
Phylogeographic Relationships Reveal the Origin of an Introduced Population of the Dalmatian Algyroides (Reptilia: Lacertidae) into Southern Italy

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Laboratory Procedures
2.2. Phylogenetic and Demographic Analyses
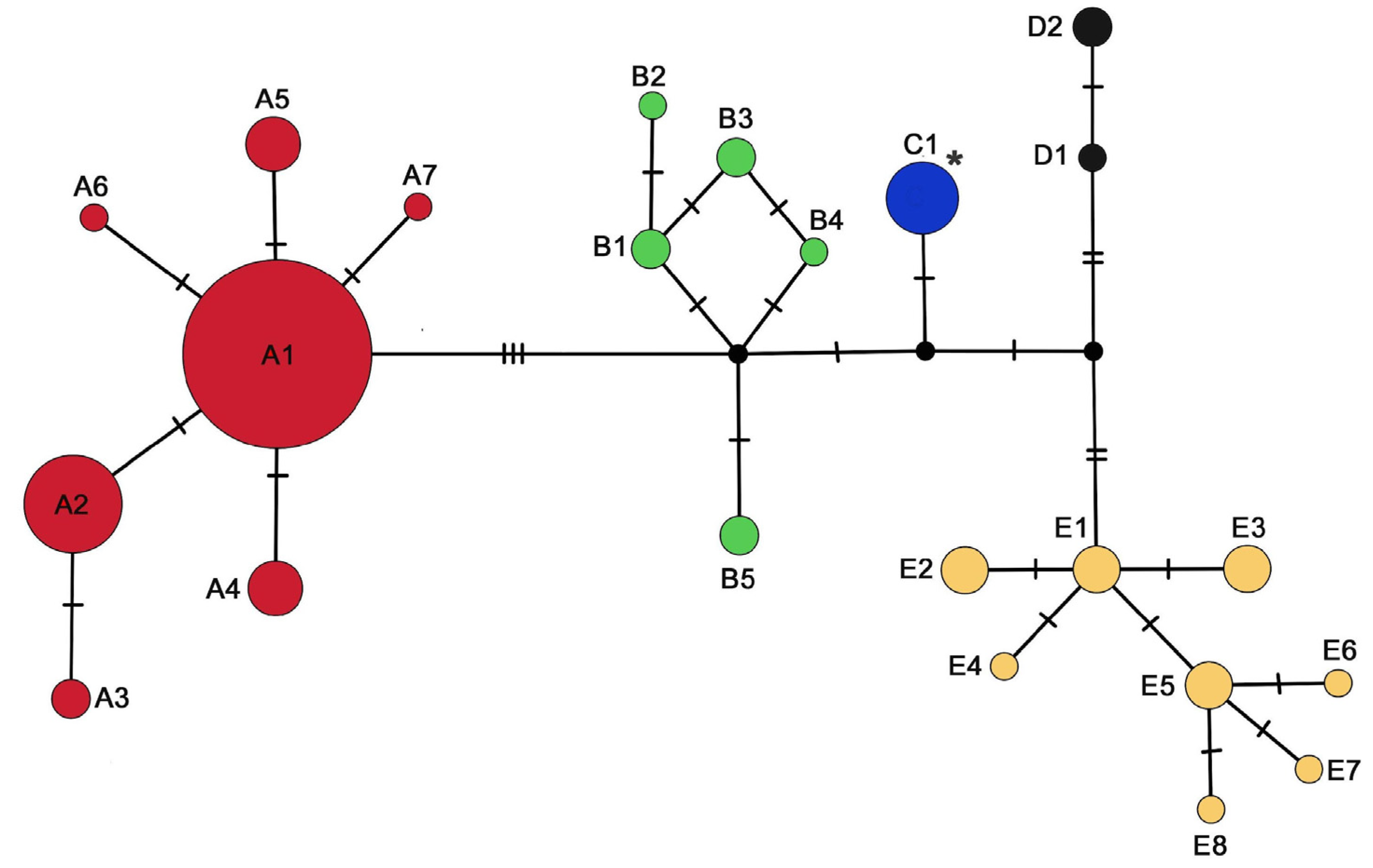
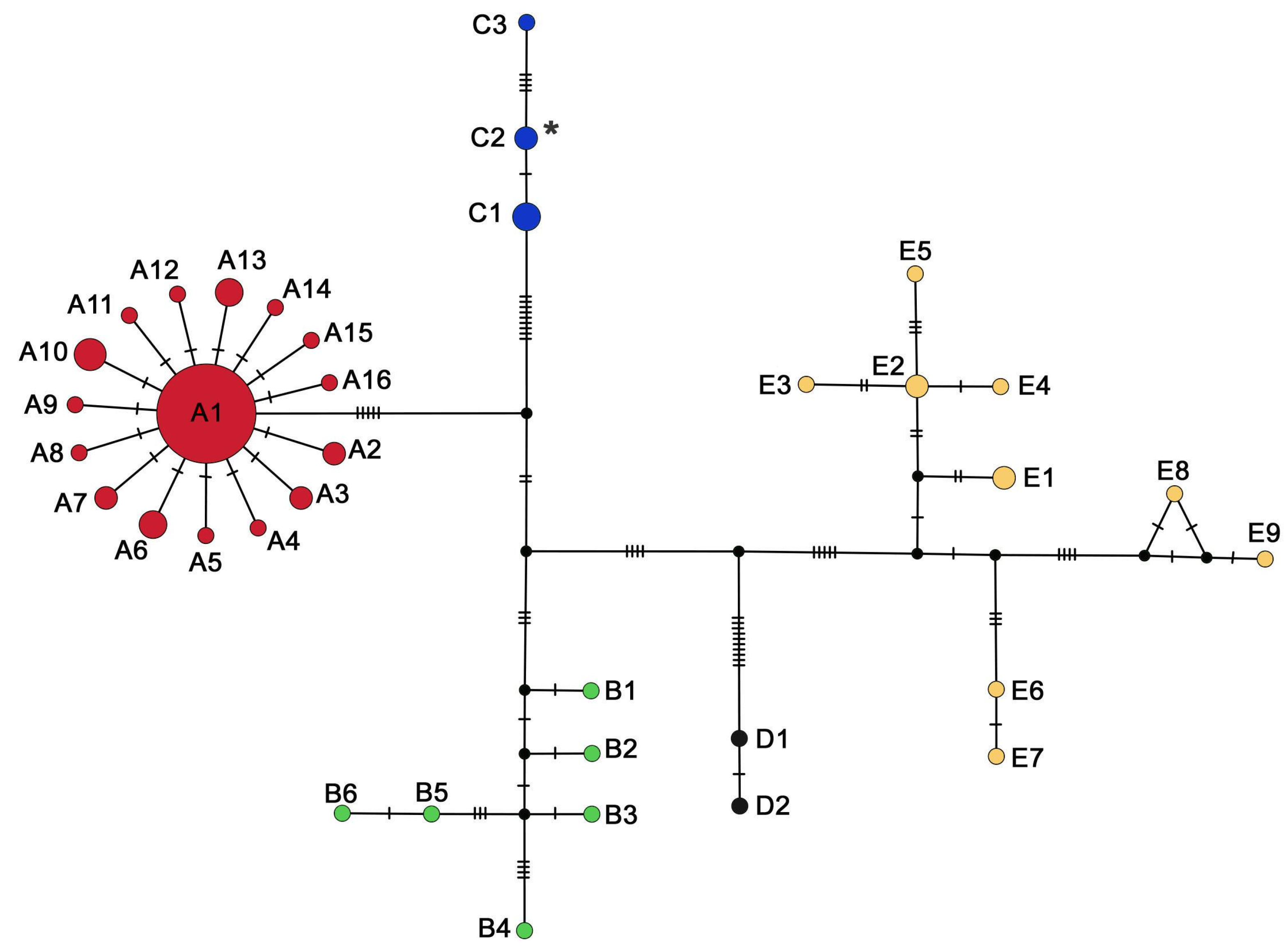
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hewitt, G.M. Some genetic consequences of ice ages, and their role in divergence and speciation. Biol. J. Linn. Soc. 1996, 58, 247–276. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 2, 305–332. [Google Scholar] [CrossRef]
- Allendorf, F.W.; Lundquist, L.L. Introduction: Population biology, evolution, and control of invasive species. Conserv. Biol. 2003, 17, 24–30. [Google Scholar] [CrossRef]
- Storfer, A.; Murphy, M.A.; Spear, S.F.; Holderegger, R.; Waits, L.P. Landscape genetics: Where are we now? Mol. Ecol. 2010, 19, 3496–3514. [Google Scholar] [CrossRef]
- Vila, M.; Basnou, C.; Pysek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef]
- Crispo, E.; Moore, J.S.; Lee-Yaw, J.A.; Gray, S.M.; Haller, B.C. Broken barriers: Human-induced changes to gene flow and introgression in animals: An examination of the ways in which humans increase genetic exchange among populations and species and the consequences for biodiversity. BioEssays 2011, 33, 508–518. [Google Scholar] [CrossRef]
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef]
- Kraus, F. Alien Reptiles and Amphibians: A Scientific Compendium and Analysis; Invading Nature—Springer Series in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2007; Volume 4, pp. 133–369. [Google Scholar] [CrossRef]
- Silva-Rocha, I.R.; Salvi, D.; Carretero, M.A.; Ficetola, G.F. Alien reptiles on Mediterranean Islands: A model for invasion biogeography. Divers. Distrib. 2019, 25, 995–1005. [Google Scholar] [CrossRef]
