Mar. Biol. 1978, 49, 377–386. [Google Scholar] [CrossRef]
- Cannicci, S.; Burrows, D.; Fratini, S.; Smith, T.J., III; Offenberg, J.; Dahdouh-Guebas, F. Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquat. Bot. 2008, 89, 186–200. [Google Scholar] [CrossRef]
- Chen, G.; Ye, Y. Changes in properties of mangrove sediment due to foraging on Kandelia obovata leaves by crabs Parasesarmea plicatum (Grapsidae: Sesarminae). Mar. Ecol. Prog. Ser. 2010, 419, 137–145. [Google Scholar] [CrossRef]
- Smith III, T.J.; Boto, K.G.; Frusher, S.D.; Giddins, R.L. Keystone species and mangrove forest dynamics: The influence of burrowing by crabs on soil nutrient status and forest productivity. Estuar. Coast. Shelf Sci. 1991, 33, 419–432. [Google Scholar] [CrossRef]
- Kristensen, E. Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. J. Sea Res. 2008, 59, 30–43. [Google Scholar] [CrossRef]
- Shunyang, C.; Chen, G.; Chen, B.; Yong, Y.; Zhiyuan, M.A. Feeding ecology of Sesarmid crabs in mangroves. Acta Ecol. Sin. 2014, 34, 5349–5359. [Google Scholar] [CrossRef]
- Harada, Y.; Lee, S.Y. Foraging behavior of the mangrove sesarmid crab Neosarmatium trispinosum enhances food intake and nutrient retention in a low-quality food environment. Estuar. Coast. Shelf Sci. 2016, 174, 41–48. [Google Scholar] [CrossRef]
- Micheli, F. Feeding ecology of mangrove crabs in North Eastern Australia: Mangrove litter consumption by Sesarma messa and Sesarma smithii. J. Exp. Mar. Biol. Ecol. 1993, 171, 165–186. [Google Scholar] [CrossRef]
- Ashton, E.C. Mangrove Sesarmid Crab Feeding Experiments in Peninsular Malaysia. J. Exp. Mar. Biol. Ecol. 2002, 273, 97–119. [Google Scholar] [CrossRef]
- Ya, B.P.; Yeo, D.C.J.; Todd, P.A. Feeding Ecology of Two Species of Perisesarma (Crustacea: Decapoda: Brachyura: Sesarmidae) in Mandai Mangroves, Singapore. J. Crustac. Biol. 2008, 28, 480–484. Available online: https://www.jstor.org/stable/20487759 (accessed on 10 September 2023). [CrossRef]
