
Genetic Variation in the Pallas’s Cat (Otocolobus manul) in Zoo-Managed and Wild Populations

 ,
, 
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
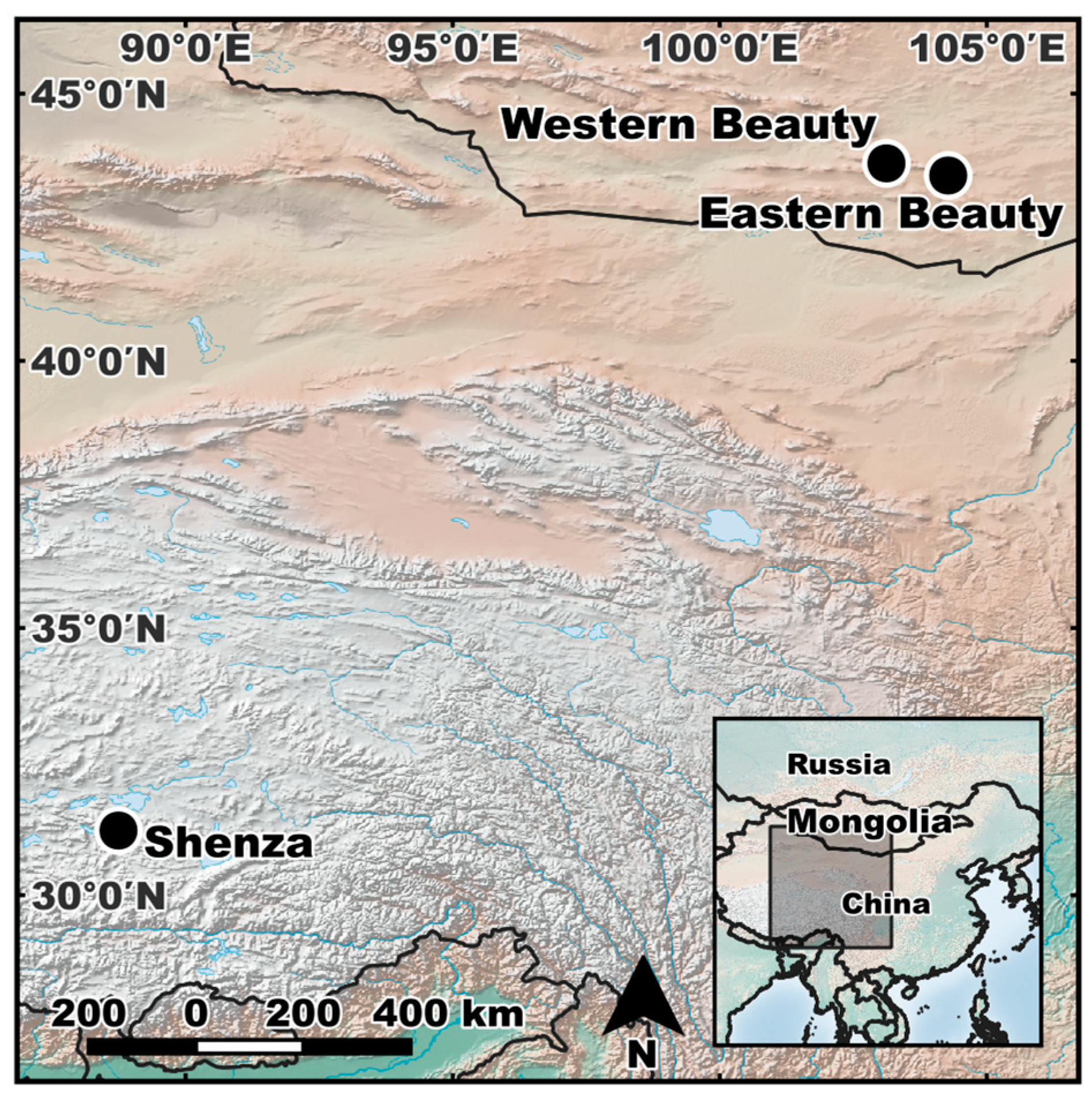
2.1. Sample Collection
2.2. Genotyping of Microsatellites
2.3. General Analysis of Microsatellites
2.4. Microsatellite Comparison with Snow Leopards
2.5. Microsatellite Analysis of the Zoo-Managed Population
2.6. Sequencing & Analysis of MT-RNR1
2.7. Sequencing & Analysis of EPAS1
3. Results
3.1. General Analysis of Microsatellites
3.2. Microsatellite Comparison with Snow Leopards
3.3. Microsatellite Analysis of the Zoo-Managed Population
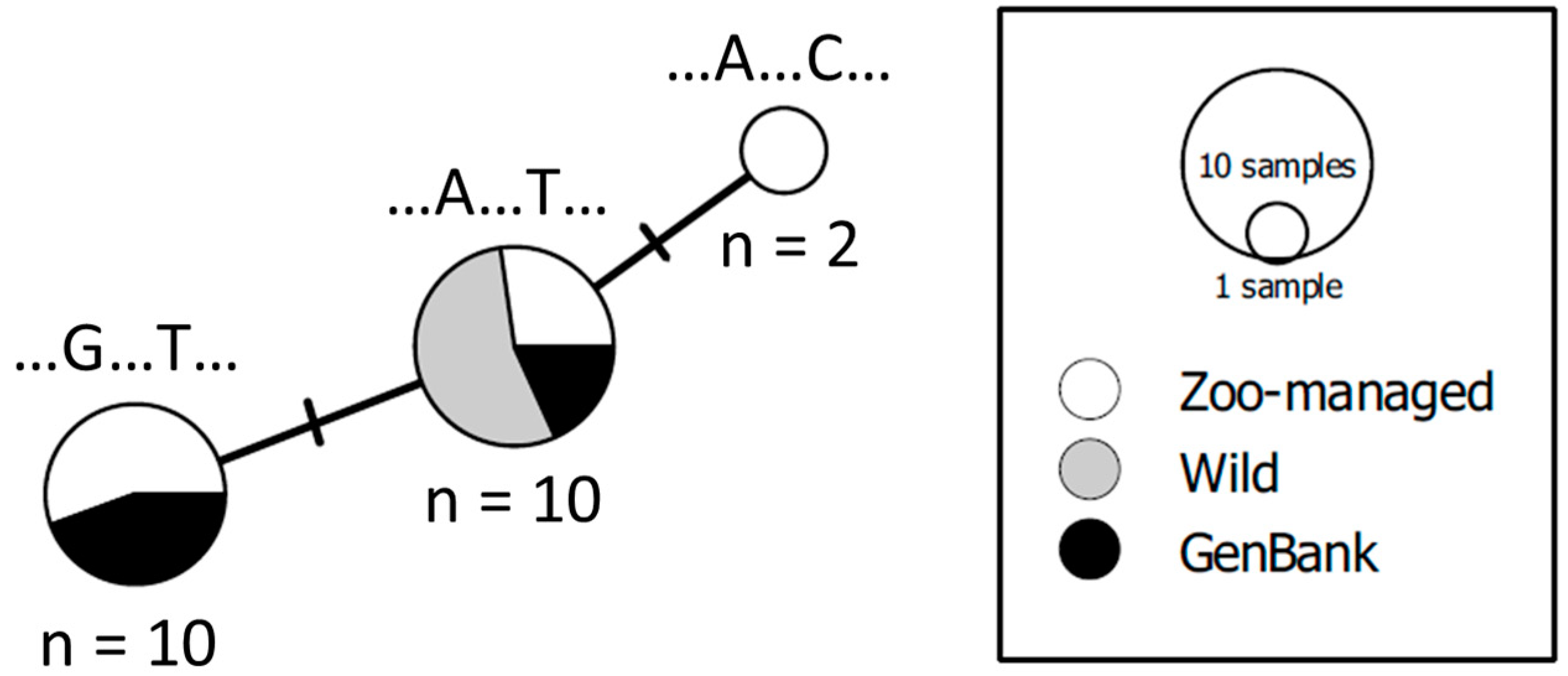
3.4. Analysis of MT-RNR1
3.5. Analysis of EPAS1
4. Discussion
4.1. Summary of Microsatellite Loci
4.2. Initial Estimates of Genetic Diversity in the Pallas’s Cat
4.3. Diversity in the Zoo-Managed Population
4.4. EPAS1 Variant in the Pallas’s Cat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tensen, L. Biases in wildlife and conservation research, using felids and canids as a case study. Glob. Ecol. Conserv. 2018, 15, e00423. [Google Scholar] [CrossRef]
- Ross, S. Providing an Ecological Basis for the Conservation of the Pallas’s Cat (Otocolobus manul). Ph.D. Thesis, University of Bristol, Bristol, UK, 2009. [Google Scholar] [CrossRef]
- Ross, S.; Barashkova, A.; Kirilyuk, V.; Naidenko, S. The behaviour and ecology of the manul. Cat News 2019, 13, 9–13. [Google Scholar]
- Ross, S.; Barashkova, A.; Dhendup, T.; Munkhtsog, B.; Smelansky, I.; Barclay, D.; Moqanaki, E. Otocolobus manul (errata version published in 2020). The IUCN Red List of Threatened Species. 2020. Available online: https://www.researchgate.net/publication/342945167_Otocolobus_manul_errata_version_published_in_2020_The_IUCN_Red_List_of_Threatened_Species (accessed on 11 November 2022).