- Fitzpatrick, B.M.; Fordyce, J.A.; Niemiller, M.L.; Reynolds, R.G. What can DNA tell us about biological invasions? Biol. Invasions 2012, 14, 245–253. [Google Scholar] [CrossRef]
- Cristescu, M.E. Genetic reconstructions of invasion history. In: Invasion Genetics: The Baker and Stebbins Legacy. Mol. Ecol. 2016, 24, 267–282. [Google Scholar] [CrossRef]
- Oficialdegui, F.J.; Clavero, M.; Sánchez, M.I.; Green, A.J.; Boyero, L.; Michot, T.C.; Klose, K.; Kawai, T.; Lejeusne, C. Unravelling the global invasion routes of a worldwide invader, the red swamp crayfish (Procambarus clarkii). Freshw. Biol. 2019, 64, 1382–1400. [Google Scholar] [CrossRef]
- Eales, J.; Thorpe, R.S. Revealing the geographic origin of an invasive lizard: The problem of native population genetic diversity. Biol. Invasions 2010, 12, 77–86. [Google Scholar] [CrossRef]
- Michaelides, S.; While, G.M.; Bell, C.; Uller, T. Human introductions create opportunities for intra-specific hybridization in an alien lizard. Biol. Invasions 2013, 15, 1101–1112. [Google Scholar] [CrossRef]
- Santos, J.L.; Žagar, A.; Drašler, K.; Rato, C.; Ayres, C.; Harris, D.J.; Carretero, M.A.; Salvi, D. Phylogeographic evidence for multiple long-distance introductions of the common wall lizard associated with human trade and transport. Amphib.-Reptil. 2019, 40, 121–127. [Google Scholar] [CrossRef]
- Silva-Rocha, I.R.; Salvi, D.; Carretero, M.A. Genetic data reveal a multiple origin for the populations of the Italian wall lizard Podarcis sicula (Squamata: Lacertidae) introduced in the Iberian Peninsula and Balearic islands. Ital. J. Zool. 2012, 79, 502–510. [Google Scholar] [CrossRef]