- Forgeron, S.J.; Quadros, A.F.; Zimmer, M. Crab-driven processing does not explain leaf litter-deposition in mangrove crab burrows. Ecol. Evol. 2021, 11, 8856–8862. [Google Scholar] [CrossRef]
- Leh, C.M.U.; Sasekumar, A. The food of sesarmid crabs in Malaysian mangrove forests. Malay. Nat. J. 1985, 39, 135–145. [Google Scholar]
- Gao, X.; Lee, S.Y. Feeding strategies of mangrove leaf-eating crabs for meeting their nitrogen needs on a low-nutrient diet. Front. Mar. Sci. 2022, 9, 872272. [Google Scholar] [CrossRef]
- Ferriera, A.C.; Alencar, C.E.R.D.; Bezera, L.E.A. Interrelationships among ecological factors of brachyuran crabs, trees and soil in mangrove community assemblage in Northeast Brazil. Community Ecol. 2019, 20, 277–290. [Google Scholar] [CrossRef]
- Moser, S.M.; Macintosh, D.J. Diurnal and lunar patterns of larval recruitment of Brachyura into a mangrove estuary system in Ranong Province, Thailand. Mar. Biol. 2001, 138, 827–841. [Google Scholar] [CrossRef]
- Macintosh, D.J.; Goncalves, F.; Soares, A.M.V.M.; Moser, S.M.; Paphavisit, N. Transport mechanisms of crab megalopae in mangrove ecosystem, with special reference to a mangrove estuary in Ranong, Thailand. In Mud Crab Aquaculture and Biology, Proceedings of the International Scientific Forum, Darwin, Australia, 21–24 April 1997; Keenan, C.P., Blackshaw, A., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1999; ACIAR Proceedings No. 78; pp. 178–186. [Google Scholar]
- Morgan, S.G.; Christy, J.H. Adaptive significance of the timing of larval release by crabs. Am. Nat. 1995, 145, 457–479. Available online: https://stri-sites.si.edu/docs/publications/pdfs/Christy_1995_Am_Nat_wSteve.pdf (accessed on 25 January 2024). [CrossRef]
- Thong, K.L.; Sasekumar, A. The trophic relationships of the fish community of the Angsa Bank, Selangor, Malaysia. In Proceedings of the Asian Symposium on Mangrove Environment, Research and Management; Soepadmo, E., Rao, A.N., Macintosh, D.J., Eds.; University of Malaya and UNESCO: Kuala Lumpur, Malaysia, 1984; pp. 385–399. [Google Scholar]