- Lanz, T.; Breitenmoser-Würsten, C.; Barclay, D.; Nygren, E.; Samelius, G.; Breitenmoser, U. Prologue: Why care about Otocolobus manul? Cat News 2019, 13, 5–8. [Google Scholar]
- Ross, S.; Munkhtsog, B.; Harris, S. Determinants of mesocarnivore range use: Relative effects of prey and habitat properties on Pallas’s cat home-range Size. J. Mammal. 2012, 93, 1292–1300. [Google Scholar] [CrossRef]
- Barashkova, A.N.; Kirilyuk, V.E.; Smelansky, I.E. Significance of protected areas for the Pallas’s cat (Otocolobus manul: Felidae) conservation in Russia. Nat. Conserv. Res. 2017, 2, 113–124. [Google Scholar] [CrossRef]
- Greenspan, E.; Giordano, A.J. A rangewide distribution model for the Pallas’s cat (Otocolobus manul): Identifying potential new survey regions for an understudied small cat. Mammalia 2021, 85, 574–587. [Google Scholar] [CrossRef]
- Chalani, M.; Ghoddousi, A.; Ghadirian, T.; Goljani, R. First Pallas’s cat photo-trapped in Khojir National Park, Iran. Cat News 2008, 49, 7. [Google Scholar]
- Pal, R.; Bhattacharya, T.; Sathyakumar, S. First record of Pallas’s cat in Uttarakhand, Nelang Valley, Gangotri National Park, India. Cat News 2019, 69, 24–25. [Google Scholar]
- Barashkova, A.; Smelansky, I.; Kirilyuk, V.; Naidenko, S.; Antonevich, A.; Gritsina, M.; Uulu, K.Z.; Koshkin, M.; Battogtokh, N.; Otgonbayar, B.; et al. Distribution and status of the manul in central Asia and adjacent areas. Cat News 2019, 13, 14–23. [Google Scholar]
- Swanson, W.F. Application of assisted reproduction for population management in felids: The potential and reality for conservation of small cats. Theriogenology 2006, 66, 49–58. [Google Scholar] [CrossRef]
- Kayser, S.; Bladow, R. Population Analysis & Breeding and Transfer Plan—Pallas’ Cat (Otocolobus manul). In AZA Species Survival Plan Red Program; AZA’s Population Management Center, Lincoln Park Zoo: Chicago, IL, USA, 2021; 44p, Available online: https://www.aza.org/population-management-center (accessed on 16 February 2024).
- Leberg, P.L.; Firmin, B.D. Role of inbreeding depression and purging in captive breeding and restoration programmes. Mol. Ecol. 2008, 17, 334–343. [Google Scholar] [CrossRef]
- Kayser, S. AZA Regional Studbook—Pallas’ Cat (Otocolobus manul). 2021; 73p. Available online: http://www.aza.org (accessed on 16 February 2024).