- Silva-Rocha, I.R.; Salvi, D.; Harris, D.J.; Freits, S.; Davis, C.; Foster, J.; Deichsel, G.; Adamopoulou, C.; Carretero, M.A. Molecular assessment of Podarcis sicula populations in Britain, Greece and Turkey reinforces a multiple-origin invasion pattern in this species. Acta Herpetol. 2014, 9, 253–258. [Google Scholar] [CrossRef]
- Oskyrko, O.; Sreelatha, L.B.; Hanke, G.F.; Deichsel, G.; Carretero, M.A. Origin of introduced Italian wall lizards, Podarcis siculus (rafinesque-schmaltz, 1810) (Squamata: Lacertidae), in North America. BioInvasions Rec. 2022, 11, 1095–1106. [Google Scholar] [CrossRef]
- Senczuk, G.; Colangelo, P.; De Simone, E.; Aloise, G.; Castiglia, R. A combination of long term fragmentation and glacial persistence drove the evolutionary history of the Italian wall lizard Podarcis siculus. BMC Evol. Biol. 2017, 17, 6. [Google Scholar] [CrossRef] [PubMed]
- Carlino, P.; Pauwels, O.S.G. Taxonomic identity of an extralimital population of Algyroides Lizards (Squamata: Lacertidae) from Apulia Region in Southern Italy. Bull. Chic. Herpetol. Soc. 2016, 51, 149–151. [Google Scholar]
- Deimezis-Tsikoutas, A.; Capsalas, G.; Antonopoulos, A.; Strachinis, I.; Pafilis, P. Algyroides nigropunctatus (Squamata: Lacertidae) in the city of Athens: An unexpected finding. Russ. J. Herpetol. 2020, 27, 172–174. [Google Scholar] [CrossRef]
- Arnold, E.N.; Ovenden, D. A Field Guide to the Reptiles and Amphibians of Britain and Europe, 2nd ed.; Collins: London, UK, 1978. [Google Scholar]
- Chondropoulos, B.P. A checklist of the Greek reptiles. I. The lizards. Amphib.-Reptil. 1986, 7, 217–235. [Google Scholar] [CrossRef]
- Valakos, E.; Pafilis, P.; Sotiropoulos, K.; Lymberakis, P.; Maragou, P.; Foufopoulos, J. The Amphibians and Reptiles of Greece; Chimaira: Frankfurt, Germany, 2008. [Google Scholar]