- Leh, M.U.C.; Sasekumar, A.; Chew, L.L. Feeding biology of eel catfish Plotosus canius Hamilton in a Malaysian mangrove estuary and mudflat. Raffles Bull. Zool. 2012, 60, 551–557. [Google Scholar]
- Sasekumar, A.; Ong, T.L.; Thong, K.L. Predation of mangrove fauna by marine fishes. In Proceedings of the Asian Symposium on Mangrove Environment, Research and Management; Soepadmo, E., Rao, A.N., Macintosh, D.J., Eds.; University of Malaya and UNESCO: Kuala Lumpur, Malaysia, 1984; pp. 378–384. [Google Scholar]
- Sheaves, M.; Mohony, B. Short-circuit in the mangrove food chain. Mar. Ecol. Prog. Ser. 2000, 199, 97–109. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
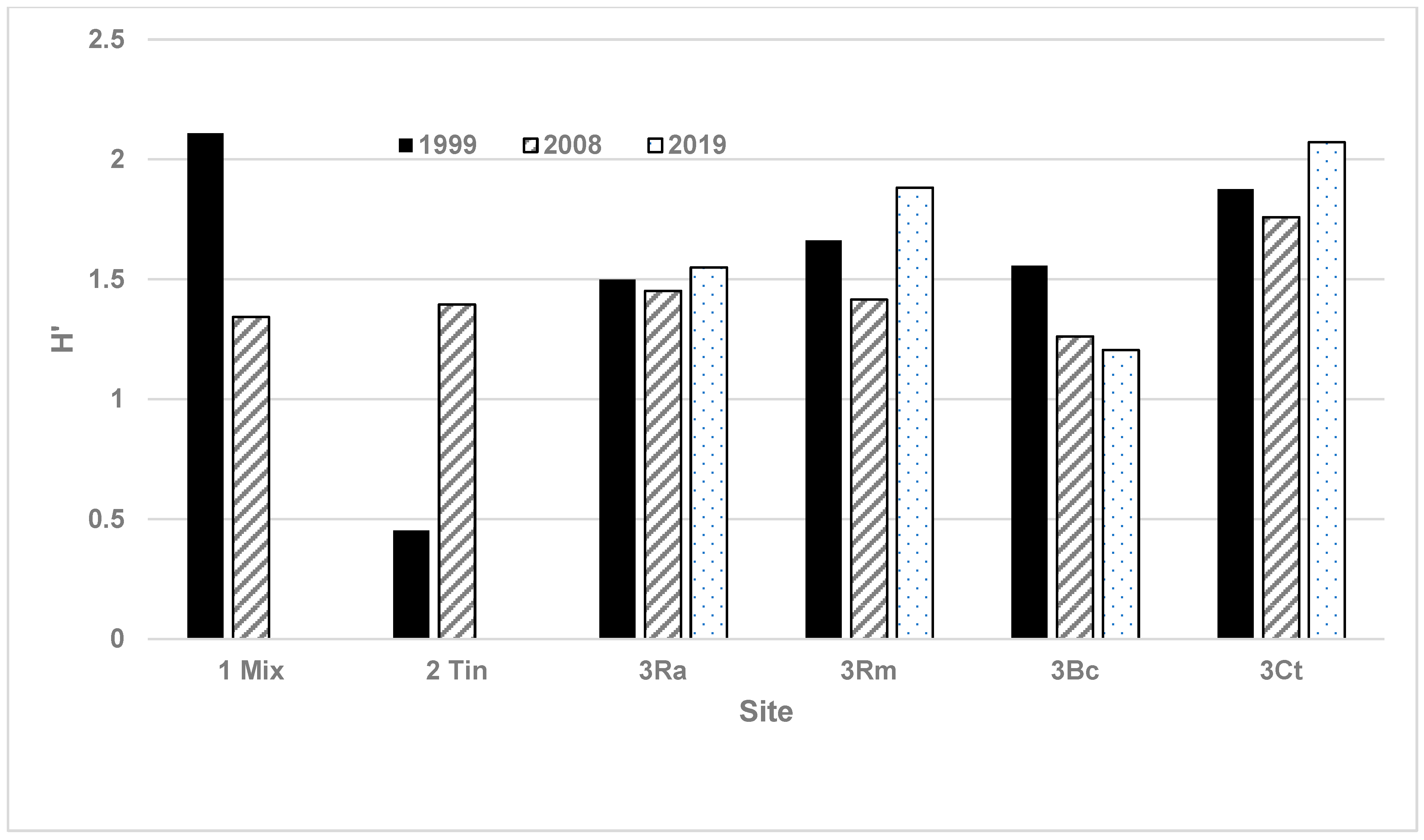{kind=link}
{kind=link}
| Site | 1 | 2 | 3Ra | 3Rm | 3Bc | 3Ct |
|---|---|---|---|---|---|---|
| History | Mixed conservation forest (Rhizophora-dominated) | Tin mining until 1985 | Charcoal concession forest clear felled in 1994 | |||
| Replanted | natural | 1985 | 1994 | 1994 | 1995 | 1995 |
| TH (cm) | 342 ± 0 | 260 ± 0 | 319 ± 9 | 327 ± 2 | 322 ± 10 | 299 ± 10 |
| XF (days) | 295 | 365 | 334 | 323 | 325 | 352 |
| H’ (99) | 1.36 | 0.93 | 0 | 0 | 0.15 | 0 |
| H’ (08) | 1.19 | 0.49 | 0.93 | 0 | 0.13 | 0.44 |
| H’ (19) | - | - | 0.11 | 0 | 0 | 0 |
| H (99) | 8.9 ± 4.7 | 6.9 ± 1.7 | 3.2 ± 0.3 | 4.0 ± 0.8 | 1.3 ± 0.4 | 1.5 ± 0.2 |
| H (08) | 11.9 ± 7.7 | 3.5 ± 1.4 | 10.7 ± 2.9 | 12.9 ± 1.6 | 5.0 ± 0.02 | 4.9 ± 0.5 |
| H (19) | - | - | 12.1 ± 3.8 | 19.6 ± 2.3 | 12.9 ± 0.9 | 12.2 ± 0.6 |
| DBH (99) | 9.0 ± 9.3 | 6.1 ± 3.2 | 2.7 ± 0.6 | 3.4 ± 0.8 | 1.4 ± 0.7 | 1.8 ± 0.8 |
| DBH (08) | 12.7 ± 11.2 | 4.7 ± 1.9 | 7.6 ± 1.9 | 10.1 ± 2.6 | 5.2 ± 1.8 | 6.4 ± 2.4 |
| DBH (19) | - | - | 7.9 ± 2.7 | 9.1 ± 6.7 | 7.5 ± 3.4 | 8.6 ± 3.4 |
| A (99) | 33 | 22 | 38 | 32 | 29 | 55 |
| A (08) | 25 | 14 | 39 | 32 | 35 | 53 |