- Hutchins, M.; Wiese, R.J. Beyond genetic and demographic management: The future of the species survival plan and related AAZPA conservation efforts. Zoo Biol. 1991, 10, 285–292. [Google Scholar] [CrossRef]
- Fienieg, E.S.; Galbusera, P. The use and integration of molecular DNA information in conservation breeding programmes: A Review. J. Zoo Aquar. Res. 2013, 1, 44–51. [Google Scholar]
- Tian, H.; Mcknight, S.L.; Russell, D.W. Endothelial PAS domain protein 1 (EPAS1), a transcription factor selectively expressed in endothelial cells. Genes Dev. 1997, 11, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Janecka, J.E.; Hacker, C.; Broderick, J.; Pulugulla, S.; Auron, P.; Ringling, M.; Nelson, B.; Munkhtsog, B.; Hussain, S.; Davis, B.; et al. Noninvasive Genetics and Genomics Shed Light on the Status, Phylogeography, and Evolution of the Elusive Snow Leopard. In Conservation Genetics in Mammals: Integrative Research Using Novel Approaches; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 83–120. ISBN 9783030333348. [Google Scholar]
- Cho, Y.S.; Hu, L.; Hou, H.; Lee, H.; Xu, J.; Kwon, S.; Oh, S.; Kim, H.M.; Jho, S.; Kim, S.; et al. The tiger genome and comparative analysis with lion and snow leopard genomes. Nat. Commun. 2013, 4, 2433. [Google Scholar] [CrossRef] [PubMed]
- Werhahn, G.; Sherchan, A.M.; Manandhar, P. Eurasian lynx and Pallas’s cat in Dolpa District of Nepal: Genetics, distribution and diet. Cat News 2018, 67, 34–36. [Google Scholar]
- Zhao, D.; Yang, C.; Ma, J.; Zhang, X.; Ran, J. Vertebrate prey composition analysis of the Pallas’s cat (Otocolobus manul) in the Gongga Mountain Nature Reserve, based on fecal DNA. Mammalia 2020, 84, 449–457. [Google Scholar] [CrossRef]
- Hacker, C.E.; Cong, W.; Xue, Y.; Li, J.; Zhang, Y.; Wu, L.; Ji, Y.; Dai, Y.; Li, Y.; Jin, L.; et al. Dietary diversity and niche partitioning of carnivores across the Qinghai–Tibetan Plateau of China using DNA metabarcoding. J. Mammal. 2022, 103, 1005–1018. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, J.; Jiang, E.; Xu, Y.; Ning, F.; Du, Z.; Bai, X. The complete mitochondrial genome of Pallas’s cat (Otocolobus manul). Mitochondrial. DNA B Resour. 2019, 4, 658–659. [Google Scholar] [CrossRef]
- Flack, N.; Drown, M.; Walls, C.; Pratte, J.; Mclain, A.; Faulk, C. Chromosome-level, nanopore-only genome and allele-specific DNA methylation of Pallas’s cat, Otocolobus manul. bioRxiv 2022. [Google Scholar] [CrossRef]
- Kitchener, A.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the felidae. The final report of the cat classification task force of the IUCN/SSC Cat Specialist Group. Cat News 2017, 11, 1–80. [Google Scholar]
- Arif, I.A.; Khan, H.A.; Khan, H.A. Molecular markers for biodiversity analysis of wildlife animals: A brief review. Anim. Biodivers. Conserv. 2009, 32, 9–17. [Google Scholar] [CrossRef]
- Shrestha, B.; Kindlmann, P. Implications of landscape genetics and connectivity of snow leopard in the Nepalese Himalayas for its conservation. Sci. Rep. 2020, 10, 19853. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.G.; Hazzouri, K.M.; Choi, J.Y.; Delaney, P.; Al-Kharafi, M.; Howells, E.J.; Aranda, M.; Burt, J.A. Signatures of selection underpinning rapid coral adaptation to the world’s warmest reefs. Sci. Adv. 2022, 8, 7287. [Google Scholar] [CrossRef] [PubMed]
- DeYoung, R.W.; Honeycutt, R.L. The molecular toolbox: Genetic techniques in wildlife ecology and management. J. Wildl. Manag. 2005, 69, 1362–1384. [Google Scholar] [CrossRef]
- Janecka, J.E.; Zhang, Y.; Li, D.; Munkhtsog, B.; Bayaraa, M.; Galsandorj, N.; Wangchuk, T.R.; Karmacharya, D.; Li, J.; Lu, Z.; et al. Range-wide snow leopard phylogeography supports three subspecies. J. Hered. 2017, 108, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Hacker, C.E.; Jevit, M.; Hussain, S.; Muhammad, G.; Munkhtsog, B.; Munkhtsog, B.; Zhang, Y.; Li, D.; Liu, Y.; Farrington, J.D.; et al. Regional comparison of snow leopard (Panthera uncia) diet using DNA metabarcoding. Biodivers. Conserv. 2021, 30, 797–817. [Google Scholar] [CrossRef]