- Wilson, M.; Stille, B.; Stille, M. Herpetofauna of Paxos, Ionian Islands, Greece, including two species new to the island. Herpetozoa 2014, 27, 108–112. [Google Scholar]
- Stille, B.; Stille, M. The Herpetofauna of Corfu and Adjacent Islands; Chimaira: Frankfurt, Germany, 2017. [Google Scholar]
- Griffiths, H.I.; Krystufek, B.; Reed, J.M. Balkan Biodiversity: Pattern and Process in the European Hotspot; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Lymberakis, P.; Poulakakis, N. Three continents claiming an archipelago: The evolution of Aegean’s herpetofaunal diversity. Diversity 2010, 2, 233–255. [Google Scholar] [CrossRef]
- Hewitt, G.M. Mediterranean peninsulas: The evolution of hotspots. In Biodiversity Hotspots: Distribution and Protection of Conservation Priority Areas; Zachos, F., Habel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 123–147. [Google Scholar]
- Psonis, N.; Antoniou, A.; Karameta, E.; Leaché, A.D.; Kotsakiozi, P.; Darriba, D.; Kozlov, A.; Stamatakis, A.; Poursanidis, D.; Kukushkin, O.; et al. Resolving complex phylogeographic patterns in the Balkan Peninsula using closely related wall-lizard species as a model system. Mol. Phylogenet. Evol. 2018, 125, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Podnar, M.; Mayer, W. First insights into the mitochondrial DNA diversity of Dalmatian Algyroides, Algyroides nigropunctatus (Lacertidae). Period. Biol. 2006, 108, 85–87. [Google Scholar]
- Strachinis, I.; Poulakakis, N.; Karaiskou, N.; Patronidis, P.; Patramanis, I.; Poursanidis, D.; Jablonski, D.; Triantafyllidis, A. Phylogeography and systematics of Algyroides (Sauria: Lacertidae) of the Balkan Peninsula. Zool. Scr. 2021, 50, 282–299. [Google Scholar] [CrossRef]
- QGIS.org. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2023. Available online: http://qgis.org (accessed on 12 April 2024).