| A (19) | - | - | 39 | 32 | 40 | 44 |
| BA (99) | 43 | 8.1 | 2.6 | 3.8 | 0.9 | 2.4 |
| BA (08) | 60 | 5.7 | 20 | 33 | 15 | 29 |
| BA (19) | - | - | 32 | 50 | 33 | 38 |
| Site | 1 | 1 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Main Plant Species | Mix | Mix | Rm | Rm | Ra | Ra | Ra | Ra | Ra | Ra | Bc | Bc | Bc | Ct | Ct | Ct |
| Year | 99 | 08 | 99 | 08 | 99 | 08 | 19 | 99 | 08 | 19 | 99 | 08 | 19 | 99 | 08 | 19 |
| Superfamily Grapsoidea | ||||||||||||||||
| Family Grapsidae | ||||||||||||||||
| Metograpsus latifrons (White, 1847) | + | + | ||||||||||||||
| Family Sesarmidae | ||||||||||||||||
| Clistocoeloma merguiense De Man, 1888 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Episesarma versicolor (Tweedie, 1940) | + | + | + | + | + | + | + | |||||||||
| Nanosesarma batavicum (Moreira, 1903) | + | + | + | |||||||||||||
| Neosarmatium sp. (Serène and Soh, 1970) | + | |||||||||||||||
| Parasesarma lenzii (De Man, 1895) | + | + | + | + | + | + | + | + | + | + | ||||||
| Parasesarma melissa (De Man, 1888) | + | + | + | + | + | |||||||||||
| Parasesarma rutilimanum (Tweedie, 1936) | + | + | + | + | + | + | ||||||||||
| Parasesarma sp 1 | + | + | + | + | + | + | + | + | + | |||||||
| Parasesarma sp 2 | + | |||||||||||||||
| Perisesarma darwinense (Campbell, 1967) | + | + | + | + | + | |||||||||||
| Perisesarma eumolpe (De Man, 1895) | + | + | + | + | + | + | + | |||||||||
| Perisesarma onychophorum (De Man, 1895) | + | + | + | + | + | + | + | + | ||||||||
| Perisesarma sp. | + | |||||||||||||||
| Sarmatium sp. | + | + | ||||||||||||||
| Sesarmoides kraussi (De Man, 1888) | + | + | + | + | + | + | + | |||||||||
| Family Varunidae | ||||||||||||||||
| Metaplax elegans (De Man, 1888) | + | + | + | + | + | + | + | + | ||||||||
| Metaplax sheni (Gordon, 1930) | + | + | + | + | + | + | ||||||||||
| Super Family Ocypodoidea | ||||||||||||||||
| Family Camptandriidae | ||||||||||||||||