- Menotti-Raymond, M.; David, V.A.; Lyons, L.A.; Schä, A.A.; Tomlin, J.F.; Hutton, M.K.; O’brien, S.J. A genetic linkage map of microsatellites in the domestic cat (Felis catus). Genomics 1999, 57, 9–23. [Google Scholar] [CrossRef]
- Menotti-Raymond, M.; David, V.A.; Chen, Z.Q.; Menotti, K.A.; Sun, S.; Schaffer, A.A.; Agarwala, R.; Tomlin, J.F.; O’Brien, S.J.; Murphy, W.J. Second-generation integrated genetic linkage/radiation hybrid maps of the domestic cat (Felis catus). J. Hered. 2003, 94, 95–106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic Analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Waits, L.P.; Luikart, G.; Taberlet, P. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol. 2001, 10, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. An estimator for pairwise relatedness using molecular markers. Genetics 2002, 160, 1203–1215. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Coancestry: A program for simulating, estimating and analysing relatedness and inbreeding coefficients. Mol. Ecol. Resour. 2011, 11, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Ritland, K. Estimation of pairwise relatedness with molecular markers. Genetics 1999, 152, 1753–1766. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Estimating pairwise relatedness in a small sample of individuals. Heredity 2017, 119, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Riaz, T.; Shehzad, W.; Viari, A.; Pompanon, F.; Taberlet, P.; Coissac, E. EcoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis. Nucleic Acids Res. 2011, 39, e145. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bischof, R.; Gregersen, E.R.; Brøseth, H.; Ellegren, H.; Flagstad, Ø. Noninvasive genetic sampling reveals intrasex Territoriality in wolverines. Ecol. Evol. 2016, 6, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Treloar, S.; Lohr, C.; Hopkins, A.J.M.; Ottewell, K.; McArthur, S.; Davis, R.A. Scat DNA as a non-invasive method for estimating the abundance of the vulnerable mala (Lagorchestes hirsutus). Wildl. Res. 2023, 51, WR22122. [Google Scholar] [CrossRef]
- Taberlet, P.; Waits, L.P.; Luikart, G. Noninvasive genetic sampling: Look before you leap. Trends Ecol. Evol. 1999, 14, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Huisman, J.; Kruuk, L.E.B.; Ellis, P.A.; Clutton-Brock, T.; Pemberton, J.M. Inbreeding depression across the lifespan in a wild mammal population. Proc. Natl. Acad. Sci. USA 2016, 113, 3585–3590. [Google Scholar] [CrossRef] [PubMed]
- Prochowska, S.; Niżański, W.; Snoeck, F.; Wydooghe, E.; Van Soom, A.; Kochan, J.; Stefanyk, V. How can we introduce ART into wild felid conservation in practice? Joint experience in semen collection from captive wild felids in Europe. Animals 2022, 12, 871. [Google Scholar] [CrossRef] [PubMed]
- Swanson, W.F.; Stoops, M.A.; Magarey, G.M.; Herrick, J.R. Sperm Cryopreservation in Endangered Felids: Developing Linkage of in Situ-Ex Situ Populations. In Spermatology; Roldan, E., Gomenido, M., Eds.; Nottingham University Press: Nottingham, UK, 2007; pp. 417–432. [Google Scholar]
- Boakes, E.H.; Wang, J.; Amos, W. An investigation of inbreeding depression and purging in captive pedigreed populations. Heredity 2007, 98, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.G.R.; Woolhouse, M.E.J.; Tapio, M.; Mbole-Kariuki, M.N.; Sonstegard, T.S.; Thumbi, S.M.; Jennings, A.E.; Van Wyk, I.C.; Chase-Topping, M.; Kiara, H.; et al. Genetic susceptibility to infectious disease in East African shorthorn zebu: A genome-wide analysis of the effect of heterozygosity and exotic introgression. BMC Evol. Biol. 2013, 13, 246. [Google Scholar] [CrossRef]