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [PubMed]
- Arèvalo, E.; Davis, S.K.; Sites, J.W. Mitochondrial DNA sequence divergence and phylogenetic relationships among eight chromosome races of the Sceloporus grammicus complex (Phrynosomatidae) in central Mexico. Syst. Biol. 1994, 43, 387–418. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR, version 2.0; University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Pavlicev, M.; Mayer, W. Fast radiation of the subfamily Lacertinae (Reptilia: Lacertidae): History or methodical artefact? Mol. Phylogenet. Evol. 2009, 52, 727–734. [Google Scholar] [CrossRef]
- Salvi, D.; Harris, D.J.; Perera, A.; Bologna, M.A.; Carretero, M.A. Preliminary survey on genetic variation within the Pygmy Algyroides, Algyroides fitzingeri, across Corsica and Sardinia. Amphib.-Reptil. 2011, 32, 281–286. [Google Scholar]
- Greenbaum, E.; Villanueva, C.O.; Kusamba, C.; Aristote, M.M.; Branch, W.R. A molecular phylogeny of Equatorial African Lacertidae, with the description of a new genus and species from eastern Democratic Republic of the Congo. Zool. J. Linn. Soc. 2011, 163, 913–942. [Google Scholar] [CrossRef] [PubMed]
- Kirchhof, S.; Lyra, M.L.; Rodríguez, A.; Ineich, I.; Müller, J.; Rödel, M.O.; Trape, J.F.; Vences, M.; Boissinot, S. Mitogenome analyses elucidate the evolutionary relationships of a probable Eocene wet tropics relic in the xerophilic lizard genus Acanthodactylus. Sci. Rep. 2021, 11, 4858. [Google Scholar] [CrossRef] [PubMed]
- Rato, C.; Sillero, N.; Ceacero, F.; Muñoz, E.G.; Carretero, M.A. A survival story: Evolutionary history of the Iberian Algyroides (Squamata: Lacertidae), an endemic lizard relict. Biodivers. Conserv. 2021, 30, 2707–2729. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post- analysis of large phylogenies. Bioinformatics 2014, 22, 1312–1313. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Bressi, N. Algyroides nigropunctatus nigropunctatus in Italy: Notes on ecology, habitat selection and conservation (Reptilia, Lacertidae). Ital. J. Zool. J. 2004, 71, 113–116. [Google Scholar] [CrossRef]
- Zakkak, S.; Halley, J.M.; Akriotis, T.; Kati, V. Lizards along an agricultural land abandonment gradient in Pindos Mountains, Greece. Amphib.-Reptil. 2015, 36, 253–264. [Google Scholar] [CrossRef]
- Carneiro, D.; García-Muñoz, E.; Žagar, A.; Pafilis, P.; Carretero, M.A. Is ecophysiology congruent with the present-day relictual distribution of a lizard group? Evidence from preferred temperatures and water loss rates. Herpetol. J. 2017, 27, 47–56. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.U.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or Bootstrap? A Simulation Study Comparing the Performance of Bayesian Markov Chain Monte Carlo Sampling and Bootstrapping in Assessing Phylogenetic Confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef]
- Maddison, W.P. Gene trees in species trees. Syst. Biol. 1997, 46, 523–536. [Google Scholar] [CrossRef]
- Maddison, W.P.; Knowles, L.L. Inferring phylogeny despite incomplete lineage sorting. Syst. Biol. 2006, 55, 21–30. [Google Scholar] [CrossRef]
- Pinho, C.; Harris, D.J.; Ferrand, N. Contrasting patterns of population subdivision and historical demography in three western Mediterranean lizard species inferred from mitochondrial DNA variation. Mol. Ecol. 2007, 16, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Savić, I.R. Diversification of the Balkan fauna: Its origin, historical development and present status. Adv. Arachnol. Dev. Biol. 2008, 12, 57–78. [Google Scholar]