| Paracleistostoma depressum (De Man, 1895) | + | + | ||||||||||||||
| Tylodiplax tetratylophora (De Man, 1895) | + | + | + | |||||||||||||
| Family Dotiliidae | ||||||||||||||||
| Ilyoplax delsmani (De Man, 1926) | + | |||||||||||||||
| Ilyoplax obliqua (Tweedie, 1935) | + | |||||||||||||||
| Ilyoplax punctata (Tweedie, 1935) | + | + | + | |||||||||||||
| Ilyoplax sp. | + | |||||||||||||||
| Family Ocypodidae | ||||||||||||||||
| Austruca lactea (De Haan, 1835) | + | + | + | + | + | + | ||||||||||
| Austruca triangularis (A. Milne-Edwards, 1873) | + | + | + | + | + | + | + | + | + | + | + | + | + | |||
| Tubuca coarctata (H Milne Edwards, 1852) | + | |||||||||||||||
| Tubuca dussumieri (H Milne Edwards, 1852) | + | |||||||||||||||
| Tubuca forcipata (Adams and White, 1849) | + | |||||||||||||||
| Tubuca rosea (Tweedie, 1937) | + | + | + | + | + | + | + | + | + | + | + | + | ||||
| Family Macrophthalmidae | ||||||||||||||||
| Ilyograpsus paludicola (Rathbun, 1909) | + | |||||||||||||||
| Super Family Pilumnoidea | ||||||||||||||||
| Family Pilumnidae | ||||||||||||||||
| Heteropanope glabra (Stimpson, 1858) | + | + | + | + | + | + | + | + | + | |||||||
| Heteropanope sp. | + | |||||||||||||||
| Super Family Paguroidea | ||||||||||||||||
| Family Diogenidae | ||||||||||||||||
| Clibanarius padavensis De Man, 1888 | + | + | + | + | + | + | ||||||||||
| Clibanarius sp. | + | + | ||||||||||||||
| Family Thalassinidae | ||||||||||||||||
| Thalassina sp. | + | |||||||||||||||
| Family Upogebiidae | ||||||||||||||||
| Upogebia sp. | + |
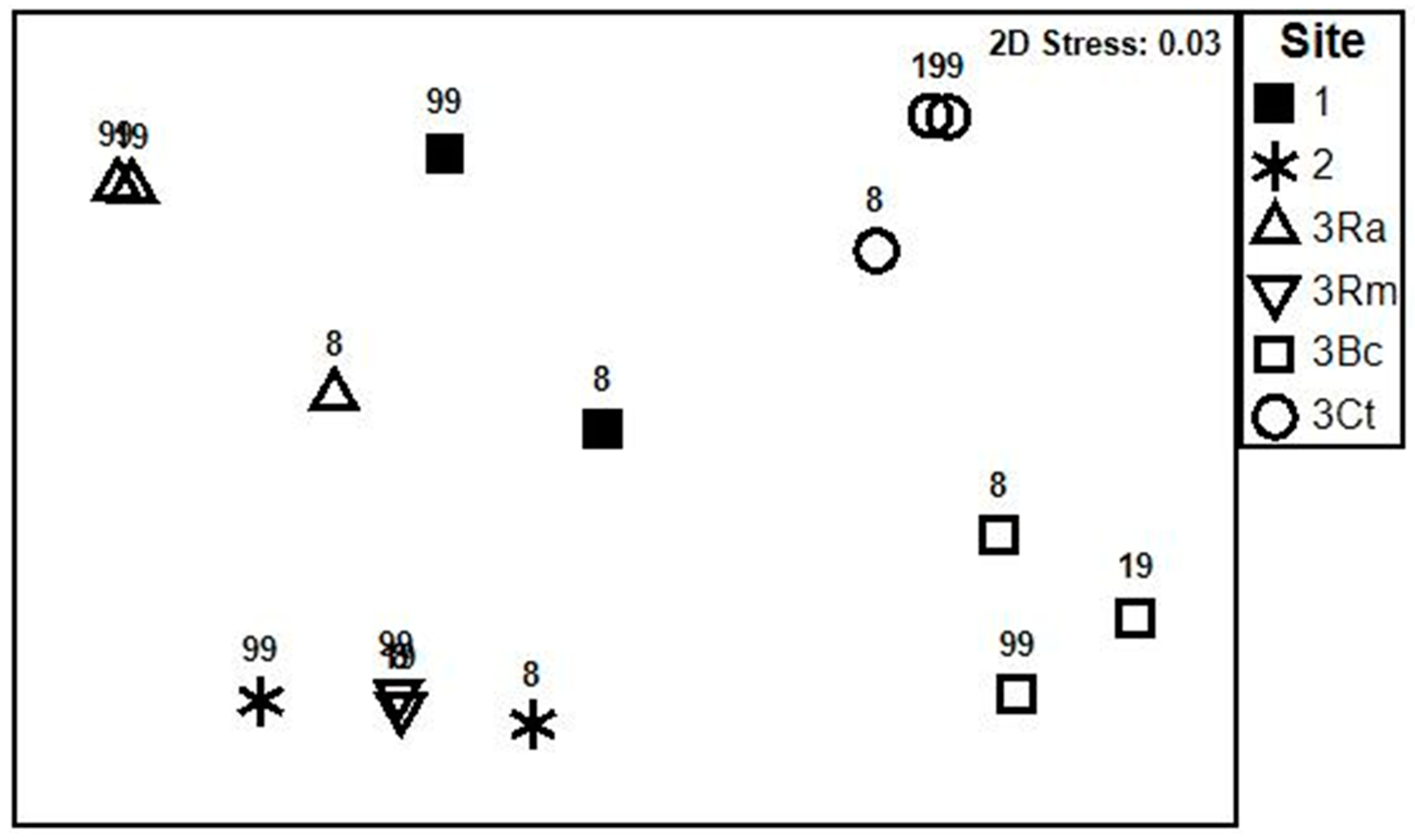
| Pairwise Comparisons between Sites | Global R | Significance P% |
|---|---|---|
| 1, 2 | 1.0 | 0.3 |
| 1, 3Ra | 0.54 | 0.2 |
| 1, 3Rm | 0.68 | 0.1 |
| 1, 3Bc | 0.93 | 0.1 |
| 1, 3Ct | 0.85 | 0.1 |
| 2, 3Ra | 0.97 | 0.1 |
| 2, 3Rm | 0.98 | 0.2 |
| 2, 3Bc | 0.99 | 0.1 |
| 2, 3Ct | 0.98 | 0.1 |
| 3Ra, 3Rm | 0.08 | 21.1 |
| 3Ra, 3Bc | 0.29 | 0.5 |
| 3Ra, 3Ct | 0.24 | 1.8 |