- Pavlova, E.V.; Kirilyuk, V.E.; Naidenko, S.V. Patterns of seroprevalence of feline viruses among domestic cats (Felis catus) and Pallas’ cats (Otocolobus manul) in Daursky Reserve, Russia. Can. J. Zool. 2015, 93, 849–855. [Google Scholar] [CrossRef]
- Lücht, M.; Stagegaard, J.; Conraths, F.J.; Schares, G. Toxoplasma gondii in small exotic felids from zoos in Europe and the Middle East: Serological prevalence and risk factors. Parasit Vectors 2019, 12, 449. [Google Scholar] [CrossRef]
- Naidenko, S.; Demina, T. Pathogens and parasites as potential threats for Pallas’s cats. Cat News 2019, 13, 52–54. [Google Scholar]
- Voet, D.; Voet, J.G.; Pratt, C.W. Fundamentals of Biochemistry; John Wiley & Sons, Inc.: New York, NY, USA, 1999. [Google Scholar]
- Morris, J.; Hartl, D.; Knoll, A.; Lue, R.; Michael, M.; Berry, A.; Biewener, A.; Farrell, R.; Holbrook, N.M.; Heitz, J.; et al. Biology How Life Works, 3rd ed.; W. H. Freeman and Company: New York, NY, USA, 2013. [Google Scholar]
- Deagle, B.E.; Eveson, J.P.; Jarman, S.N. Quantification of damage in DNA recovered from highly degraded samples—A case study on DNA in faeces. Front. Zool. 2006, 3, 11. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Origin of Samples | n | AN | AE | HO | HE | FI | AT | AP |
|---|---|---|---|---|---|---|---|---|---|
| Otocolobus manul | Wild and zoo-managed populations | 16 | 5.103 ± 0.356 | 3.235 ± 0.239 | 0.571 ± 0.045 | 0.623 ± 0.038 | 0.083 ± 0.039 | 148 | N/A |
| Otocolobus manul | Zoo-managed population only | 10 | 3.552 ± 0.241 | 2.432 ± 0.167 | 0.590 ± 0.048 | 0.535 ± 0.033 | −0.093 ± 0.049 | 103 | 34 |
| Otocolobus manul | Wild population only | 6 | 3.931 ± 0.276 | 3.057 ± 0.241 | 0.540 ± 0.054 | 0.595 ± 0.041 | 0.105 ± 0.056 | 114 | 45 |
| Panthera uncia | Range-wide | 70 | 5.724 ± 0.412 | 2.753 ± 0.224 | 0.425 ± 0.034 | 0.572 ± 0.034 | 0.264 ± 0.029 | 166 | N/A |
| Oman11 | Oman12 | Oman13 | Oman14 | Oman15 | Oman16 | Oman17 | Oman18 | Oman19 | Oman20 | |
|---|---|---|---|---|---|---|---|---|---|---|
| N/A | Oman11 | |||||||||
| 0.091 | N/A | Oman12 | ||||||||
| 0.061 | −0.251 | N/A | Oman13 | |||||||
| 0.143 | −0.112 | −0.268 | N/A | Oman14 | ||||||
| 0.341 | −0.071 | −0.225 | 0.428 | N/A | Oman15 | |||||
| 0.069 | 0.077 | 0.339 | 0.147 | 0.061 | N/A | Oman16 | ||||
| 0.055 | −0.186 | 0.342 | −0.407 | −0.171 | −0.105 | N/A | Oman17 | |||
| 0.094 | 0.159 | −0.126 | −0.046 | 0.094 | 0.141 | −0.118 | N/A | Oman18 | ||
| 0.044 | 0.553 | −0.165 | −0.015 | −0.069 | 0.269 | −0.017 | 0.255 | N/A | Oman19 | |
| 0.128 | 0.209 | −0.136 | 0.182 | 0.168 | 0.309 | −0.429 | 0.406 | 0.167 | N/A | Oman20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, J.J.; Crichlow, A.D.; Hacker, C.E.; Munkhtsog, B.; Munkhtsog, B.; Zhang, Y.; Swanson, W.F.; Lyons, L.A.; Janecka, J.E. Genetic Variation in the Pallas’s Cat (Otocolobus manul) in Zoo-Managed and Wild Populations. Diversity 2024, 16, 228. https://doi.org/10.3390/d16040228
Robinson JJ, Crichlow AD, Hacker CE, Munkhtsog B, Munkhtsog B, Zhang Y, Swanson WF, Lyons LA, Janecka JE. Genetic Variation in the Pallas’s Cat (Otocolobus manul) in Zoo-Managed and Wild Populations. Diversity. 2024; 16(4):228. https://doi.org/10.3390/d16040228
Chicago/Turabian StyleRobinson, Joshua J., Alexis D. Crichlow, Charlotte E. Hacker, Bariushaa Munkhtsog, Bayaraa Munkhtsog, Yuguang Zhang, William F. Swanson, Leslie A. Lyons, and Jan E. Janecka. 2024. "Genetic Variation in the Pallas’s Cat (Otocolobus manul) in Zoo-Managed and Wild Populations" Diversity 16, no. 4: 228. https://doi.org/10.3390/d16040228
APA StyleRobinson, J. J., Crichlow, A. D., Hacker, C. E., Munkhtsog, B., Munkhtsog, B., Zhang, Y., Swanson, W. F., Lyons, L. A., & Janecka, J. E. (2024). Genetic Variation in the Pallas’s Cat (Otocolobus manul) in Zoo-Managed and Wild Populations. Diversity, 16(4), 228. https://doi.org/10.3390/d16040228