- Canestrelli, D.; Aloise, G.; Cecchetti, S.; Nascetti, G. Birth of a hotspot of intraspecific genetic diversity: Notes from the underground. Mol. Ecol. 2010, 19, 5432–5451. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: Retrospect and prospect. J. Biogeogr. 2009, 36, 3–15. [Google Scholar] [CrossRef]
- Ursenbacher, S.; Schweiger, S.; Tomović, L.; Crnobrnja-Isailović, J.; Fumagalli, L.; Mayer, W. Molecular phylogeography of the nose-horned viper (Vipera ammodytes, Linnaeus (1758)): Evidence for high genetic diversity and multiple refugia in the Balkan peninsula. Mol. Phylogenet. Evol. 2008, 46, 1116–1128. [Google Scholar] [CrossRef] [PubMed]
- Kotsakiozi, P.; Parmakelis, A.; Giokas, S.; Papanikolaou, I.; Valakos, E.D. Mitochondrial phylogeny and biogeographic history of the Greek endemic land-snail genus Codringtonia Kobelt 1898 (Gastropoda, Pulmonata, Helicidae). Mol. Phylogenet. Evol. 2012, 62, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Sagonas, K.; Poulakakis, N.; Lymberakis, P.; Parmakelis, A.; Pafilis, P.; Valakos, E.D. Molecular systematics and historical biogeography of the green lizards (Lacerta) in Greece: Insights from mitochondrial and nuclear DNA. Mol. Phylogenet. Evol. 2014, 76, 144–154. [Google Scholar] [CrossRef]
- Kornilios, P.; Thanou, E.; Kapli, P.; Parmakelis, D.; Chatzaki, M. Peeking through the trapdoor: Historical biogeography of the Aegean endemic spider Cyrtocarenum Ausserer, 1871 with an estimation of mtDNA substitution rates for Mygalomorphae. Mol. Phylogenet. Evol. 2016, 98, 300–313. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Lineage A | Lineage B | Lineage C | Lineage D | Linage E | |
| Lineage A | 0.1/0.1 | 2.0 | 2.2 | 3.3 | 2.9 |
| Lineage B | 1.0 | 0.4/0.7 | 2.7 | 3.1 | 2.9 |
| Lineage C | 1.0 | 0.7 | 0.0/0.3 | 2.6 | 2.9 |
| Lineage D | 1.3 | 1.3 | 1.0 | 0.1/0.1 | 3.0 |
| Lineage E | 1.8 | 1.3 | 1.0 | 1.2 | 0.3/1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toli, E.-A.; Sergiadou, D.; Carlino, P.; Bounas, A.; Carretero, M.A.; Castiglia, R.; Harris, D.J.; Papadaki, C.; Pauwels, O.S.G.; Leković, L.; et al. Phylogeographic Relationships Reveal the Origin of an Introduced Population of the Dalmatian Algyroides (Reptilia: Lacertidae) into Southern Italy. Diversity 2024, 16, 406. https://doi.org/10.3390/d16070406
Toli E-A, Sergiadou D, Carlino P, Bounas A, Carretero MA, Castiglia R, Harris DJ, Papadaki C, Pauwels OSG, Leković L, et al. Phylogeographic Relationships Reveal the Origin of an Introduced Population of the Dalmatian Algyroides (Reptilia: Lacertidae) into Southern Italy. Diversity. 2024; 16(7):406. https://doi.org/10.3390/d16070406
Chicago/Turabian StyleToli, Elisavet-Aspasia, Dimitra Sergiadou, Piero Carlino, Anastasios Bounas, Miguel A. Carretero, Riccardo Castiglia, D. James Harris, Chrysoula Papadaki, Olivier S. G. Pauwels, Lidija Leković, and et al. 2024. "Phylogeographic Relationships Reveal the Origin of an Introduced Population of the Dalmatian Algyroides (Reptilia: Lacertidae) into Southern Italy" Diversity 16, no. 7: 406. https://doi.org/10.3390/d16070406
APA StyleToli, E.-A., Sergiadou, D., Carlino, P., Bounas, A., Carretero, M. A., Castiglia, R., Harris, D. J., Papadaki, C., Pauwels, O. S. G., Leković, L., & Sotiropoulos, K. (2024). Phylogeographic Relationships Reveal the Origin of an Introduced Population of the Dalmatian Algyroides (Reptilia: Lacertidae) into Southern Italy. Diversity, 16(7), 406. https://doi.org/10.3390/d16070406