| 3Rm, 3Bc | 0.05 | 29.1 |
| 3Rm, 3Ct | 0.06 | 23.0 |
| 3Bc, 3Ct | 0.05 | 25.9 |
| Year | 99 | 08 | 99 | 08 | 99 | 08 | 19 | 99 | 08 | 19 | 99 | 08 | 19 | 99 | 08 | 19 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | 1 | 1 | 2 | 2 | 3Ra | 3Ra | 3Ra | 3 Rm | 3 Rm | 3 Rm | 3Bc | 3Bc | 3Bc | 3Ct | 3Ct | 3Ct |
| Cm | 4.67 | 5.25 | 0.67 | 0.25 | 3 | 2.5 | 0.25 | 2.67 | 0.25 | 0.25 | 0.5 | 1.25 | 0.25 | 1.17 | 2.25 | 0.5 |
| Pl | 2.67 | 3.33 | 0.67 | 0.25 | 0.67 | 1 | 0.25 | 0.25 | 0.5 | 0.5 | ||||||
| Pr | 0.67 | 1.25 | 0.17 | 0.67 | 0.25 | 0.17 | ||||||||||
| Nb | 0.33 | 0.25 | 0.17 | |||||||||||||
| Sk | 0.67 | 1 | 0.17 | 0.17 | 0.25 | 0.33 | 0.5 | |||||||||
| Me | 37.3 | 18 | 0.5 | 0.33 | 0.25 | 0.25 | 0.33 | 0.5 | ||||||||
| Tt | 1.33 | 1.25 | 0.25 | |||||||||||||
| Ml | 0.33 | 0.5 | ||||||||||||||
| Tr | 0.33 | 3 | 3.17 | 0.25 | 8.17 | 1.5 | 1.25 | 8 | 5.25 | 2.75 | 8 | 1.25 | ||||
| At | 1 | 0.25 | 24.2 | 5.75 | 20.7 | 8 | 1.5 | 32.3 | 17 | 2 | 16.2 | 4 | 1 | |||
| Pe | 0.25 | 0.5 | 0.5 | 0.25 | 0.5 | 0 | 0.33 | 0.25 | ||||||||
| Po | 1 | 0.25 | 2.83 | 0.25 | 0.25 | 5.17 | 3.67 | 4.17 | ||||||||
| Hg | 0.25 | 0.33 | 0.5 | 0.5 | 0.25 | 0.1 7 | 0.25 | 0.17 | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashton, E.C.; Macintosh, D.J. Mangrove Rehabilitation and Brachyuran Crab Biodiversity in Ranong, Thailand. Diversity 2024, 16, 92. https://doi.org/10.3390/d16020092
Ashton EC, Macintosh DJ. Mangrove Rehabilitation and Brachyuran Crab Biodiversity in Ranong, Thailand. Diversity. 2024; 16(2):92. https://doi.org/10.3390/d16020092
Chicago/Turabian StyleAshton, Elizabeth C., and Donald J. Macintosh. 2024. "Mangrove Rehabilitation and Brachyuran Crab Biodiversity in Ranong, Thailand" Diversity 16, no. 2: 92. https://doi.org/10.3390/d16020092
APA StyleAshton, E. C., & Macintosh, D. J. (2024). Mangrove Rehabilitation and Brachyuran Crab Biodiversity in Ranong, Thailand. Diversity, 16(2), 92. https://doi.org/10.3390/d16020092